
Structure, Function, and Applications of Soybean Calcium Transporters


Abstract
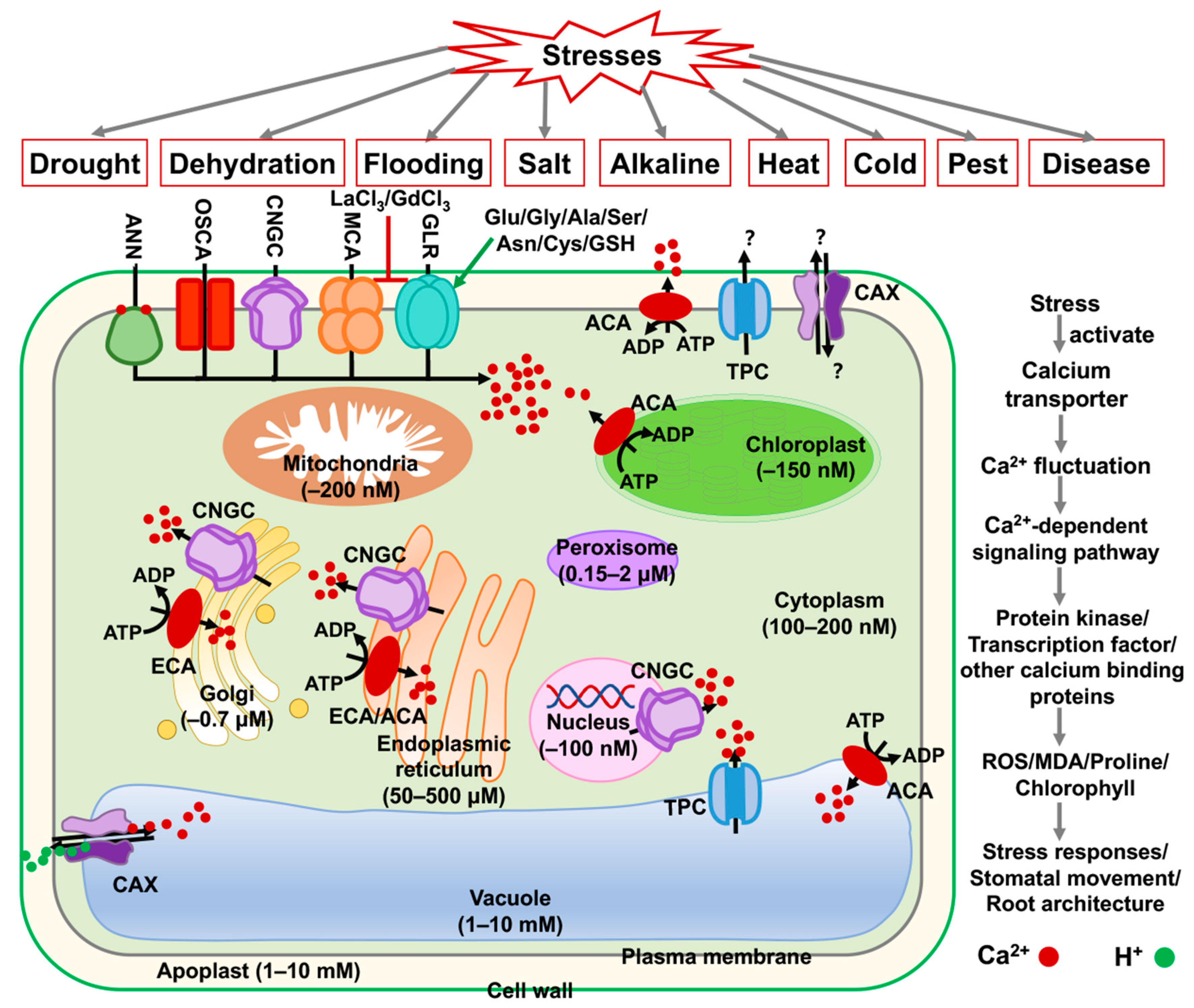
:1. Introduction
2. Calcium Efflux Transporters
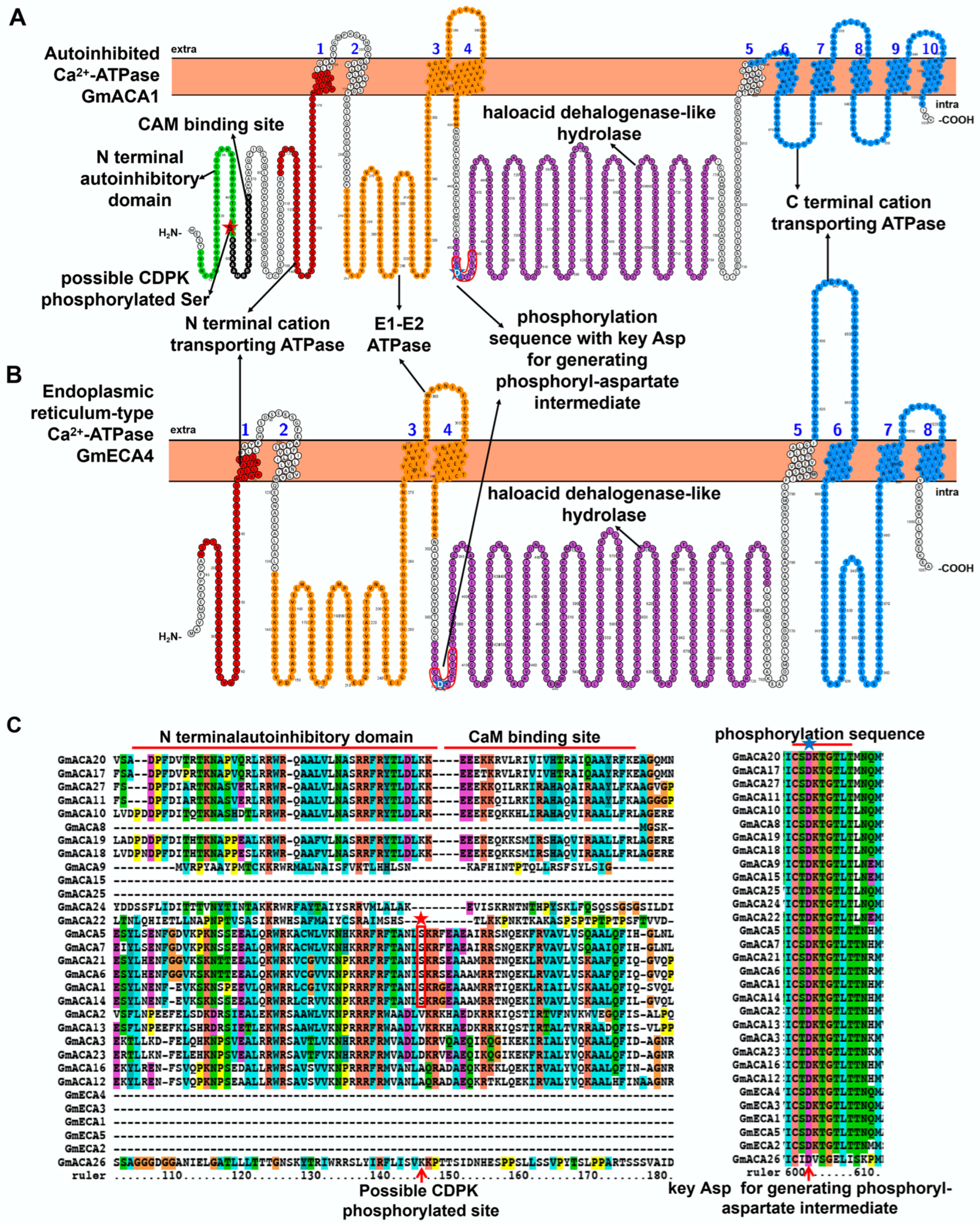
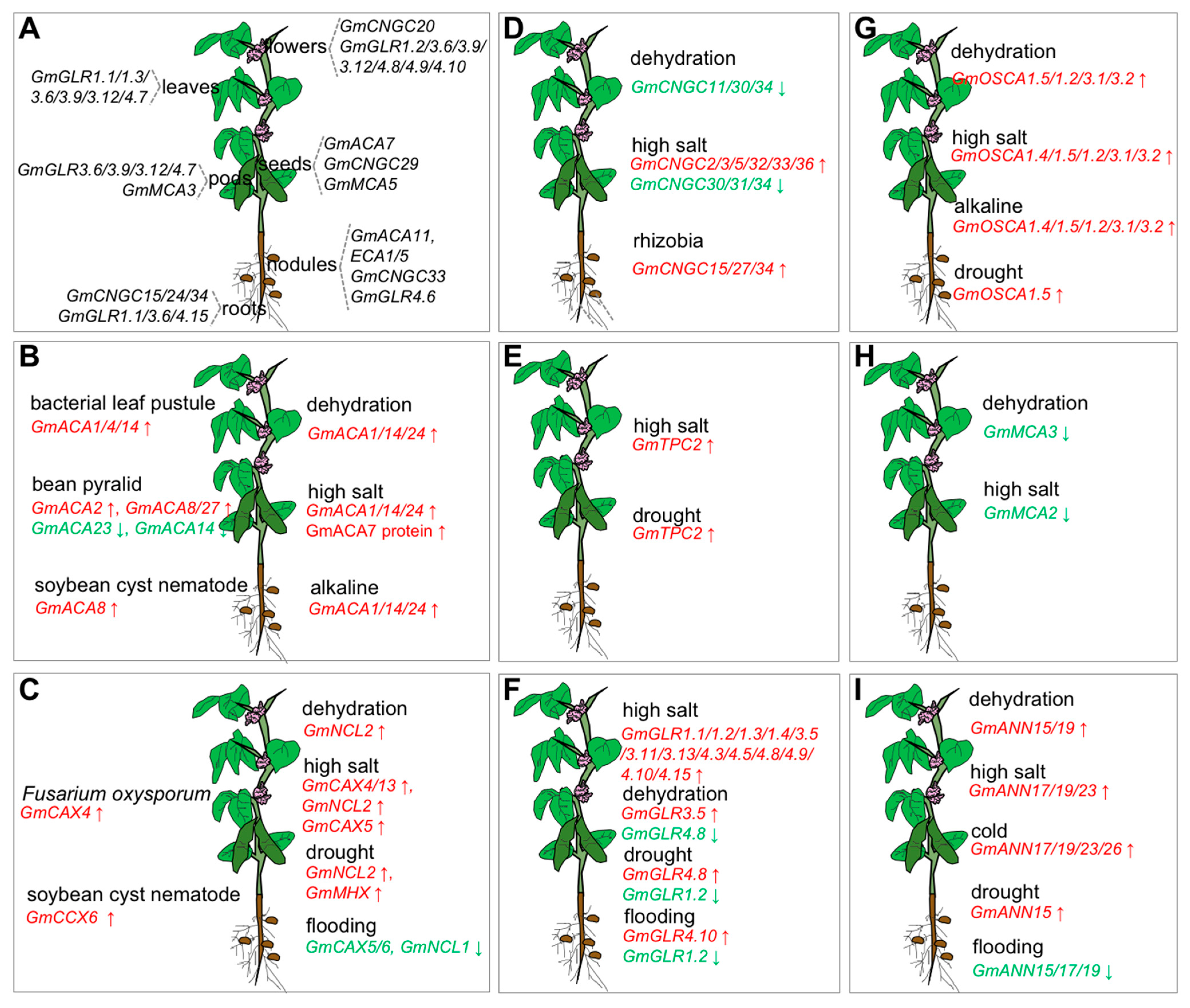
2.1. Characteristic Features and Roles of Ca2+-ATPase Involved in Soybean Stress Response
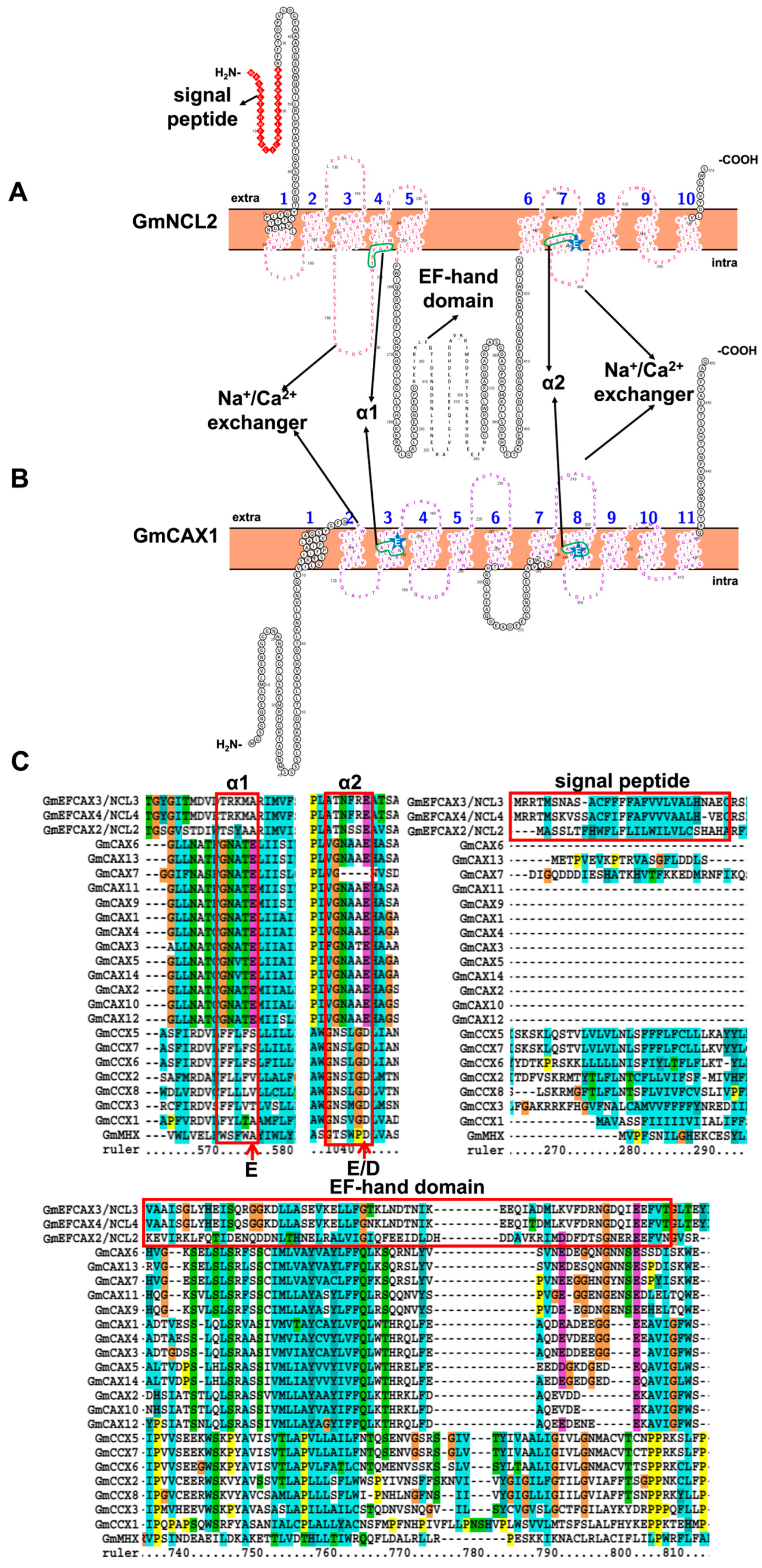
2.2. Characteristic Features and Roles of Ca2+/Cation Antiporter Involved in Soybean Stress Response
3. Calcium Influx Transporters
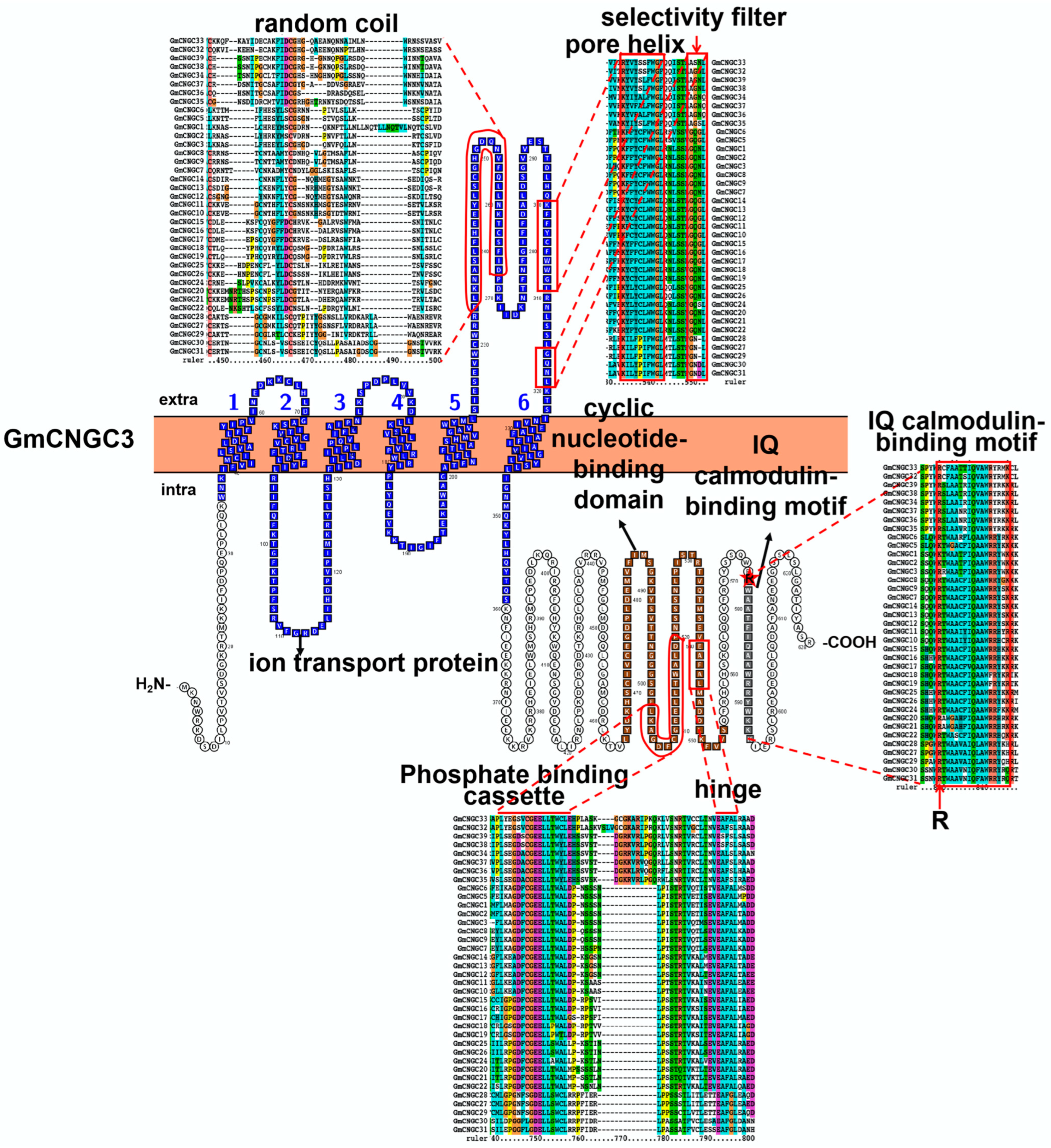
3.1. Characteristic Features and Roles of Cyclic Nucleotide-Gated Ion Channel (CNGC) Involved in Soybean Stress Response
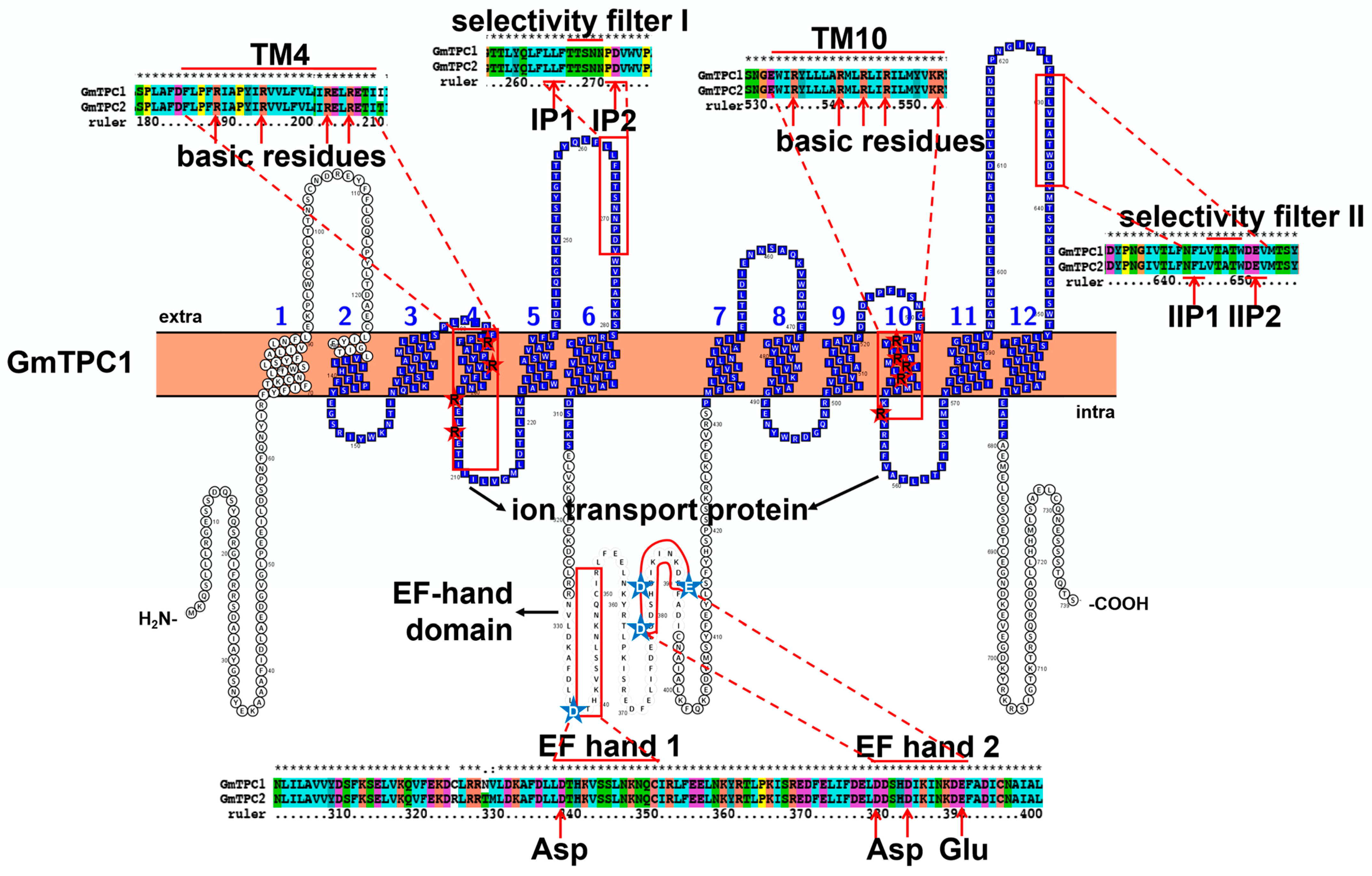
3.2. Characteristic Features and Roles of Two-Pore Cation (TPC) Channel Involved in Soybean Stress Response
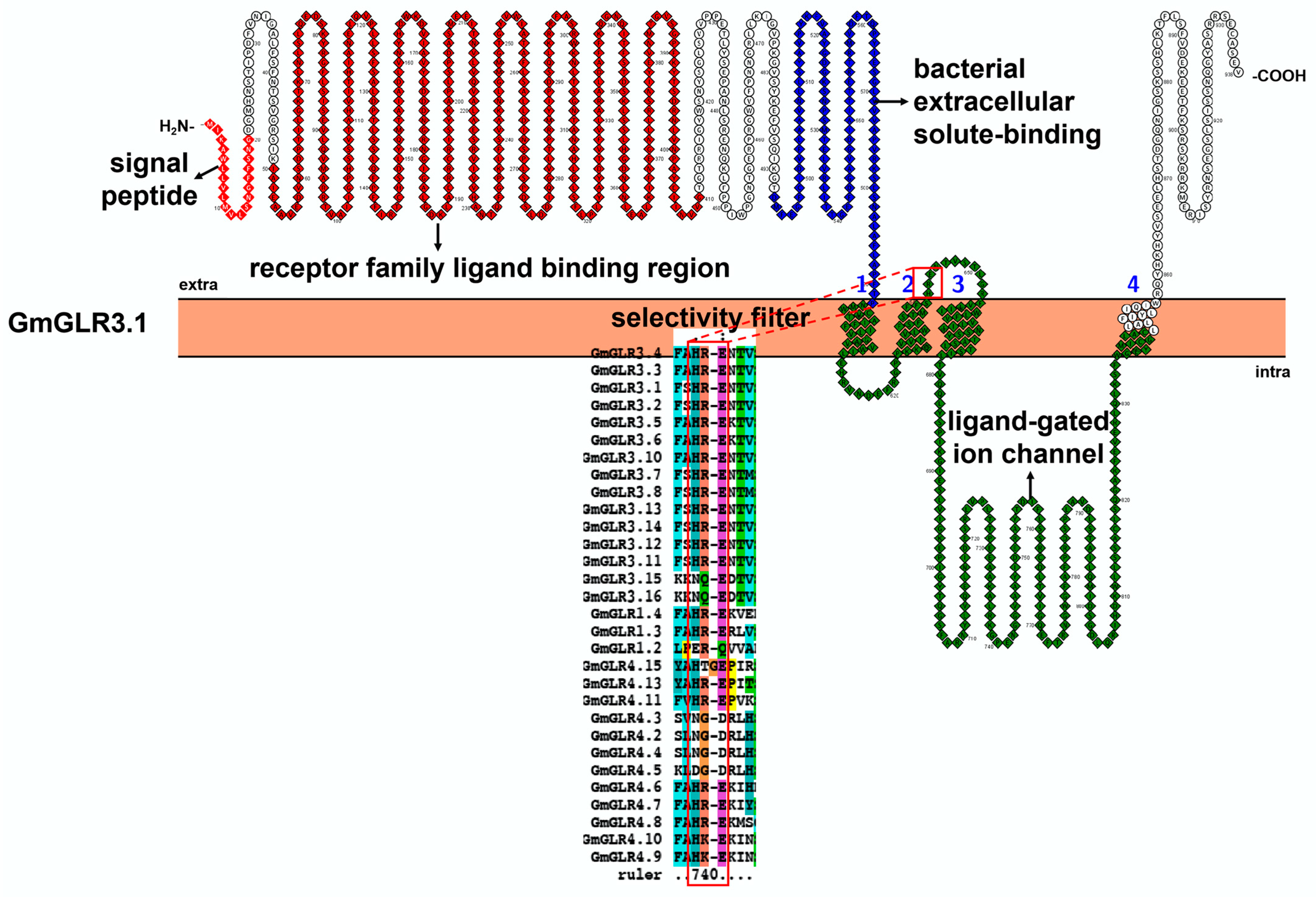
3.3. Characteristic Features and Roles of Glutamate Receptor-like (GLR) Protein Involved in Soybean Stress Response
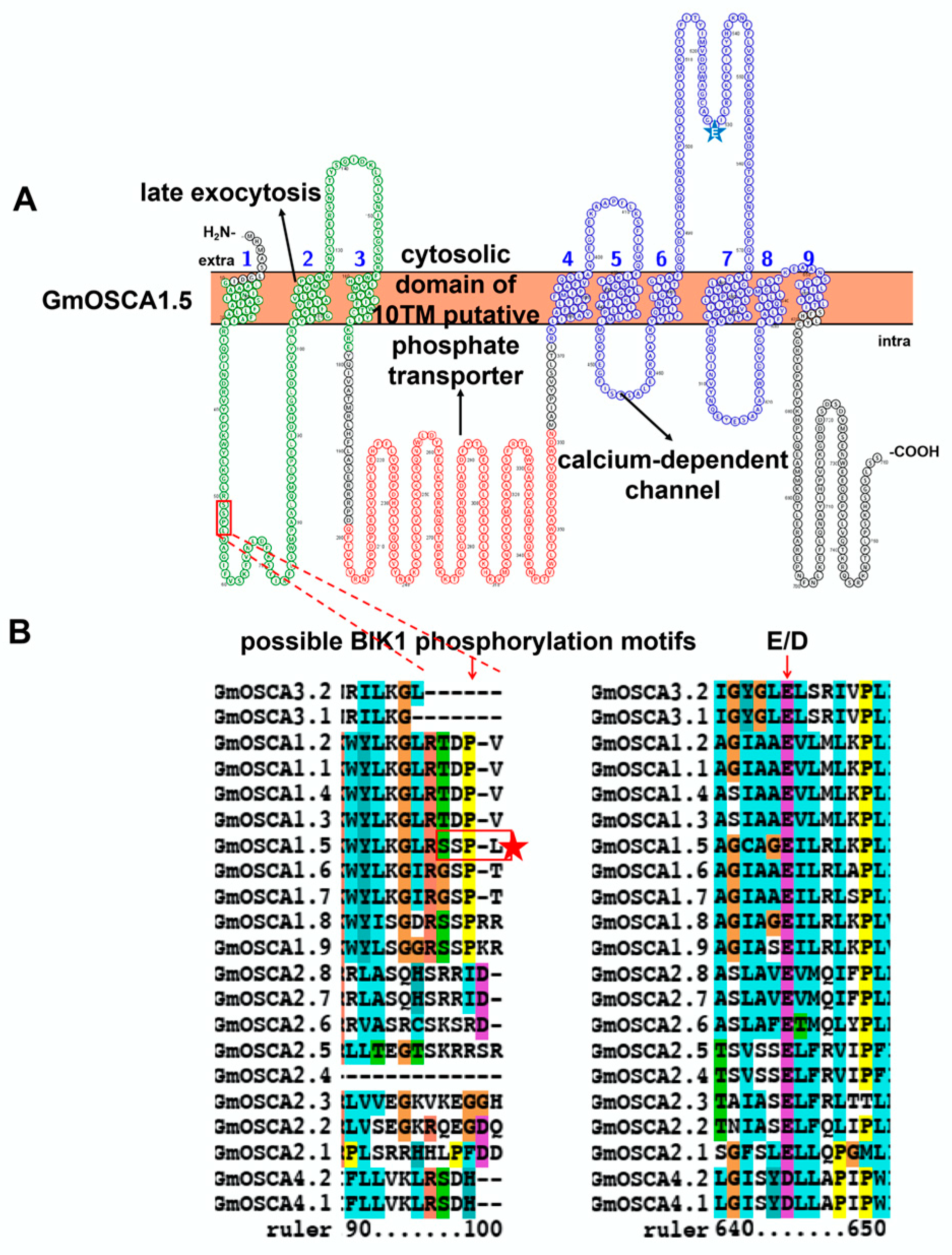
3.4. Characteristic Features and Roles of Hyperosmolality-Gated Calcium-Permeable Channel (OSCA) Involved in Soybean Stress Response
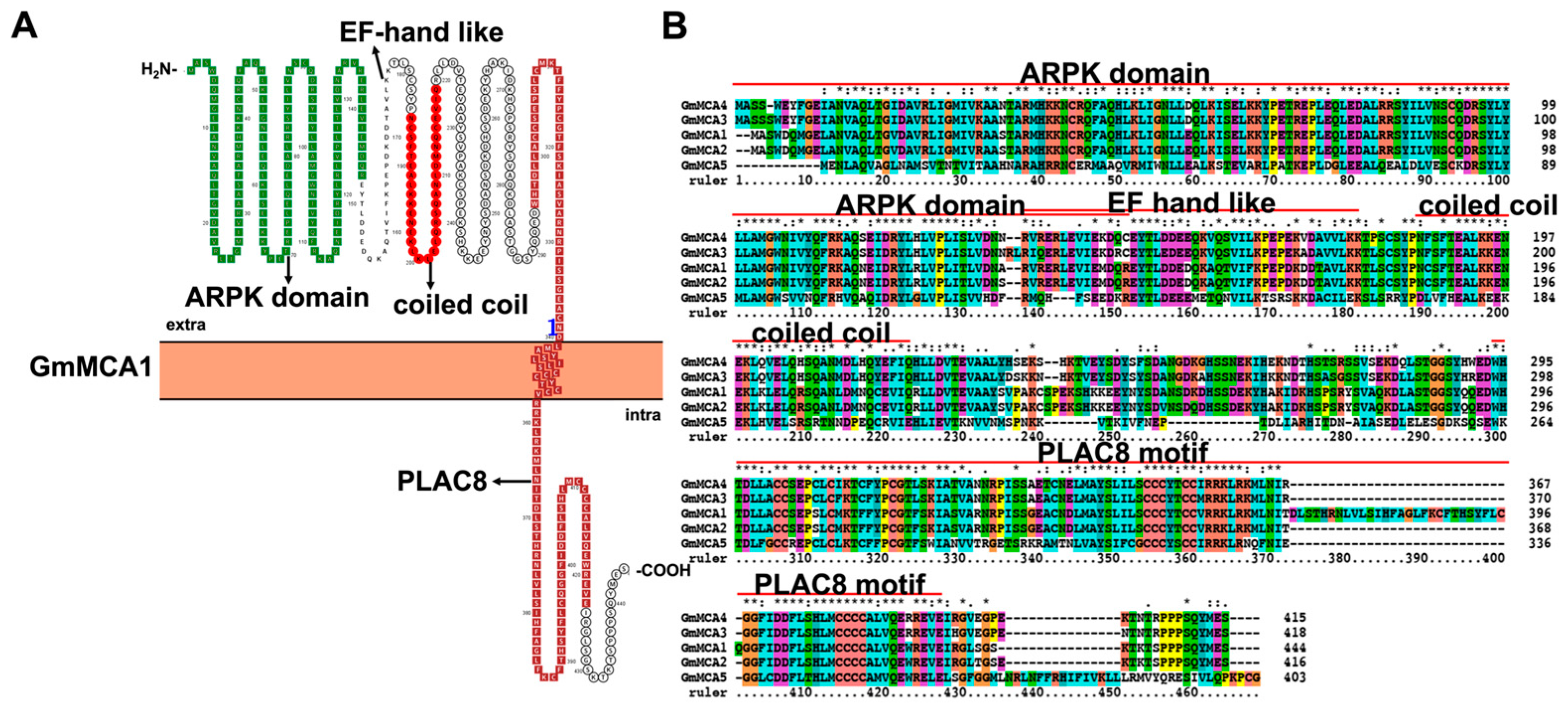
3.5. Characteristic Features and Roles of Mid1-Complementing Activity (MCA) Protein Antiporter Involved in Soybean Stress Response
3.6. Characteristic Features and Roles of Annexins Antiporter (ANNs) Involved in Soybean Stress Response
4. Conclusions and Prospect
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- White, P.J.; Broadley, M.R. Calcium in plants. Ann Bot 2003, 92, 487–511. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Wang, C.; Gao, Q.; Li, L.; Luan, S. Calcium spikes, waves and oscillations in plant development and biotic interactions. Nat Plants 2020, 6, 750–759. [Google Scholar] [CrossRef]
- Gao, Q.; Wang, C.; Xi, Y.; Shao, Q.; Luan, S. A receptor-channel trio conducts Ca2+ signalling for pollen tube reception. Nature 2022, 607, 534–539. [Google Scholar] [CrossRef]
- Dindas, J.; Dreyer, I.; Huang, S.; Hedrich, R.; Roelfsema, M.R.G. A voltage-dependent Ca2+ homeostat operates in the plant vacuolar membrane. New Phytol. 2021, 230, 1449–1460. [Google Scholar] [CrossRef]
- Roelfsema, M.R.; Hedrich, R. Making sense out of Ca2+ signals: Their role in regulating stomatal movements. Plant Cell Environ. 2010, 33, 305–321. [Google Scholar] [CrossRef] [PubMed]
- Stael, S.; Wurzinger, B.; Mair, A.; Mehlmer, N.; Vothknecht, U.C.; Teige, M. Plant organellar calcium signalling: An emerging field. J. Exp. Bot. 2012, 63, 1525–1542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Connorton, J.M.; Hirschi, K.D.; Pittman, J.K. Mechanism and evolution of calcium transport across the plant plasma membrane. In The Plant Plasma Membrane; Murphy, A.S., Schulz, B., Peer, W., Eds. Springer: Berlin/Heidelberg, Germany, 2011; pp. 275–289. [Google Scholar]
- Zeng, H.; Zhao, B.; Wu, H.; Zhu, Y.; Chen, H. Comprehensive in silico characterization and expression profiling of nine gene families associated with calcium transport in soybean. Agronomy 2020, 10, 1539. [Google Scholar] [CrossRef]
- Chiu, J.C.; Brenner, E.D.; DeSalle, R.; Nitabach, M.N.; Holmes, T.C.; Coruzzi, G.M. Phylogenetic and expression analysis of the Glutamate-Receptor-Like gene family in Arabidopsis thaliana. Mol. Biol. Evol. 2002, 19, 1066–1082. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Yuan, F.; Wen, Z.; Li, Y.; Wang, F.; Zhu, T.; Zhuo, W.; Jin, X.; Wang, Y.; Zhao, H.; et al. Genome-wide survey and expression analysis of the OSCA gene family in rice. BMC Plant Biol. 2015, 15, 261. [Google Scholar] [CrossRef] [Green Version]
- Jami, S.K.; Clark, G.B.; Ayele, B.T.; Roux, S.J.; Kirti, P.B. Identification and characterization of annexin gene family in rice. Plant Cell Rep. 2012, 31, 813–825. [Google Scholar] [CrossRef]
- Ding, S.-Y.; Feng, X.; Du, H.; Wang, H. Genome-wide analysis of maize OSCA family members and their involvement in drought stress. PeerJ 2019, 7, e6765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taneja, M.; Tyagi, S.; Sharma, S.; Upadhyay, S.K. Ca2+/Cation Antiporters (CaCA): Identification, characterization and expression profiling in bread wheat (Triticum aestivum L.). Front. Plant Sci. 2016, 7, 1775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clemente, T.E.; Cahoon, E.B. Soybean oil: Genetic approaches for modification of functionality and total content. Plant Physiol. 2009, 151, 1030–1040. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Gu, X.; Wu, Z.; Qin, T.; Guo, L.; Wang, T.; Zhang, L.; Jiang, G. Assessing the effects of elevated ozone on physiology, growth, yield and quality of soybean in the past 40 years: A meta-analysis. Ecotoxicol. Environ. Saf. 2021, 208, 111644. [Google Scholar] [CrossRef]
