

Conjunctival Fluid Secretion Impairment via CaCC-CFTR Dysfunction Is the Key Mechanism in Environmental Dry Eye
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
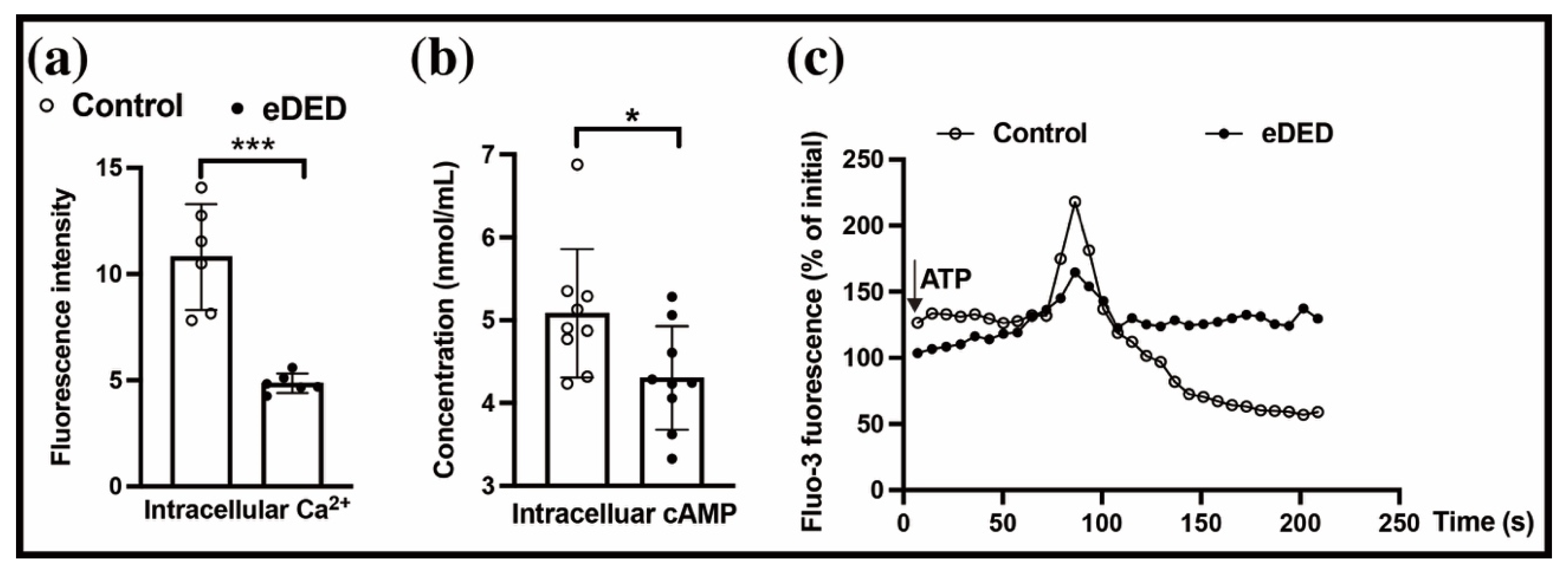
2.1. eDED Has Ocular Surface Injury and Conjunctival Fluid Secretion Dysfunction
2.2. P2Y2R Plays a Key Role in eDED
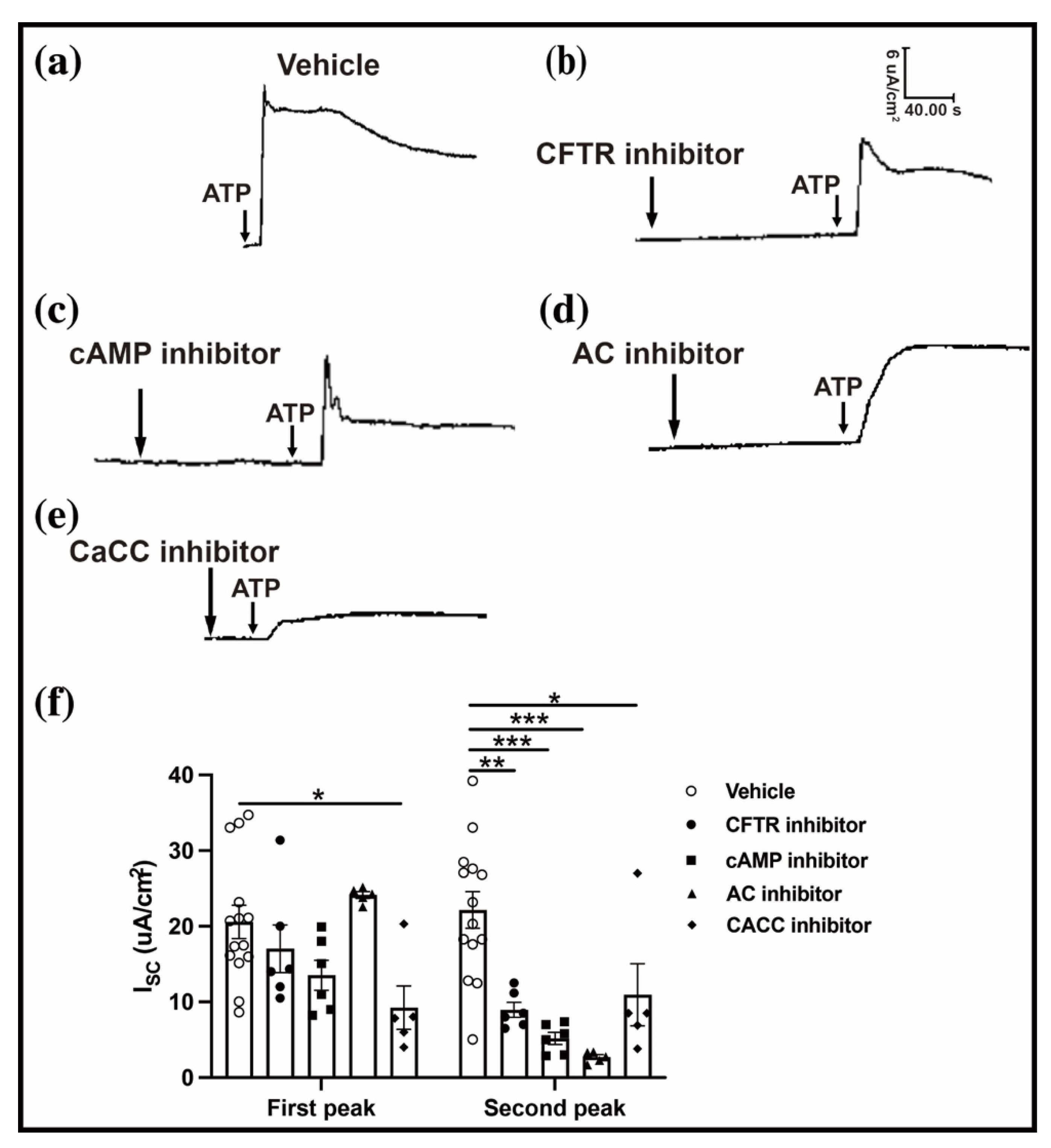
2.3. ATP-Induced Conjunctival Fluid Secretion Is Related to the Anion Channel
2.4. CaCC-CFTR Pathway Participates in the ATP-Induced Conjunctival Fluid Secretion
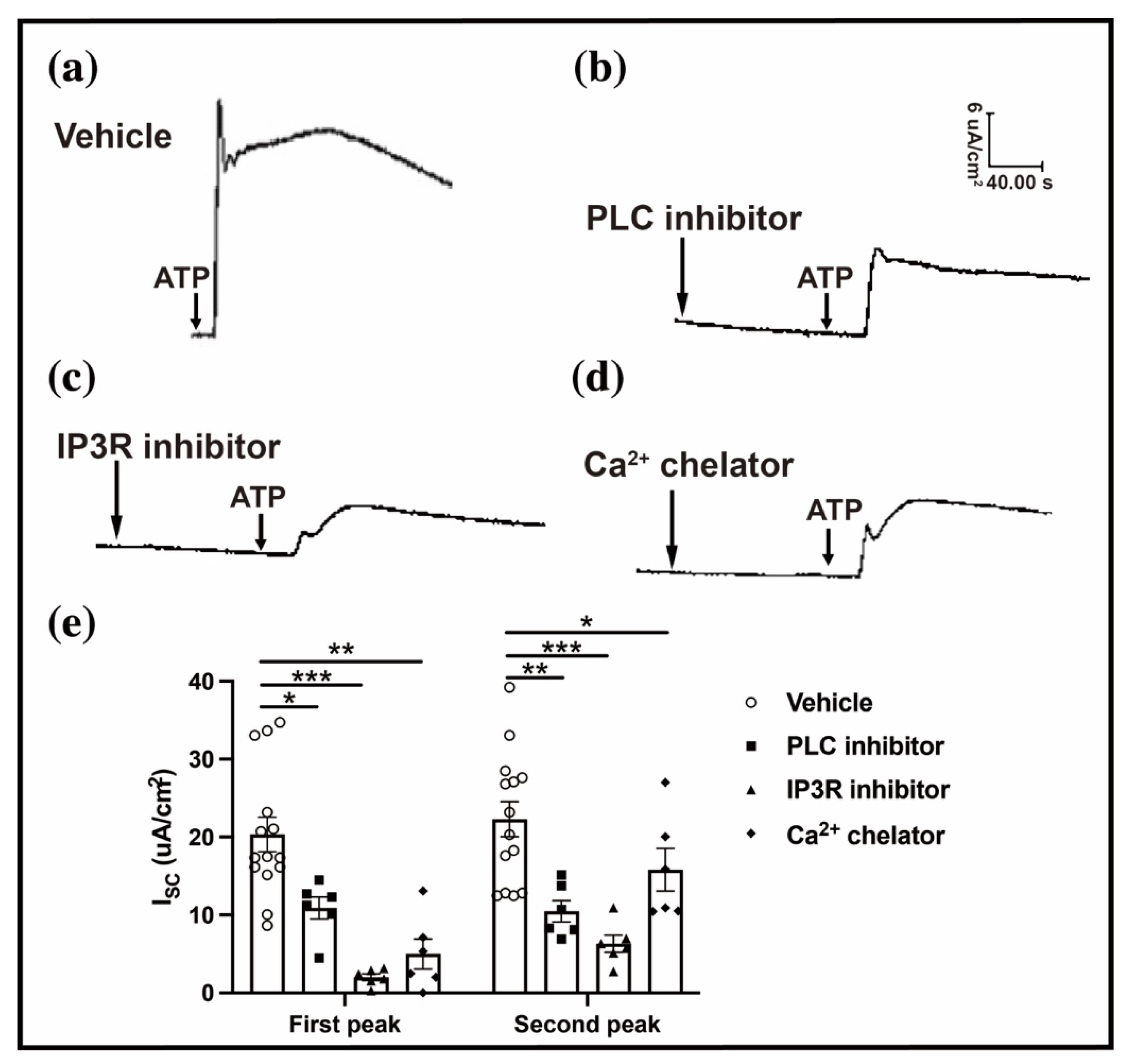
2.5. Intracellular Ca2+ Regulates the ATP-Induced Conjunctival Fluid Secretion
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Environmental Dry Eye Disease Model
4.3. Intervention of Rabbit Eyes with P2Y2R siRNA
4.4. Short-Circuit Current (Isc) Measurements
4.5. Measurement of Intracellular cAMP
4.6. Measurement of Intracellular Ca2+
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stapleton, F.; Alves, M.; Bunya, V.Y.; Jalbert, I.; Lekhanont, K.; Malet, F.; Na, K.S.; Schaumberg, D.; Uchino, M.; Vehof, J.; et al. TFOS DEWS II Epidemiology Report. Ocul. Surf. 2017, 15, 334–365. [Google Scholar] [CrossRef] [PubMed]
- Bron, A.J.; de Paiva, C.S.; Chauhan, S.K.; Bonini, S.; Gabison, E.E.; Jain, S.; Knop, E.; Markoulli, M.; Ogawa, Y.; Perez, V.; et al. TFOS DEWS II pathophysiology report. Ocul Surf. 2017, 15, 438–510. [Google Scholar] [CrossRef]
- Wolffsohn, J.S.; Arita, R.; Chalmers, R.; Djalilian, A.; Dogru, M.; Dumbleton, K.; Gupta, P.K.; Karpecki, P.; Lazreg, S.; Pult, H.; et al. TFOS DEWS II Diagnostic Methodology report. Ocul. Surf. 2017, 15, 539–574. [Google Scholar] [CrossRef]
- Uchino, M.; Schaumberg, D.A.; Dogru, M.; Uchino, Y.; Fukagawa, K.; Shimmura, S.; Satoh, T.; Takebayashi, T.; Tsubota, K. Prevalence of dry eye disease among Japanese visual display terminal users. Ophthalmology 2008, 115, 1982–1988. [Google Scholar] [CrossRef] [PubMed]
