
Berbamine Reduces Chloroquine-Induced Itch in Mice through Inhibition of MrgprX1


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
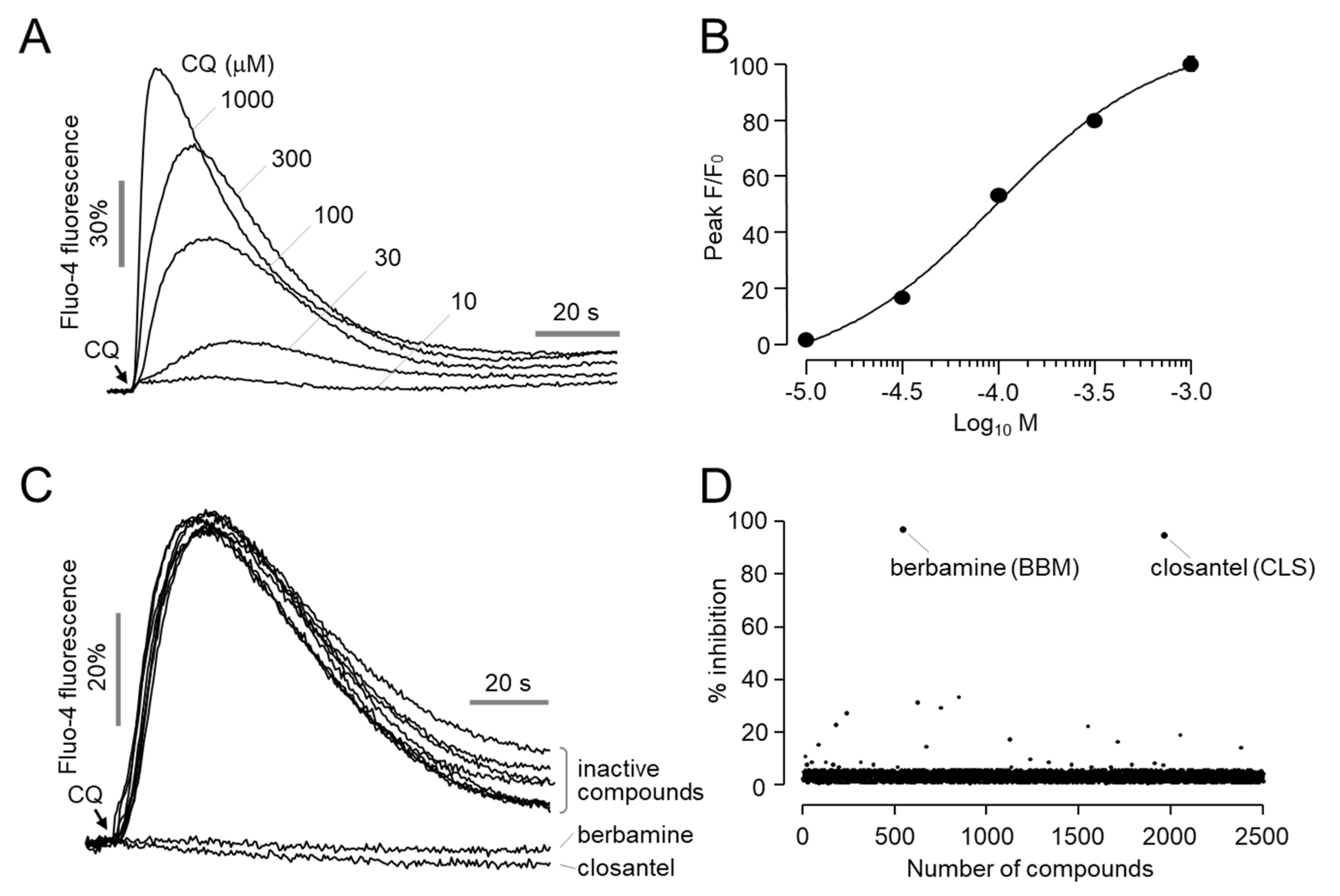
2.1. Berbamine, a Novel Antagonist of MrgprX1
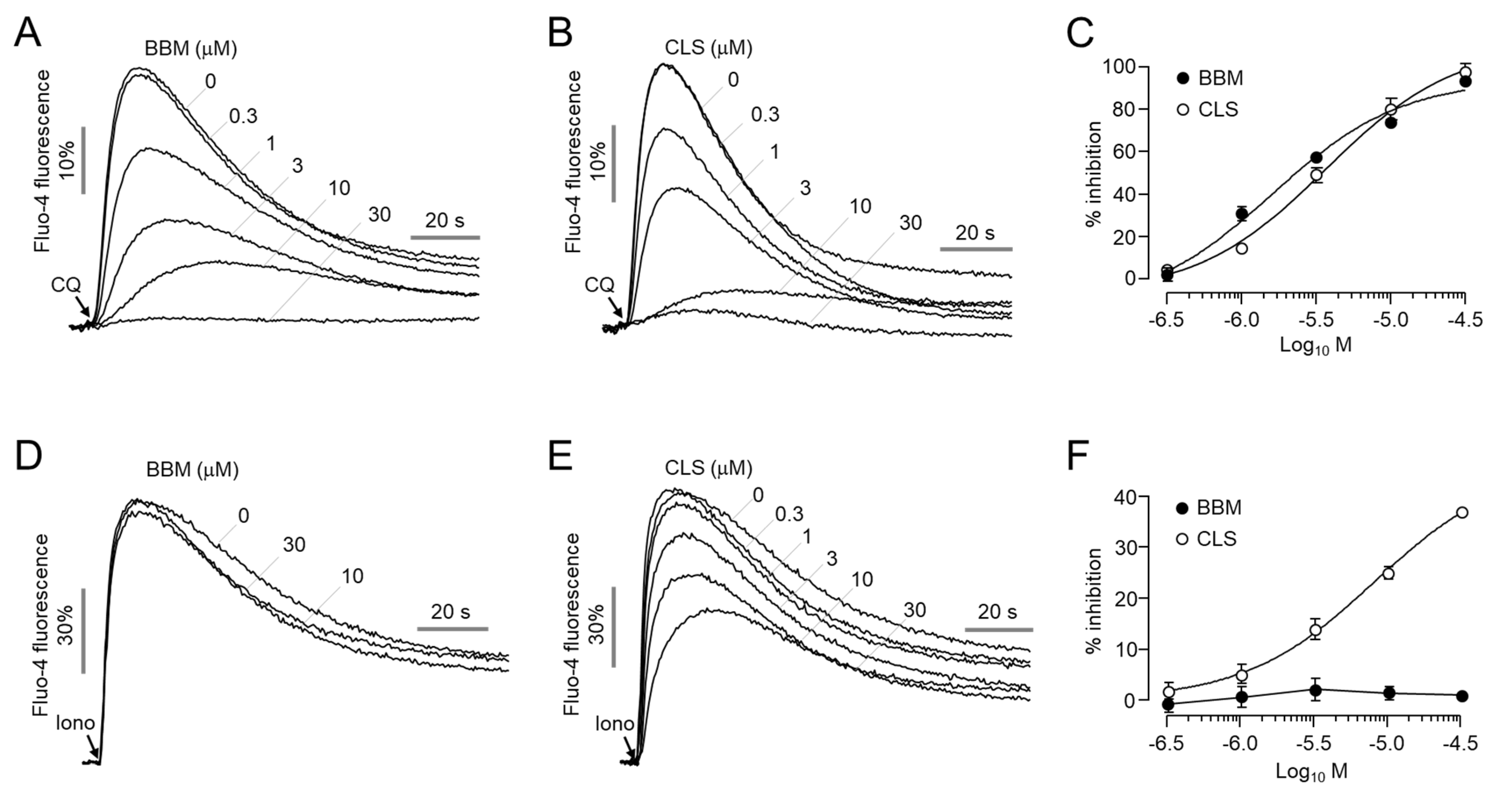
2.2. Effect of Berbamine and Closantel on CQ- and Ionomycin-Induced Increases in Intracellular Calcium Levels
2.3. Berbamine Has No Effect on Other Pruritogenic GPCRs except for MrgprX1
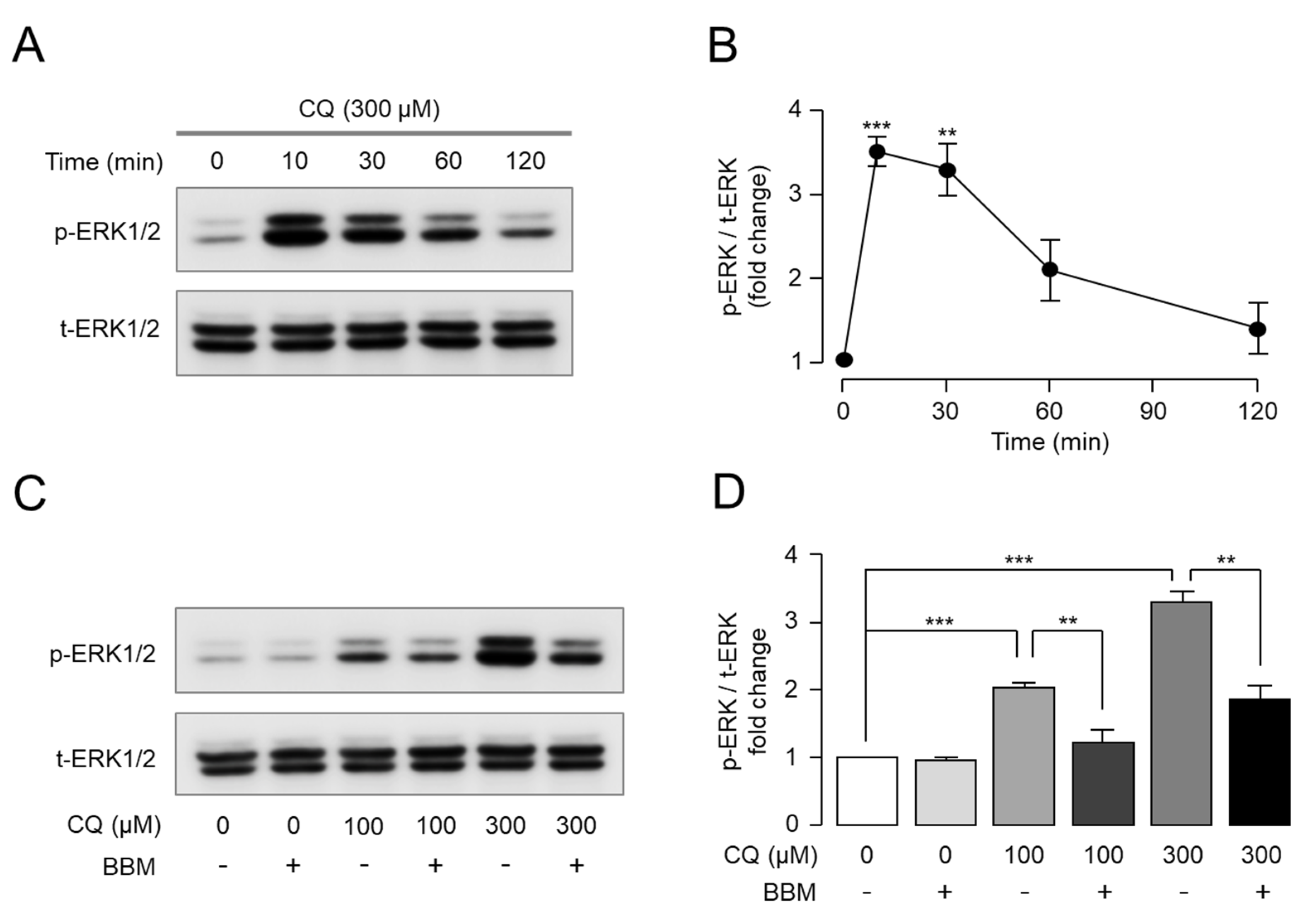
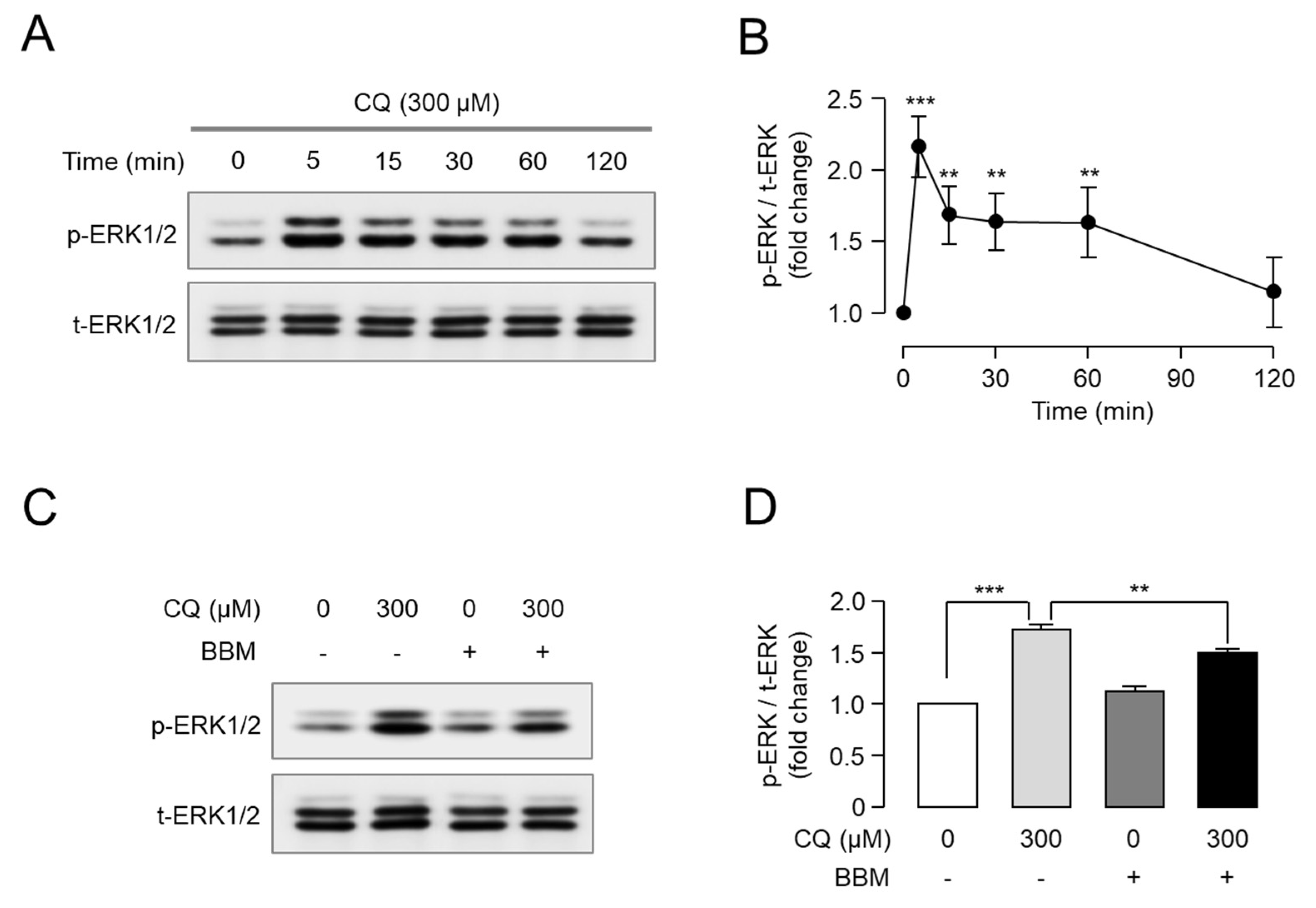
2.4. Berbamine Inhibits CQ-Induced ERK1/2 Phosphorylation through MrgprX1
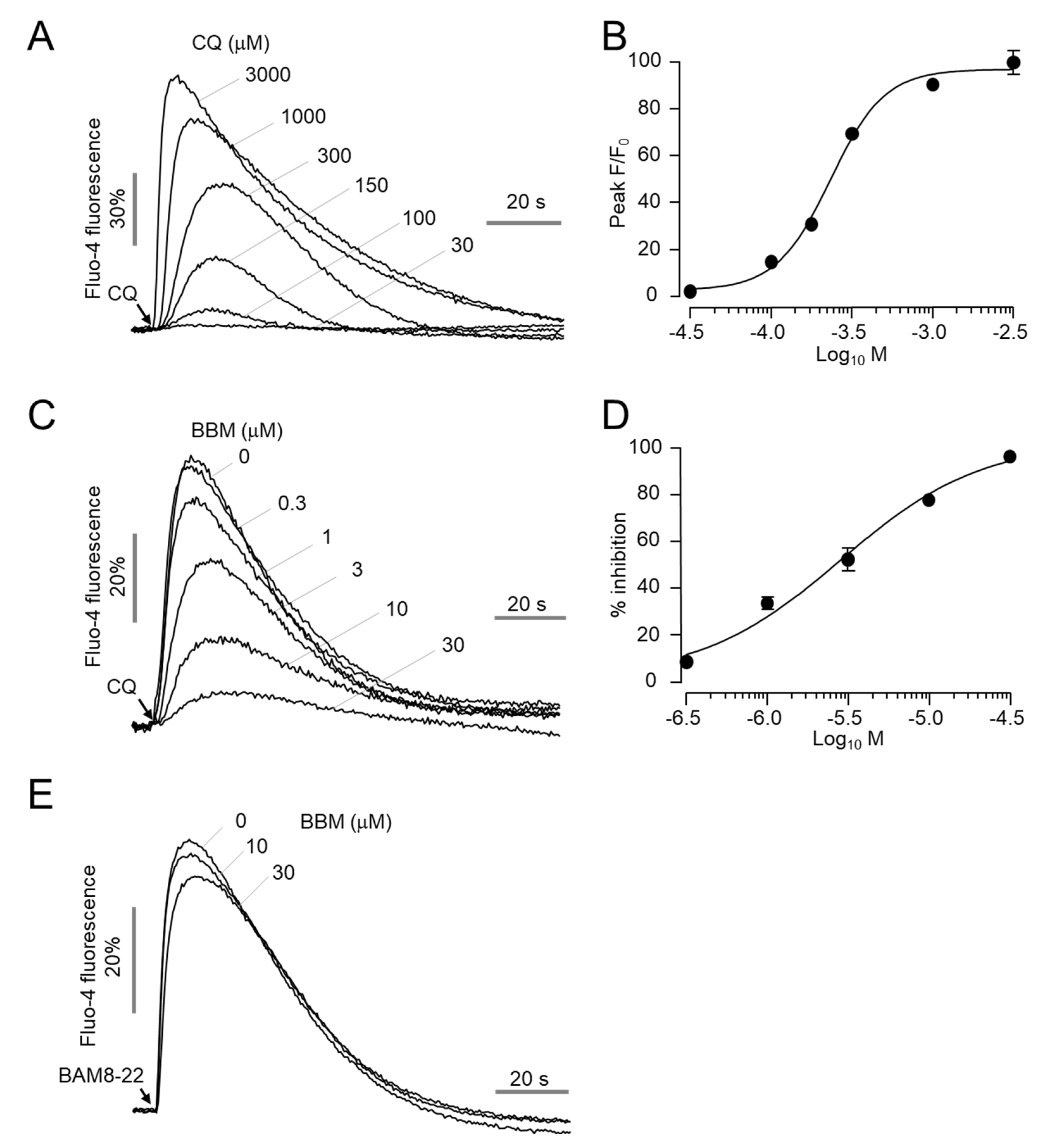
2.5. Berbamine Potently Inhibits MrgprA3-Mediated Calcium Signaling and ERK1/2 Phosphorylation
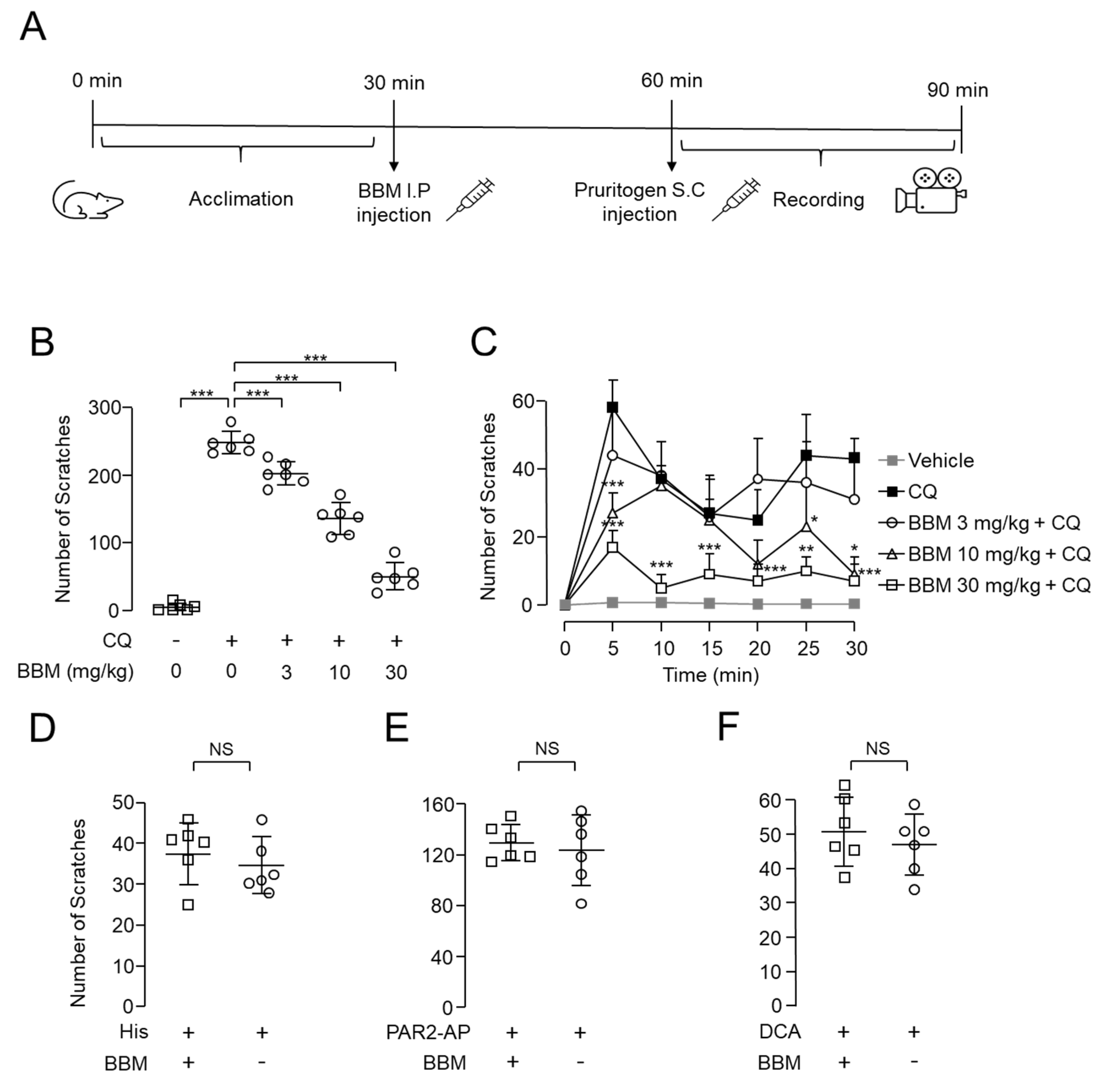
2.6. Antipruritic Effect of Berbamine on CQ-Induced Pruritus Mouse Model
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Intracellular Calcium Measurement
4.3. YFP Fluorescence Quenching Assay
4.4. Immunoblot Analysis
4.5. Acute Pruritogen-Induced Pruritus
4.6. Statistical Analysis
4.7. Study Approval
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CQ | Chloroquine |
| Mrgpr | Mas-related G Protein-coupled receptor |
| PAR | Protease-activated receptor |
| TGR5 | Takeda G protein-coupled receptor 5 |
| CLS | Transmembrane member 16A |
| BBM | Berbamine |
