

Derivatives of Trimethoxybenzoic Acid and Gallic Acid as Potential Efflux Pump Inhibitors: In Silico and In Vitro Studies
 , , , , ,
, , , , ,  ,
,  and
and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Chemistry
2.2. Molecular Docking
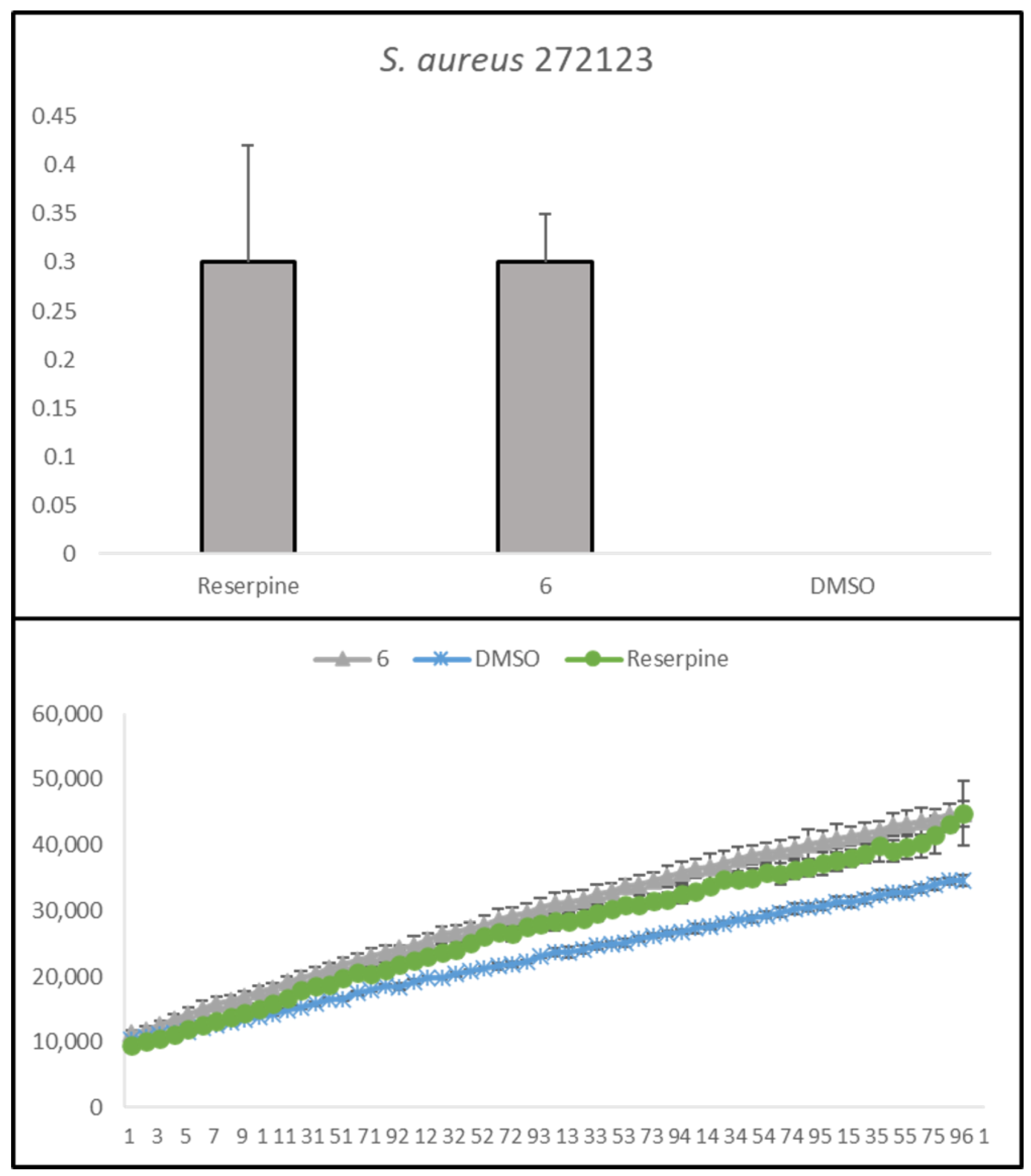
2.3. Antibacterial Activity and Antibiotic Potentiation
2.4. Efflux Pump Inhibition
2.5. Inhibition of Biofilm Formation and Quorum-Sensing Assays
2.6. Antifungal Activity
2.7. In Silico ADME Properties
3. Materials and Methods
3.1. Chemistry
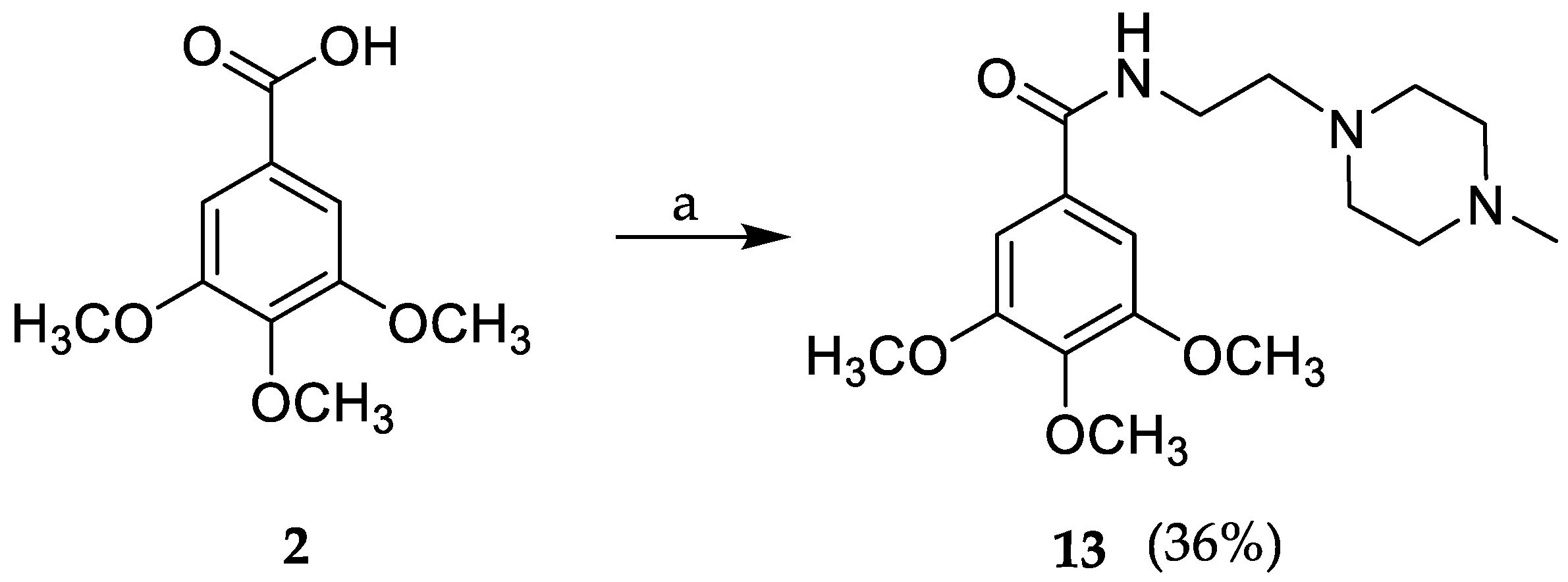
3.1.1. General Procedure for the Synthesis of Compounds 3–13
3,4,5-Trimethoxy-N-(2-(4-methylpiperazin-1-yl)ethyl)benzamide (13)
3.2. Molecular Docking
3.3. Culture Media and Chemicals
3.4. Microorganisms
3.5. Antibacterial and Antibiotic Potentiation Assays
3.6. Efflux Pump Inhibition
3.7. Inhibition of Biofilm Formation
3.8. Quorum-Sensing Inhibition
3.9. Antifungal Assays
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gaynes, R.P. The Discovery of Penicillin—New Insights After More Than 75 Years of Clinical Use. Emerg. Infect. Dis. 2017, 23, 849–853. [Google Scholar] [CrossRef]
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Andrei, S.; Droc, G.; Stefan, G. FDA approved antibacterial drugs: 2018-2019. Discoveries 2019, 7, e102. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, P.S. Antibacterial Discovery: 21st Century Challenges. Antibiotics 2020, 9, 213. [Google Scholar] [CrossRef] [PubMed]
- Donadio, S.; Maffioli, S.; Monciardini, P.; Sosio, M.; Jabes, D. Antibiotic discovery in the twenty-first century: Current trends and future perspectives. J. Antibiot. 2010, 63, 423–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durães, F.; Pinto, M.; Sousa, E. Medicinal Chemistry Updates on Bacterial Efflux Pump Modulators. Curr. Med. Chem. 2018, 25, 6030–6069. [Google Scholar] [CrossRef]
- Neves, A.R.; Boas, C.V.; Gonçalves, C.; Vasconcelos, V.; Pinto, M.; Silva, E.R.; Sousa, E.; Almeida, J.R.; Correia-da-Silva, M. Gallic acid derivatives as inhibitors of mussel (Mytilus galloprovincialis) larval settlement: Lead optimization, biological evaluation and use in antifouling coatings. Bioorganic Chem. 2022, 126, 105911. [Google Scholar] [CrossRef]
