

Protective Effect of Resveratrol in an Experimental Model of Salicylate-Induced Tinnitus




Abstract
:1. Introduction
2. Results
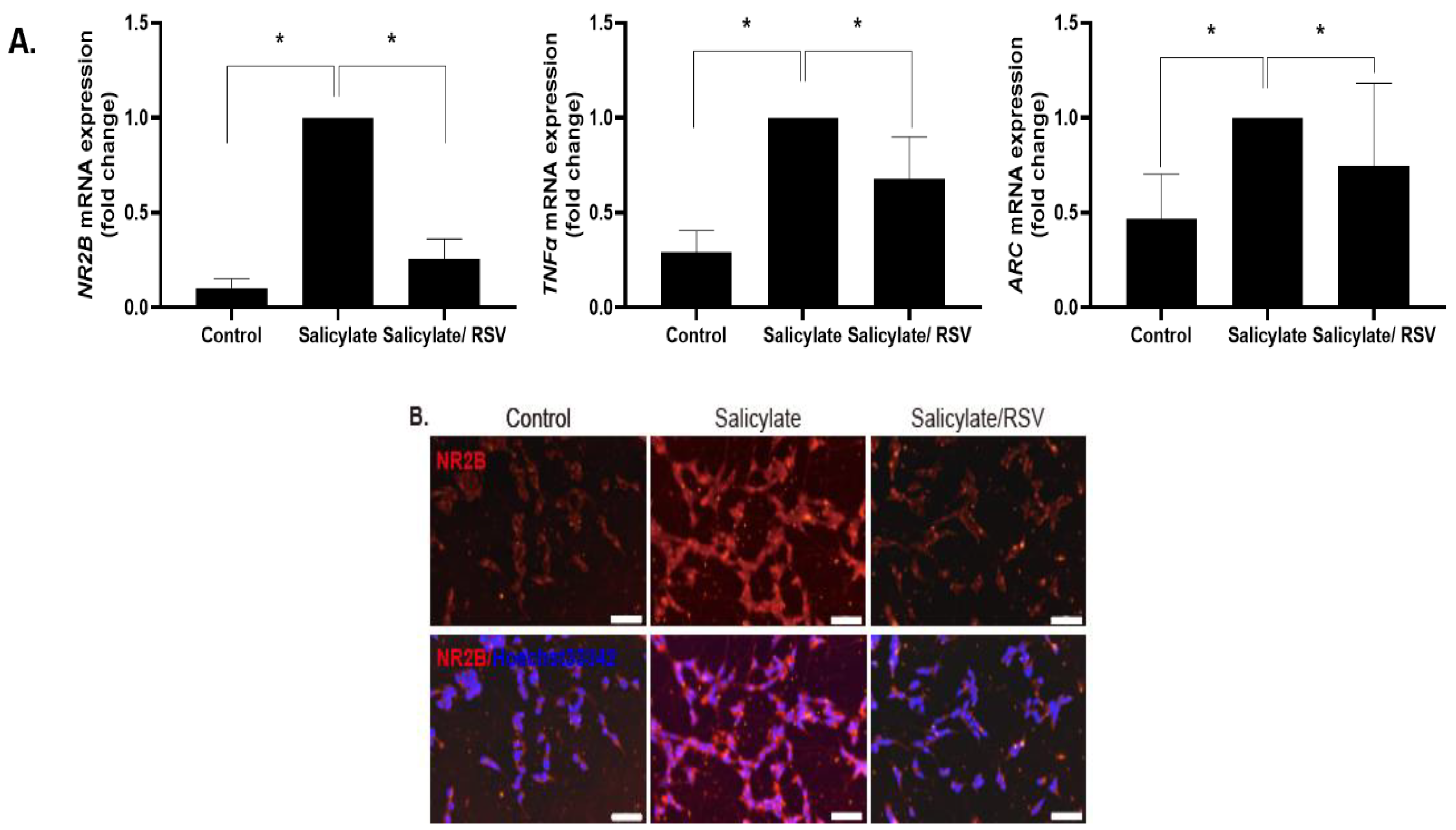
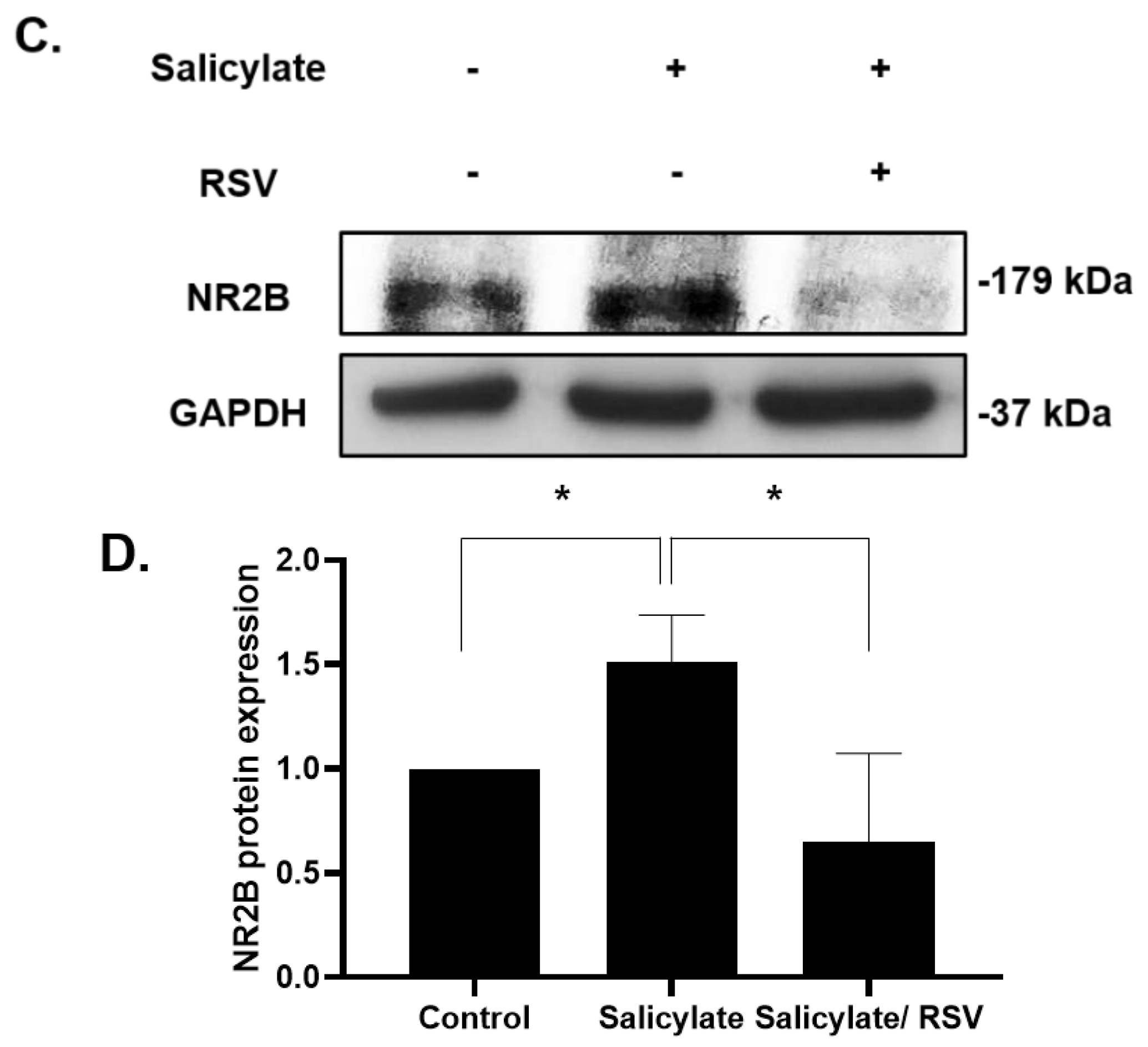
2.1. RSV Induces a Decrease in the NR2B Expression in Neuronal Cells
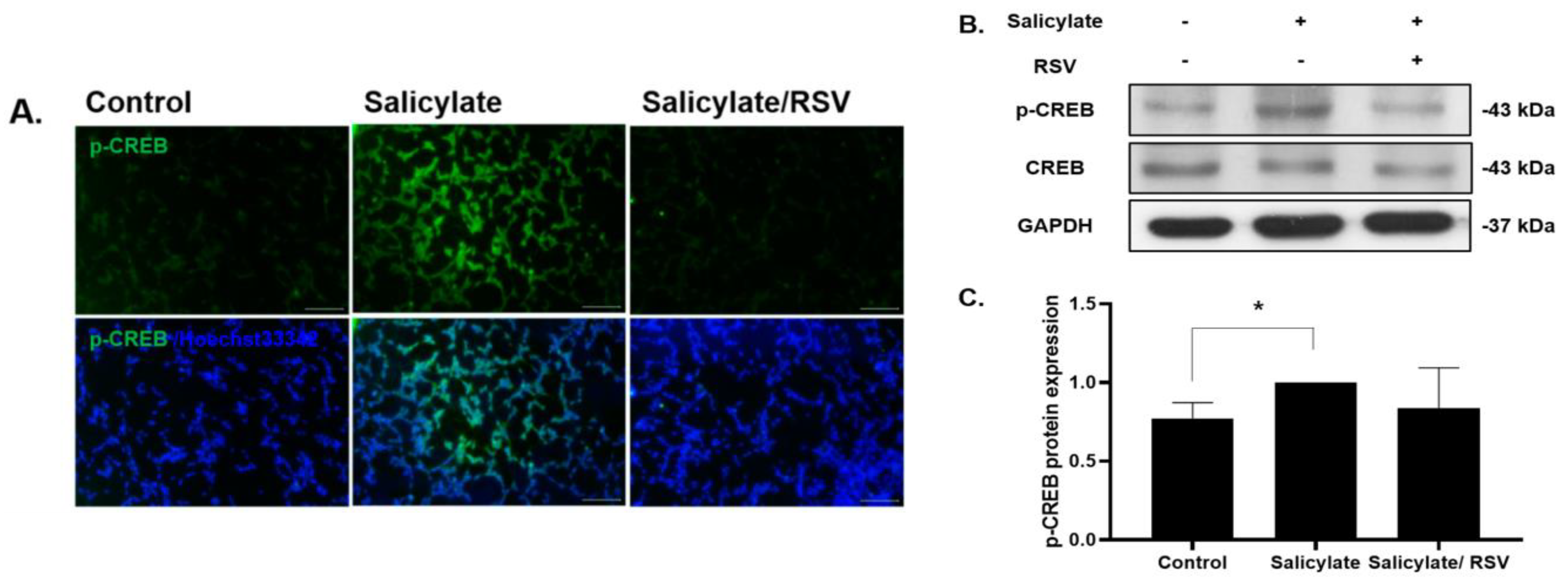
2.2. RSV Pretreatment Decreases Phosphorylated CREB in Salicylate-Treated Neuronal Cells
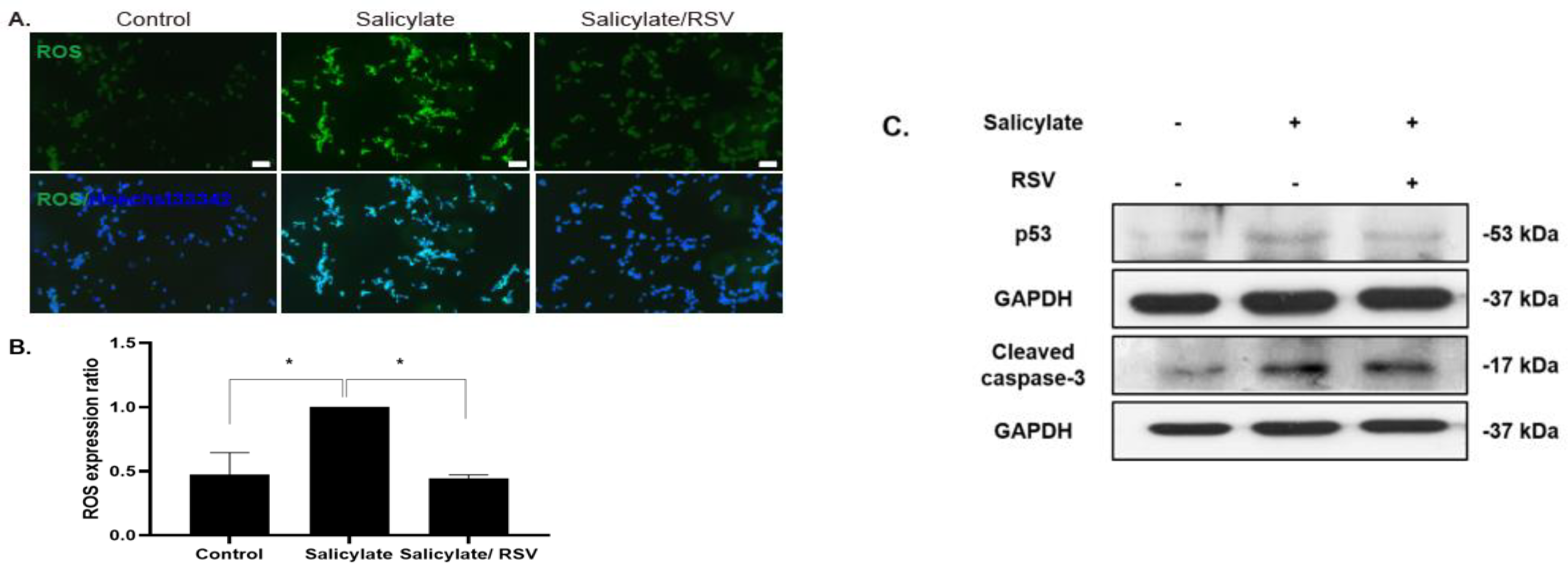
2.3. RSV Inhibits a ROS Increase in Salicylate-Treated Neuronal Cells
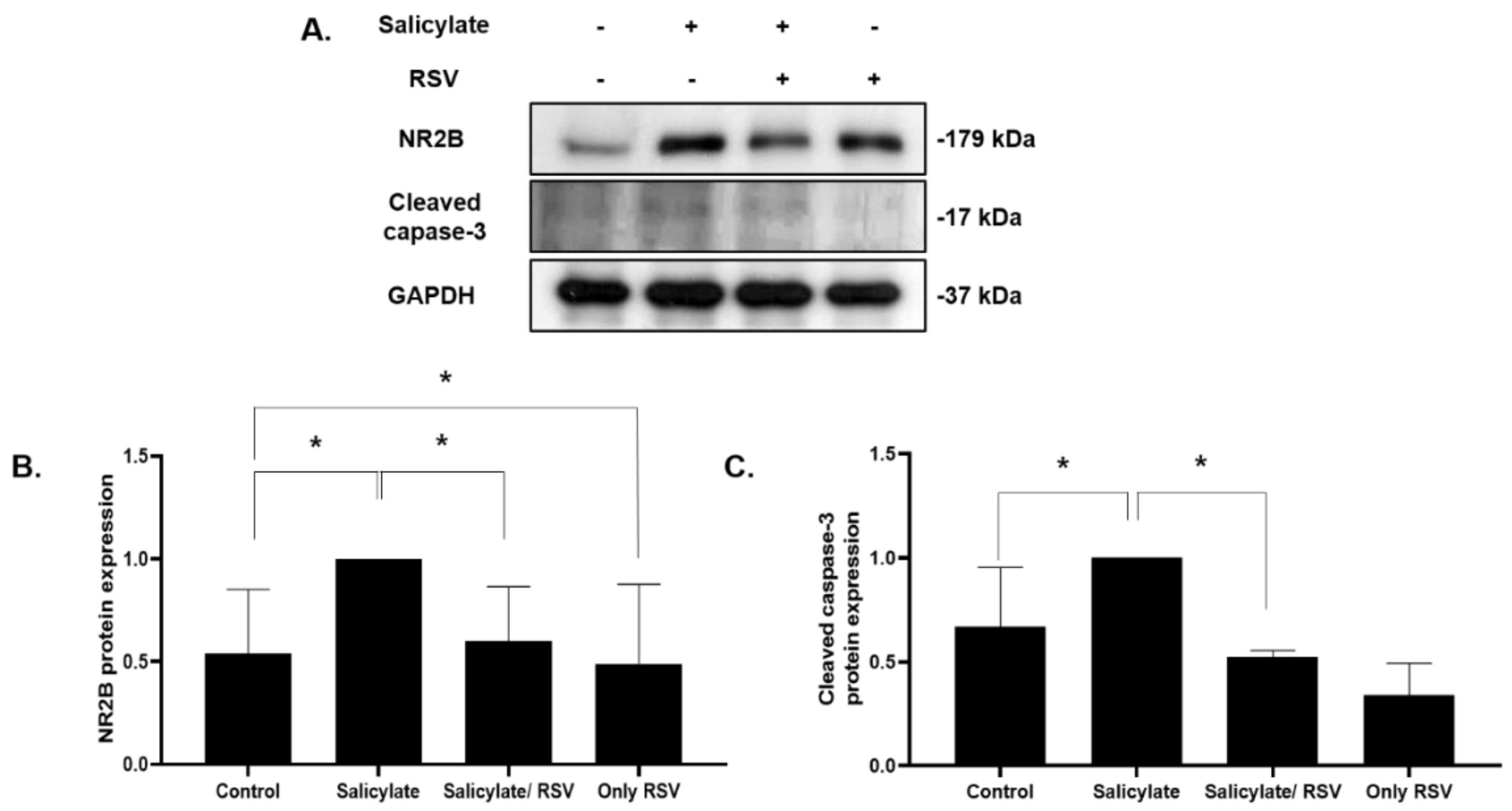
2.4. Effect of RSV on Salicylate-Treated Rat Cortical Neurons
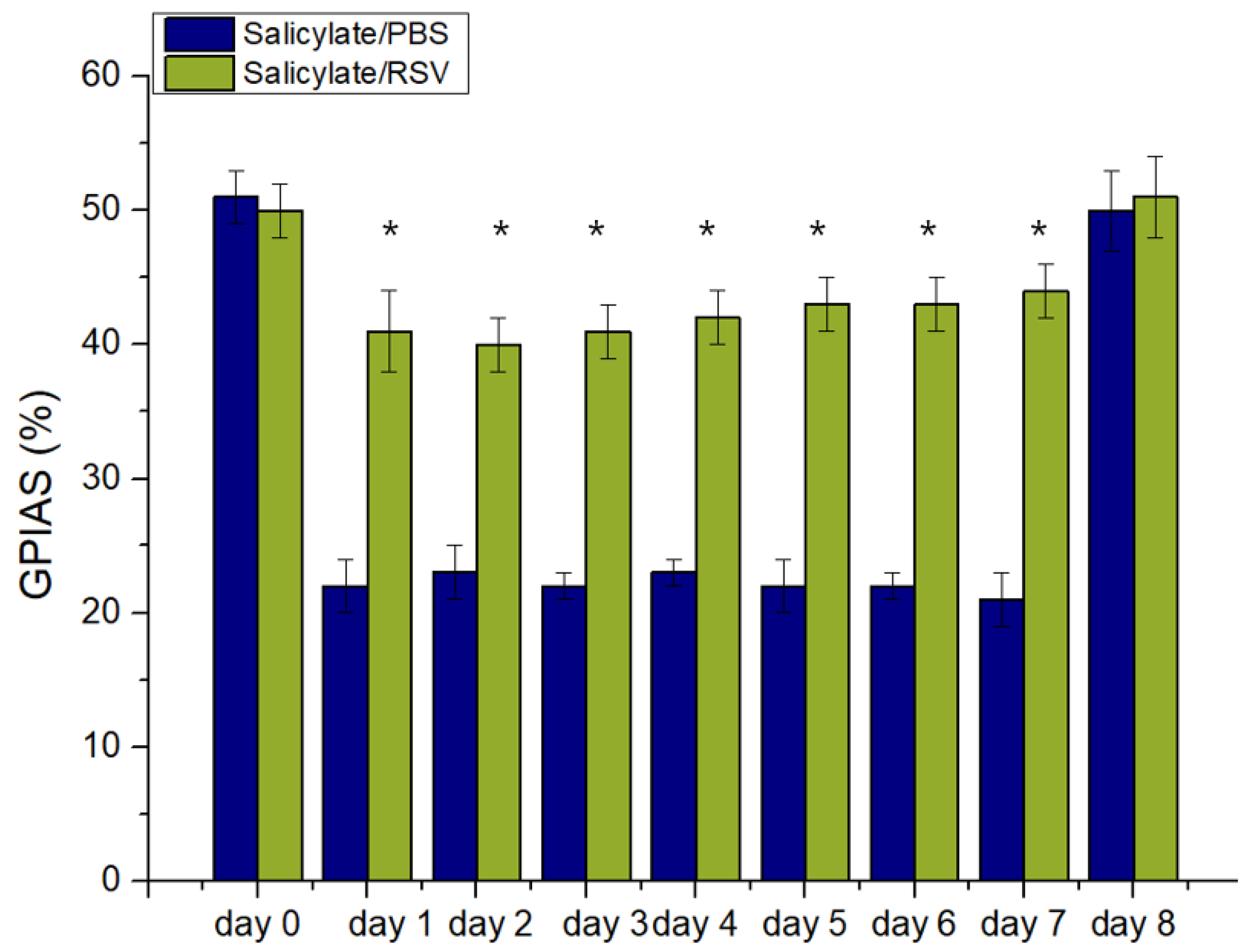
2.5. RSV Attenuates a Salicylate-Induced GPIAS Decrease in Rats
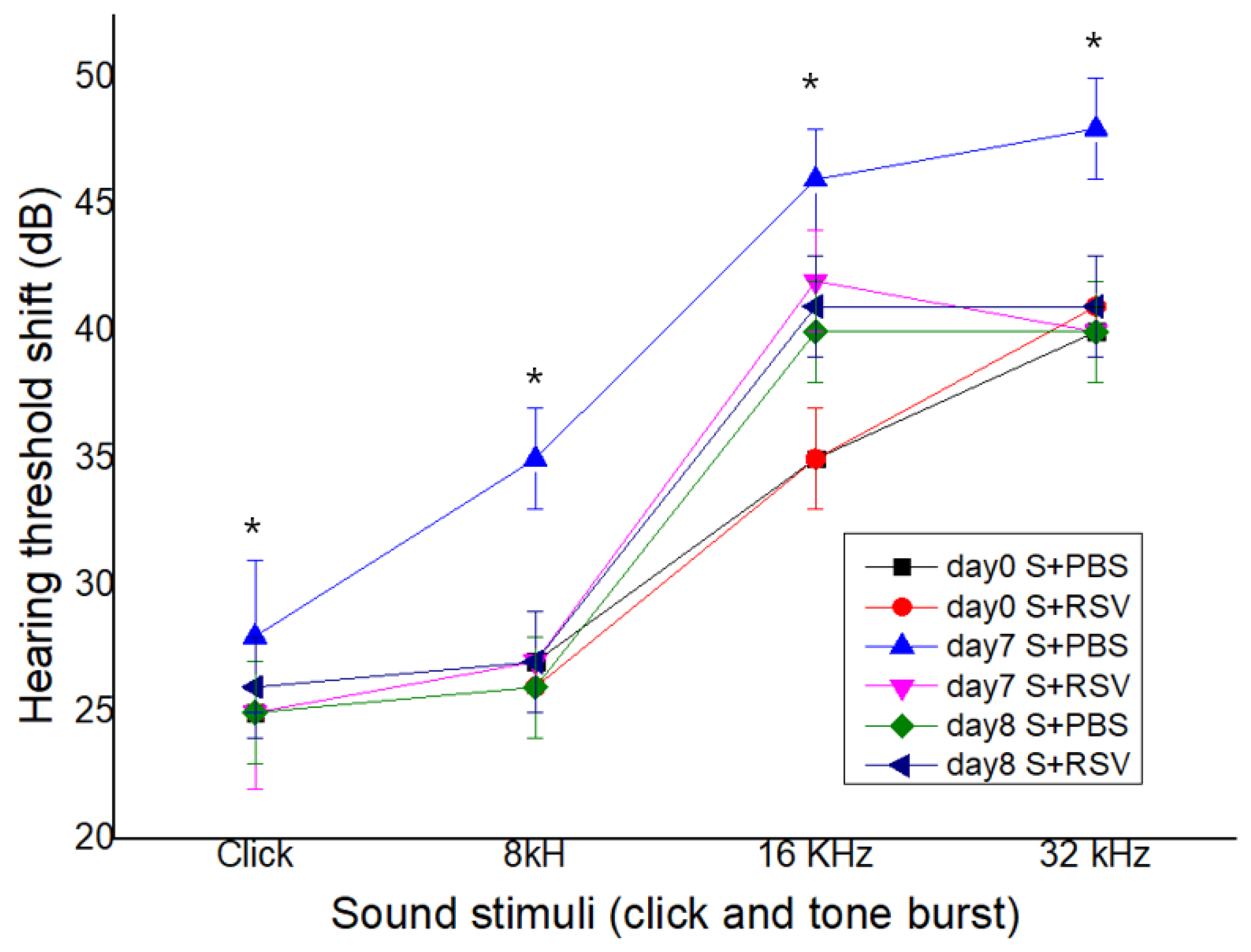
2.6. RSV Attenuates a Salicylate-Induced Elevation of the ABR Threshold in Rats
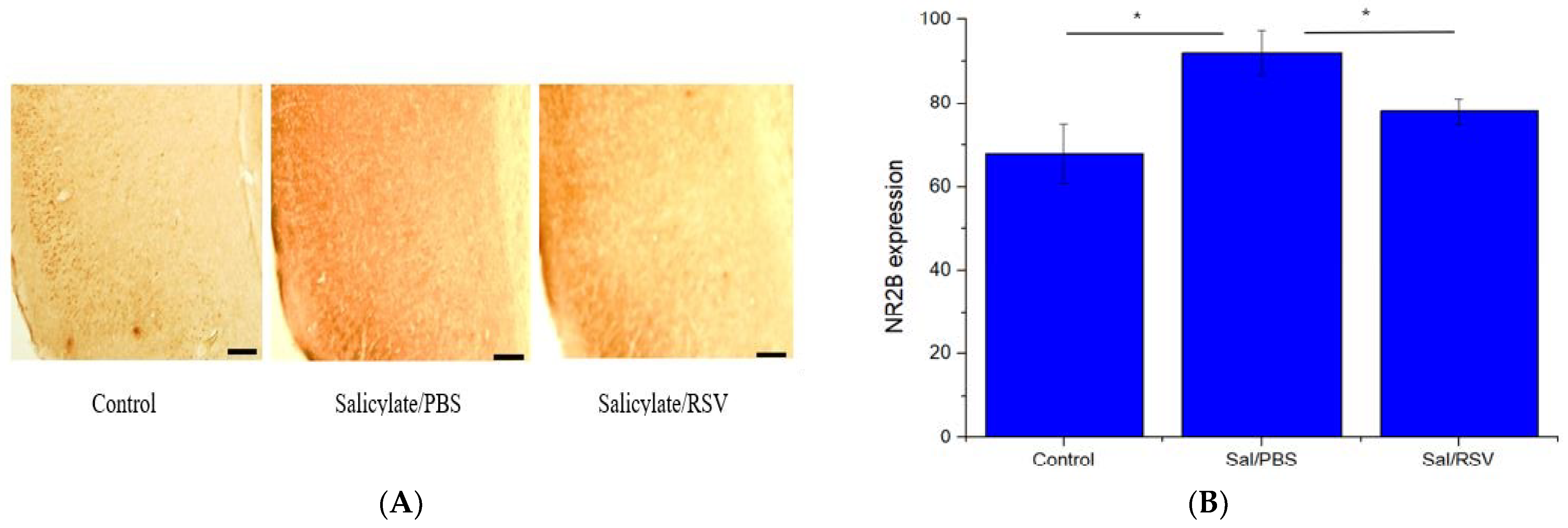
2.7. Spatial Expression of NR2B in the Auditory Cortex after Salicylate and RSV Treatments
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Treatment
4.2. Quantitative Polymerase Chain Reaction (qPCR)
4.3. Immunocytochemistry Staining
4.4. Immunoblotting
4.5. ROS Assay
4.6. In Vivo Experiments
4.6.1. Animals
4.6.2. GPIAS Measurement
4.6.3. ABR Measurement
4.6.4. Preparation of Free-Floating Sections
4.6.5. Immunohistochemistry
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tang, D.; Li, H.; Chen, L. Advances in Understanding, Diagnosis, and Treatment of Tinnitus. Adv. Exp. Med. Biol. 2019, 1130, 109–128. [Google Scholar] [CrossRef] [PubMed]
