


Proteomic Analysis Reveals That Mitochondria Dominate the Hippocampal Hypoxic Response in Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
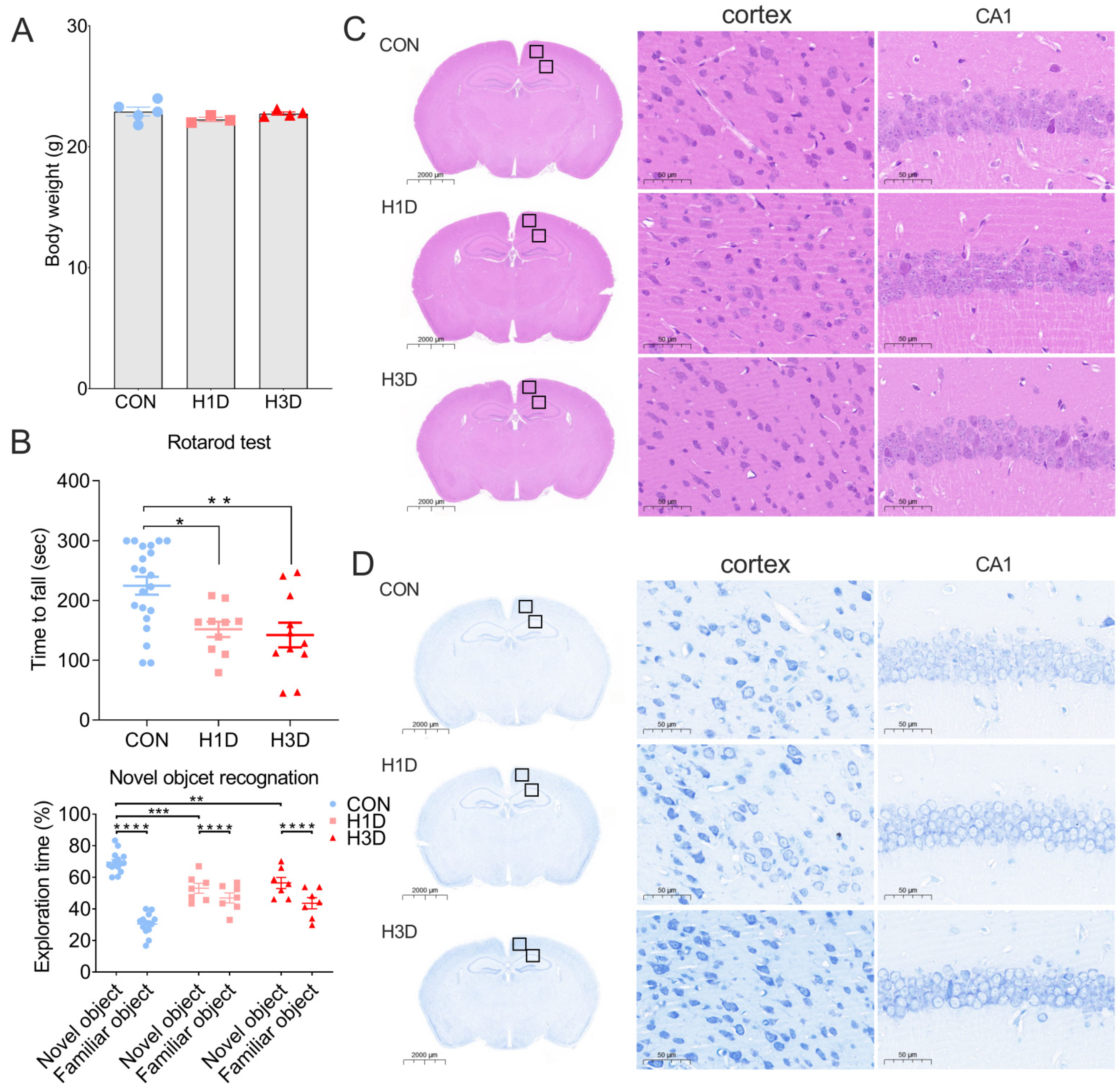
2.1. Hypoxic Stress Impaired Cognitive and Motor Function but Did Not Alter the Morphology or Structure of Hippocampal Neurons
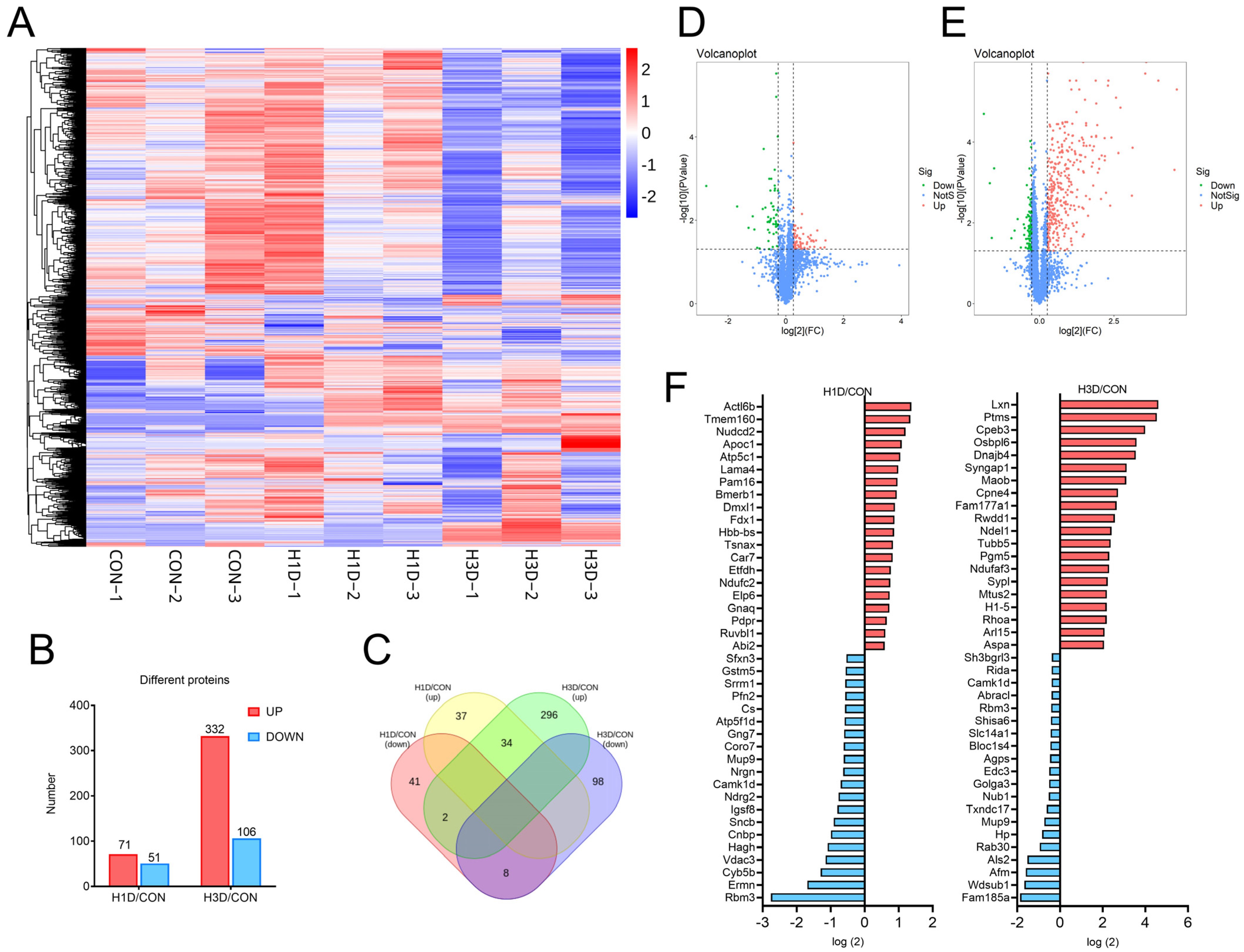
2.2. Global Proteomic Signatures of the Hippocampus under Hypoxic Stress
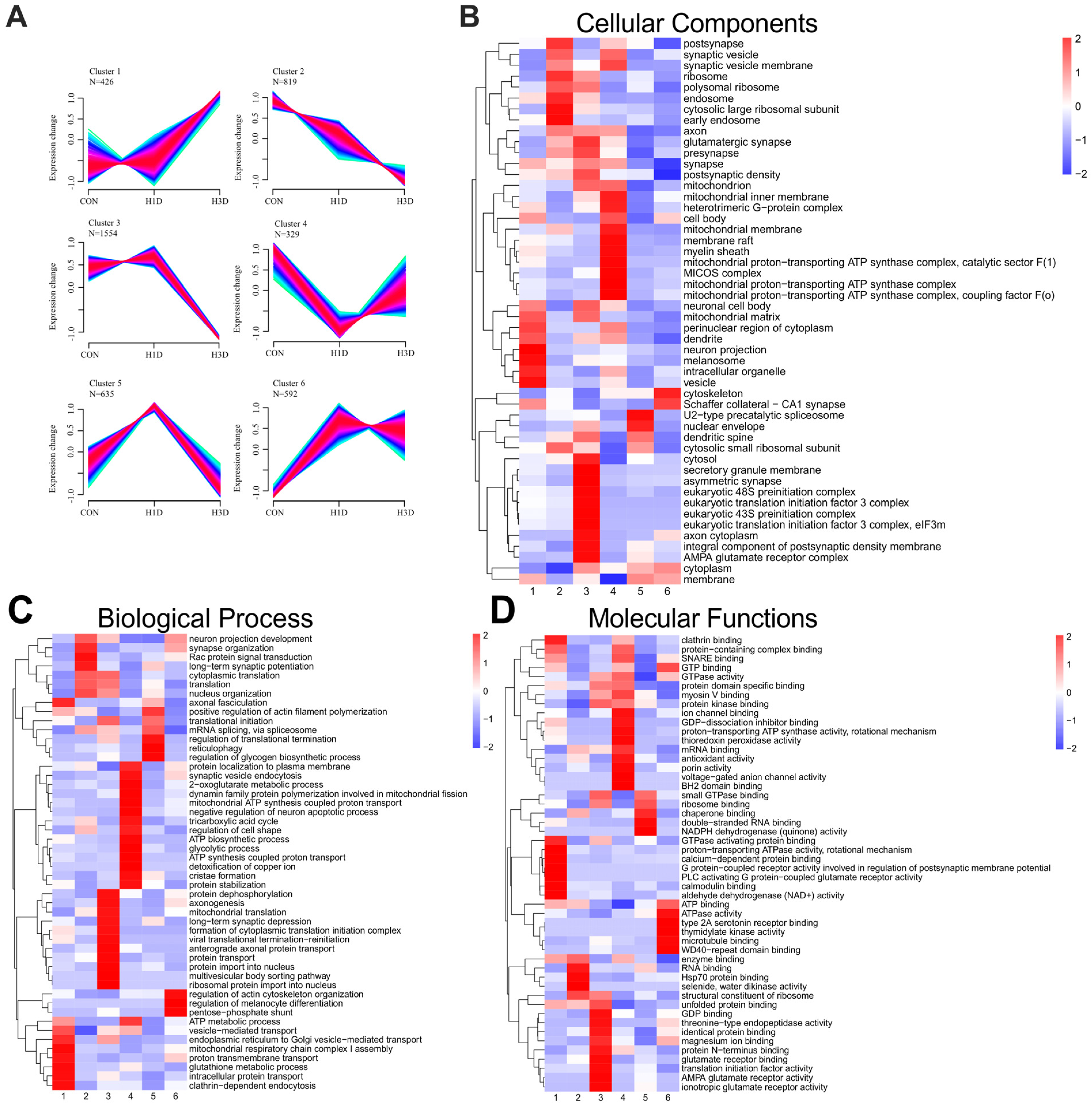
2.3. Hippocampal Proteome Dynamic Alterations Induced by Hypoxic Stress
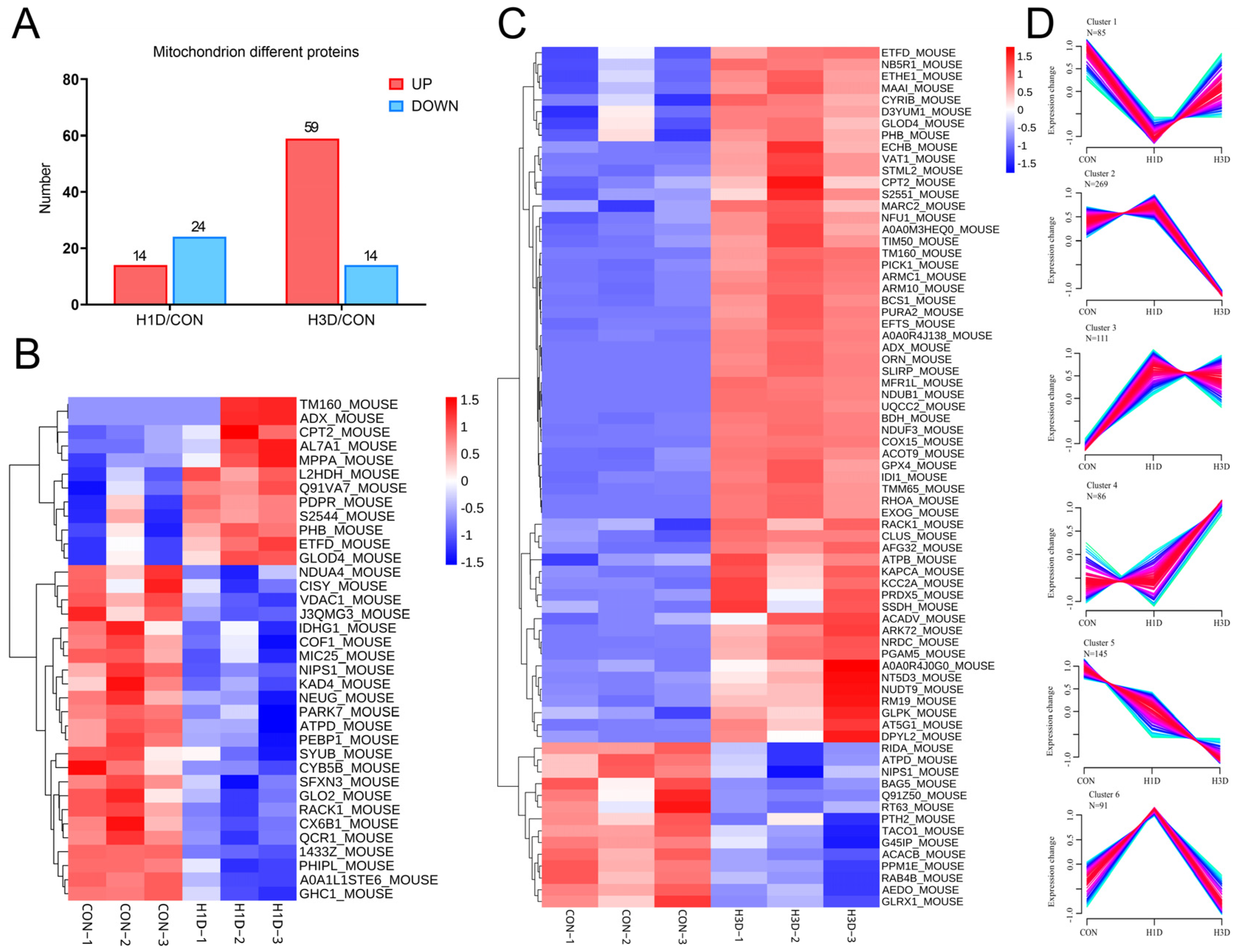
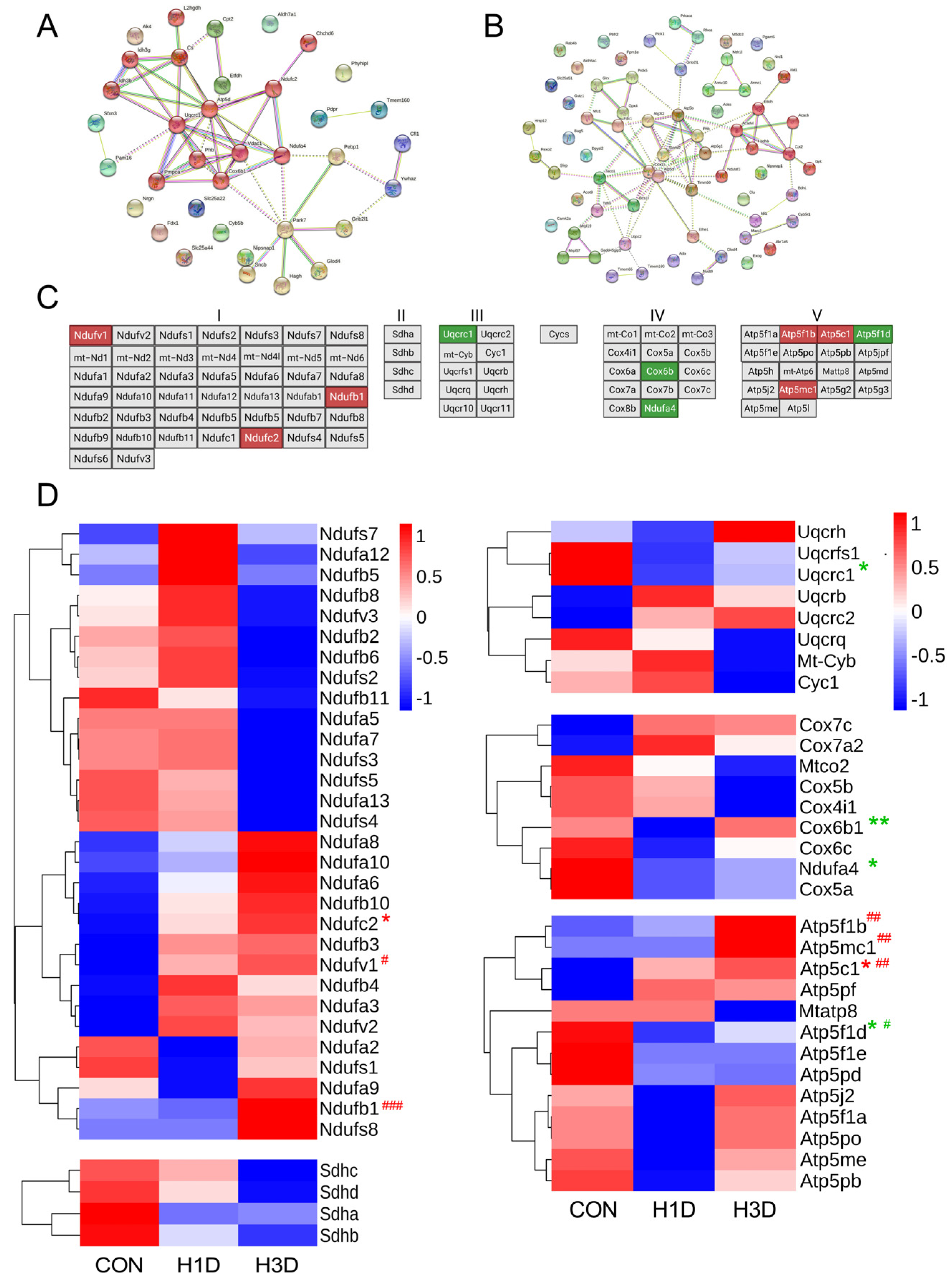
2.4. Hypoxic Stress Mainly Caused Mitochondrial Dysfunction
2.5. Hypoxic Stress Impaired Mitochondrial Oxidative Phosphorylation by Suppressing Mitochondrial Complex IV
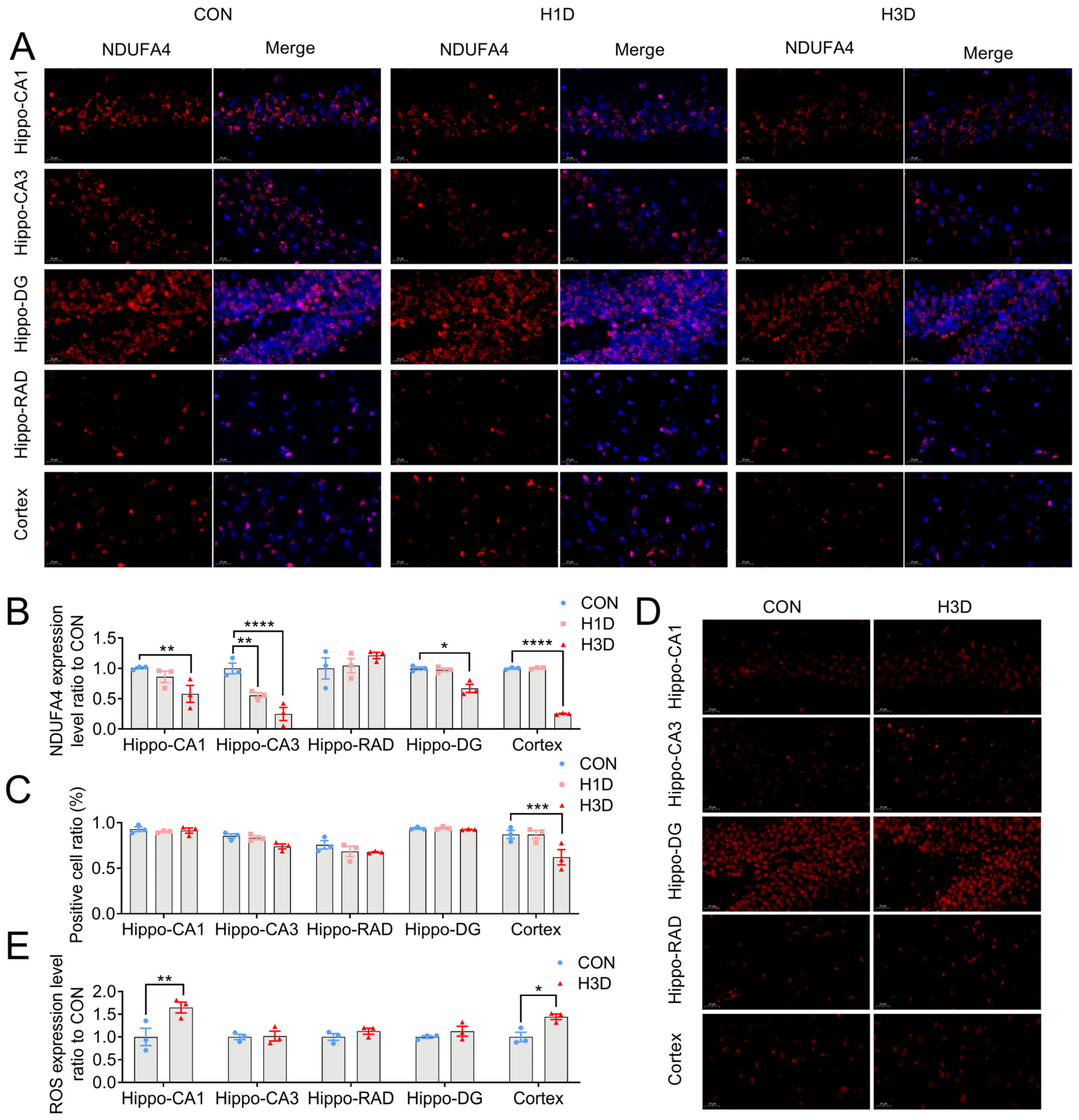
2.6. Hypoxic Stress Caused Mitochondrial Complex IV Dysfunction by Downregulating NDUFA4 Expression
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Hypoxia Treatment
4.3. Rotarod Test
4.4. Novel Object Recognition
4.5. HE and Nissl Staining
4.5.1. HE Staining
4.5.2. Nissl Staining
4.6. Immunofluorescence Staining for ROS Detection
4.7. Immunofluorescence Labeling
4.8. Proteomic Analysis
4.8.1. Sample Preparation
4.8.2. TMT Labeling
4.8.3. Peptide Identification by Nano UPLC-MS/MS
4.8.4. Protein Identification
4.8.5. Bioinformatic Analysis
4.9. Statistical Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CI | NADH dehydrogenase; |
| CII | Succinate dehydrogenase; |
| CIV | Mitochondrial complex IV; |