- Neves, A.R.; Gomes, L.C.; Faria, S.I.; Sousa, J.; Ruivo, R.; Páscoa, I.; Pinto, M.; Sousa, E.; Santos, M.M.; Silva, E.R.; et al. Antifouling Marine Coatings with a Potentially Safer and Sustainable Synthetic Polyphenolic Derivative. Mar. Drugs 2022, 20, 507. [Google Scholar] [CrossRef]
- Perron, Y.G. Therapeutic Agents. US2870145A, 20 January 1959. [Google Scholar]
- Zimmermann, S.; Klinger-Strobel, M.; Bohnert, J.A.; Wendler, S.; Rödel, J.; Pletz, M.W.; Löffler, B.; Tuchscherr, L. Clinically Approved Drugs Inhibit the Staphylococcus aureus Multidrug NorA Efflux Pump and Reduce Biofilm Formation. Front. Microbiol. 2019, 10, 2762. [Google Scholar] [CrossRef] [Green Version]
- Shaheen, A.; Afridi, W.A.; Mahboob, S.; Sana, M.; Zeeshan, N.; Ismat, F.; Mirza, O.; Iqbal, M.; Rahman, M. Reserpine Is the New Addition into the Repertoire of AcrB Efflux Pump Inhibitors. Mol. Biol. 2019, 53, 596–605. [Google Scholar] [CrossRef]
- Shamon, S.D.; Perez, M.I. Blood pressure-lowering efficacy of reserpine for primary hypertension. Cochrane Database Syst. Rev. 2016, 12, Cd007655. [Google Scholar] [CrossRef] [PubMed]
- Hoenders, H.J.R.; Bartels-Velthuis, A.A.; Vollbehr, N.K.; Bruggeman, R.; Knegtering, H.; de Jong, J. Natural Medicines for Psychotic Disorders: A Systematic Review. J. Nerv. Ment. Dis. 2018, 206, 81–101. [Google Scholar] [CrossRef] [Green Version]
- Marcus, S.M.; Borzelleca, J.F. Observations on reserpine-induced bradycardia. Arch. Int. Pharmacodyn. Ther. 1968, 174, 12–16. [Google Scholar] [PubMed]
- Mladěnka, P.; Applová, L.; Patočka, J.; Costa, V.M.; Remiao, F.; Pourová, J.; Mladěnka, A.; Karlíčková, J.; Jahodář, L.; Vopršalová, M.; et al. Comprehensive review of cardiovascular toxicity of drugs and related agents. Med. Res. Rev. 2018, 38, 1332–1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, X.; Chen, M.; Yu, Z.; Bell, J.M.; Wang, H.; Forrester, I.; Villarreal, H.; Jakana, J.; Du, D.; Luisi, B.F.; et al. In situ structure and assembly of the multidrug efflux pump AcrAB-TolC. Nat. Commun. 2019, 10, 2635. [Google Scholar] [CrossRef] [Green Version]