- Yi, B.; Hu, S.; Zuo, C.; Jiao, F.; Lv, J.; Chen, D.; Ma, Y.; Chen, J.; Mei, L.; Wang, X.; et al. Effects of long-term salicylate administration on synaptic ultrastructure and metabolic activity in the rat CNS. Sci. Rep. 2016, 6, 24428. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, J.T.; Schafer, S.T.; Gage, F.H. Adult Neurogenesis in the Hippocampus: From Stem Cells to Behavior. Cell 2016, 167, 897–914. [Google Scholar] [CrossRef] [PubMed]
- Alvan, G.; Berninger, E.; Gustafsson, L.L.; Karlsson, K.K.; Paintaud, G.; Wakelkamp, M. Concentration-Response Relationship of Hearing Impairment Caused by Quinine and Salicylate: Pharmacological Similarities but Different Molecular Mechanisms. Basic. Clin. Pharmacol. Toxicol. 2017, 120, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Palmer, B.F.; Clegg, D.J. Salicylate Toxicity. N. Engl. J. Med. 2020, 382, 2544–2555. [Google Scholar] [CrossRef]
- Wu, C.; Wu, X.; Yi, B.; Cui, M.; Wang, X.; Wang, Q.; Wu, H.; Huang, Z. Changes in GABA and glutamate receptors on auditory cortical excitatory neurons in a rat model of salicylate-induced tinnitus. Am. J. Transl. Res. 2018, 10, 3941–3955. [Google Scholar]
- Bing, D.; Lee, S.C.; Campanelli, D.; Xiong, H.; Matsumoto, M.; Panford-Walsh, R.; Wolpert, S.; Praetorius, M.; Zimmermann, U.; Chu, H.; et al. Cochlear NMDA receptors as a therapeutic target of noise-induced tinnitus. Cell Physiol. Biochem. 2015, 35, 1905–1923. [Google Scholar] [CrossRef]
- Noreña, A.J.; Farley, B.J. Tinnitus-related neural activity: Theories of generation, propagation, and centralization. Hear. Res. 2013, 295, 161–171. [Google Scholar] [CrossRef]