| CLS | Closantel |
| CFTR | Cystic fibrosis transmembrane conductance regulator |
| H1R | Histamine H1 receptor |
| H4R | Histamine H4 receptor |
| DRG | Dorsal root ganglion |
| DCA | Deoxycholic acid |
References
- World Health Organization. World Malaria Report 2021; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- Takala-Harrison, S.; Laufer, M.K. Antimalarial drug resistance in Africa: Key lessons for the future. Ann. N. Y. Acad. Sci. 2015, 1342, 62–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bussaratid, V.; Walsh, D.S.; Wilairatana, P.; Krudsood, S.; Silachamroon, U.; Looareesuwan, S. Frequency of Pruritus in Plasmodium Vivax Malaria Patients Treated with Chloroquine in Thailand. Trop. Dr. 2000, 30, 211–214. [Google Scholar] [CrossRef] [PubMed]
- Mnyika, K.S.; Kihamia, C.M. Chloroquine-induced pruritus: Its impact on chloroquine utilization in malaria control in Dar es Salaam. J. Trop. Med. Hyg. 1991, 94, 27–31. [Google Scholar] [PubMed]
- Abila, B.; Ezeamuzie, I.C.; Igbigbi, P.S.; Ambakederemo, A.W.; Asomugha, L. Effects of two antihistamines on chloroquine and histamine induced weal and flare in healthy African volunteers. Afr. J. Med. Med. Sci. 1994, 23, 139–142. [Google Scholar]
