
Identification of Canine Pyometra-Associated Metabolites Using Untargeted Metabolomics
Abstract
:1. Introduction
2. Results
2.1. Clinical Evaluation of Dogs
2.2. Ultrasonography and Histopathology
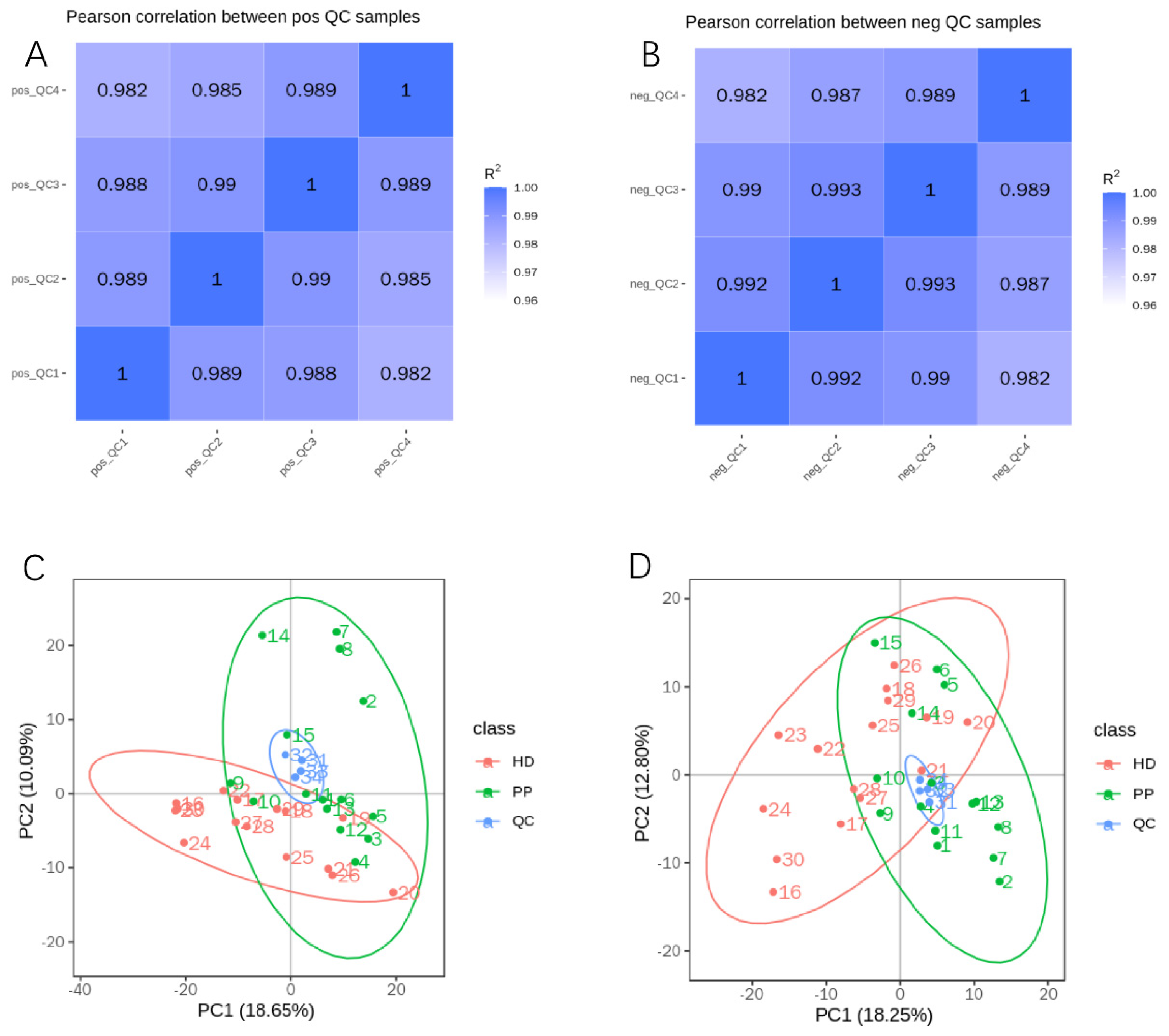
2.3. Quality Control of Untargeted Metabolic Profiling
2.4. Untargeted Metabolic Profiling of Endometrium in Dogs with Pyometra
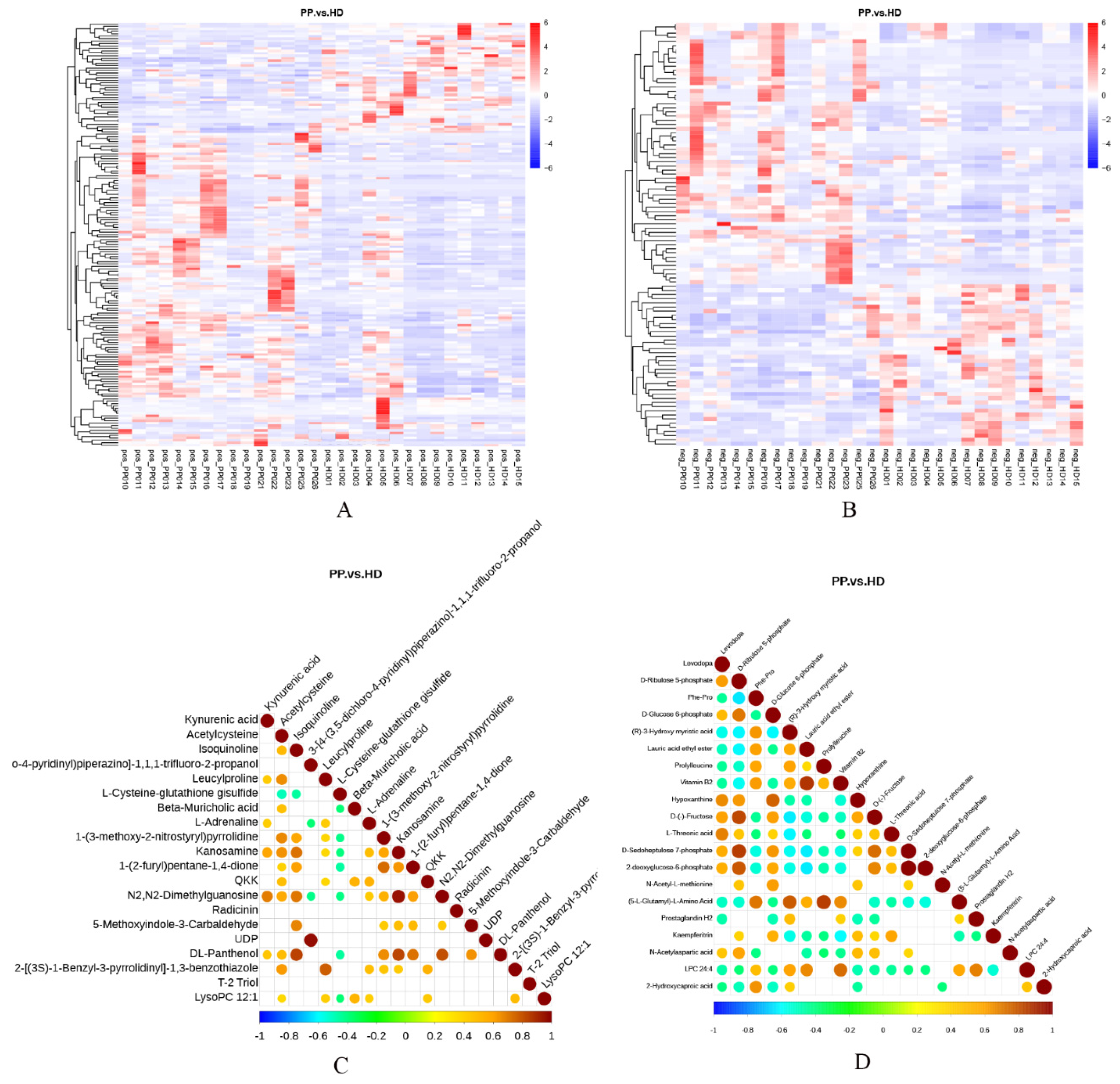
2.5. Screening of Differential Metabolites of Endometrium in Dogs with Pyometra
2.6. Analysis of Differential Metabolites
2.7. Metabolic Pathway Analysis
2.8. Receiver Operating Characteristic of Differential Metabolites Related to the Canine Pyometra
3. Discussion
4. Materials and Methods
4.1. Animals and Samplings
4.2. Ultrasonography Examinations
4.3. Blood Examinations
4.4. Histopathology Examinations
4.5. Metabolites Extraction
4.6. UHPLC-MS/MS Analysis
4.7. Data Processing and Metabolite Identification
4.8. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hagman, R. Pyometra in Small Animals. Vet. Clin. N. Am. Small Anim. Pr. 2018, 48, 639–661. [Google Scholar] [CrossRef] [PubMed]
- Jitpean, S.; Hagman, R.; Ström Holst, B.; Höglund, O.V.; Pettersson, A.; Egenvall, A. Breed variations in the incidence of pyometra and mammary tumours in Swedish dogs. Reprod. Domest. Anim. 2012, 47 (Suppl. S6), 347–350. [Google Scholar] [CrossRef] [PubMed]
- Jitpean, S.; Ström-Holst, B.; Emanuelson, U.; Höglund, O.V.; Pettersson, A.; Alneryd-Bull, C.; Hagman, R. Outcome of pyometra in female dogs and predictors of peritonitis and prolonged postoperative hospitalization in surgically treated cases. BMC Vet. Res. 2014, 10, 6. [Google Scholar] [CrossRef] [Green Version]
- Smith, F.O. Canine pyometra. Theriogenology 2006, 66, 610–612. [Google Scholar] [CrossRef] [PubMed]
- Quartuccio, M.; Liotta, L.; Cristarella, S.; Lanteri, G.; Ieni, A.; D’Arrigo, T.; De Majo, M. Contrast-Enhanced Ultrasound in Cystic Endometrial Hyperplasia-Pyometra Complex in the Bitch: A Preliminary Study. Animals 2020, 10, 1368. [Google Scholar] [CrossRef] [PubMed]