- Cui, W.; Wang, H.; Cheng, Y.; Ma, X.; Lei, Y.; Ruan, X.; Shi, L.; Lv, M. Long-term treatment with salicylate enables NMDA receptors and impairs AMPA receptors in C57BL/6J mice inner hair cell ribbon synapse. Mol. Med. Rep. 2019, 19, 51–58. [Google Scholar] [CrossRef]
- Hong, J.; Chen, Y.; Zhang, Y.; Li, J.; Ren, L.; Yang, L.; Shi, L.; Li, A.; Zhang, T.; Li, H.; et al. N-Methyl-D-Aspartate Receptors Involvement in the Gentamicin-Induced Hearing Loss and Pathological Changes of Ribbon Synapse in the Mouse Cochlear Inner Hair Cells. Neural. Plast. 2018, 2018, 3989201. [Google Scholar] [CrossRef]
- Segal, J.A.; Harris, B.D.; Kustova, Y.; Basile, A.; Skolnick, P. Aminoglycoside neurotoxicity involves NMDA receptor activation. Brain Res. 1999, 815, 270–277. [Google Scholar] [CrossRef]
- Xia, N.; Daiber, A.; Förstermann, U.; Li, H. Antioxidant effects of resveratrol in the cardiovascular system. Br. J. Pharmacol. 2017, 174, 1633–1646. [Google Scholar] [CrossRef]
- Karthick, C.; Periyasamy, S.; Jayachandran, K.S.; Anusuyadevi, M. Intrahippocampal Administration of Ibotenic Acid Induced Cholinergic Dysfunction via NR2A/NR2B Expression: Implications of Resveratrol against Alzheimer Disease Pathophysiology. Front. Mol. Neurosci. 2016, 9, 28. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Pang, Y.; Wei, W.; Shao, A.; Deng, C.; Li, X.; Chang, H.; Hu, P.; Liu, X.; Zhang, X. Resveratrol protects retinal ganglion cell axons through regulation of the SIRT1-JNK pathway. Exp. Eye Res. 2020, 200, 108249. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Kim, H.S.; An, Y.S.; Chang, J.; Choi, J.; Im, G.J. Protective effect of resveratrol against cisplatin-induced ototoxicity in HEI-OC1 auditory cells. Int. J. Pediatr. Otorhinolaryngol. 2015, 79, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Jastreboff, P.J.; Brennan, J.F.; Coleman, J.K.; Sasaki, C.T. Phantom auditory sensation in rats: An animal model for tinnitus. Behav. Neurosci. 1988, 102, 811–822. [Google Scholar] [CrossRef] [PubMed]
- Qin, D.; Liu, P.; Chen, H.; Huang, X.; Ye, W.; Lin, X.; Wei, F.; Su, J. Salicylate-Induced Ototoxicity of Spiral Ganglion Neurons: Ca2+/CaMKII-Mediated Interaction Between NMDA Receptor and GABA(A) Receptor. Neurotox. Res. 2019, 35, 838–847. [Google Scholar] [CrossRef]
- Sheppard, A.; Hayes, S.H.; Chen, G.D.; Ralli, M.; Salvi, R. Review of salicylate-induced hearing loss, neurotoxicity, tinnitus and neuropathophysiology. Acta Otorhinolaryngol. Ital. 2014, 34, 79–93. [Google Scholar]
- Gupta, K.; Hardingham, G.E.; Chandran, S. NMDA receptor-dependent glutamate excitotoxicity in human embryonic stem cell-derived neurons. Neurosci. Lett. 2013, 543, 95–100. [Google Scholar] [CrossRef]
- Nunes, S.; Danesi, F.; Del Rio, D.; Silva, P. Resveratrol and inflammatory bowel disease: The evidence so far. Nutr. Res. Rev. 2018, 31, 85–97. [Google Scholar] [CrossRef]
- Shaito, A.; Posadino, A.M.; Younes, N.; Hasan, H.; Halabi, S.; Alhababi, D.; Al-Mohannadi, A.; Abdel-Rahman, W.M.; Eid, A.H.; Nasrallah, G.K.; et al. Potential Adverse Effects of Resveratrol: A Literature Review. Int. J. Mol. Sci. 2020, 21, 2084. [Google Scholar] [CrossRef] [PubMed]