- Steinhoff, M.; Neisius, U.; Ikoma, A.; Fartasch, M.; Heyer, G.; Skov, P.S.; Luger, T.A.; Schmelz, M. Proteinase-activated receptor-2 mediates itch: A novel pathway for pruritus in human skin. J. Neurosci. 2003, 23, 6176–6180. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Tang, Z.; Surdenikova, L.; Kim, S.; Patel, K.N.; Kim, A.; Ru, F.; Guan, Y.; Weng, H.J.; Geng, Y.; et al. Sensory neuron-specific GPCR Mrgprs are itch receptors mediating chloroquine-induced pruritus. Cell 2009, 139, 1353–1365. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Sikand, P.; Ma, C.; Tang, Z.; Han, L.; Li, Z.; Sun, S.; LaMotte, R.H.; Dong, X. Mechanisms of itch evoked by β-alanine. J. Neurosci. 2012, 32, 14532–14537. [Google Scholar] [CrossRef] [Green Version]
- Meixiong, J.; Vasavda, C.; Snyder, S.H.; Dong, X. MRGPRX4 is a G protein-coupled receptor activated by bile acids that may contribute to cholestatic pruritus. Proc. Natl. Acad. Sci. USA 2019, 116, 10525–10530. [Google Scholar] [CrossRef] [Green Version]
- Alemi, F.; Kwon, E.; Poole, D.P.; Lieu, T.; Lyo, V.; Cattaruzza, F.; Cevikbas, F.; Steinhoff, M.; Nassini, R.; Materazzi, S.; et al. The TGR5 receptor mediates bile acid-induced itch and analgesia. J. Clin. Investig. 2013, 123, 1513–1530. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Han, S.-k.; Zylka, M.J.; Simon, M.I.; Anderson, D.J. A diverse family of GPCRs expressed in specific subsets of nociceptive sensory neurons. Cell 2001, 106, 619–632. [Google Scholar] [CrossRef] [Green Version]