- Woźna-Wysocka, M.; Rybska, M.; Błaszak, B.; Jaśkowski, B.M.; Kulus, M.; Jaśkowski, J.M. Morphological changes in bitches endometrium affected by cystic endometrial hyperplasia—Pyometra complex—The value of histopathological examination. BMC Vet. Res. 2021, 17, 174. [Google Scholar] [CrossRef] [PubMed]
- Fieni, F.; Topie, E.; Gogny, A. Medical treatment for pyometra in dogs. Reprod. Domest. Anim. 2014, 49 (Suppl. S2), 28–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hart, B.L.; Hart, L.A.; Thigpen, A.P.; Willits, N.H. Long-term health effects of neutering dogs: Comparison of Labrador Retrievers with Golden Retrievers. PLoS ONE 2014, 9, e102241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, A.N. The role of neutering in cancer development. Vet. Clin. N. Am. Small Anim. Pr. 2014, 44, 965–975. [Google Scholar] [CrossRef]
- Root Kustritz, M.V. Effects of surgical sterilization on canine and feline health and on society. Reprod. Domest. Anim. 2012, 47 (Suppl. S4), 214–222. [Google Scholar] [CrossRef]
- Rota, A.; Corrò, M.; Patuzzi, I.; Milani, C.; Masia, S.; Mastrorilli, E.; Petrin, S.; Longo, A.; Del Carro, A.; Losasso, C. Effect of sterilization on the canine vaginal microbiota: A pilot study. BMC Vet. Res. 2020, 16, 455. [Google Scholar] [CrossRef] [PubMed]
- Lund, E.M.; Armstrong, P.J.; Kirk, C.A.; Klausner, J.S. Prevalence and risk factors for obesity in adult dogs from private US veterinary practices. J. Appl. Res. Vet. Med. 2006, 4, 177–186. [Google Scholar]
- Ko, H.Y.; Kim, J.; Geum, M.; Kim, H.J. Cystatin C and Neutrophil Gelatinase-Associated Lipocalin as Early Biomarkers for Chronic Kidney Disease in Dogs. Top. Companion Anim. Med. 2021, 45, 100580. [Google Scholar] [CrossRef] [PubMed]
- Heilmann, R.M.; Steiner, J.M. Clinical utility of currently available biomarkers in inflammatory enteropathies of dogs. J. Vet. Intern. Med. 2018, 32, 1495–1508. [Google Scholar] [CrossRef] [PubMed]
