

Phytochromes and Their Role in Diurnal Variations of ROS Metabolism and Plant Proteome

 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Mutant Plants Were Slightly Paler, but Did Not Display Significant Differences as Compared to Col-0
2.2. Plant Analysis Did Not Show Striking Differences in Total Proteome Composition
2.3. Night–Day Variation in Plant Proteomes Highlighted Differences in Mutants
2.4. Light-Dependent Accumulation of Protein Found in Col-0 Was Predominantly Lost in All Mutant Genotypes
2.5. Mutation in the PhyB-Modulated Accumulation Patterns of Proteins Involved in Both Primary and Secondary Metabolism
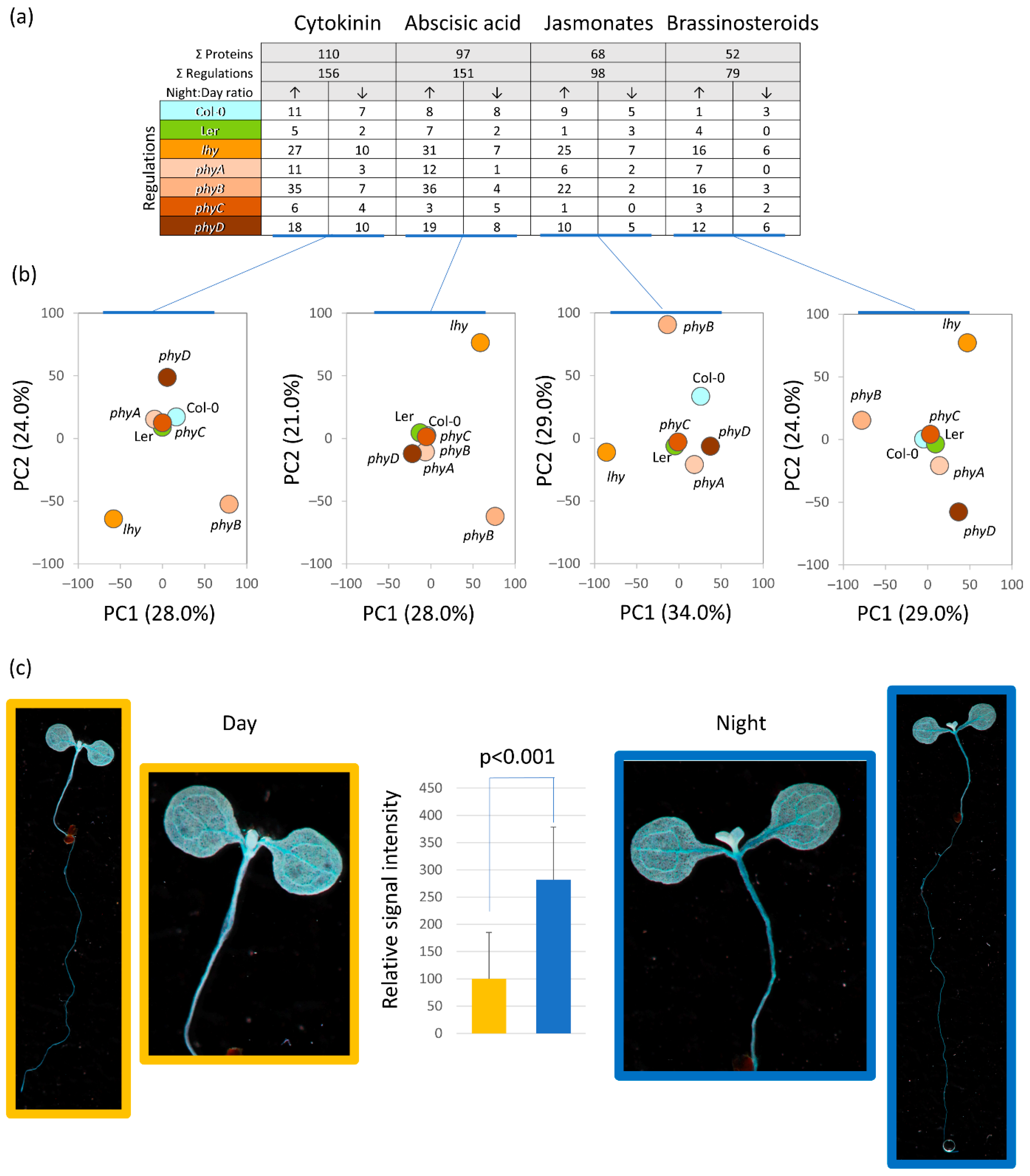
2.6. Mutation in LHY Impacted Hormone Metabolism and Signaling
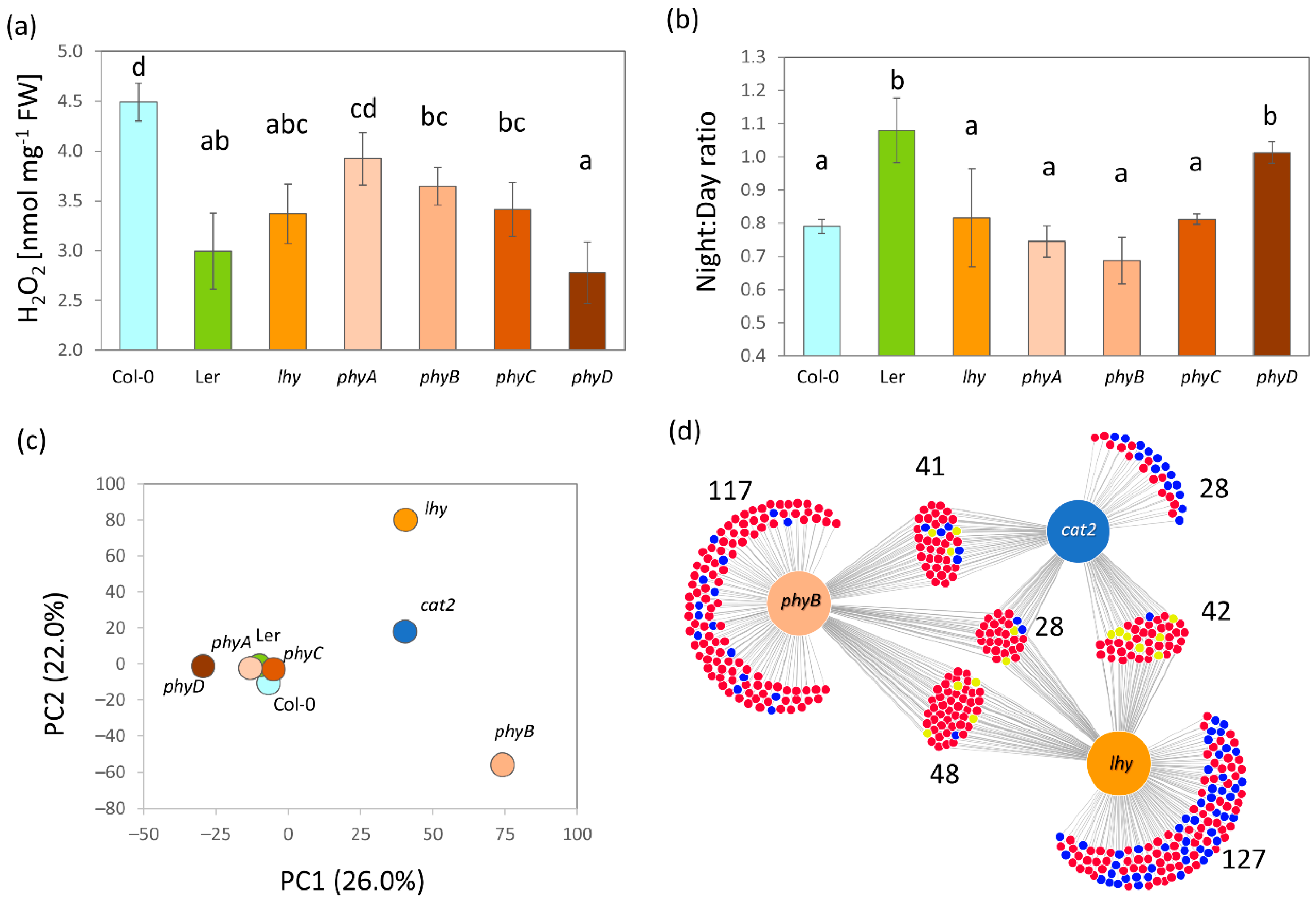
2.7. The Alteration in Metabolism Confirms the Critical Role of ROS in Night–Day Protein Accumulation Patterns
2.8. Cytokinin Signaling Affects Diurnal Variation in Plant Proteome
3. Discussion
3.1. Identification of Novel Targets of Diurnal Regulations
3.2. The Observed Variations in Mutant Proteomes Could Be Associated with a Disruption in Hormonal Metabolism and Signaling
3.3. ROS Metabolism Oscillation and Its Role in the Regulation of Plant Proteome
3.4. Implications of Observed Differences and Similarities in Mutant Genotypes
4. Materials and Methods
4.1. Plant Material
4.2. Plant Growth Conditions
4.3. Proteome Analysis
4.4. Hydrogen Peroxide Determination
4.5. GUS Activity Staining and Quantitation
4.6. Data Analysis and Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Covington, M.F.; Maloof, J.N.; Straume, M.; Kay, S.A.; Harmer, S.L. Global transcriptome analysis reveals circadian regulation of key pathways in plant growth and development. Genome Biol. 2008, 9, R130. [Google Scholar] [CrossRef] [Green Version]
- Hazen, S.P.; Naef, F.; Quisel, T.; Gendron, J.M.; Chen, H.; Ecker, J.R.; Borevitz, J.O.; Kay, S.A. Exploring the transcriptional landscape of plant circadian rhythms using genome tiling arrays. Genome Biol. 2009, 10, R17. [Google Scholar] [CrossRef] [Green Version]
- Paajanen, P.; Lane de Barros Dantas, L.; Dodd, A.N. Layers of crosstalk between circadian regulation and environmental signalling in plants. Curr. Biol. 2021, 31, R399–R413. [Google Scholar] [CrossRef]
- Harmer, S.L. The Circadian System in Higher Plants. Annu. Rev. Plant Biol. 2009, 60, 357–377. [Google Scholar] [CrossRef] [Green Version]
- Gil, K.; Park, C. Thermal adaptation and plasticity of the plant circadian clock. New Phytol. 2019, 221, 1215–1229. [Google Scholar] [CrossRef] [Green Version]
- Dubois, M.; Claeys, H.; Van den Broeck, L.; Inzé, D. Time of day determines Arabidopsis transcriptome and growth dynamics under mild drought. Plant. Cell Environ. 2017, 40, 180–189. [Google Scholar] [CrossRef] [Green Version]
- Blair, E.J.; Bonnot, T.; Hummel, M.; Hay, E.; Marzolino, J.M.; Quijada, I.A.; Nagel, D.H. Contribution of time of day and the circadian clock to the heat stress responsive transcriptome in Arabidopsis. Sci. Rep. 2019, 9, 4814. [Google Scholar] [CrossRef] [Green Version]
- Hua, J. Modulation of plant immunity by light, circadian rhythm, and temperature. Curr. Opin. Plant Biol. 2013, 16, 406–413. [Google Scholar] [CrossRef]
- Hermans, C.; Vuylsteke, M.; Coppens, F.; Craciun, A.; Inzé, D.; Verbruggen, N. Early transcriptomic changes induced by magnesium deficiency in Arabidopsis thaliana reveal the alteration of circadian clock gene expression in roots and the triggering of abscisic acid-responsive genes. New Phytol. 2010, 187, 119–131. [Google Scholar] [CrossRef]