- Yan, N. Structural advances for the major facilitator superfamily (MFS) transporters. Trends Biochem. Sci. 2013, 38, 151–159. [Google Scholar] [CrossRef]
- Thai, K.-M.; Ngo, T.-D.; Phan, T.-V.; Tran, T.-D.; Nguyen, N.-V.; Nguyen, T.-H.; Le, M.-T. Virtual Screening for Novel Staphylococcus Aureus NorA Efflux Pump Inhibitors From Natural Products. Med. Chem. 2015, 11, 135–155. [Google Scholar] [CrossRef]
- Yin, Y.; He, X.; Szewczyk, P.; Nguyen, T.; Chang, G. Structure of the Multidrug Transporter EmrD from Escherichia coli. Science 2006, 312, 741–744. [Google Scholar] [CrossRef]
- Alav, I.; Sutton, J.M.; Rahman, K.M. Role of bacterial efflux pumps in biofilm formation. J. Antimicrob. Chemother. 2018, 73, 2003–2020. [Google Scholar] [CrossRef] [Green Version]
- Costa, S.S.; Sobkowiak, B.; Parreira, R.; Edgeworth, J.D.; Viveiros, M.; Clark, T.G.; Couto, I. Genetic Diversity of norA, Coding for a Main Efflux Pump of Staphylococcus aureus. Front. Genet. 2019, 9. [Google Scholar] [CrossRef]
- Bruce, S.A.; Smith, J.T.; Mydosh, J.L.; Ball, J.; Needle, D.B.; Gibson, R.; Andam, C.P. Shared antibiotic resistance and virulence genes in Staphylococcus aureus from diverse animal hosts. Sci. Rep. 2022, 12, 4413. [Google Scholar] [CrossRef] [PubMed]
- Durães, F.; Resende, D.I.S.P.; Palmeira, A.; Szemerédi, N.; Pinto, M.M.M.; Spengler, G.; Sousa, E. Xanthones Active against Multidrug Resistance and Virulence Mechanisms of Bacteria. Antibiotics 2021, 10, 600. [Google Scholar] [CrossRef] [PubMed]
- Parai, D.; Banerjee, M.; Dey, P.; Mukherjee, S.K. Reserpine attenuates biofilm formation and virulence of Staphylococcus aureus. Microb. Pathog. 2020, 138, 103790. [Google Scholar] [CrossRef] [PubMed]
- Benomar, S.; Evans, K.C.; Unckless, R.L.; Chandler, J.R. Efflux Pumps in Chromobacterium Species Increase Antibiotic Resistance and Promote Survival in a Coculture Competition Model. Appl. Environ. Microbiol. 2019, 85, e00908–e00919. [Google Scholar] [CrossRef] [Green Version]