- Zylka, M.J.; Dong, X.; Southwell, A.L.; Anderson, D.J. Atypical expansion in mice of the sensory neuron-specific Mrg G protein-coupled receptor family. Proc. Natl. Acad. Sci. USA 2003, 100, 10043–10048. [Google Scholar] [CrossRef] [PubMed]
- He, S.Q.; Li, Z.; Chu, Y.X.; Han, L.; Xu, Q.; Li, M.; Yang, F.; Liu, Q.; Tang, Z.; Wang, Y.; et al. MrgC agonism at central terminals of primary sensory neurons inhibits neuropathic pain. Pain 2014, 155, 534–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunapuli, P.; Lee, S.; Zheng, W.; Alberts, M.; Kornienko, O.; Mull, R.; Kreamer, A.; Hwang, J.I.; Simon, M.I.; Strulovici, B. Identification of small molecule antagonists of the human mas-related gene-X1 receptor. Anal. Biochem. 2006, 351, 50–61. [Google Scholar] [CrossRef]
- Choi, D.-S.; Ji, Y.; Jang, Y.; Lee, W.-J.; Shim, W.-S. Crotamiton, an anti-scabies agent, suppresses histamine-and chloroquine-induced itch pathways in sensory neurons and alleviates scratching in mice. Biomol. Ther. 2020, 28, 569. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.; Mun, C.H.; Park, S.-H.; Jeon, D.; Kim, S.J.; Yoon, T.; Ko, E.; Jo, S.; Park, Y.-B.; Namkung, W.; et al. Punicalagin Ameliorates Lupus Nephritis via Inhibition of PAR2. Int. J. Mol. Sci. 2020, 21, 4975. [Google Scholar] [CrossRef]