- Chen, Y.-Y.; Wang, Y.; Shin, L.-J.; Wu, J.-F.; Shanmugam, V.; Tsednee, M.; Lo, J.-C.; Chen, C.-C.; Wu, S.-H.; Yeh, K.-C. Iron Is Involved in the Maintenance of Circadian Period Length in Arabidopsis. Plant Physiol. 2013, 161, 1409–1420. [Google Scholar] [CrossRef]
- Kolmos, E.; Chow, B.Y.; Pruneda-Paz, J.L.; Kay, S.A. HsfB2b-mediated repression of PRR7 directs abiotic stress responses of the circadian clock. Proc. Natl. Acad. Sci. USA 2014, 111, 16172–16177. [Google Scholar] [CrossRef] [Green Version]
- Marcolino-Gomes, J.; Rodrigues, F.A.; Fuganti-Pagliarini, R.; Bendix, C.; Nakayama, T.J.; Celaya, B.; Molinari, H.B.C.; de Oliveira, M.C.N.; Harmon, F.G.; Nepomuceno, A. Diurnal Oscillations of Soybean Circadian Clock and Drought Responsive Genes. PLoS ONE 2014, 9, e86402. [Google Scholar] [CrossRef] [Green Version]
- Staiger, D.; Köster, T. Spotlight on post-transcriptional control in the circadian system. Cell. Mol. Life Sci. 2011, 68, 71–83. [Google Scholar] [CrossRef]
- Romanowski, A.; Schlaen, R.G.; Perez-Santangelo, S.; Mancini, E.; Yanovsky, M.J. Global transcriptome analysis reveals circadian control of splicing events in Arabidopsis thaliana. Plant J. 2020, 103, 889–902. [Google Scholar] [CrossRef]
- Vogel, C.; Marcotte, E.M. Insights into the regulation of protein abundance from proteomic and transcriptomic analyses. Nat. Rev. Genet. 2012, 13, 227–232. [Google Scholar] [CrossRef] [Green Version]
- Hwang, H.; Cho, M.-H.; Hahn, B.-S.; Lim, H.; Kwon, Y.-K.; Hahn, T.-R.; Bhoo, S.H. Proteomic identification of rhythmic proteins in rice seedlings. Biochim. Biophys. Acta Proteins Proteom. 2011, 1814, 470–479. [Google Scholar] [CrossRef]
- Choudhary, M.K.; Nomura, Y.; Shi, H.; Nakagami, H.; Somers, D.E. Circadian Profiling of the Arabidopsis Proteome Using 2D-DIGE. Front. Plant Sci. 2016, 7, 1007. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, M.K.; Nomura, Y.; Wang, L.; Nakagami, H.; Somers, D.E. Quantitative Circadian Phosphoproteomic Analysis of Arabidopsis Reveals Extensive Clock Control of Key Components in Physiological, Metabolic, and Signaling Pathways. Mol. Cell. Proteom. 2015, 14, 2243–2260. [Google Scholar] [CrossRef] [Green Version]
- Graf, A.; Coman, D.; Uhrig, R.G.; Walsh, S.; Flis, A.; Stitt, M.; Gruissem, W. Parallel analysis of Arabidopsis circadian clock mutants reveals different scales of transcriptome and proteome regulation. Open Biol. 2017, 7, 160333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seaton, D.D.; Graf, A.; Baerenfaller, K.; Stitt, M.; Millar, A.J.; Gruissem, W. Photoperiodic control of the Arabidopsis proteome reveals a translational coincidence mechanism. Mol. Syst. Biol. 2018, 14, e7962. [Google Scholar] [CrossRef]
- Uhrig, R.G.; Echevarría-Zomeño, S.; Schlapfer, P.; Grossmann, J.; Roschitzki, B.; Koerber, N.; Fiorani, F.; Gruissem, W. Diurnal dynamics of the Arabidopsis rosette proteome and phosphoproteome. Plant. Cell Environ. 2021, 44, 821–841. [Google Scholar] [CrossRef] [PubMed]
- Verhage, L. Isotope labeling to measure protein synthesis rates throughout the diurnal cycle—The technique explained. Plant J. 2022, 109, 743–744. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Yu, Y.; Wang, X.; Qin, Y.; Su, C.; Wang, L. Aschoff’s rule on circadian rhythms orchestrated by blue light sensor CRY2 and clock component PRR9. Nat. Commun. 2022, 13, 5869. [Google Scholar] [CrossRef] [PubMed]