- Gajdács, M.; Spengler, G. The Role of Drug Repurposing in the Development of Novel Antimicrobial Drugs: Non-Antibiotic Pharmacological Agents as Quorum Sensing-Inhibitors. Antibiotics 2019, 8, 270. [Google Scholar] [CrossRef] [Green Version]
- Mardanova, A.M.; Bogomol’naya, L.M.; Romanova, Y.D.; Sharipova, M.R. Efflux systems in Serratia marcescens. Microbiology 2013, 82, 668–679. [Google Scholar] [CrossRef]
- Wang, S.; Dong, G.; Sheng, C. Structural simplification: An efficient strategy in lead optimization. Acta Pharm. Sin. B 2019, 9, 880–901. [Google Scholar] [CrossRef]
- Murakami, S.; Nakashima, R.; Yamashita, E.; Matsumoto, T.; Yamaguchi, A. Crystal structures of a multidrug transporter reveal a functionally rotating mechanism. Nature 2006, 443, 173–179. [Google Scholar] [CrossRef]
- Mikolosko, J.; Bobyk, K.; Zgurskaya, H.I.; Ghosh, P. Conformational flexibility in the multidrug efflux system protein AcrA. Structure 2006, 14, 577–587. [Google Scholar] [CrossRef] [Green Version]
- Koronakis, V.; Sharff, A.; Koronakis, E.; Luisi, B.; Hughes, C. Crystal structure of the bacterial membrane protein TolC central to multidrug efflux and protein export. Nature 2000, 405, 914–919. [Google Scholar] [CrossRef]
- Sussman, J.L.; Lin, D.; Jiang, J.; Manning, N.O.; Prilusky, J.; Ritter, O.; Abola, E.E. Protein Data Bank (PDB): Database of Three-Dimensional Structural Information of Biological Macromolecules. Acta Crystallogr. Sect. D 1998, 54, 1078–1084. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [Green Version]
- Apweiler, R.; Bairoch, A.; Wu, C.H.; Barker, W.C.; Boeckmann, B.; Ferro, S.; Gasteiger, E.; Huang, H.; Lopez, R.; Magrane, M.; et al. UniProt: The universal protein knowledgebase. Nucleic Acids Res. 2017, 45, D158–D169. [Google Scholar] [CrossRef] [PubMed]
- Zárate, S.G.; Morales, P.; Świderek, K.; Bolanos-Garcia, V.M.; Bastida, A. A Molecular Modeling Approach to Identify Novel Inhibitors of the Major Facilitator Superfamily of Efflux Pump Transporters. Antibiotics 2019, 8, 25. [Google Scholar] [CrossRef] [Green Version]
- Bessa, L.J.; Barbosa-Vasconcelos, A.; Mendes, A.; Vaz-Pires, P.; Martins da Costa, P. High prevalence of multidrug-resistant Escherichia coli and Enterococcus spp. in river water, upstream and downstream of a wastewater treatment plant. J. Water Health 2014, 12, 426–435. [Google Scholar] [CrossRef] [PubMed]