- Corpas, R.; Griñán-Ferré, C.; Palomera-Ávalos, V.; Porquet, D.; García de Frutos, P.; Franciscato Cozzolino, S.M.; Rodríguez-Farré, E.; Pallàs, M.; Sanfeliu, C.; Cardoso, B.R. Melatonin induces mechanisms of brain resilience against neurodegeneration. J. Pineal. Res. 2018, 65, e12515. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Wang, S.J. Inhibitory effect of glutamate release from rat cerebrocortical nerve terminals by resveratrol. Neurochem. Int. 2009, 54, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Scasso, F.; Sprio, A.E.; Canobbio, L.; Scanarotti, C.; Manini, G.; Berta, G.N.; Bassi, A.M. Dietary supplementation of coenzyme Q10 plus multivitamins to hamper the ROS mediated cisplatin ototoxicity in humans: A pilot study. Heliyon 2017, 3, e00251. [Google Scholar] [CrossRef]
- Tabuchi, K.; Nishimura, B.; Nakamagoe, M.; Hayashi, K.; Nakayama, M.; Hara, A. Ototoxicity: Mechanisms of cochlear impairment and its prevention. Curr. Med. Chem. 2011, 18, 4866–4871. [Google Scholar] [CrossRef]
- Deng, L.; Ding, D.; Su, J.; Manohar, S.; Salvi, R. Salicylate selectively kills cochlear spiral ganglion neurons by paradoxically up-regulating superoxide. Neurotox. Res. 2013, 24, 307–319. [Google Scholar] [CrossRef]
- Pardo, L.; Valor, L.M.; Eraso-Pichot, A.; Barco, A.; Golbano, A.; Hardingham, G.E.; Masgrau, R.; Galea, E. CREB Regulates Distinct Adaptive Transcriptional Programs in Astrocytes and Neurons. Sci. Rep. 2017, 7, 6390. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, B.; Wang, X.; Shang, X. Up-regulation of Ca(2+)/CaMKII/CREB signaling in salicylate-induced tinnitus in rats. Mol. Cell Biochem. 2018, 448, 71–76. [Google Scholar] [CrossRef]