| CIII | Bc1-complex; |
| COX | Cytochrome c oxidase; |
| COX4I1 | COX4 subtype 1; |
| COX4I2 | COX4 subtype 2; |
| ETC | Electron transfer chain; |
| HIF | Hypoxia-inducible Factor; |
| GO | Gene Ontology; |
| KEGG | Kyoto Encyclopedia of Genes and Genomes; |
| LDHA | Lactate dehydrogenase A; |
| NDUFA4 | NADH dehydrogenase (ubiquinone) 1 alpha subcomplex 4; |
| OXPHOS | Oxidative phosphorylation; |
| PDH | Pyruvate dehydrogenase; |
| PDK1 | Pyruvate dehydrogenase kinase 1; |
| PHDS | Prolyl hydroxylases; |
| ROS | Reactive oxygen species; |
| TCA | Tricarboxylic acid cycle; |
References
- Trzepizur, W.; Blanchard, M.; Ganem, T.; Balusson, F.; Feuilloy, M.; Girault, J.M.; Meslier, N.; Oger, E.; Paris, A.; Pigeanne, T.; et al. Sleep Apnea-Specific Hypoxic Burden, Symptom Subtypes, and Risk of Cardiovascular Events and All-Cause Mortality. Am. J. Respir. Crit. Care Med. 2022, 205, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Kaczynska, K.; Orlowska, M.E.; Andrzejewski, K. Respiratory Abnormalities in Parkinson’s Disease: What Do We Know from Studies in Humans and Animal Models? Int. J. Mol. Sci. 2022, 23, 3499. [Google Scholar] [CrossRef] [PubMed]
- Kazim, S.F.; Sharma, A.; Saroja, S.R.; Seo, J.H.; Larson, C.S.; Ramakrishnan, A.; Wang, M.; Blitzer, R.D.; Shen, L.; Pena, C.J.; et al. Chronic Intermittent Hypoxia Enhances Pathological Tau Seeding, Propagation, and Accumulation and Exacerbates Alzheimer-like Memory and Synaptic Plasticity Deficits and Molecular Signatures. Biol. Psychiatry 2022, 91, 346–358. [Google Scholar] [CrossRef] [PubMed]