- Kaszak, I.; Ruszczak, A.; Kanafa, S.; Kacprzak, K.; Król, M.; Jurka, P. Current biomarkers of canine mammary tumors. Acta Vet. Scand. 2018, 60, 66. [Google Scholar] [CrossRef] [Green Version]
- Tabrez, S.; Shait Mohammed, M.R.; Jabir, N.R.; Khan, M.I. Identification of novel cardiovascular disease associated metabolites using untargeted metabolomics. Biol. Chem. 2021, 402, 749–757. [Google Scholar] [CrossRef]
- Bauermeister, A.; Mannochio-Russo, H.; Costa-Lotufo, L.V.; Jarmusch, A.K.; Dorrestein, P.C. Mass spectrometry-based metabolomics in microbiome investigations. Nat. Rev. Microbiol. 2022, 20, 143–160. [Google Scholar] [CrossRef]
- Cui, L.; Lu, H.; Lee, Y.H. Challenges and emergent solutions for LC-MS/MS based untargeted metabolomics in diseases. Mass Spectrom. Rev. 2018, 37, 772–792. [Google Scholar] [CrossRef]
- Johnson, C.H.; Ivanisevic, J.; Siuzdak, G. Metabolomics: Beyond biomarkers and towards mechanisms. Nat. Rev. Mol. Cell Biol. 2016, 17, 451–459. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Ma, D.; Yang, Y.S.; Yang, F.; Ding, J.H.; Gong, Y.; Jiang, L.; Ge, L.P.; Wu, S.Y.; Yu, Q.; et al. Comprehensive metabolomics expands precision medicine for triple-negative breast cancer. Cell Res. 2022, 32, 477–490. [Google Scholar] [CrossRef]
- Chen, Z.Z.; Gerszten, R.E. Metabolomics and Proteomics in Type 2 Diabetes. Circ. Res. 2020, 126, 1613–1627. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Liu, Z.; Gong, L. Biomarker identification and pathway analysis of rheumatoid arthritis based on metabolomics in combination with ingenuity pathway analysis. Proteomics 2021, 21, e2100037. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Sun, H.; Yan, G.; Wang, P.; Wang, X. Metabolomics for Biomarker Discovery: Moving to the Clinic. Biomed. Res. Int. 2015, 2015, 354671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortiz, C.N.; Torres-Reverón, A.; Appleyard, C.B. Metabolomics in endometriosis: Challenges and perspectives for future studies. Reprod. Fertil. 2021, 2, R35–R50. [Google Scholar] [CrossRef]