- Oakenfull, R.J.; Davis, S.J. Shining a light on the Arabidopsis circadian clock. Plant. Cell Environ. 2017, 40, 2571–2585. [Google Scholar] [CrossRef] [Green Version]
- Legris, M.; Ince, Y.Ç.; Fankhauser, C. Molecular mechanisms underlying phytochrome-controlled morphogenesis in plants. Nat. Commun. 2019, 10, 5219. [Google Scholar] [CrossRef] [Green Version]
- Schaffer, R.; Ramsay, N.; Samach, A.; Corden, S.; Putterill, J.; Carré, I.A.; Coupland, G. The late elongated hypocotyl Mutation of Arabidopsis Disrupts Circadian Rhythms and the Photoperiodic Control of Flowering. Cell 1998, 93, 1219–1229. [Google Scholar] [CrossRef]
- Park, M.-J.; Kwon, Y.-J.; Gil, K.-E.; Park, C.-M. LATE ELONGATED HYPOCOTYL regulates photoperiodic flowering via the circadian clock in Arabidopsis. BMC Plant Biol. 2016, 16, 114. [Google Scholar] [CrossRef] [Green Version]
- Kyung, J.; Jeon, M.; Jeong, G.; Shin, Y.; Seo, E.; Yu, J.; Kim, H.; Park, C.-M.; Hwang, D.; Lee, I. The two clock proteins CCA1 and LHY activate VIN3 transcription during vernalization through the vernalization-responsive cis-element. Plant Cell 2022, 34, 1020–1037. [Google Scholar] [CrossRef]
- Yang, S.W.; Jang, I.-C.; Henriques, R.; Chua, N.-H. FAR-RED ELONGATED HYPOCOTYL1 and FHY1-LIKE Associate with the Arabidopsis Transcription Factors LAF1 and HFR1 to Transmit Phytochrome A Signals for Inhibition of Hypocotyl Elongation. Plant Cell 2009, 21, 1341–1359. [Google Scholar] [CrossRef] [Green Version]
- Nagatani, A.; Reed, J.W.; Chory, J. Isolation and Initial Characterization of Arabidopsis Mutants That Are Deficient in Phytochrome A. Plant Physiol. 1993, 102, 269–277. [Google Scholar] [CrossRef]
- Salomé, P.A.; Michael, T.P.; Kearns, E.V.; Fett-Neto, A.G.; Sharrock, R.A.; McClung, C.R. The out of phase 1 Mutant Defines a Role for PHYB in Circadian Phase Control in Arabidopsis. Plant Physiol. 2002, 129, 1674–1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, J.-H.; Domijan, M.; Klose, C.; Biswas, S.; Ezer, D.; Gao, M.; Khattak, A.K.; Box, M.S.; Charoensawan, V.; Cortijo, S.; et al. Phytochromes function as thermosensors in Arabidopsis. Science 2016, 354, 886–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arico, D.; Legris, M.; Castro, L.; Garcia, C.F.; Laino, A.; Casal, J.J.; Mazzella, M.A. Neighbour signals perceived by phytochrome B increase thermotolerance in Arabidopsis. Plant. Cell Environ. 2019, 42, 2554–2566. [Google Scholar] [CrossRef] [PubMed]
- Monte, E.; Alonso, J.M.; Ecker, J.R.; Zhang, Y.; Li, X.; Young, J.; Austin-Phillips, S.; Quail, P.H. Isolation and Characterization of phyC Mutants in Arabidopsis Reveals Complex Crosstalk between Phytochrome Signaling Pathways. Plant Cell 2003, 15, 1962–1980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, K.D.; Guerineau, F.; Devlin, P.F.; Millar, A.J. Low-temperature-specific effects of PHYTOCHROME C on the circadian clock in Arabidopsis suggest that PHYC underlies natural variation in biological timing. bioRxiv 2015, 30577. [Google Scholar] [CrossRef] [Green Version]
- Devlin, P.F.; Robson, P.R.H.; Patel, S.R.; Goosey, L.; Sharrock, R.A.; Whitelam, G.C. Phytochrome D Acts in the Shade-Avoidance Syndrome in Arabidopsis by Controlling Elongation Growth and Flowering Time1. Plant Physiol. 1999, 119, 909–916. [Google Scholar] [CrossRef] [Green Version]
- Aukerman, M.J.; Hirschfeld, M.; Wester, L.; Weaver, M.; Clack, T.; Amasino, R.M.; Sharrock, R.A. A deletion in the PHYD gene of the Arabidopsis Wassilewskija ecotype defines a role for phytochrome D in red/far-red light sensing. Plant Cell 1997, 9, 1317–1326. [Google Scholar] [CrossRef]