- Curila, K.; Jurak, P.; Halamek, J.; Prinzen, F.; Waldauf, P.; Karch, J.; Stros, P.; Plesinger, F.; Mizner, J.; Susankova, M.; et al. Ventricular activation pattern assessment during right ventricular pacing: Ultra-high-frequency ECG study. J. Cardiovasc. Electrophysiol. 2021, 32, 1385–1394. [Google Scholar] [CrossRef]
- Wilson, M.H.; Newman, S.; Imray, C.H. The cerebral effects of ascent to high altitudes. Lancet Neurol. 2009, 8, 175–191. [Google Scholar] [CrossRef]
- Tesler, N.; Latshang, T.D.; Lo Cascio, C.M.; Stadelmann, K.; Stoewhas, A.C.; Kohler, M.; Bloch, K.E.; Achermann, P.; Huber, R. Ascent to moderate altitude impairs overnight memory improvements. Physiol. Behav. 2015, 139, 121–126. [Google Scholar] [CrossRef]
- Hota, S.K.; Sharma, V.K.; Hota, K.; Das, S.; Dhar, P.; Mahapatra, B.B.; Srivastava, R.B.; Singh, S.B. Multi-domain cognitive screening test for neuropsychological assessment for cognitive decline in acclimatized lowlanders staying at high altitude. Indian J. Med. Res. 2012, 136, 411–420. [Google Scholar]
- Markolovic, S.; Wilkins, S.E.; Schofield, C.J. Protein Hydroxylation Catalyzed by 2-Oxoglutarate-dependent Oxygenases. J. Biol. Chem. 2015, 290, 20712–20722. [Google Scholar] [CrossRef] [Green Version]
- Lee, P.; Chandel, N.S.; Simon, M.C. Cellular adaptation to hypoxia through hypoxia inducible factors and beyond. Nat. Rev. Mol. Cell Biol. 2020, 21, 268–283. [Google Scholar] [CrossRef]