- Phang, T.H.; Shao, G.; Lam, H.M. Salt tolerance in soybean. J. Integr. Plant Biol. 2008, 50, 1196–1212. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, S.; Luthria, D.; Bae, H.; Lakshman, D.; Mitra, A. Transgenic soybeans and soybean protein analysis: An overview. J. Agric. Food Chem. 2013, 61, 11736–11743. [Google Scholar] [CrossRef]
- Pedersen, C.N.; Axelsen, K.B.; Harper, J.F.; Palmgren, M.G. Evolution of plant p-type ATPases. Front. Plant Sci. 2012, 3, 31. [Google Scholar] [CrossRef] [Green Version]
- Fuglsang, A.T.; Palmgren, M. Proton and calcium pumping P-type ATPases and their regulation of plant responses to the environment. Plant Physiol. 2021, 187, 1856–1875. [Google Scholar] [CrossRef]
- Chung, W.S.; Lee, S.H.; Kim, J.C.; Heo, W.D.; Kim, M.C.; Park, C.Y.; Park, H.C.; Lim, C.O.; Kim, W.B.; Harper, J.F.; et al. Identification of a calmodulin-regulated soybean Ca2+-ATPase (SCA1) that is located in the plasma membrane. Plant Cell 2000, 12, 1393–1407. [Google Scholar] [CrossRef] [Green Version]
- Liang, F.; Cunningham, K.W.; Harper, J.F.; Sze, H. ECA1 complements yeast mutants defective in Ca2+ pumps and encodes an endoplasmic reticulum-type Ca2+-ATPase in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 1997, 94, 8579–8584. [Google Scholar] [CrossRef]
- Geisler, M.; Frangne, N.; Gomès, E.; Martinoia, E.; Palmgren, M.G. The ACA4 gene of Arabidopsis encodes a vacuolar membrane calcium pump that improves salt tolerance in yeast. Plant Physiol. 2000, 124, 1814–1827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.Y.; Seo, S.B.; Lee, S.J.; Na, J.G.; Kim, Y.J. Mutation in PMR1, a Ca2+-ATPase in Golgi, confers salt tolerance in Saccharomyces cerevisiae by inducing expression of PMR2, an Na+-ATPase in plasma membrane. J. Biol. Chem. 2001, 276, 28694–28699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, M.; Jia, B.; Cui, N.; Wen, Y.; Duanmu, H.; Yu, Q.; Xiao, J.; Sun, X.; Zhu, Y. Functional characterization of a Glycine soja Ca2+-ATPase in salt-alkaline stress responses. Plant Mol. Biol. 2016, 90, 419–434. [Google Scholar] [CrossRef] [PubMed]
- Omasits, U.; Ahrens, C.H.; Müller, S.; Wollscheid, B. Protter: Interactive protein feature visualization and integration with experimental proteomic data. Bioinformatics 2014, 30, 884–886. [Google Scholar] [CrossRef] [Green Version]
- Hwang, I.; Harper, J.F.; Liang, F.; Sze, H. Calmodulin activation of an endoplasmic reticulum-located calcium pump involves an interaction with the N-terminal autoinhibitory domain. Plant Physiol. 2000, 122, 157–168. [Google Scholar] [CrossRef] [Green Version]
- Hwang, I.; Sze, H.; Harper, J.F. A calcium-dependent protein kinase can inhibit a calmodulin-stimulated Ca2+ pump (ACA2) located in the endoplasmic reticulum of Arabidopsis. Proc. Natl. Acad. Sci. USA 2000, 97, 6224–6229. [Google Scholar] [CrossRef] [Green Version]
- Mills, R.F.; Doherty, M.L.; Lopez-Marques, R.L.; Weimar, T.; Dupree, P.; Palmgren, M.G.; Pittman, J.K.; Williams, L.E. ECA3, a Golgi-localized P2A-type ATPase, plays a crucial role in manganese nutrition in Arabidopsis. Plant Physiol. 2008, 146, 116–128. [Google Scholar] [CrossRef] [Green Version]
- Pi, E.; Qu, L.; Hu, J.; Huang, Y.; Qiu, L.; Lu, H.; Jiang, B.; Liu, C.; Peng, T.; Zhao, Y.; et al. Mechanisms of soybean roots’ tolerances to salinity revealed by proteomic and phosphoproteomic comparisons between two cultivars. Mol Cell Proteom. 2016, 15, 266–288. [Google Scholar] [CrossRef] [Green Version]
- Malle, S.; Morrison, M.; Belzile, F. Identification of loci controlling mineral element concentration in soybean seeds. BMC Plant Biol. 2020, 20, 419. [Google Scholar] [CrossRef]
- Kim, K.H.; Kang, Y.J.; Kim, D.H.; Yoon, M.Y.; Moon, J.K.; Kim, M.Y.; Van, K.; Lee, S.H. RNA-Seq analysis of a soybean near-isogenic line carrying bacterial leaf pustule-resistant and -susceptible alleles. DNA Res. Int. J. Rapid Publ. Rep. Genes Genomes 2011, 18, 483–497. [Google Scholar] [CrossRef]
- Zeng, W.; Sun, Z.; Cai, Z.; Chen, H.; Lai, Z.; Yang, S.; Tang, X. Comparative transcriptome analysis of soybean response to bean pyralid larvae. BMC Genom. 2017, 18, 871. [Google Scholar] [CrossRef] [PubMed]
- Kofsky, J.; Zhang, H.; Song, B.H. Novel resistance strategies to soybean cyst nematode (SCN) in wild soybean. Sci. Rep. 2021, 11, 7967. [Google Scholar] [CrossRef] [PubMed]
- Clarke, V.C.; Loughlin, P.C.; Gavrin, A.; Chen, C.; Brear, E.M.; Day, D.A.; Smith, P.M. Proteomic analysis of the soybean symbiosome identifies new symbiotic proteins. Mol. Cell Proteom. 2015, 14, 1301–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, B.; Ichida, A.; Wang, Y.; Gens, J.S.; Pickard, B.G.; Harper, J.F. Identification of a calmodulin-regulated Ca2+-ATPase in the endoplasmic reticulum. Plant Physiol. 1999, 119, 1165–1176. [Google Scholar] [CrossRef] [Green Version]