- Tóth, R.; Kevei, E.; Hall, A.; Millar, A.J.; Nagy, F.; Kozma-Bognár, L. Circadian Clock-Regulated Expression of Phytochrome and Cryptochrome Genes in Arabidopsis. Plant Physiol. 2001, 127, 1607–1616. [Google Scholar] [CrossRef]
- Ferrari, C.; Proost, S.; Janowski, M.; Becker, J.; Nikoloski, Z.; Bhattacharya, D.; Price, D.; Tohge, T.; Bar-Even, A.; Fernie, A.; et al. Kingdom-wide comparison reveals the evolution of diurnal gene expression in Archaeplastida. Nat. Commun. 2019, 10, 737. [Google Scholar] [CrossRef] [Green Version]
- Kircher, S.; Gil, P.; Kozma-Bognár, L.; Fejes, E.; Speth, V.; Husselstein-Muller, T.; Bauer, D.; Ádám, É.; Schäfer, E.; Nagy, F. Nucleocytoplasmic Partitioning of the Plant Photoreceptors Phytochrome A, B, C, D, and E Is Regulated Differentially by Light and Exhibits a Diurnal Rhythm. Plant Cell 2002, 14, 1541–1555. [Google Scholar] [CrossRef]
- Liu, Y.; Sun, Y.; Yao, H.; Zheng, Y.; Cao, S.; Wang, H. Arabidopsis Circadian Clock Repress Phytochrome A Signaling. Front. Plant Sci. 2022, 13, 809563. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, A.; Nagel, M.-K.; Popp, C.; Wüst, F.; Bindics, J.; Viczián, A.; Hiltbrunner, A.; Nagy, F.; Kunkel, T.; Schäfer, E. Interaction with plant transcription factors can mediate nuclear import of phytochrome B. Proc. Natl. Acad. Sci. USA 2012, 109, 5892–5897. [Google Scholar] [CrossRef] [Green Version]
- Fulton, D.C.; Stettler, M.; Mettler, T.; Vaughan, C.K.; Li, J.; Francisco, P.; Gil, M.; Reinhold, H.; Eicke, S.; Messerli, G.; et al. β-AMYLASE4, a Noncatalytic Protein Required for Starch Breakdown, Acts Upstream of Three Active β-Amylases in Arabidopsis Chloroplasts. Plant Cell 2008, 20, 1040–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charron, J.-B.F.; Ouellet, F.; Houde, M.; Sarhan, F. The plant Apolipoprotein D ortholog protects Arabidopsis against oxidative stress. BMC Plant Biol. 2008, 8, 86. [Google Scholar] [CrossRef] [Green Version]
- Chi, W.-T.; Fung, R.W.M.; Liu, H.-C.; Hsu, C.-C.; Charng, Y.-Y. Temperature-induced lipocalin is required for basal and acquired thermotolerance in Arabidopsis. Plant. Cell Environ. 2009, 32, 917–927. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.H.; Piao, H.L.; Kim, H.-Y.; Choi, S.M.; Jiang, F.; Hartung, W.; Hwang, I.; Kwak, J.M.; Lee, I.-J.; Hwang, I. Activation of Glucosidase via Stress-Induced Polymerization Rapidly Increases Active Pools of Abscisic Acid. Cell 2006, 126, 1109–1120. [Google Scholar] [CrossRef] [Green Version]
- Larkin, R.M.; Stefano, G.; Ruckle, M.E.; Stavoe, A.K.; Sinkler, C.A.; Brandizzi, F.; Malmstrom, C.M.; Osteryoung, K.W. REDUCED CHLOROPLAST COVERAGE genes from Arabidopsis thaliana help to establish the size of the chloroplast compartment. Proc. Natl. Acad. Sci. USA 2016, 113, E1116–E1125. [Google Scholar] [CrossRef] [Green Version]
- Papp, I.; Mur, L.; Dalmadi, Á.; Dulai, S.; Koncz, C. A mutation in the Cap Binding Protein 20 gene confers drought. Plant Mol. Biol. 2004, 55, 679–686. [Google Scholar] [CrossRef]
- Luo, Y.; Wang, Z.; Ji, H.; Fang, H.; Wang, S.; Tian, L.; Li, X. An Arabidopsis homolog of importin β1 is required for ABA response and drought tolerance. Plant J. 2013, 75, 377–389. [Google Scholar] [CrossRef]
- Zheng, B.S.; Rönnberg, E.; Viitanen, L.; Salminen, T.A.; Lundgren, K.; Moritz, T.; Edqvist, J. Arabidopsis sterol carrier protein-2 is required for normal development of seeds and seedlings. J. Exp. Bot. 2008, 59, 3485–3499. [Google Scholar] [CrossRef]