- Mookerjee, S.A.; Gerencser, A.A.; Nicholls, D.G.; Brand, M.D. Quantifying intracellular rates of glycolytic and oxidative ATP production and consumption using extracellular flux measurements. J. Biol. Chem. 2017, 292, 7189–7207. [Google Scholar] [CrossRef] [Green Version]
- Papandreou, I.; Cairns, R.A.; Fontana, L.; Lim, A.L.; Denko, N.C. HIF-1 mediates adaptation to hypoxia by actively downregulating mitochondrial oxygen consumption. Cell Metab. 2006, 3, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Dong, S.; Liu, W.; Wang, M.; Tian, S.; Ai, Y.; Wang, H. Accumulated ROS Activates HIF-1alpha-Induced Glycolysis and Exerts a Protective Effect on Sensory Hair Cells Against Noise-Induced Damage. Front. Mol. Biosci. 2021, 8, 806650. [Google Scholar] [CrossRef] [PubMed]
- Aragones, J.; Schneider, M.; Van Geyte, K.; Fraisl, P.; Dresselaers, T.; Mazzone, M.; Dirkx, R.; Zacchigna, S.; Lemieux, H.; Jeoung, N.H.; et al. Deficiency or inhibition of oxygen sensor Phd1 induces hypoxia tolerance by reprogramming basal metabolism. Nat. Genet. 2008, 40, 170–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scharping, N.E.; Rivadeneira, D.B.; Menk, A.V.; Vignali, P.D.A.; Ford, B.R.; Rittenhouse, N.L.; Peralta, R.; Wang, Y.; Wang, Y.; DePeaux, K.; et al. Mitochondrial stress induced by continuous stimulation under hypoxia rapidly drives T cell exhaustion. Nat. Immunol. 2021, 22, 205–215. [Google Scholar] [CrossRef]
- Vercellino, I.; Sazanov, L.A. The assembly, regulation and function of the mitochondrial respiratory chain. Nat. Rev. Mol. Cell Biol. 2022, 23, 141–161. [Google Scholar] [CrossRef] [PubMed]