- Solinski, H.J.; Petermann, F.; Rothe, K.; Boekhoff, I.; Gudermann, T.; Breit, A. Human Mas-related G protein-coupled receptors-X1 induce chemokine receptor 2 expression in rat dorsal root ganglia neurons and release of chemokine ligand 2 from the human LAD-2 mast cell line. PLoS ONE 2013, 8, e58756. [Google Scholar] [CrossRef] [Green Version]
- Hanboonkunupakarn, B.; White, N.J. The threat of antimalarial drug resistance. Trop. Dis. Travel Med. Vaccines 2016, 2, 10. [Google Scholar] [CrossRef] [Green Version]
- Adam, M.E.; El Fatih, I.; Elkadaru, A.Y.; Ibrahim, K.E.; Berger, B.J.; Wiese, M.; Babiker, H.A. Imipramine induced complete reversal of chloroquine resistance in plasmodium falciparum infections in Sudan. Saudi Pharm. J. 2004, 12, 130–135. [Google Scholar]
- Evans, J.; May, J.; Tominski, D.; Eggelte, T.; Marks, F.; Abruquah, H.; Meyer, C.; Timmann, C.; Agbenyega, T.; Horstmann, R. Pre-treatment with chloroquine and parasite chloroquine resistance in Ghanaian children with severe malaria. QJM 2005, 98, 789–796. [Google Scholar] [CrossRef]
- Onyeji, C.O.; Ogunbona, F.A. Pharmacokinetic aspects of chloroquine-induced pruritus: Influence of dose and evidence for varied extent of metabolism of the drug. Eur. J. Pharm. Sci. 2001, 13, 195–201. [Google Scholar] [CrossRef]
- Dencker, L.; Lindquist, N.G.; Ullberg, S. Distribution of an 125I-labelled chloroquine analogue in a pregnant macaca monkey. Toxicology 1975, 5, 255–265. [Google Scholar] [CrossRef]