- Diaz, C.; Kusano, M.; Sulpice, R.; Araki, M.; Redestig, H.; Saito, K.; Stitt, M.; Shin, R. Determining novel functions of Arabidopsis14-3-3 proteins in central metabolic processes. BMC Syst. Biol. 2011, 5, 192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Link, S.; Engelmann, K.; Meierhoff, K.; Westhoff, P. The Atypical Short-Chain Dehydrogenases HCF173 and HCF244 Are Jointly Involved in Translational Initiation of the psbA mRNA of Arabidopsis. Plant Physiol. 2012, 160, 2202–2218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christians, M.J.; Larsen, P.B. Mutational loss of the prohibitin AtPHB3 results in an extreme constitutive ethylene response phenotype coupled with partial loss of ethylene-inducible gene expression in Arabidopsis seedlings. J. Exp. Bot. 2007, 58, 2237–2248. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Ries, A.; Wu, K.; Yang, A.; Crawford, N.M. The Arabidopsis Prohibitin Gene PHB3 Functions in Nitric Oxide–Mediated Responses and in Hydrogen Peroxide–Induced Nitric Oxide Accumulation. Plant Cell 2010, 22, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Clark, S.M.; Di Leo, R.; Dhanoa, P.K.; Van Cauwenberghe, O.R.; Mullen, R.T.; Shelp, B.J. Biochemical characterization, mitochondrial localization, expression, and potential functions for an Arabidopsis γ-aminobutyrate transaminase that utilizes both pyruvate and glyoxylate. J. Exp. Bot. 2009, 60, 1743–1757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, G.; Kim, H.; Kim, D.H.; Hanh, H.; Yoon, Y.; Singaram, I.; Wijesinghe, K.J.; Johnson, K.A.; Zhuang, X.; Liang, Z.; et al. SH3 Domain-Containing Protein 2 Plays a Crucial Role at the Step of Membrane Tubulation during Cell Plate Formation. Plant Cell 2017, 29, 1388–1405. [Google Scholar] [CrossRef] [Green Version]
- Niehaus, T.D.; Patterson, J.A.; Alexander, D.C.; Folz, J.S.; Pyc, M.; MacTavish, B.S.; Bruner, S.D.; Mullen, R.T.; Fiehn, O.; Hanson, A.D. The metabolite repair enzyme Nit1 is a dual-targeted amidase that disposes of damaged glutathione in Arabidopsis. Biochem. J. 2019, 476, 683–697. [Google Scholar] [CrossRef]
- Sibout, R.; Eudes, A.; Mouille, G.; Pollet, B.; Lapierre, C.; Jouanin, L.; Séguin, A. CINNAMYL ALCOHOL DEHYDROGENASE-C and -D Are the Primary Genes Involved in Lignin Biosynthesis in the Floral Stem of Arabidopsis. Plant Cell 2005, 17, 2059–2076. [Google Scholar] [CrossRef] [Green Version]
- Burow, M.; Zhang, Z.-Y.; Ober, J.A.; Lambrix, V.M.; Wittstock, U.; Gershenzon, J.; Kliebenstein, D.J. ESP and ESM1 mediate indol-3-acetonitrile production from indol-3-ylmethyl glucosinolate in Arabidopsis. Phytochemistry 2008, 69, 663–671. [Google Scholar] [CrossRef]
- Černý, M.; Novák, J.; Habánová, H.; Cerna, H.; Brzobohatý, B. Role of the proteome in phytohormonal signaling. Biochim. Biophys. Acta Proteins Proteom. 2016, 1864, 1003–1015. [Google Scholar] [CrossRef]
- Ancín, M.; Fernandez-Irigoyen, J.; Santamaria, E.; Larraya, L.; Fernández-San Millán, A.; Veramendi, J.; Farran, I. New In Vivo Approach to Broaden the Thioredoxin Family Interactome in Chloroplasts. Antioxidants 2022, 11, 1979. [Google Scholar] [CrossRef] [PubMed]