- Abusharha, A.A.; Pearce, E.I. The effect of low humidity on the human tear film. Cornea 2013, 32, 429–434. [Google Scholar] [CrossRef] [PubMed]
- LI, Y.; Kuang, K.; Yerxa, B.; Wen, Q.; Rosskothen, H.; Fischbarg, J. Rabbit conjunctival epithelium transports fluid, and P2Y22 receptor agonists stimulate Cl2 and fluid secretion. Am. J. Physiol. Cell Physiol. 2001, 281, C595–C602. [Google Scholar] [CrossRef] [Green Version]
- Nandoskar, P.; Wang, Y.; Wei, R.; Liu, Y.; Zhao, P.; Lu, M.; Huang, J. Changes of chloride channels in the lacrimal glands of a rabbit model of Sjgren syndrome. Cornea 2011, 31, 7. [Google Scholar]
- Lee, H.K.; Park, J.; Kim, B.R.; Jun, I.; Kim, T.I.; Namkung, W. Isorhamnetin Ameliorates Dry Eye Disease via CFTR Activation in Mice. Int J Mol Sci 2021, 22, 3954. [Google Scholar] [CrossRef]
- Flores, A.M.; Casey, S.D.; Felix, C.M.; Phuan, P.W.; Verkman, A.S.; Levin, M.H. Small-molecule CFTR activators increase tear secretion and prevent experimental dry eye disease. FASEB J. 2016, 30, 1789–1797. [Google Scholar]
- Botelho, S.Y.; Goldstein, A.M.; Rosenlund, M.L. Tear sodium, potassium, chloride, and calcium at various flow rates: Children with cystic fibrosis and unaffected siblings with and without corneal staining. J Pediatr. 1973, 83, 601–606. [Google Scholar] [CrossRef]
- Murakami, T.; Fujihara, T.; Nakamura, M.; Nakata, K. P2Y(2) receptor stimulation increases tear fluid secretion in rabbits. Curr. Eye Res. 2000, 21, 782–787. [Google Scholar] [CrossRef]
- Dominguez-Godinez, C.; Carracedo, G.; Pintor, J. Diquafosol Delivery from Silicone Hydrogel Contact Lenses: Improved Effect on Tear Secretion. J. Ocul. Pharmacol. Ther. 2018, 34, 170–176. [Google Scholar] [CrossRef]
- Son, M.; Ito, Y.; Sato, S.; Ishikawa, T.; Kondo, M.; Nakayama, S.; Shimokata, K.; Kume, H. Apical and basolateral ATP-induced anion secretion in polarized human airway epithelia. Am. J. Respir. Cell Mol. Biol. 2004, 30, 411–419. [Google Scholar] [CrossRef]
