
The Pivotal Role of Protein Phosphatase 2A (PP2A) in Brain Tumors


 , , ,
, , ,  ,
, 

Abstract
:1. Introduction
2. Role of PP2A in Cancer
3. Role of PP2A in GBM
4. PP2A Inhibitors in GBM
- LB100LB100 is a molecule derived from the synthetic anticancer compound norcantharidin [30,31]. LB100 competitively inhibits PP2A by directly binding to PP2A-C and reducing its catalytic activity [32]. Combination treatment with LB100 and radiation significantly delayed tumor growth, prolonging survival [33]. Cui et al. showed that LB100 enhanced the anti-CAIX CAR-T cell cytotoxic activity against GBM tumor cells [34]. Lu et al. demonstrated that LB100 inhibited PP2A and caused dose-dependent antiproliferative activity in GBM cell lines [35]. In another study conducted by the same group, Lu et al. studied LB-102, a lipid-soluble homolog of LB100, demonstrating that it combined with temozolomide, a DNA-methylating chemotherapeutic drug used to treat glioblastoma multiforme, causes complete regression of glioblastoma multiforme (GBM) xenografts. In addition, combined with doxorubicin (DOX), a DNA-intercalating agent used as an anticancer drug, LB-102 causes marked GBM xenograft regression, whereas DOX alone only slows growth. These findings indicate that inhibition of PP2A by LB-102 blocks cell-cycle arrest and increases progression of the cell cycle in the presence of TMZ or DOX [36].
- PME-1Protein phosphatase methylesterase 1 (PME-1) is an endogenous PP2A inhibitor protein that regulates PP2Ac activity by demethylating the highly conserved carboxyl-terminal tail and binding directly to the PP2Ac catalytic site. Kaur et al. demonstrated that overexpression of the PP2A inhibitor protein PME-1 drives glioma cell resistance to various multikinase inhibitors through specific PP2A complexes and a decrease in histone deacetylase 4 cytoplasmic activity. PME-1 and HDAC4 are associated with the progression of human glioma [27].
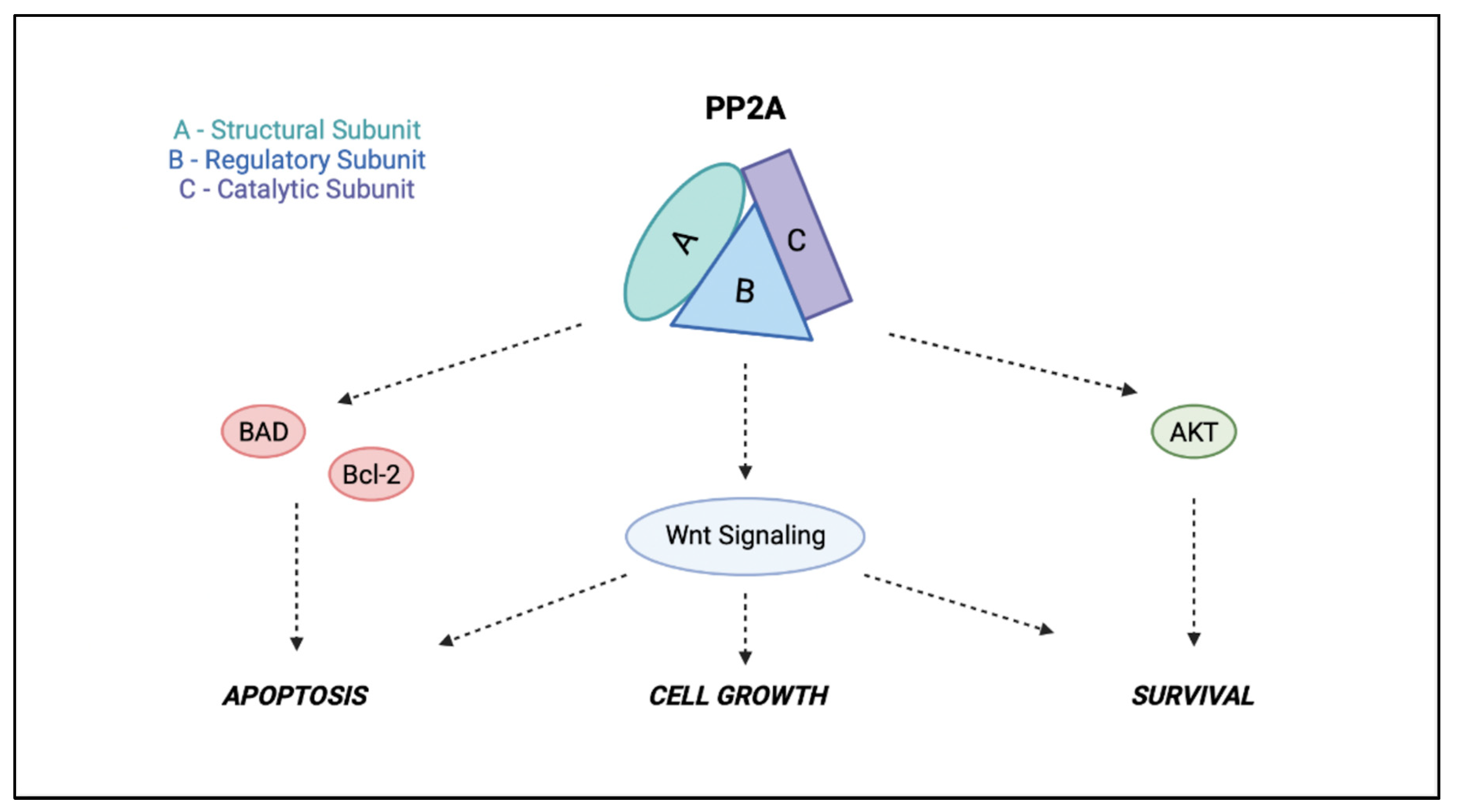
- SETOne of the endogenous inhibitors of PP2A is SET nuclear proto-oncogene (SET) [37]. In the context of GBM, numerous sources of evidence suggest that SET could represent a potential carcinogenic factor. Through the upregulation of Bcl-2 gene expression and downregulation of Bax and caspase-3 expression, it is able to regulate cell proliferation and apoptosis of GBM cells [38].
- CIP2AAnother endogenous inhibitor of PP2A is the cancerous inhibitor of PP2A (CIP2A), an oncoprotein upregulated in several peripheral tumors. CIP2A is able to promote the growth of cancer cells through the inhibition of dephosphorylation of PP2A substrates involved in cancer development. Moreover, CIP2A is found to be upregulated in mouse brain astrocytes (causing reactive astrogliosis), which promote synaptic degeneration and cognitive deficits [29]. CIP2A, directly interacting with the oncogenic transcription factor c-Myc, inhibits the activity of PP2A toward c-Myc serine 62 (S62) and, thus, prevents c-Myc proteolytic degradation. In addition, CIP2A promotes in vivo tumor formation and anchorage-independent cell growth [29].