- Mergner, J.; Frejno, M.; List, M.; Papacek, M.; Chen, X.; Chaudhary, A.; Samaras, P.; Richter, S.; Shikata, H.; Messerer, M.; et al. Mass-spectrometry-based draft of the Arabidopsis proteome. Nature 2020, 579, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.L.; Nemhauser, J.L. Do Trees Grow on Money? Auxin as the Currency of the Cellular Economy. Cold Spring Harb. Perspect. Biol. 2010, 2, a001420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, M.; Mas, P. A Functional Connection between the Circadian Clock and Hormonal Timing in Arabidopsis. Genes 2018, 9, 567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawat, R.; Schwartz, J.; Jones, M.A.; Sairanen, I.; Cheng, Y.; Andersson, C.R.; Zhao, Y.; Ljung, K.; Harmer, S.L. REVEILLE1, a Myb-like transcription factor, integrates the circadian clock and auxin pathways. Proc. Natl. Acad. Sci. USA 2009, 106, 16883–16888. [Google Scholar] [CrossRef] [Green Version]
- Covington, M.F.; Harmer, S.L. The Circadian Clock Regulates Auxin Signaling and Responses in Arabidopsis. PLoS Biol. 2007, 5, e222. [Google Scholar] [CrossRef] [Green Version]
- Voß, U.; Wilson, M.H.; Kenobi, K.; Gould, P.D.; Robertson, F.C.; Peer, W.A.; Lucas, M.; Swarup, K.; Casimiro, I.; Holman, T.J.; et al. The circadian clock rephases during lateral root organ initiation in Arabidopsis thaliana. Nat. Commun. 2015, 6, 7641. [Google Scholar] [CrossRef] [Green Version]
- Adams, S.; Grundy, J.; Veflingstad, S.R.; Dyer, N.P.; Hannah, M.A.; Ott, S.; Carré, I.A. Circadian control of abscisic acid biosynthesis and signalling pathways revealed by genome-wide analysis of LHY binding targets. New Phytol. 2018, 220, 893–907. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Takahashi, D.; Kawamura, Y.; Uemura, M. Comparison of Plasma Membrane Proteomic Changes of Arabidopsis Suspension-Cultured Cells (T87 Line) after Cold and ABA Treatment in Association with Freezing Tolerance Development. Plant Cell Physiol. 2012, 53, 543–554. [Google Scholar] [CrossRef] [Green Version]
- Ezer, D.; Jung, J.-H.; Lan, H.; Biswas, S.; Gregoire, L.; Box, M.S.; Charoensawan, V.; Cortijo, S.; Lai, X.; Stöckle, D.; et al. The evening complex coordinates environmental and endogenous signals in Arabidopsis. Nat. Plants 2017, 3, 17087. [Google Scholar] [CrossRef]
- Nováková, M.; Motyka, V.; Dobrev, P.I.; Malbeck, J.; Gaudinová, A.; Vanková, R. Diurnal variation of cytokinin, auxin and abscisic acid levels in tobacco leaves. J. Exp. Bot. 2005, 56, 2877–2883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanano, S.; Domagalska, M.A.; Nagy, F.; Davis, S.J. Multiple phytohormones influence distinct parameters of the plant circadian clock. Genes Cells 2006, 11, 1381–1392. [Google Scholar] [CrossRef] [PubMed]
- Salome, P.A.; To, J.P.C.; Kieber, J.J.; McClung, R. Arabidopsis Response Regulators ARR3 and ARR4 Play Cytokinin-Independent Roles in the Control of Circadian Period. Plant Cell Online 2006, 18, 55–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nitschke, S.; Cortleven, A.; Iven, T.; Feussner, I.; Havaux, M.; Riefler, M.; Schmülling, T. Circadian Stress Regimes Affect the Circadian Clock and Cause Jasmonic Acid-Dependent Cell Death in Cytokinin-Deficient Arabidopsis Plants. Plant Cell 2016, 28, 1616–1639. [Google Scholar] [CrossRef] [Green Version]
- Pavlů, J.; Novák, J.; Koukalová, V.; Luklová, M.; Brzobohatý, B.; Černý, M. Cytokinin at the Crossroads of Abiotic Stress Signalling Pathways. Int. J. Mol. Sci. 2018, 19, 2450. [Google Scholar] [CrossRef] [Green Version]
- Zheng, B.; Deng, Y.; Mu, J.; Ji, Z.; Xiang, T.; Niu, Q.-W.; Chua, N.-H.; Zuo, J. Cytokinin affects circadian-clock oscillation in a phytochrome B- and Arabidopsis response regulator 4-dependent manner. Physiol. Plant. 2006, 127, 277–292. [Google Scholar] [CrossRef]