- Centeio, R.; Ousingsawat, J.; Schreiber, R.; Kunzelmann, K. CLCA1 Regulates Airway Mucus Production and Ion Secretion Through TMEM16A. Int. J. Mol. Sci. 2021, 22, 5133. [Google Scholar] [CrossRef] [PubMed]
- Shiue, M.H.; Kim, K.J.; Lee, V.H. Modulation of chloride secretion across the pigmented rabbit conjunctiva. Exp. Eye Res. 1998, 66, 275–282. [Google Scholar] [CrossRef]
- Yu, D.; Thelin, W.R.; Rogers, T.D.; Stutts, M.J.; Randell, S.H.; Grubb, B.R.; Boucher, R.C. Regional differences in rat conjunctival ion transport activities. Am. J. Physiol. Cell Physiol. 2012, 303, C767–C780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faria, D.; Schreiber, R.; Kunzelmann, K. CFTR is activated through stimulation of purinergic P2Y2 receptors. Pflugers Arch. 2009, 457, 1373–1380. [Google Scholar] [CrossRef] [PubMed]
- Rajagopal, M.; Kathpalia, P.P.; Thomas, S.V.; Pao, A.C. Activation of P2Y1 and P2Y2 receptors induces chloride secretion via calcium-activated chloride channels in kidney inner medullary collecting duct cells. Am. J. Physiol. Renal. Physiol. 2011, 301, F544–F553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutta, A.K.; Woo, K.; Doctor, R.B.; Fitz, J.G.; Feranchak, A.P. Extracellular nucleotides stimulate Cl- currents in biliary epithelia through receptor-mediated IP3 and Ca2+ release. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295, G1004–G1015. [Google Scholar] [CrossRef] [Green Version]
- Mikolajewicz, N.; Smith, D.; Komarova, S.V.; Khadra, A. High-affinity P2Y2 and low-affinity P2X7 receptor interaction modulates ATP-mediated calcium signaling in murine osteoblasts. PLoS Comput. Biol. 2021, 17, e1008872. [Google Scholar] [CrossRef]
- Peng, L.; Gao, D.D.; Xu, J.W.; Xu, J.B.; Ke, L.J.; Qiu, Z.E.; Zhu, Y.X.; Zhang, Y.L.; Zhou, W.L. Cellular mechanisms underlying carbon monoxide stimulated anion secretion in rat epididymal epithelium. Nitric Oxide 2020, 100, 30–37. [Google Scholar] [CrossRef]