- Yi, B.; Wu, C.; Shi, R.; Han, K.; Sheng, H.; Li, B.; Mei, L.; Wang, X.; Huang, Z.; Wu, H. Long-term Administration of Salicylate-induced Changes in BDNF Expression and CREB Phosphorylation in the Auditory Cortex of Rats. Otol. Neurotol. 2018, 39, e173–e180. [Google Scholar] [CrossRef]
- Son, Y.; Byun, S.J.; Pae, H.O. Involvement of heme oxygenase-1 expression in neuroprotection by piceatannol, a natural analog and a metabolite of resveratrol, against glutamate-mediated oxidative injury in HT22 neuronal cells. Amino Acids 2013, 45, 393–401. [Google Scholar] [CrossRef]
- Hosoda, R.; Hamada, H.; Uesugi, D.; Iwahara, N.; Nojima, I.; Horio, Y.; Kuno, A. Different Antioxidative and Antiapoptotic Effects of Piceatannol and Resveratrol. J. Pharmacol. Exp. Ther. 2021, 376, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Jang, C.H.; Lee, S.; Park, I.Y.; Song, A.; Moon, C.; Cho, G.W. Memantine Attenuates Salicylate-induced Tinnitus Possibly by Reducing NR2B Expression in Auditory Cortex of Rat. Exp. Neurobiol. 2019, 28, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Galazyuk, A.; Hébert, S. Gap-Prepulse Inhibition of the Acoustic Startle Reflex (GPIAS) for Tinnitus Assessment: Current Status and Future Directions. Front. Neurol. 2015, 6, 88. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.D.; Stolzberg, D.; Lobarinas, E.; Sun, W.; Ding, D.; Salvi, R. Salicylate-induced cochlear impairments, cortical hyperactivity and re-tuning, and tinnitus. Hear. Res. 2013, 295, 100–113. [Google Scholar] [CrossRef] [PubMed]