- Olatunde, I. Chloroquine concentrations in the skin of rabbits and man. Br. J. Pharmacol. 1971, 43, 335. [Google Scholar] [PubMed]
- Sams, W.; Epstein, J.H. The affinity of melanin for chloroquine. J. Investig. Dermatol. 1965, 45, 482–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, M.; Takashina, H.; Tsutsumi, S. Comparative assessment of ocular tissue distribution of drug-related radioactivity after chronic oral administration of 14C-levofloxacin and 14C-chloroquine in pigmented rats. J. Pharm. Pharmacol. 2004, 56, 977–983. [Google Scholar] [CrossRef] [PubMed]
- Carstens, E.; Akiyama, T. Central mechanisms of itch. Itch-Manag. Clin. Pract. 2016, 50, 11–17. [Google Scholar]
- Ji, R.-R.; Kohno, T.; Moore, K.A.; Woolf, C.J. Central sensitization and LTP: Do pain and memory share similar mechanisms? Trends Neurosci. 2003, 26, 696–705. [Google Scholar] [CrossRef]
- Ji, R.-R. Neuroimmune interactions in itch: Do chronic itch, chronic pain, and chronic cough share similar mechanisms? Pulm. Pharmacol. Ther. 2015, 35, 81–86. [Google Scholar] [CrossRef] [Green Version]
- Ständer, S.; Schmelz, M. Chronic itch and pain–similarities and differences. Eur. J. Pain 2006, 10, 473–478. [Google Scholar] [CrossRef]