- Szabadkai, G.; Bianchi, K.; Varnai, P.; De Stefani, D.; Wieckowski, M.R.; Cavagna, D.; Nagy, A.I.; Balla, T.; Rizzuto, R. Chaperone-mediated coupling of endoplasmic reticulum and mitochondrial Ca2+ channels. J. Cell Biol. 2006, 175, 901–911. [Google Scholar] [CrossRef]
- Fischbarg, J. Water channels and their roles in some ocular tissues. Mol. Aspects Med. 2012, 33, 638–641. [Google Scholar] [CrossRef]
- Kulkarni, A.A.; Trousdale, M.D.; Stevenson, D.; Gukasyan, H.J.; Shiue, M.H.; Kim, K.J.; Read, R.W.; Lee, V.H. Nucleotide-induced restoration of conjunctival chloride and fluid secretion in adenovirus type 5-infected pigmented rabbit eyes. J. Pharmacol. Exp. Ther. 2003, 305, 1206–1211. [Google Scholar] [CrossRef] [Green Version]
- Kompella, U.B.; Kim, K.J.; Lee, V.H. Active chloride transport in the pigmented rabbit conjunctiva. Curr. Eye Res. 1993, 12, 1041–1048. [Google Scholar] [CrossRef]
- Shi, X.P.; Candia, O.A. Active sodium and chloride transport across the isolated rabbit conjunctiva. Curr. Eye Res. 1995, 14, 927–935. [Google Scholar] [CrossRef]
- Rajagopal, M.; Kathpalia, P.P.; Widdicombe, J.H.; Pao, A.C. Differential effects of extracellular ATP on chloride transport in cortical collecting duct cells. Am. J. Physiol. Renal. Physiol. 2012, 303, F483–F491. [Google Scholar] [CrossRef] [Green Version]
- Du, J.Y.; Zuo, W.L.; Ruan, Y.C.; Yang, Z.H.; Chen, M.H.; Chen, S.L.; Li, S.; Wu, Z.L.; Xiang, H.; Zhou, W.L. Stimulating effects of dopamine on chloride transport across the rat caudal epididymal epithelium in culture. Biol. Reprod. 2009, 80, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.M.; Park, S.; Ji, H.; Kim, T.I.; Kim, E.K.; Kang, K.K.; Shin, D.M. DA-6034 Induces [Ca(2+)]i Increase in Epithelial Cells. Korean J. Physiol. Pharmacol. 2014, 18, 89–94. [Google Scholar] [CrossRef] [Green Version]