5. CIP2A
6. Role of CIP2A in Glioma and GBM
7. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Meeusen, B.; Janssens, V. Tumor suppressive protein phosphatases in human cancer: Emerging targets for therapeutic intervention and tumor stratification. Int. J. Biochem. Cell Biol. 2018, 96, 98–134. [Google Scholar] [CrossRef] [PubMed]
- Mazhar, S.; Taylor, S.E.; Sangodkar, J.; Narla, G. Targeting PP2A in cancer: Combination therapies. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Brautigan, D.L.; Farrington, C.; Narla, G. Targeting protein phosphatase PP2A for cancer therapy: Development of allosteric pharmaceutical agents. Clin. Sci. (Lond.) 2021, 135, 1545–1556. [Google Scholar] [CrossRef]
- Lambrecht, C.; Libbrecht, L.; Sagaert, X.; Pauwels, P.; Hoorne, Y.; Crowther, J.; Louis, J.V.; Sents, W.; Sablina, A.; Janssens, V. Loss of protein phosphatase 2A regulatory subunit B56delta promotes spontaneous tumorigenesis in vivo. Oncogene 2018, 37, 544–552. [Google Scholar] [CrossRef] [PubMed]
- Seshacharyulu, P.; Pandey, P.; Datta, K.; Batra, S.K. Phosphatase: PP2A structural importance, regulation and its aberrant expression in cancer. Cancer Lett. 2013, 335, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, C.E.; Li, S.; Vizeacoumar, F.S.; Bhanumathy, K.K.; Lee, J.S.; Parameswaran, S.; Furber, L.; Abuhussein, O.; Paul, J.M.; McDonald, M.; et al. Therapeutic relevance of the protein phosphatase 2A in cancer. Oncotarget 2016, 7, 61544–61561. [Google Scholar] [CrossRef] [Green Version]
- Baskaran, R.; Velmurugan, B.K. Protein phosphatase 2A as therapeutic targets in various disease models. Life Sci. 2018, 210, 40–46. [Google Scholar] [CrossRef]
- Perrotti, D.; Neviani, P. Protein phosphatase 2A: A target for anticancer therapy. Lancet Oncol. 2013, 14, e229–e238. [Google Scholar] [CrossRef] [Green Version]
- Arroyo, J.D.; Hahn, W.C. Involvement of PP2A in viral and cellular transformation. Oncogene 2005, 24, 7746–7755. [Google Scholar] [CrossRef] [Green Version]
- Eichhorn, P.J.; Creyghton, M.P.; Bernards, R. Protein phosphatase 2A regulatory subunits and cancer. Biochim. Biophys. Acta 2009, 1795, 1–15. [Google Scholar] [CrossRef]
- Janssens, V.; Goris, J. Protein phosphatase 2A: A highly regulated family of serine/threonine phosphatases implicated in cell growth and signalling. Biochem. J. 2001, 353, 417–439. [Google Scholar] [CrossRef] [PubMed]
- Takagi, Y.; Futamura, M.; Yamaguchi, K.; Aoki, S.; Takahashi, T.; Saji, S. Alterations of the PPP2R1B gene located at 11q23 in human colorectal cancers. Gut 2000, 47, 268–271. [Google Scholar] [CrossRef] [PubMed]