- Dobisova, T.; Hrdinova, V.; Cuesta, C.; Michlickova, S.; Urbankova, I.; Hejatkova, R.; Zadnikova, P.; Pernisova, M.; Benkova, E.; Hejatko, J. Light Controls Cytokinin Signaling via Transcriptional Regulation of Constitutively Active Sensor Histidine Kinase CKI1. Plant Physiol. 2017, 174, 387–404. [Google Scholar] [CrossRef] [Green Version]
- Mizoguchi, T.; Wheatley, K.; Hanzawa, Y.; Wright, L.; Mizoguchi, M.; Song, H.-R.; Carré, I.A.; Coupland, G. LHY and CCA1 Are Partially Redundant Genes Required to Maintain Circadian Rhythms in Arabidopsis. Dev. Cell 2002, 2, 629–641. [Google Scholar] [CrossRef] [Green Version]
- Černý, M.; Habánová, H.; Berka, M.; Luklová, M.; Brzobohatý, B. Hydrogen Peroxide: Its Role in Plant Biology and Crosstalk with Signalling Networks. Int. J. Mol. Sci. 2018, 19, 2812. [Google Scholar] [CrossRef] [Green Version]
- Lai, A.G.; Doherty, C.J.; Mueller-Roeber, B.; Kay, S.A.; Schippers, J.H.M.; Dijkwel, P.P. CIRCADIAN CLOCK-ASSOCIATED 1 regulates ROS homeostasis and oxidative stress responses. Proc. Natl. Acad. Sci. USA 2012, 109, 17129–17134. [Google Scholar] [CrossRef]
- Román, Á.; Li, X.; Deng, D.; Davey, J.W.; James, S.; Graham, I.A.; Haydon, M.J. Superoxide is promoted by sucrose and affects amplitude of circadian rhythms in the evening. Proc. Natl. Acad. Sci. USA 2021, 118, e2020646118. [Google Scholar] [CrossRef]
- Gallé, Á.; Czékus, Z.; Bela, K.; Horváth, E.; Ördög, A.; Csiszár, J.; Poór, P. Plant Glutathione Transferases and Light. Front. Plant Sci. 2019, 9, 1944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallé, Á.; Czékus, Z.; Bela, K.; Horváth, E.; Csiszár, J.; Poór, P. Diurnal changes in tomato glutathione transferase activity and expression. Acta Biol. Hung. 2018, 69, 505–509. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.-W.; Liu, M.-J.; Chen, I.-C.; Huang, C.-H.; Chao, L.-Y.; Hsieh, H.-L. A Glutathione S -Transferase Regulated by Light and Hormones Participates in the Modulation of Arabidopsis Seedling Development. Plant Physiol. 2010, 154, 1646–1658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zechmann, B. Diurnal changes of subcellular glutathione content in Arabidopsis thaliana. Biol. Plant. 2017, 61, 791–796. [Google Scholar] [CrossRef]
- Pavlů, J.; Kerchev, P.; Černý, M.; Novák, J.; Berka, M.; Jobe, T.O.; López Ramos, J.M.; Saiz-Fernández, I.; Michael Rashotte, A.; Kopriva, S.; et al. Cytokinin modulates sulfur and glutathione metabolic network. J. Exp. Bot. 2022. [Google Scholar] [CrossRef]
- van Zanten, M.; Snoek, L.B.; Proveniers, M.C.G.; Peeters, A.J.M. The many functions of ERECTA. Trends Plant Sci. 2009, 14, 214–218. [Google Scholar] [CrossRef]
- McCarthy, A.; Chung, M.; Ivanov, A.G.; Krol, M.; Inman, M.; Maxwell, D.P.; Hüner, N.P.A. An established Arabidopsis thaliana var. Landsberg erecta cell suspension culture accumulates chlorophyll and exhibits a stay-green phenotype in response to high external sucrose concentrations. J. Plant Physiol. 2016, 199, 40–51. [Google Scholar] [CrossRef]
- Li, X.; Wang, H.; Wang, Y.; Zhang, L.; Wang, Y. Comparison of Metabolic Profiling of Arabidopsis Inflorescences Between Landsberg erecta and Columbia, and Meiosis-Defective Mutants by 1H-NMR Spectroscopy. Phenomics 2021, 1, 73–89. [Google Scholar] [CrossRef]
- Burgie, E.S.; Gannam, Z.T.K.; McLoughlin, K.E.; Sherman, C.D.; Holehouse, A.S.; Stankey, R.J.; Vierstra, R.D. Differing biophysical properties underpin the unique signaling potentials within the plant phytochrome photoreceptor families. Proc. Natl. Acad. Sci. USA 2021, 118, e2105649118. [Google Scholar] [CrossRef]