- Iwano, M.; Igarashi, M.; Tarutani, Y.; Kaothien-Nakayama, P.; Nakayama, H.; Moriyama, H.; Yakabe, R.; Entani, T.; Shimosato-Asano, H.; Ueki, M.; et al. A pollen coat-inducible autoinhibited Ca2+-ATPase expressed in stigmatic papilla cells is required for compatible pollination in the Brassicaceae. Plant Cell 2014, 26, 636–649. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Yan, J.; Du, X.; Hua, J. Overlapping and differential roles of plasma membrane calcium ATPases in Arabidopsis growth and environmental responses. J. Exp. Bot. 2018, 69, 2693–2703. [Google Scholar] [CrossRef] [Green Version]
- Limonta, M.; Romanowsky, S.; Olivari, C.; Bonza, M.C.; Luoni, L.; Rosenberg, A.; Harper, J.F.; De Michelis, M.I. ACA12 is a deregulated isoform of plasma membrane Ca2+-ATPase of Arabidopsis thaliana. Plant Mol. Biol. 2014, 84, 387–397. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Berkelman, T.; Franklin, A.E.; Hoffman, N.E. Characterization of a gene encoding a Ca2+-ATPase-like protein in the plastid envelope. Proc. Natl. Acad. Sci. USA 1993, 90, 10066–10070. [Google Scholar] [CrossRef] [Green Version]
- Schiøtt, M.; Romanowsky, S.M.; Bækgaard, L.; Jakobsen, M.K.; Palmgren, M.G.; Harper, J.F. A plant plasma membrane Ca2+ pump is required for normal pollen tube growth and fertilization. Proc. Natl. Acad. Sci. USA 2004, 101, 9502–9507. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhang, X.; Wang, R.; Li, W. The plasma membrane-localised Ca2+-ATPase ACA8 plays a role in sucrose signalling involved in early seedling development in Arabidopsis. Plant Cell Rep. 2014, 33, 755–766. [Google Scholar] [CrossRef]
- Nguyen, H.H.; Lee, M.H.; Song, K.; Ahn, G.; Lee, J.; Hwang, I. The A/ENTH Domain-Containing Protein AtECA4 Is an Adaptor Protein Involved in Cargo Recycling from the trans-Golgi Network/Early Endosome to the Plasma Membrane. Mol. Plant 2018, 11, 568–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, G.Z.; Wang, H.W.; Huang, J.; Tian, A.G.; Wang, Y.J.; Zhang, J.S.; Chen, S.Y. A putative plasma membrane cation/proton antiporter from soybean confers salt tolerance in Arabidopsis. Plant Mol. Biol. 2005, 59, 809–820. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Connorton, J.M.; Guo, Y.; Li, X.; Shigaki, T.; Hirschi, K.D.; Pittman, J.K. Functional studies of split Arabidopsis Ca2+/H+ exchangers. J. Biol. Chem. 2009, 284, 34075–34083. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Yang, A.; James, A.T. Comparative proteomic analysis of drought response in roots of two soybean genotypes. Crop Pasture Sci. 2017, 68, 609–619. [Google Scholar] [CrossRef]
- Pittman, J.K.; Hirschi, K.D. CAX-ing a wide net: Cation/H+ transporters in metal remediation and abiotic stress signalling. Plant Biol. 2016, 18, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Emery, L.; Whelan, S.; Hirschi, K.; Pittman, J. Protein phylogenetic analysis of Ca2+/cation antiporters and insights into their evolution in plants. Front. Plant Sci. 2012, 3, 1. [Google Scholar] [CrossRef] [Green Version]
- Pittman, J.K.; Hirschi, K.D. Phylogenetic analysis and protein structure modelling identifies distinct Ca2+/Cation antiporters and conservation of gene family structure within Arabidopsis and rice species. Rice 2016, 9, 3. [Google Scholar] [CrossRef] [Green Version]
- Amagaya, K.; Shibuya, T.; Nishiyama, M.; Kato, K.; Kanayama, Y. Characterization and expression analysis of the Ca2+/cation antiporter gene family in tomatoes. Plants 2019, 9, 25. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Shigaki, T.; Mei, H.; Guo, Y.Q.; Cheng, N.H.; Hirschi, K.D. Interaction between Arabidopsis Ca2+/H+ exchangers CAX1 and CAX3. J. Biol. Chem. 2009, 284, 4605–4615. [Google Scholar] [CrossRef] [Green Version]
- Hocking, B.; Conn, S.J.; Manohar, M.; Xu, B.; Athman, A.; Stancombe, M.A.; Webb, A.R.; Hirschi, K.D.; Gilliham, M. Heterodimerization of Arabidopsis calcium/proton exchangers contributes to regulation of guard cell dynamics and plant defense responses. J. Exp. Bot. 2017, 68, 4171–4183. [Google Scholar] [CrossRef]