- Kuleš, J.; Horvatić, A.; Guillemin, N.; Ferreira, R.F.; Mischke, R.; Mrljak, V.; Chadwick, C.C.; Eckersall, P.D. The plasma proteome and the acute phase protein response in canine pyometra. J. Proteom. 2020, 223, 103817. [Google Scholar] [CrossRef]
- Hagman, R. Pyometra in Small Animals 2.0. Vet. Clin. N. Am. Small Anim Pr. 2022, 52, 631–657. [Google Scholar] [CrossRef]
- Franco-Martínez, L.; Horvatić, A.; Gelemanović, A.; Samardžija, M.; Mrljak, V.; Contreras-Aguilar, M.D.; Martínez-Subiela, S.; Dąbrowski, R.; Tvarijonaviciute, A. Changes in the Salivary Proteome Associated with Canine Pyometra. Front. Vet. Sci. 2020, 7, 277. [Google Scholar] [CrossRef]
- Haas, M.; Kaup, F.J.; Neumann, S. Canine pyometra: A model for the analysis of serum CXCL8 in inflammation. J. Vet. Med. Sci. 2016, 78, 375–381. [Google Scholar] [CrossRef] [Green Version]
- Hillström, A.; Hagman, R.; Tvedten, H.; Kjelgaard-Hansen, M. Validation of a commercially available automated canine-specific immunoturbidimetric method for measuring canine C-reactive protein. Vet. Clin. Pathol. 2014, 43, 235–243. [Google Scholar] [CrossRef]
- Troisi, J.; Sarno, L.; Landolfi, A.; Scala, G.; Martinelli, P.; Venturella, R.; Di Cello, A.; Zullo, F.; Guida, M. Metabolomic Signature of Endometrial Cancer. J. Proteome Res. 2018, 17, 804–812. [Google Scholar] [CrossRef]
- Andörfer, L.; Holtfreter, B.; Weiss, S.; Matthes, R.; Pitchika, V.; Schmidt, C.O.; Samietz, S.; Kastenmüller, G.; Nauck, M.; Völker, U.; et al. Salivary metabolites associated with a 5-year tooth loss identified in a population-based setting. BMC Med. 2021, 19, 161. [Google Scholar] [CrossRef]
- Jia, W.; Fan, Z.; Shi, Q.; Zhang, R.; Wang, X.; Shi, L. LC-MS-based metabolomics reveals metabolite dynamic changes during irradiation of goat meat. Food Res. Int. 2021, 150, 110721. [Google Scholar] [CrossRef] [PubMed]