- Jiang, G.-Y.; Dai, M.-H.; Huang, K.; Chai, G.-D.; Chen, J.-Y.; Chen, L.; Lang, B.; Wang, Q.-X.; St Clair, D.; McCaig, C. Neurochemical characterization of pERK-expressing spinal neurons in histamine-induced itch. Sci. Rep. 2015, 5, 12787. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Jiang, G.-Y.; Song, N.-J.; Huang, Y.; Chen, J.-Y.; Wang, Q.-X.; Ding, Y.-Q. Extracellular signal-regulated kinase (ERK) activation is required for itch sensation in the spinal cord. Mol. Brain 2014, 7, 25. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Wang, C.; Hu, D.; Zhang, X.; Shen, R.; Zhou, Y.; Yang, Y.; Zhu, C.; Tang, Z.; Yu, G. mMrgprA3/mMrgprC11/hMrgprX1: Potential therapeutic targets for allergic contact dermatitis–induced pruritus in mice and humans. Contact Dermat. 2022, 86, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Zhang, C.; Hu, P.; Zhu, M.; Hu, M.; Gao, S. An UPLC-MS/MS method for quantifying tetrandrine and its metabolite berbamine in human blood: Application to a human pharmacokinetic study. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2017, 1070, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Van Dyke, K.; Yang, B. Interaction of berbamine and chloroquine or artemisinin against chloroquine-sensitive and-resistant plasmodium falciparum in vitro. Drug Dev. Res. 1993, 30, 229–237. [Google Scholar] [CrossRef]