- Kadenbach, B. Complex IV—The regulatory center of mitochondrial oxidative phosphorylation. Mitochondrion 2021, 58, 296–302. [Google Scholar] [CrossRef]
- Dutta, S.; Sengupta, P. Men and mice: Relating their ages. Life Sci. 2016, 152, 244–248. [Google Scholar] [CrossRef]
- Abrous, D.N.; Koehl, M.; Lemoine, M. A Baldwin interpretation of adult hippocampal neurogenesis: From functional relevance to physiopathology. Mol. Psychiatry 2022, 27, 383–402. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Zong, S.; Wu, M.; Gu, J.; Liu, T.; Guo, R.; Yang, M. Structure of the intact 14-subunit human cytochrome c oxidase. Cell Res. 2018, 28, 1026–1034. [Google Scholar] [CrossRef] [Green Version]
- Jain, I.H.; Zazzeron, L.; Goli, R.; Alexa, K.; Schatzman-Bone, S.; Dhillon, H.; Goldberger, O.; Peng, J.; Shalem, O.; Sanjana, N.E.; et al. Hypoxia as a therapy for mitochondrial disease. Science 2016, 352, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Tello, D.; Balsa, E.; Acosta-Iborra, B.; Fuertes-Yebra, E.; Elorza, A.; Ordonez, A.; Corral-Escariz, M.; Soro, I.; Lopez-Bernardo, E.; Perales-Clemente, E.; et al. Induction of the mitochondrial NDUFA4L2 protein by HIF-1alpha decreases oxygen consumption by inhibiting Complex I activity. Cell Metab. 2011, 14, 768–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaelin, W.G., Jr.; Ratcliffe, P.J. Oxygen sensing by metazoans: The central role of the HIF hydroxylase pathway. Mol. Cell 2008, 30, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Tchernyshyov, I.; Semenza, G.L.; Dang, C.V. HIF-1-mediated expression of pyruvate dehydrogenase kinase: A metabolic switch required for cellular adaptation to hypoxia. Cell Metab. 