- Calin, G.A.; di Iasio, M.G.; Caprini, E.; Vorechovsky, I.; Natali, P.G.; Sozzi, G.; Croce, C.M.; Barbanti-Brodano, G.; Russo, G.; Negrini, M. Low frequency of alterations of the alpha (PPP2R1A) and beta (PPP2R1B) isoforms of the subunit A of the serine-threonine phosphatase 2A in human neoplasms. Oncogene 2000, 19, 1191–1195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.S.; Esplin, E.D.; Li, J.L.; Huang, L.; Gazdar, A.; Minna, J.; Evans, G.A. Alterations of the PPP2R1B gene in human lung and colon cancer. Science 1998, 282, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Campbell, I.G.; Manolitsas, T. Absence of PPP2R1B gene alterations in primary ovarian cancers. Oncogene 1999, 18, 6367–6369. [Google Scholar] [CrossRef] [Green Version]
- Schonthal, A.H. Role of serine/threonine protein phosphatase 2A in cancer. Cancer Lett. 2001, 170, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wlodarchak, N.; Xing, Y. PP2A as a master regulator of the cell cycle. Crit. Rev. Biochem. Mol. Biol. 2016, 51, 162–184. [Google Scholar] [CrossRef]
- Yang, D.; Okamura, H.; Morimoto, H.; Teramachi, J.; Haneji, T. Protein phosphatase 2A Calpha regulates proliferation, migration, and metastasis of osteosarcoma cells. Lab. Investig. 2016, 96, 1050–1062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, G.; Ruediger, R. Mouse model for probing tumor suppressor activity of protein phosphatase 2A in diverse signaling pathways. Cell Cycle 2012, 11, 451–459. [Google Scholar] [CrossRef] [Green Version]
- Basilotta, R.; Lanza, M.; Casili, G.; Chisari, G.; Munao, S.; Colarossi, L.; Cucinotta, L.; Campolo, M.; Esposito, E.; Paterniti, I. Potential Therapeutic Effects of PPAR Ligands in Glioblastoma. Cells 2022, 11, 621. [Google Scholar] [CrossRef]
- Scuderi, S.A.; Casili, G.; Ardizzone, A.; Forte, S.; Colarossi, L.; Sava, S.; Paterniti, I.; Esposito, E.; Cuzzocrea, S.; Campolo, M. KYP-2047, an Inhibitor of Prolyl-Oligopeptidase, Reduces GlioBlastoma Proliferation through Angiogenesis and Apoptosis Modulation. Cancers 2021, 13, 3444. [Google Scholar] [CrossRef] [PubMed]
- Tomiyama, A.; Kobayashi, T.; Mori, K.; Ichimura, K. Protein Phosphatases-A Touchy Enemy in the Battle Against Glioblastomas: A Review. Cancers 2019, 11, 241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dedobbeleer, M.; Willems, E.; Freeman, S.; Lombard, A.; Goffart, N.; Rogister, B. Phosphatases and solid tumors: Focus on glioblastoma initiation, progression and recurrences. Biochem. J. 2017, 474, 2903–2924. [Google Scholar] [CrossRef] [PubMed]