- Gong, N.; Zhang, M.; Zhang, X.B.; Chen, L.; Sun, G.C.; Xu, T.L. The aspirin metabolite salicylate enhances neuronal excitation in rat hippocampal CA1 area through reducing GABAergic inhibition. Neuropharmacology 2008, 54, 454–463. [Google Scholar] [CrossRef]
- Liu, J.; Li, X.; Wang, L.; Dong, Y.; Han, H.; Liu, G. Effects of salicylate on serotoninergic activities in rat inferior colliculus and auditory cortex. Hear. Res. 2003, 175, 45–53. [Google Scholar] [CrossRef]
- Kim, H.T.; Ohn, T.; Jeong, S.G.; Song, A.; Jang, C.H.; Cho, G.W. Oxidative stress-induced aberrant G9a activation disturbs RE-1-containing neuron-specific genes expression, leading to degeneration in human SH-SY5Y neuroblastoma cells. Korean J. Physiol. Pharmacol. 2021, 25, 51–58. [Google Scholar] [CrossRef]
- Ralli, M.; Troiani, D.; Podda, M.V.; Paciello, F.; Eramo, S.L.; de Corso, E.; Salvi, R.; Paludetti, G.; Fetoni, A.R. The effect of the NMDA channel blocker memantine on salicylate-induced tinnitus in rats. Acta Otorhinolaryngol. Ital. 2014, 34, 198–204. [Google Scholar]
- Lobarinas, E.; Hayes, S.H.; Allman, B.L. The gap-startle paradigm for tinnitus screening in animal models: Limitations and optimization. Hear. Res. 2013, 295, 150–160. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′→3′) | Reverse Primer (3′→5′) | Gene Accession |
|---|---|---|---|
| Human β-actin | ATCCGCAAAGACCTGTACGC | TCTTCATTGTGCTGGGTGCC | NM_001101 |