- Shaul, O.; Hilgemann, D.W.; de-Almeida-Engler, J.; Van Montagu, M.; Inz, D.; Galili, G. Cloning and characterization of a novel Mg2+/H+ exchanger. EMBO J. 1999, 18, 3973–3980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, J.; Tian, H.; Park, S.; Sreevidya, C.S.; Ward, J.M.; Hirschi, K.D. AtCCX3 is an Arabidopsis endomembrane H+-dependent K+ transporter. Plant Physiol. 2008, 148, 1474–1486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Zhang, G.; Gonzales, N.; Guo, Y.; Hu, H.; Park, S.; Zhao, J. Ca2+-regulated and diurnal rhythm-regulated Na+/Ca2+ exchanger AtNCL affects flowering time and auxin signalling in Arabidopsis. Plant Cell Environ. 2016, 39, 377–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corso, M.; Doccula, F.G.; de Melo, J.R.F.; Costa, A.; Verbruggen, N. Endoplasmic reticulum-localized CCX2 is required for osmotolerance by regulating ER and cytosolic Ca2+ dynamics in Arabidopsis. Proc. Natl. Acad. Sci. USA 2018, 115, 3966–3971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, X.; Lytton, J. The cation/Ca2+ exchanger superfamily: Phylogenetic analysis and structural implications. Mol. Biol. Evol. 2004, 21, 1692–1703. [Google Scholar] [CrossRef] [Green Version]
- Verret, F.; Wheeler, G.; Taylor, A.R.; Farnham, G.; Brownlee, C. Calcium channels in photosynthetic eukaryotes: Implications for evolution of calcium-based signalling. New Phytol. 2010, 187, 23–43. [Google Scholar] [CrossRef]
- Nawaz, Z.; Kakar, K.U.; Saand, M.A.; Shu, Q.Y. Cyclic nucleotide-gated ion channel gene family in rice, identification, characterization and experimental analysis of expression response to plant hormones, biotic and abiotic stresses. BMC Genom. 2014, 15, 853. [Google Scholar] [CrossRef] [Green Version]
- Mori, I.C.; Nobukiyo, Y.; Nakahara, Y.; Shibasaka, M.; Furuichi, T.; Katsuhara, M. A Cyclic Nucleotide-Gated Channel, HvCNGC2-3, is activated by the co-presence of Na+ and K+ and permeable to Na+ and K+ non-selectively. Plants 2018, 7, 61. [Google Scholar] [CrossRef] [Green Version]
- Nawaz, Z.; Kakar, K.U.; Ullah, R.; Yu, S.; Zhang, J.; Shu, Q.Y.; Ren, X.L. Genome-wide identification, evolution and expression analysis of cyclic nucleotide-gated channels in tobacco (Nicotiana tabacum L.). Genomics 2019, 111, 142–158. [Google Scholar] [CrossRef]
- Bridges, D.; Fraser, M.E.; Moorhead, G.B. Cyclic nucleotide binding proteins in the Arabidopsis thaliana and Oryza sativa genomes. BMC Bioinform. 2005, 6, 6. [Google Scholar] [CrossRef]
- Saand, M.A.; Xu, Y.P.; Munyampundu, J.P.; Li, W.; Zhang, X.R.; Cai, X.Z. Phylogeny and evolution of plant cyclic nucleotide-gated ion channel (CNGC) gene family and functional analyses of tomato CNGCs. DNA Res. Int. J. Rapid Publ. Rep. Genes Genomes 2015, 22, 471–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saand, M.A.; Xu, Y.P.; Li, W.; Wang, J.P.; Cai, X.Z. Cyclic nucleotide gated channel gene family in tomato: Genome-wide identification and functional analyses in disease resistance. Front. Plant Sci. 2015, 6, 303. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Yang, S.; Ren, J.; Ye, X.; Jiang, X.; Liu, Z. Genome-wide identification and functional analysis of the cyclic nucleotide-gated channel gene family in Chinese cabbage. 3 Biotech 2019, 9, 114. [Google Scholar] [CrossRef] [PubMed]
- Christopher, D.A.; Borsics, T.; Yuen, C.Y.; Ullmer, W.; Andème-Ondzighi, C.; Andres, M.A.; Kang, B.H.; Staehelin, L.A. The cyclic nucleotide gated cation channel AtCNGC10 traffics from the ER via Golgi vesicles to the plasma membrane of Arabidopsis root and leaf cells. BMC Plant Biol. 2007, 7, 48. [Google Scholar] [CrossRef] [Green Version]
- Yuen, C.C.Y.; Christopher, D.A. The Group IV-A cyclic nucleotide-gated channels, CNGC19 and CNGC20, localize to the vacuole membrane in Arabidopsis thaliana. AoB Plants 2013, 5, plt012. [Google Scholar] [CrossRef]
- Zelman, A.K.; Dawe, A.; Gehring, C.; Berkowitz, G.A. Evolutionary and structural perspectives of plant cyclic nucleotide-gated cation channels. Front. Plant Sci. 2012, 3, 95. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Chai, X.; Gao, Q.; Zhou, L.; Zhang, S.; Li, L.; Luan, S. Dynamic interactions of plant CNGC subunits and calmodulins drive oscillatory Ca2+ channel activities. Dev. Cell 2019, 48, 710–725.e715. [Google Scholar] [CrossRef] [Green Version]
- Chin, K.; DeFalco, T.A.; Moeder, W.; Yoshioka, K. The Arabidopsis cyclic nucleotide-gated ion channels AtCNGC2 and AtCNGC4 work in the same signaling pathway to regulate pathogen defense and floral transition. Plant Physiol. 2013, 163, 611–624. [Google Scholar] [CrossRef] [Green Version]
- Jammes, F.; Hu, H.C.; Villiers, F.; Bouten, R.; Kwak, J.M. Calcium-permeable channels in plant cells. FEBS J. 2011, 278, 4262–4276. [Google Scholar] [CrossRef]
- Pandey, G.K.; Sanyal, S.K. Plant ligand-gated channels 2: CNGC. In Functional Dissection of Calcium Homeostasis and Transport Machinery in Plants; Springer International Publishing: Cham, Switherlands, 2021; pp. 63–73. [Google Scholar]
- Fischer, C.; Kugler, A.; Hoth, S.; Dietrich, P. An IQ domain mediates the interaction with calmodulin in a plant cyclic nucleotide-gated channel. Plant Cell Physiol. 2013, 54, 573–584. [Google Scholar] [CrossRef]