- Guzman-Aranguez, A.; Perez de Lara, M.J.; Pintor, J. Hyperosmotic stress induces ATP release and changes in P2X7 receptor levels in human corneal and conjunctival epithelial cells. Purinergic Signal. 2017, 13, 249–258. [Google Scholar] [CrossRef] [Green Version]
- Pedemonte, N.; Galietta, L.J. Structure and function of TMEM16 proteins (anoctamins). Physiol. Rev. 2014, 94, 419–459. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.M.; Gao, J.M.; Luo, Y.L.; Wen, Y.Z.; Zhang, Y.L.; Hide, G.; Zhou, W.L.; Ayala, F.J.; Lun, Z.R. Infection by Toxoplasma gondii, a severe parasite in neonates and AIDS patients, causes impaired anion secretion in airway epithelia. Proc. Natl. Acad. Sci. USA 2015, 112, 4435–4440. [Google Scholar] [CrossRef]
- Khajavi, N.; Reinach, P.S.; Slavi, N.; Skrzypski, M.; Lucius, A.; Strauss, O.; Kohrle, J.; Mergler, S. Thyronamine induces TRPM8 channel activation in human conjunctival epithelial cells. Cell Signal 2015, 27, 315–325. [Google Scholar] [CrossRef]
- Marchi, S.; Patergnani, S.; Missiroli, S.; Morciano, G.; Rimessi, A.; Wieckowski, M.R.; Giorgi, C.; Pinton, P. Mitochondrial and endoplasmic reticulum calcium homeostasis and cell death. Cell Calcium 2018, 69, 62–72. [Google Scholar] [CrossRef]
- Brookes, P.S.; Yoon, Y.; Robotham, J.L.; Anders, M.W.; Sheu, S.S. Calcium, ATP, and ROS: A mitochondrial love-hate triangle. Am. J. Physiol. Cell Physiol. 2004, 287, C817–C833. [Google Scholar] [CrossRef]
- Chen, X.M.; Kuang, J.B.; Yu, H.Y.; Wu, Z.N.; Wang, S.Y.; Zhou, S.Y. A Novel Rabbit Dry Eye Model Induced by a Controlled Drying System. Transl. Vis. Sci. Technol. 2021, 10, 32. [Google Scholar] [CrossRef]
- Butovich, I.A.; Yuksel, S.; Leonard, B.; Gadek, T.; Polans, A.S.; Albert, D.M. Novel Lipids of the Rabbit Harderian Gland Improve Tear Stability in an Animal Model of Dry Eye Disease. J. Ocul. Pharmacol. Ther. 2021, 37, 545–555. [Google Scholar] [CrossRef]
- Crooke, A.; Mediero, A.; Guzman-Aranguez, A.; Pintor, J. Silencing of P2Y2 receptor delays Ap4A-corneal re-epithelialization process. Mol. Vis. 2009, 15, 1169–1178. [Google Scholar]