- Wirthgen, E.; Hoeflich, A.; Rebl, A.; Günther, J. Kynurenic Acid: The Janus-Faced Role of an Immunomodulatory Tryptophan Metabolite and Its Link to Pathological Conditions. Front. Immunol. 2017, 8, 1957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fatokun, A.A.; Hunt, N.H.; Ball, H.J. Indoleamine 2,3-dioxygenase 2 (IDO2) and the kynurenine pathway: Characteristics and potential roles in health and disease. Amino Acids 2013, 45, 1319–1329. [Google Scholar] [CrossRef] [PubMed]
- Plazas, E.; Avila, M.M.; Muñoz, D.R.; Cuca, S.L. Natural isoquinoline alkaloids: Pharmacological features and multi-target potential for complex diseases. Pharm. Res. 2022, 177, 106126. [Google Scholar] [CrossRef] [PubMed]
- Rushworth, G.F.; Megson, I.L. Existing and potential therapeutic uses for N-acetylcysteine: The need for conversion to intracellular glutathione for antioxidant benefits. Pharmacol. Ther. 2014, 141, 150–159. [Google Scholar] [CrossRef]
- Cox, J.E. Progestagens in bitches: A review. J. Small Anim. Pr. 1970, 11, 759–778. [Google Scholar] [CrossRef]
- Liu, Y.; Yasawong, M.; Yu, B. Metabolic engineering of Escherichia coli for biosynthesis of β-nicotinamide mononucleotide from nicotinamide. Microb. Biotechnol. 2021, 14, 2581–2591. [Google Scholar] [CrossRef]
- Want, E.J.; Masson, P.; Michopoulos, F.; Wilson, I.D.; Theodoridis, G.; Plumb, R.S.; Shockcor, J.; Loftus, N.; Holmes, E.; Nicholson, J.K. Global metabolic profiling of animal and human tissues via UPLC-MS. Nat. Protoc. 2013, 8, 17–32. [Google Scholar] [CrossRef]
- Wen, B.; Mei, Z.; Zeng, C.; Liu, S. metaX: A flexible and comprehensive software for processing metabolomics data. BMC Bioinform. 2017, 18, 183. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Dogs with Pyometra | Healthy Dogs |
|---|---|---|
| Mean age (years) ± SD | 9.33 ± 3.39 | 8.47 ± 2.67 |
| Number of breeds | ||
| Teddy dog | 5 | 7 |
| Poodle | 2 | 1 |
| Border Collie | 1 | 2 |
| Pomeranian | 1 | 3 |
| Other | 6 | 2 |
| Number of somatotypes | ||
| Large | 2 | 0 |
| Medium | 2 | 3 |
| Small | 11 | 12 |
| Number of lineages | ||
| Mix breed | 0 | 0 |