2006, 3, 177–185. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, R.; Zhang, H.; Kim, J.W.; Shimoda, L.; Dang, C.V.; Semenza, G.L. HIF-1 regulates cytochrome oxidase subunits to optimize efficiency of respiration in hypoxic cells. Cell 2007, 129, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, T.; Asano, Y.; Shintani, Y.; Aoyama, H.; Kioka, H.; Tsukamoto, O.; Hikita, M.; Shinzawa-Itoh, K.; Takafuji, K.; Higo, S.; et al. Higd1a is a positive regulator of cytochrome c oxidase. Proc. Natl. Acad. Sci. USA 2015, 112, 1553–1558. [Google Scholar] [CrossRef] [Green Version]
- Ramzan, R.; Vogt, S.; Kadenbach, B. Stress-mediated generation of deleterious ROS in healthy individuals—Role of cytochrome c oxidase. J. Mol. Med. 2020, 98, 651–657. [Google Scholar] [CrossRef] [Green Version]
- Lai, R.K.; Xu, I.M.; Chiu, D.K.; Tse, A.P.; Wei, L.L.; Law, C.T.; Lee, D.; Wong, C.M.; Wong, M.P.; Ng, I.O.; et al. NDUFA4L2 Fine-tunes Oxidative Stress in Hepatocellular Carcinoma. Clin. Cancer Res. 2016, 22, 3105–3117. [Google Scholar] [CrossRef] [Green Version]
- Spinelli, J.B.; Rosen, P.C.; Sprenger, H.G.; Puszynska, A.M.; Mann, J.L.; Roessler, J.M.; Cangelosi, A.L.; Henne, A.; Condon, K.J.; Zhang, T.; et al. Fumarate is a terminal electron acceptor in the mammalian electron transport chain. Science 2021, 374, 1227–1237. [Google Scholar] [CrossRef]
- Yoshikawa, S.; Shimada, A. Reaction mechanism of cytochrome c oxidase. Chem. Rev. 2015, 115, 1936–1989. [Google Scholar] [CrossRef]