- Hofstetter, C.P.; Burkhardt, J.K.; Shin, B.J.; Gursel, D.B.; Mubita, L.; Gorrepati, R.; Brennan, C.; Holland, E.C.; Boockvar, J.A. Protein phosphatase 2A mediates dormancy of glioblastoma multiforme-derived tumor stem-like cells during hypoxia. PLoS ONE 2012, 7, e30059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, X.; Ma, N.; Du, L.; Liang, Y.; Zhang, P.; Han, Y.; Qu, B. PP2A and tumor radiotherapy. Hereditas 2020, 157, 36. [Google Scholar] [CrossRef]
- Merisaari, J.; Denisova, O.V.; Doroszko, M.; Le Joncour, V.; Johansson, P.; Leenders, W.P.J.; Kastrinsky, D.B.; Zaware, N.; Narla, G.; Laakkonen, P.; et al. Monotherapy efficacy of blood-brain barrier permeable small molecule reactivators of protein phosphatase 2A in glioblastoma. Brain Commun. 2020, 2, fcaa002. [Google Scholar] [CrossRef] [Green Version]
- Kaur, A.; Denisova, O.V.; Qiao, X.; Jumppanen, M.; Peuhu, E.; Ahmed, S.U.; Raheem, O.; Haapasalo, H.; Eriksson, J.; Chalmers, A.J.; et al. PP2A Inhibitor PME-1 Drives Kinase Inhibitor Resistance in Glioma Cells. Cancer Res. 2016, 76, 7001–7011. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, C.M.; Perl, A.; Leonard, D.; Sangodkar, J.; Narla, G. Therapeutic targeting of PP2A. Int. J. Biochem. Cell Biol. 2018, 96, 182–193. [Google Scholar] [CrossRef]
- Junttila, M.R.; Puustinen, P.; Niemela, M.; Ahola, R.; Arnold, H.; Bottzauw, T.; Ala-aho, R.; Nielsen, C.; Ivaska, J.; Taya, Y.; et al. CIP2A inhibits PP2A in human malignancies. Cell 2007, 130, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Maggio, D.; Ho, W.S.; Breese, R.; Walbridge, S.; Wang, H.; Cui, J.; Heiss, J.D.; Gilbert, M.R.; Kovach, J.S.; Lu, R.O.; et al. Inhibition of protein phosphatase-2A with LB-100 enhances antitumor immunity against glioblastoma. J. Neurooncol. 2020, 148, 231–244. [Google Scholar] [CrossRef]
- Otani, Y.; Sur, H.P.; Rachaiah, G.; Namagiri, S.; Chowdhury, A.; Lewis, C.T.; Shimizu, T.; Gangaplara, A.; Wang, X.; Vezina, A.; et al. Inhibiting protein phosphatase 2A increases the antitumor effect of protein arginine methyltransferase 5 inhibition in models of glioblastoma. Neuro-oncology 2021, 23, 1481–1493. [Google Scholar] [CrossRef] [PubMed]
- Bryant, J.P.; Levy, A.; Heiss, J.; Banasavadi-Siddegowda, Y.K. Review of PP2A Tumor Biology and Antitumor Effects of PP2A Inhibitor LB100 in the Nervous System. Cancers 2021, 13, 87. [Google Scholar] [CrossRef] [PubMed]
- Gordon, I.K.; Lu, J.; Graves, C.A.; Huntoon, K.; Frerich, J.M.; Hanson, R.H.; Wang, X.; Hong, C.S.; Ho, W.; Feldman, M.J.; et al. Protein Phosphatase 2A Inhibition with LB100 Enhances Radiation-Induced Mitotic Catastrophe and Tumor Growth Delay in Glioblastoma. Mol. Cancer Ther. 2015, 14, 1540–1547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, J.; Wang, H.; Medina, R.; Zhang, Q.; Xu, C.; Indig, I.H.; Zhou, J.; Song, Q.; Dmitriev, P.; Sun, M.Y.; et al. Inhibition of PP2A with LB-100 Enhances Efficacy of CAR-T Cell Therapy Against Glioblastoma. Cancers 2020, 12, 139. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Zhuang, Z.; Song, D.K.; Mehta, G.U.; Ikejiri, B.; Mushlin, H.; Park, D.M.; Lonser, R.R. The effect of a PP2A inhibitor on the nuclear receptor corepressor pathway in glioma. J. Neurosurg. 2010, 113, 225–233. [Google Scholar] [CrossRef]