| Pure breed | 15 | 15 |
| Items | Dogs with Pyometra | Healthy Dogs | Reference | Unit | p-Value | ||||
|---|---|---|---|---|---|---|---|---|---|
| Min | Max | Mean | Min | Max | Mean | ||||
| Blood routine | |||||||||
| White blood cell (WBC) | 7.13 | 48.23 | 19.25 ± 12.70 | 6.92 | 13.96 | 9.61 ± 2.00 | 6.00–17.00 | 109/L | 0.007 |
| Neutrophils (Neu) | 4.79 | 44.64 | 16.33 ± 11.73 | 4.53 | 9.11 | 6.46 ± 1.56 | 3.62–12.30 | 109/L | 0.003 |
| Blood biochemical examination | |||||||||
| Blood urea nitrogen (BUN) | 3.43 | 15.90 | 9.67 ± 4.02 | 2.98 | 6.94 | 5.22 ± 1.11 | 2.5–9.6 | mmol/L | 0.0002 |
| Globulins (GLO) | 29.80 | 67.20 | 52.05 ± 11.86 | 25.2 | 40.8 | 33.87 ± 4.43 | 23–52 | g/L | 5.97 × 10−6 |
| C-reactive protein | |||||||||
| c-CRP | 11.00 | 47.10 | 22.20 ± 11.22 | 1.1 | 9.9 | 4.33 ± 2.34 | 16–218 | mg/L | 3.48 × 10−6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, H.-H.; Du, C.-T.; Zhang, Y.-Z.; Yu, C.; Huang, R.-L.; Tang, X.-Y.; Xie, G.-H. Identification of Canine Pyometra-Associated Metabolites Using Untargeted Metabolomics. Int. J. Mol. Sci. 2022, 23, 14161. https://doi.org/10.3390/ijms232214161
Zheng H-H, Du C-T, Zhang Y-Z, Yu C, Huang R-L, Tang X-Y, Xie G-H. Identification of Canine Pyometra-Associated Metabolites Using Untargeted Metabolomics. International Journal of Molecular Sciences. 2022; 23(22):14161. https://doi.org/10.3390/ijms232214161
Chicago/Turabian StyleZheng, Hui-Hua, Chong-Tao Du, Yu-Zhu Zhang, Chao Yu, Rong-Lei Huang, Xin-Yue Tang, and Guang-Hong Xie. 2022. "Identification of Canine Pyometra-Associated Metabolites Using Untargeted Metabolomics" International Journal of Molecular Sciences 23, no. 22: 14161. https://doi.org/10.3390/ijms232214161
APA StyleZheng, H. -H., Du, C. -T., Zhang, Y. -Z., Yu, C., Huang, R. -L., Tang, X. -Y., & Xie, G. -H. (2022). Identification of Canine Pyometra-Associated Metabolites Using Untargeted Metabolomics. International Journal of Molecular Sciences, 23(22), 14161. https://doi.org/10.3390/ijms232214161