- Chin, K.; Moeder, W.; Abdel-Hamid, H.; Shahinas, D.; Gupta, D.; Yoshioka, K. Importance of the alphaC-helix in the cyclic nucleotide binding domain for the stable channel regulation and function of cyclic nucleotide gated ion channels in Arabidopsis. J. Exp. Bot. 2010, 61, 2383–2393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Zeng, W.; Jiang, Y. Tuning the ion selectivity of two-pore channels. Proc. Natl. Acad. Sci. USA 2017, 114, 1009–1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaślan, D.; Mueller, T.D.; Becker, D.; Schultz, J.; Cuin, T.A.; Marten, I.; Dreyer, I.; Schönknecht, G.; Hedrich, R. Gating of the two-pore cation channel AtTPC1 in the plant vacuole is based on a single voltage-sensing domain. Plant Biol. 2016, 18, 750–760. [Google Scholar] [CrossRef] [PubMed]
- Schulze, C.; Sticht, H.; Meyerhoff, P.; Dietrich, P. Differential contribution of EF-hands to the Ca2+-dependent activation in the plant two-pore channel TPC1. Plant J. Cell Mol. Biol. 2011, 68, 424–432. [Google Scholar] [CrossRef]
- Peiter, E.; Maathuis, F.J.; Mills, L.N.; Knight, H.; Pelloux, J.; Hetherington, A.M.; Sanders, D. The vacuolar Ca2+-activated channel TPC1 regulates germination and stomatal movement. Nature 2005, 434, 404–408. [Google Scholar] [CrossRef]
- Dadacz-Narloch, B.; Kimura, S.; Kurusu, T.; Farmer, E.E.; Becker, D.; Kuchitsu, K.; Hedrich, R. On the cellular site of two-pore channel TPC1 action in the Poaceae. New Phytol. 2013, 200, 663–674. [Google Scholar] [CrossRef]
- Hashimoto, K.; Koselski, M.; Tsuboyama, S.; Dziubinska, H.; Trebacz, K.; Kuchitsu, K. Functional analyses of the two distinctive types of Two-Pore Channels and the Slow Vacuolar Channel in Marchantia polymorpha. Plant Cell Physiol. 2022, 63, 163–175. [Google Scholar] [CrossRef]
- Islam, M.M.; Munemasa, S.; Hossain, M.A.; Nakamura, Y.; Mori, I.C.; Murata, Y. Roles of AtTPC1, vacuolar two pore channel 1, in Arabidopsis stomatal closure. Plant Cell Physiol. 2010, 51, 302–311. [Google Scholar] [CrossRef]
- Kurusu, T.; Yagala, T.; Miyao, A.; Hirochika, H.; Kuchitsu, K. Identification of a putative voltage-gated Ca2+ channel as a key regulator of elicitor-induced hypersensitive cell death and mitogen-activated protein kinase activation in rice. Plant J. Cell Mol. Biol. 2005, 42, 798–809. [Google Scholar] [CrossRef]
- Kurusu, T.; Hamada, H.; Koyano, T.; Kuchitsu, K. Intracellular localization and physiological function of a rice Ca2+-permeable channel OsTPC1. Plant Signal. Behav. 2012, 7, 1428–1430. [Google Scholar] [CrossRef]
- De Bortoli, S.; Teardo, E.; Szabò, I.; Morosinotto, T.; Alboresi, A. Evolutionary insight into the ionotropic glutamate receptor superfamily of photosynthetic organisms. Biophys. Chem. 2016, 218, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Price, M.B.; Jelesko, J.; Okumoto, S. Glutamate receptor homologs in plants: Functions and evolutionary origins. Front. Plant Sci. 2012, 3, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aouini, A.; Hernould, M.; Ariizumi, T.; Matsukura, C.; Ezura, H.; Asamizu, E. Overexpression of the tomato glutamate receptor-like genes SlGLR1.1 and SlGLR3.5 hinders Ca2+ utilization and promotes hypersensitivity to Na+ and K+ stresses. Plant Biotechnol. 2012, 29, 229–235. [Google Scholar] [CrossRef] [Green Version]
- Ni, J.; Yu, Z.; Du, G.; Zhang, Y.; Taylor, J.L.; Shen, C.; Xu, J.; Liu, X.; Wang, Y.; Wu, Y. Heterologous expression and functional analysis of rice GLUTAMATE RECEPTOR-LIKE family indicates its role in glutamate triggered calcium flux in rice roots. Rice 2016, 9, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabrice, M.R.; Jing, Y.; Jiang, X.; Xiong, C.; Liu, X.; Chen, J.; Jiao, H.; Zhou, H.; Zhao, Z.; Zhang, S.; et al. PbGLR3.3 regulates pollen tube growth in the mediation of Ca2+ influx in Pyrus bretschneideri. J. Plant Biol. 2018, 61, 217–226. [Google Scholar] [CrossRef]
- Philippe, F.; Verdu, I.; Morere-Le Paven, M.C.; Limami, A.M.; Planchet, E. Involvement of Medicago truncatula glutamate receptor-like channels in nitric oxide production under short-term water deficit stress. J. Plant Physiol. 2019, 236, 1–6. [Google Scholar] [CrossRef]
- Alfieri, A.; Doccula, F.G.; Pederzoli, R.; Grenzi, M.; Bonza, M.C.; Luoni, L.; Candeo, A.; Romano Armada, N.; Barbiroli, A.; Valentini, G.; et al. The structural bases for agonist diversity in an Arabidopsis thaliana glutamate receptor-like channel. Proc. Natl. Acad. Sci. USA 2020, 117, 752–760. [Google Scholar] [CrossRef]
- Yoshida, R.; Mori, I.C.; Kamizono, N.; Shichiri, Y.; Shimatani, T.; Miyata, F.; Honda, K.; Iwai, S. Glutamate functions in stomatal closure in Arabidopsis and fava bean. J. Plant Res. 2016, 129, 39–49. [Google Scholar] [CrossRef] [Green Version]