- Gajdács, M.; Spengler, G. Standard operating procedure (SOP) for disk diffusion-based quorum sensing inhibition assays. Acta Pharm. Hung. 2020, 89, 117–125. [Google Scholar] [CrossRef] [Green Version]
- CLSI. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically, 11th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Spengler, G.; Takács, D.; Horváth, A.; Szabó, A.M.; Riedl, Z.; Hajós, G.; Molnár, J.; Burián, K. Efflux pump inhibiting properties of racemic phenothiazine derivatives and their enantiomers on the bacterial AcrAB-TolC system. In Vivo 2014, 28, 1071–1075. [Google Scholar]
- Peeters, E.; Nelis, H.J.; Coenye, T. Comparison of multiple methods for quantification of microbial biofilms grown in microtiter plates. J. Microbiol. Methods 2008, 72, 157–165. [Google Scholar] [CrossRef]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts; Approved standard—CLSI document M27-A3; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2008. [Google Scholar]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi; Approved Standard—Second Edition. CLSI document M38-A2; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2008. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Docking Score | ||||
|---|---|---|---|---|---|
| AcrB | AcrA | TolC | NorA | ||
| HH | LD | ||||
| 1 | −5.9 | −5.1 | −4.5 | −5.5 | −4.1 |
| 2 | −5.6 | −4.3 | −3.7 | −5.5 | −4.0 |
| 3 | −6.5 | −4.9 | −4.3 | −6.0 | −4.5 |
| 4 | −6.2 | −5.5 | −4.6 | −5.9 | −4.2 |
| 5 | −7.3 | −5.7 | −4.7 | −6.4 | −7.3 |
| 6 | −7.6 | −6.4 | −5.4 | −6.7 | −6.2 |
| 7 | −6.8 | −5.0 | −6.0 | −5.7 | −4.4 |
| 8 | −6.7 | −5.0 | −5.8 | −5.9 | −4.7 |
| 9 | −8.9 | −5.8 | −6.7 | −6.5 | −5.9 |
| 10 | −9.8 | −6.5 | −7.1 | −7.3 | −6.2 |
| 11 | −6.6 | −4.7 | −5.3 | −5.7 | −3.9 |
| 12 | −7.0 | −5.0 | −5.0 | −5.9 | −4.8 |
| 13 | −7.1 | −5.0 | −5.6 | −5.9 | −4.8 |
| Reserpine | −8.1 | 5.6 | 4.6 | −7.5 | −4.1 |
| Compound | RFI ± SD | |
|---|---|---|
| S. aureus 272123 | S. Typhimurium SL1344 | |
| 3 | ND | ND |
| 4 | ND | ND |
| 5 | 0.08 ± 0.07 | 0.22 ± 0.02 |