- Lu, J.; Kovach, J.S.; Johnson, F.; Chiang, J.; Hodes, R.; Lonser, R.; Zhuang, Z. Inhibition of serine/threonine phosphatase PP2A enhances cancer chemotherapy by blocking DNA damage induced defense mechanisms. Proc. Natl. Acad. Sci. USA 2009, 106, 11697–11702. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Gu, Y.; Wang, H.; Yin, J.; Zheng, G.; Zhang, Z.; Lu, M.; Wang, C.; He, Z. Overexpression of PP2A inhibitor SET oncoprotein is associated with tumor progression and poor prognosis in human non-small cell lung cancer. Oncotarget 2015, 6, 14913–14925. [Google Scholar] [CrossRef] [Green Version]
- He, K.; Shi, L.; Jiang, T.; Li, Q.; Chen, Y.; Meng, C. Association between SET expression and glioblastoma cell apoptosis and proliferation. Oncol. Lett. 2016, 12, 2435–2444. [Google Scholar] [CrossRef] [Green Version]
- Yi, F.; Ni, W.; Liu, W.; Bai, J.; Li, W. Expression and biological role of CIP2A in human astrocytoma. Mol. Med. Rep. 2013, 7, 1376–1380. [Google Scholar] [CrossRef] [Green Version]
- Birkman, E.M.; Elzagheid, A.; Jokilehto, T.; Avoranta, T.; Korkeila, E.; Kulmala, J.; Syrjanen, K.; Westermarck, J.; Sundstrom, J. Protein phosphatase 2A (PP2A) inhibitor CIP2A indicates resistance to radiotherapy in rectal cancer. Cancer Med. 2018, 7, 698–706. [Google Scholar] [CrossRef]
- Kim, M.O.; Choe, M.H.; Yoon, Y.N.; Ahn, J.; Yoo, M.; Jung, K.Y.; An, S.; Hwang, S.G.; Oh, J.S.; Kim, J.S. Antihelminthic drug niclosamide inhibits CIP2A and reactivates tumor suppressor protein phosphatase 2A in non-small cell lung cancer cells. Biochem. Pharmacol. 2017, 144, 78–89. [Google Scholar] [CrossRef] [PubMed]
- Laine, A.; Sihto, H.; Come, C.; Rosenfeldt, M.T.; Zwolinska, A.; Niemela, M.; Khanna, A.; Chan, E.K.; Kahari, V.M.; Kellokumpu-Lehtinen, P.L.; et al. Senescence sensitivity of breast cancer cells is defined by positive feedback loop between CIP2A and E2F1. Cancer Discov. 2013, 3, 182–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, D.; Nyalali, A.M.K.; Hou, Y.; Xu, Y.; Zhou, J.; Zhao, W.; Huang, B.; Li, F. 2,5-Dimethyl Celecoxib Inhibits Proliferation and Cell Cycle and Induces Apoptosis in Glioblastoma by Suppressing CIP2A/PP2A/Akt Signaling Axis. J. Mol. Neurosci. 2021, 71, 1703–1713. [Google Scholar] [CrossRef] [PubMed]
- Khanna, A.; Pimanda, J.E.; Westermarck, J. Cancerous inhibitor of protein phosphatase 2A, an emerging human oncoprotein and a potential cancer therapy target. Cancer Res. 2013, 73, 6548–6553. [Google Scholar] [CrossRef] [Green Version]