- Kurusu, T.; Kuchitsu, K.; Nakano, M.; Nakayama, Y.; Iida, H. Plant mechanosensing and Ca2+ transport. Trends Plant Sci. 2013, 18, 227–233. [Google Scholar] [CrossRef]
- Yuan, F.; Yang, H.; Xue, Y.; Kong, D.; Ye, R.; Li, C.; Zhang, J.; Theprungsirikul, L.; Shrift, T.; Krichilsky, B.; et al. OSCA1 mediates osmotic-stress-evoked Ca2+ increases vital for osmosensing in Arabidopsis. Nature 2014, 514, 367–371. [Google Scholar] [CrossRef]
- Hou, C.; Tian, W.; Kleist, T.J.; He, K.; Garcia, V.J.; Bai, F.; Hao, Y.; Luan, S.; Li, L. DUF221 proteins are a family of osmosensitive calcium-permeable cation channels conserved across eukaryotes. Cell Res. 2014, 24, 632–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Yang, J.; Jia, B.; Sun, M.; Liu, Y.; Yin, K.; Sun, X. Evolution and expression analysis of OSCA gene family in soybean. Chin. J. Oil Crop Sci. 2017, 39, 589–599. [Google Scholar]
- Edel, K.H.; Kudla, J. Increasing complexity and versatility: How the calcium signaling toolkit was shaped during plant land colonization. Cell Calcium 2015, 57, 231–246. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, J.; Sun, L. Structure of the hyperosmolality-gated calcium-permeable channel OSCA1.2. Nat. Commun. 2018, 9, 5060. [Google Scholar] [CrossRef] [Green Version]
- Thor, K.; Jiang, S.; Michard, E.; George, J.; Scherzer, S.; Huang, S.; Dindas, J.; Derbyshire, P.; Leitão, N.; DeFalco, T.A.; et al. The calcium-permeable channel OSCA1.3 regulates plant stomatal immunity. Nature 2020, 585, 569–573. [Google Scholar] [CrossRef]
- Yang, X.; Xu, Y.; Yang, F.; Magwanga, R.O.; Cai, X.; Wang, X.; Wang, Y.; Hou, Y.; Wang, K.; Liu, F.; et al. Genome-wide identification of OSCA gene family and their potential function in the regulation of dehydration and salt stress in Gossypium hirsutum. J. Cotton Res. 2019, 2, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.; Zhang, P.; Lu, X.; Wang, G.; Wang, Z.; Zhang, Q.; Zhang, X.; Wei, X.; Mei, F.; Wei, L.; et al. Systematic analysis of the maize OSCA genes revealing ZmOSCA family members involved in osmotic stress and ZmOSCA2.4 confers enhanced drought tolerance in transgenic Arabidopsis. Int. J. Mol. Sci. 2020, 21, 351. [Google Scholar] [CrossRef] [Green Version]
- Furuichi, T.; Iida, H.; Sokabe, M.; Tatsumi, H. Expression of Arabidopsis MCA1 enhanced mechanosensitive channel activity in the Xenopus laevis oocyte plasma membrane. Plant Signal. Behav. 2012, 7, 1022–1026. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, Y.; Katagiri, T.; Shinozaki, K.; Qi, Z.; Tatsumi, H.; Furuichi, T.; Kishigami, A.; Sokabe, M.; Kojima, I.; Sato, S.; et al. Arabidopsis plasma membrane protein crucial for Ca2+ influx and touch sensing in roots. Proc. Natl. Acad. Sci. USA 2007, 104, 3639–3644. [Google Scholar] [CrossRef] [Green Version]
- Nakano, M.; Iida, K.; Nyunoya, H.; Iida, H. Determination of structural regions important for Ca2+ uptake activity in Arabidopsis MCA1 and MCA2 expressed in yeast. Plant Cell Physiol. 2011, 52, 1915–1930. [Google Scholar] [CrossRef] [Green Version]
- Shigematsu, H.; Iida, K.; Nakano, M.; Chaudhuri, P.; Iida, H.; Nagayama, K. Structural characterization of the mechanosensitive channel candidate MCA2 from Arabidopsis thaliana. PLoS ONE 2014, 9, e87724. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, T.; Nakagawa, Y.; Mori, K.; Nakano, M.; Imamura, T.; Kataoka, H.; Terashima, A.; Iida, K.; Kojima, I.; Katagiri, T.; et al. MCA1 and MCA2 that mediate Ca2+ uptake have distinct and overlapping roles in Arabidopsis. Plant Physiol. 2010, 152, 1284–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, K.; Renhu, N.; Naito, M.; Nakamura, A.; Shiba, H.; Yamamoto, T.; Suzaki, T.; Iida, H.; Miura, K. Ca2+-permeable mechanosensitive channels MCA1 and MCA2 mediate cold-induced cytosolic Ca2+ increase and cold tolerance in Arabidopsis. Sci. Rep. 2018, 8, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hattori, T.; Otomi, Y.; Nakajima, Y.; Soga, K.; Wakabayashi, K.; Iida, H.; Hoson, T. MCA1 and MCA2 are involved in the response to hypergravity in Arabidopsis hypocotyls. Plants 2020, 9, 590. [Google Scholar] [CrossRef]
- Kurusu, T.; Yamanaka, T.; Nakano, M.; Takiguchi, A.; Ogasawara, Y.; Hayashi, T.; Iida, K.; Hanamata, S.; Shinozaki, K.; Iida, H.; et al. Involvement of the putative Ca2+-permeable mechanosensitive channels, NtMCA1 and NtMCA2, in Ca2+ uptake, Ca2+-dependent cell proliferation and mechanical stress-induced gene expression in tobacco (Nicotiana tabacum) BY-2 cells. J. Plant Res. 2012, 125, 555–568. [Google Scholar] [CrossRef]