| 6 | 0.30 ± 0.05 | 0.22 ± 0.11 |
| 7 | −0.02 ± 0.01 | −0.07 ± 0.03 |
| 8 | −0.07 ± 0.03 | −0.07 ± 0.05 |
| 9 | −0.35 ± 0.01 | −0.41 ± 0.01 |
| 10 | −0.21 ± 0.03 | −0.36 ± 0.01 |
| 11 | −0.03 ± 0.03 | −0.06 ± 0.01 |
| 12 | 0.17 ± 0.03 | −0.01 ± 0.01 |
| 13 | 0.14 ± 0.02 | 0.06 ± 0.01 |
| Reserpine | 0.30 ± 0.12 | 0.04 ± 0.07 |
| CCCP | ND | 0.16 ± 0.06 |
| Compound | Molecular Weight | # Rotatable Bonds | # H-Bond Acceptors | # H-Bond Donors | ilogP |
|---|---|---|---|---|---|
| Reserpine | 608.68 | 10 | 10 | 1 | 5.16 |
| 5 | 330.38 | 9 | 4 | 2 | 3.12 |
| 6 | 355.39 | 8 | 5 | 2 | 2.46 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neves, A.R.; Durães, F.; Freitas-Silva, J.; Szemerédi, N.; Martins-da-Costa, P.; Pinto, E.; Correia-da-Silva, M.; Spengler, G.; Sousa, E. Derivatives of Trimethoxybenzoic Acid and Gallic Acid as Potential Efflux Pump Inhibitors: In Silico and In Vitro Studies. Int. J. Mol. Sci. 2022, 23, 14468. https://doi.org/10.3390/ijms232214468
Neves AR, Durães F, Freitas-Silva J, Szemerédi N, Martins-da-Costa P, Pinto E, Correia-da-Silva M, Spengler G, Sousa E. Derivatives of Trimethoxybenzoic Acid and Gallic Acid as Potential Efflux Pump Inhibitors: In Silico and In Vitro Studies. International Journal of Molecular Sciences. 2022; 23(22):14468. https://doi.org/10.3390/ijms232214468
Chicago/Turabian StyleNeves, Ana Rita, Fernando Durães, Joana Freitas-Silva, Nikoletta Szemerédi, Paulo Martins-da-Costa, Eugénia Pinto, Marta Correia-da-Silva, Gabriella Spengler, and Emília Sousa. 2022. "Derivatives of Trimethoxybenzoic Acid and Gallic Acid as Potential Efflux Pump Inhibitors: In Silico and In Vitro Studies" International Journal of Molecular Sciences 23, no. 22: 14468. https://doi.org/10.3390/ijms232214468
APA StyleNeves, A. R., Durães, F., Freitas-Silva, J., Szemerédi, N., Martins-da-Costa, P., Pinto, E., Correia-da-Silva, M., Spengler, G., & Sousa, E. (2022). Derivatives of Trimethoxybenzoic Acid and Gallic Acid as Potential Efflux Pump Inhibitors: In Silico and In Vitro Studies. International Journal of Molecular Sciences, 23(22), 14468. https://doi.org/10.3390/ijms232214468