| Human NMDA (NR2B) | GGAGAGGTGGTCATGAAGAG | CATTGCTGCGTGACACCATG | NM_000834.4 |
| Human TNFα | GTTGTAGCAAACCCTCAAGCTG | CCAGCTGGTTATCTCTCAGCTC | NM_000594.3 |
| Human ARC | ACAACAGGTCTCAAGGTTCCC | AGCCGACTCCTCTCTGTAGC | NM_015193.4 |
| Rat GAPDH | CTGCCACTCAGAAGACTGTGG | TTCAGCTCTGGGATGACCTTG | NM_017008.4 |
| Rat NMDA (Nr2b) | GGAGATGGAAGAACTGGAAGCTC | GACACCTGCCATATTGTCGATG | NM_012574.1 |
| Rat TNFα | CCACCACGCTCTTCTGTCTAC | GATGATCTGAGTGTGAGGGTCTG | NM_012675.3 |
| Rat ARC | GTCTGCTGCATAGAAGGAACCAG | AGGGTGCCCACCACATACTGA | NM_019361.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, A.; Cho, G.-W.; Moon, C.; Park, I.; Jang, C.H. Protective Effect of Resveratrol in an Experimental Model of Salicylate-Induced Tinnitus. Int. J. Mol. Sci. 2022, 23, 14183. https://doi.org/10.3390/ijms232214183
Song A, Cho G-W, Moon C, Park I, Jang CH. Protective Effect of Resveratrol in an Experimental Model of Salicylate-Induced Tinnitus. International Journal of Molecular Sciences. 2022; 23(22):14183. https://doi.org/10.3390/ijms232214183
Chicago/Turabian StyleSong, Anji, Gwang-Won Cho, Changjong Moon, Ilyong Park, and Chul Ho Jang. 2022. "Protective Effect of Resveratrol in an Experimental Model of Salicylate-Induced Tinnitus" International Journal of Molecular Sciences 23, no. 22: 14183. https://doi.org/10.3390/ijms232214183
APA StyleSong, A., Cho, G. -W., Moon, C., Park, I., & Jang, C. H. (2022). Protective Effect of Resveratrol in an Experimental Model of Salicylate-Induced Tinnitus. International Journal of Molecular Sciences, 23(22), 14183. https://doi.org/10.3390/ijms232214183