- Kurusu, T.; Iida, H.; Kuchitsu, K. Roles of a putative mechanosensitive plasma membrane Ca2+-permeable channel OsMCA1 in generation of reactive oxygen species and hypo-osmotic signaling in rice. Plant Signal. Behav. 2012, 7, 796–798. [Google Scholar] [CrossRef] [Green Version]
- Kurusu, T.; Nishikawa, D.; Yamazaki, Y.; Gotoh, M.; Nakano, M.; Hamada, H.; Yamanaka, T.; Iida, K.; Nakagawa, Y.; Saji, H.; et al. Plasma membrane protein OsMCA1 is involved in regulation of hypo-osmotic shock-induced Ca2+ influx and modulates generation of reactive oxygen species in cultured rice cells. BMC Plant Biol. 2012, 12, 11. [Google Scholar] [CrossRef] [Green Version]
- Rosa, M.; Abraham-Juarez, M.J.; Lewis, M.W.; Fonseca, J.P.; Tian, W.; Ramirez, V.; Luan, S.; Pauly, M.; Hake, S. The maize mid-complementing activity homolog cell number regulator13/narrow odd dwarf coordinates organ growth and tissue patterning. Plant Cell 2017, 29, 474–490. [Google Scholar] [CrossRef] [Green Version]
- Konopka-Postupolska, D.; Clark, G. Annexins as overlooked regulators of membrane trafficking in plant cells. Int. J. Mol. Sci. 2017, 18, 863. [Google Scholar] [CrossRef] [Green Version]
- Laohavisit, A.; Shang, Z.; Rubio, L.; Cuin, T.A.; Very, A.A.; Wang, A.; Mortimer, J.C.; Macpherson, N.; Coxon, K.M.; Battey, N.H.; et al. Arabidopsis annexin1 mediates the radical-activated plasma membrane Ca2+- and K+-permeable conductance in root cells. Plant Cell 2012, 24, 1522–1533. [Google Scholar] [CrossRef] [Green Version]
- Clark, G.B.; Morgan, R.O.; Fernandez, M.P.; Roux, S.J. Evolutionary adaptation of plant annexins has diversified their molecular structures, interactions and functional roles. New Phytol. 2012, 196, 695–712. [Google Scholar] [CrossRef]
- Clark, G.B.; Sessions, A.; Eastburn, D.J.; Roux, S.J. Differential expression of members of the annexin multigene family in Arabidopsis. Plant Physiol. 2001, 126, 1072–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, R.O.; Pilar Fernandez, M. Distinct annexin subfamilies in plants and protists diverged prior to animal annexins and from a common ancestor. J. Mol. Evol. 1997, 44, 178–188. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.M.; Wei, X.K.; Liao, W.X.; Huang, L.H.; Zhang, H.; Liang, S.C.; Peng, H. Molecular analysis of the annexin gene family in soybean. Biol Plant 2013, 57, 655–662. [Google Scholar] [CrossRef]
- Jami, S.K.; Clark, G.B.; Ayele, B.T.; Ashe, P.; Kirti, P.B. Genome-wide comparative analysis of annexin superfamily in plants. PLoS ONE 2012, 7, e47801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baucher, M.; Oukouomi Lowe, Y.; Vandeputte, O.M.; Mukoko Bopopi, J.; Moussawi, J.; Vermeersch, M.; Mol, A.; El Jaziri, M.; Homblé, F.; Pérez-Morga, D. Ntann12 annexin expression is induced by auxin in tobacco roots. J. Exp. Bot. 2011, 62, 4055–4065. [Google Scholar] [CrossRef] [Green Version]
- Qiao, B.; Zhang, Q.; Liu, D.; Wang, H.; Yin, J.; Wang, R.; He, M.; Cui, M.; Shang, Z.; Wang, D.; et al. A calcium-binding protein, rice annexin OsANN1, enhances heat stress tolerance by modulating the production of H2O2. J. Exp. Bot. 2015, 66, 5853–5866. [Google Scholar] [CrossRef] [Green Version]
- Carrasco-Castilla, J.; Ortega-Ortega, Y.; J·uregui-Z˙Òiga, D.; Ju·rez-Verdayes, M.A.; Arthikala, M.-K.; Monroy-Morales, E.; Nava, N.; Santana, O.; S·nchez-LÛpez, R.; Quinto, C. Down-regulation of a Phaseolus vulgaris annexin impairs rhizobial infection and nodulation. Environ. Exp. Bot. 2018, 153, 108–119. [Google Scholar] [CrossRef]
- Wang, X.; Movahedi, A.; Wei, H.; Wu, X.; Zhang, J.-x.; Sun, W.; Li, D.; Zhuge, Q. Overexpression of PtAnnexin1 from Populus trichocarpa enhances salt and drought tolerance in transgenic poplars. Tree Genet. Genomes 2020, 16, 20. [Google Scholar] [CrossRef]
- Li, X.; Zhang, Q.; Yang, X.; Han, J.; Zhu, Z. OsANN3, a calcium-dependent lipid binding annexin is a positive regulator of ABA-dependent stress tolerance in rice. Plant Sci. Int. J. Exp. Plant Biol. 2019, 284, 212–220. [Google Scholar] [CrossRef]
- Zhang, Q.; Song, T.; Guan, C.; Gao, Y.; Ma, J.; Gu, X.; Qi, Z.; Wang, X.; Zhu, Z. OsANN4 modulates ROS production and mediates Ca2+ influx in response to ABA. BMC Plant Biol. 2021, 21, 474. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Song, T.; Han, J.; He, M.; Zhang, Q.; Zhu, Y.; Zhu, Z. A calcium-dependent lipid binding protein, OsANN10, is a negative regulator of osmotic stress tolerance in rice. Plant Sci. Int. J. Exp. Plant Biol. 2020, 293, 110420. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Ye, J.; Yang, Y.; Lin, H.; Yue, L.; Luo, J.; Long, Y.; Fu, H.; Liu, X.; Zhang, Y.; et al. The SOS2-SCaBP8 complex generates and fine-tunes an AtANN4-dependent calcium signature under salt stress. Dev. Cell 2019, 48, 697–709.e695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huh, S.M.; Noh, E.K.; Kim, H.G.; Jeon, B.W.; Bae, K.; Hu, H.C.; Kwak, J.M.; Park, O.K. Arabidopsis annexins AnnAt1 and AnnAt4 interact with each other and regulate drought and salt stress responses. Plant Cell Physiol. 2010, 51, 1499–1514. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Li, J.; Niu, X.; Deng, C.; Song, X.; Li, W.; Cheng, Z.; Xu, Q.a.; Zhang, B.; Guo, W. GhANN1 modulates the salinity tolerance by regulating ABA biosynthesis, ion homeostasis and phenylpropanoid pathway in cotton. Environ. Exp. Bot. 2021, 185, 104427. [Google Scholar] [CrossRef]