- Ramzan, R.; Dolga, A.M.; Michels, S.; Weber, P.; Culmsee, C.; Rastan, A.J.; Vogt, S. Cytochrome c Oxidase Inhibition by ATP Decreases Mitochondrial ROS Production. Cells 2022, 11, 992. [Google Scholar] [CrossRef] [PubMed]
- Pitceathly, R.D.S.; Taanman, J.W. NDUFA4 (Renamed COXFA4) Is a Cytochrome-c Oxidase Subunit. Trends Endocrinol. Metab. 2018, 29, 452–454. [Google Scholar] [CrossRef] [PubMed]
- Clayton, S.A.; Daley, K.K.; MacDonald, L.; Fernandez-Vizarra, E.; Bottegoni, G.; O’Neil, J.D.; Major, T.; Griffin, D.; Zhuang, Q.; Adewoye, A.B.; et al. Inflammation causes remodeling of mitochondrial cytochrome c oxidase mediated by the bifunctional gene C15orf48. Sci. Adv. 2021, 7, eabl5182. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Nie, S.L.; Zhou, J.M.; Liu, F.; Hu, Y.B.; Jiang, J.R.; Li, N.; Liu, J.S. Overexpression of NDUFA4L2 is associated with poor prognosis in patients with colorectal cancer. ANZ J. Surg. 2017, 87, E251–E255. [Google Scholar] [CrossRef]
- Meng, L.; Yang, X.; Xie, X.; Wang, M. Mitochondrial NDUFA4L2 protein promotes the vitality of lung cancer cells by repressing oxidative stress. Thorac. Cancer 2019, 10, 676–685. [Google Scholar] [CrossRef] [PubMed]
- Burberry, A.; Wells, M.F.; Limone, F.; Couto, A.; Smith, K.S.; Keaney, J.; Gillet, G.; van Gastel, N.; Wang, J.Y.; Pietilainen, O.; et al. C9orf72 suppresses systemic and neural inflammation induced by gut bacteria. Nature 2020, 582, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Hedayati