- Martin-Gil, A.; de Lara, M.J.; Crooke, A.; Santano, C.; Peral, A.; Pintor, J. Silencing of P2Y(2) receptors reduces intraocular pressure in New Zealand rabbits. Br. J. Pharmacol. 2012, 165, 1163–1172. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.W.; Gao, D.D.; Peng, L.; Qiu, Z.E.; Ke, L.J.; Zhu, Y.X.; Zhang, Y.L.; Zhou, W.L. The gasotransmitter hydrogen sulfide inhibits transepithelial anion secretion of pregnant mouse endometrial epithelium. Nitric Oxide 2019, 90, 37–46. [Google Scholar] [CrossRef]
- Shen, Z.Y.; Shen, W.Y.; Chen, M.H.; Shen, J.; Cai, W.J.; Yi, Z. Nitric oxide and calcium ions in apoptotic esophageal carcinoma cells induced by arsenite. World J. Gastroenterol. 2002, 8, 40–43. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Lin, L.; Chen, X.; Wang, S.; Wei, Y.; Zhou, W.; Yang, S.; Zhou, S. Conjunctival Fluid Secretion Impairment via CaCC-CFTR Dysfunction Is the Key Mechanism in Environmental Dry Eye. Int. J. Mol. Sci. 2022, 23, 14399. https://doi.org/10.3390/ijms232214399
Zhang J, Lin L, Chen X, Wang S, Wei Y, Zhou W, Yang S, Zhou S. Conjunctival Fluid Secretion Impairment via CaCC-CFTR Dysfunction Is the Key Mechanism in Environmental Dry Eye. International Journal of Molecular Sciences. 2022; 23(22):14399. https://doi.org/10.3390/ijms232214399
Chicago/Turabian StyleZhang, Jinyu, Limian Lin, Xiaomin Chen, Shuyi Wang, Yuan Wei, Wenliang Zhou, Shuangjian Yang, and Shiyou Zhou. 2022. "Conjunctival Fluid Secretion Impairment via CaCC-CFTR Dysfunction Is the Key Mechanism in Environmental Dry Eye" International Journal of Molecular Sciences 23, no. 22: 14399. https://doi.org/10.3390/ijms232214399
APA StyleZhang, J., Lin, L., Chen, X., Wang, S., Wei, Y., Zhou, W., Yang, S., & Zhou, S. (2022). Conjunctival Fluid Secretion Impairment via CaCC-CFTR Dysfunction Is the Key Mechanism in Environmental Dry Eye. International Journal of Molecular Sciences, 23(22), 14399. https://doi.org/10.3390/ijms232214399