- Wang, X.; Yang, R.; Zhou, Y.; Gu, Z. A comparative transcriptome and proteomics analysis reveals the positive effect of supplementary Ca2+ on soybean sprout yield and nutritional qualities. J. Proteom. 2016, 143, 161–172. [Google Scholar] [CrossRef]
- Sugimoto, T.; Watanabe, K.; Yoshida, S.; Aino, M.; Furiki, M.; Shiono, M.; Matoh, T.; Biggs, A.R. Field application of calcium to reduce Phytophthora stem rot of soybean, and calcium distribution in plants. Plant Dis. 2010, 94, 812–819. [Google Scholar] [CrossRef] [Green Version]
- Ceasar, S.A.; Maharajan, T.; Hillary, V.E.; Ajeesh Krishna, T.P. Insights to improve the plant nutrient transport by CRISPR/Cas system. Biotechnol. Adv. 2022, 59, 107963. [Google Scholar] [CrossRef]
- Sathee, L.; Jagadhesan, B.; Pandesha, P.H.; Barman, D.; Adavi, B.S.; Nagar, S.; Krishna, G.K.; Tripathi, S.; Jha, S.K.; Chinnusamy, V. Genome editing targets for improving nutrient use efficiency and nutrient stress adaptation. Front. Genet. 2022, 13, 900897. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classification | Gene Name | Gene ID Wm82.a4.v1 | Subcellular Localization | Arabidopsis Orthologs | Arabidopsis Orthologs Subcellular Localization | Observations |
|---|---|---|---|---|---|---|
| Ca2+-ATPase | GmACA1 | Glyma.01g193600 | Plasma membrane [20] | AT4G37640 ACA2 | Endoplasmic reticulum [27,35] | dehydration, high salt, alkaline, bacterial leaf pustule [8,24,31] |
| GmACA2 | Glyma.02g186100 | Not reported | AT3G57330 ACA11 | Vacuole [36] | bean pyralid [32] | |
| GmACA4 | No correspondence | Not reported | AT3G63380 ACA12 | Plasma membrane [37,38] | bacterial leaf pustule [31] | |
| GmACA7 | Glyma.06g046000 | Not reported | AT1G27770 ACA1 | Chloroplast [39] | high salt, bean pyralid, and a new QTL associated with the calcium content of soybean seeds [29,30] | |
| GmACA8 | Glyma.07g004300 | Not reported | AT3G21180 ACA9 | Plasma membrane [40] | bean pyralid, nematode [32,33] | |
| GmACA11 | Glyma.09g061200 | Not reported | AT5G57110 ACA8 | Plasma membrane [37,41] | soybean symbiosome [34] | |
| GmACA14 | Glyma.11g048300 | Not reported | AT4G37640 ACA2 | Endoplasmic reticulum [27,35] | dehydration, high salt, alkaline, bacterial leaf pustule, and bean pyralid [8,24,31,32] | |
| GmACA23 | Glyma.19g136400 | Not reported | AT2G41560 ACA4 | Vacuole [22] | bean pyralid [32] | |
| GmACA24 | Glyma.19g159900 | Not reported | AT3G63380 ACA12 | Plasma membrane [37,38] | dehydration, high salt, and alkaline [8,24] | |
| GmACA27 | Glyma.15g167500 | Not reported | AT4G29900 ACA10 | Not reported | bean pyralid [32] | |
| GmECA1 | Glyma.03g175200 | Not reported | AT1G07670 ECA4 | Plasma membrane, TGN, Cytosol [42] | soybean symbiosome [34] | |
| GmECA5 | Glyma.19g175900 | Not reported | AT1G07670 ECA4 | Plasma membrane, TGN, Cytosol [42] | soybean symbiosome [34] | |
| Ca2+/cation antiporter | GmCAX5 | Glyma.07g149600 | Plasma membrane [43] | AT3G51860 AtCAX3 | Tonoplast [44] | PEG, ABA, Ca2+, Na+ and Li+ treatments [43] |
| Annexins | GmANN15 | Glyma.08g136200 | Not reported | AT5G65020 annexin 2 | Not reported | salt, dehydration, drought, and flooding stresses [8,45] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, B.; Li, Y.; Sun, X.; Sun, M. Structure, Function, and Applications of Soybean Calcium Transporters. Int. J. Mol. Sci. 2022, 23, 14220. https://doi.org/10.3390/ijms232214220
Jia B, Li Y, Sun X, Sun M. Structure, Function, and Applications of Soybean Calcium Transporters. International Journal of Molecular Sciences. 2022; 23(22):14220. https://doi.org/10.3390/ijms232214220
Chicago/Turabian StyleJia, Bowei, Yuan Li, Xiaoli Sun, and Mingzhe Sun. 2022. "Structure, Function, and Applications of Soybean Calcium Transporters" International Journal of Molecular Sciences 23, no. 22: 14220. https://doi.org/10.3390/ijms232214220
APA StyleJia, B., Li, Y., Sun, X., & Sun, M. (2022). Structure, Function, and Applications of Soybean Calcium Transporters. International Journal of Molecular Sciences, 23(22), 14220. https://doi.org/10.3390/ijms232214220