- Ha, J.-H.; Kim, J.-H.; Kim, S.-G.; Sim, H.-J.; Lee, G.; Halitschke, R.; Baldwin, I.T.; Kim, J.-I.; Park, C.-M. Shoot phytochrome B modulates reactive oxygen species homeostasis in roots via abscisic acid signaling in Arabidopsis. Plant J. 2018, 94, 790–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berka, M.; Luklová, M.; Dufková, H.; Malých, V.; Novák, J.; Saiz-Fernández, I.; Rashotte, A.M.; Brzobohaty, B.; Cerny, M. Barley root proteome and metabolome in response to cytokinin and abiotic stimuli. Front. Plant Sci. 2020, 11, 1647. [Google Scholar] [CrossRef] [PubMed]
- Dufková, H.; Berka, M.; Luklová, M.; Rashotte, A.M.; Brzobohatý, B.; Černý, M. Eggplant Germination is Promoted by Hydrogen Peroxide and Temperature in an Independent but Overlapping Manner. Molecules 2019, 24, 4270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berková, V.; Kameniarová, M.; Ondrisková, V.; Berka, M.; Menšíková, S.; Kopecká, R.; Luklová, M.; Novák, J.; Spíchal, L.; Rashotte, A.M.; et al. Arabidopsis Response to Inhibitor of Cytokinin Degradation INCYDE: Modulations of Cytokinin Signaling and Plant Proteome. Plants 2020, 9, 1563. [Google Scholar] [CrossRef] [PubMed]
- Krishnakumar, V.; Contrino, S.; Cheng, C.Y.; Belyaeva, I.; Ferlanti, E.S.; Miller, J.R.; Vaughn, M.W.; Micklem, G.; Town, C.D.; Chan, A.P. Thalemine: A warehouse for Arabidopsis data integration and discovery. Plant Cell Physiol. 2017, 58, e4. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Béziat, C.; Kleine-Vehn, J.; Feraru, E. Histochemical staining of β-glucuronidase and its spatial quantification. Methods Mol. Biol. 2017, 1497, 73–80. [Google Scholar] [CrossRef]
- Pang, Z.; Chong, J.; Zhou, G.; De Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef]
- Liebermeister, W.; Noor, E.; Flamholz, A.; Davidi, D.; Bernhardt, J.; Milo, R. Visual account of protein investment in cellular functions. Proc. Natl. Acad. Sci. USA 2014, 111, 8488–8493. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Dong, S.; Ge, Y.; Fonseca, J.P.; Robinson, Z.T.; Mysore, K.S.; Mehta, P. DiVenn: An interactive and integrated web-based visualization tool for comparing gene lists. Front. Genet. 2021, 10, 421. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luklová, M.; Novák, J.; Kopecká, R.; Kameniarová, M.; Gibasová, V.; Brzobohatý, B.; Černý, M. Phytochromes and Their Role in Diurnal Variations of ROS Metabolism and Plant Proteome. Int. J. Mol. Sci. 2022, 23, 14134. https://doi.org/10.3390/ijms232214134
Luklová M, Novák J, Kopecká R, Kameniarová M, Gibasová V, Brzobohatý B, Černý M. Phytochromes and Their Role in Diurnal Variations of ROS Metabolism and Plant Proteome. International Journal of Molecular Sciences. 2022; 23(22):14134. https://doi.org/10.3390/ijms232214134
Chicago/Turabian StyleLuklová, Markéta, Jan Novák, Romana Kopecká, Michaela Kameniarová, Vladěna Gibasová, Břetislav Brzobohatý, and Martin Černý. 2022. "Phytochromes and Their Role in Diurnal Variations of ROS Metabolism and Plant Proteome" International Journal of Molecular Sciences 23, no. 22: 14134. https://doi.org/10.3390/ijms232214134
APA StyleLuklová, M., Novák, J., Kopecká, R., Kameniarová, M., Gibasová, V., Brzobohatý, B., & Černý, M. (2022). Phytochromes and Their Role in Diurnal Variations of ROS Metabolism and Plant Proteome. International Journal of Molecular Sciences, 23(22), 14134. https://doi.org/10.3390/ijms232214134