Moghadam, M.; Rezaee, S.A.R.; Hosseini, M.; Niazmand, S.; Salmani, H.; Rafatpanah, H.; Asarzadegan Dezfuli, M.; Amel Zabihi, N.; Abareshi, A.; Mahmoudabady, M. HTLV-1 infection-induced motor dysfunction, memory impairment, depression, and brain tissues oxidative damage in female BALB/c mice. Life Sci. 2018, 212, 9–19. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shao, Q.; Liu, J.; Li, G.; Gu, Y.; Guo, M.; Guan, Y.; Tian, Z.; Ma, W.; Wang, C.; Ji, X. Proteomic Analysis Reveals That Mitochondria Dominate the Hippocampal Hypoxic Response in Mice. Int. J. Mol. Sci. 2022, 23, 14094. https://doi.org/10.3390/ijms232214094
Shao Q, Liu J, Li G, Gu Y, Guo M, Guan Y, Tian Z, Ma W, Wang C, Ji X. Proteomic Analysis Reveals That Mitochondria Dominate the Hippocampal Hypoxic Response in Mice. International Journal of Molecular Sciences. 2022; 23(22):14094. https://doi.org/10.3390/ijms232214094
Chicago/Turabian StyleShao, Qianqian, Jia Liu, Gaifen Li, Yakun Gu, Mengyuan Guo, Yuying Guan, Zhengming Tian, Wei Ma, Chaoyu Wang, and Xunming Ji. 2022. "Proteomic Analysis Reveals That Mitochondria Dominate the Hippocampal Hypoxic Response in Mice" International Journal of Molecular Sciences 23, no. 22: 14094. https://doi.org/10.3390/ijms232214094
APA StyleShao, Q., Liu, J., Li, G., Gu, Y., Guo, M., Guan, Y., Tian, Z., Ma, W., Wang, C., & Ji, X. (2022). Proteomic Analysis Reveals That Mitochondria Dominate the Hippocampal Hypoxic Response in Mice. International Journal of Molecular Sciences, 23(22), 14094. https://doi.org/10.3390/ijms232214094