- Lee, H.K.; Park, J.; Kim, B.R.; Jun, I.; Kim, T.I.; Namkung, W. Isorhamnetin Ameliorates Dry Eye Disease via CFTR Activation in Mice. Int. J. Mol. Sci. 2021, 22, 3954. [Google Scholar] [CrossRef]
- Heo, Y.; Jeon, H.; Namkung, W. PAR4-Mediated PI3K/Akt and RhoA/ROCK Signaling Pathways Are Essential for Thrombin-Induced Morphological Changes in MEG-01 Cells. Int. J. Mol. Sci. 2022, 23, 776. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ryu, K.; Heo, Y.; Lee, Y.; Jeon, H.; Namkung, W. Berbamine Reduces Chloroquine-Induced Itch in Mice through Inhibition of MrgprX1. Int. J. Mol. Sci. 2022, 23, 14321. https://doi.org/10.3390/ijms232214321
Ryu K, Heo Y, Lee Y, Jeon H, Namkung W. Berbamine Reduces Chloroquine-Induced Itch in Mice through Inhibition of MrgprX1. International Journal of Molecular Sciences. 2022; 23(22):14321. https://doi.org/10.3390/ijms232214321
Chicago/Turabian StyleRyu, Kunhi, Yunkyung Heo, Yechan Lee, Hyejin Jeon, and Wan Namkung. 2022. "Berbamine Reduces Chloroquine-Induced Itch in Mice through Inhibition of MrgprX1" International Journal of Molecular Sciences 23, no. 22: 14321. https://doi.org/10.3390/ijms232214321
APA StyleRyu, K., Heo, Y., Lee, Y., Jeon, H., & Namkung, W. (2022). Berbamine Reduces Chloroquine-Induced Itch in Mice through Inhibition of MrgprX1. International Journal of Molecular Sciences, 23(22), 14321. https://doi.org/10.3390/ijms232214321