- De, P.; Carlson, J.; Leyland-Jones, B.; Dey, N. Oncogenic nexus of cancerous inhibitor of protein phosphatase 2A (CIP2A): An oncoprotein with many hands. Oncotarget 2014, 5, 4581–4602. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Ge, Z.; Liu, C.; Liu, Z.; Bjorkholm, M.; Jia, J.; Xu, D. CIP2A is overexpressed in gastric cancer and its depletion leads to impaired clonogenicity, senescence, or differentiation of tumor cells. Clin. Cancer Res. 2008, 14, 3722–3728. [Google Scholar] [CrossRef] [Green Version]
- Qu, W.; Li, W.; Wei, L.; Xing, L.; Wang, X.; Yu, J. CIP2A is overexpressed in esophageal squamous cell carcinoma. Med. Oncol. 2012, 29, 113–118. [Google Scholar] [CrossRef]
- Chen, K.F.; Yen, C.C.; Lin, J.K.; Chen, W.S.; Yang, S.H.; Jiang, J.K.; Lan, Y.T.; Lin, C.C.; Yu, H.C.; Hsu, H.M.; et al. Cancerous inhibitor of protein phosphatase 2A (CIP2A) is an independent prognostic marker in wild-type KRAS metastatic colorectal cancer after colorectal liver metastasectomy. BMC Cancer 2015, 15, 301. [Google Scholar] [CrossRef] [Green Version]
- Farrell, A.S.; Allen-Petersen, B.; Daniel, C.J.; Wang, X.; Wang, Z.; Rodriguez, S.; Impey, S.; Oddo, J.; Vitek, M.P.; Lopez, C.; et al. Targeting inhibitors of the tumor suppressor PP2A for the treatment of pancreatic cancer. Mol. Cancer Res. 2014, 12, 924–939. [Google Scholar] [CrossRef] [Green Version]
- Tang, Q.; Wang, Q.; Zeng, G.; Li, Q.; Jiang, T.; Zhang, Z.; Zheng, W.; Wang, K. Overexpression of CIP2A in clear cell renal cell carcinoma promotes cellular epithelial-mesenchymal transition and is associated with poor prognosis. Oncol. Rep. 2015, 34, 2515–2522. [Google Scholar] [CrossRef]
- Liu, J.; Wang, X.; Zhou, G.; Wang, H.; Xiang, L.; Cheng, Y.; Liu, W.; Wang, Y.; Jia, J.; Zhao, W. Cancerous inhibitor of protein phosphatase 2A is overexpressed in cervical cancer and upregulated by human papillomavirus 16 E7 oncoprotein. Gynecol. Oncol. 2011, 122, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Li, Z.; Wang, X.; Zhang, S. CIP2A is overexpressed in human ovarian cancer and regulates cell proliferation and apoptosis. Tumour Biol. 2012, 33, 2299–2306. [Google Scholar] [CrossRef] [PubMed]
- Soofiyani, S.R.; Hejazi, M.S.; Baradaran, B. The role of CIP2A in cancer: A review and update. Biomed. Pharmacother. 2017, 96, 626–633. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Liang, J.L.; Zhou, K.; Zeng, Q.L.; Ye, J.W.; Huang, M.J. Effect of CIP2A and its mechanism of action in the malignant biological behavior of colorectal cancer. Cell Commun. Signal. 2020, 18, 67. [Google Scholar] [CrossRef] [Green Version]
- Grech, G.; Baldacchino, S.; Saliba, C.; Grixti, M.P.; Gauci, R.; Petroni, V.; Fenech, A.G.; Scerri, C. Deregulation of the protein phosphatase 2A, PP2A in cancer: Complexity and therapeutic options. Tumour Biol. 2016, 37, 11691–11700. [Google Scholar] [CrossRef]
- Kim, J.S.; Kim, E.J.; Oh, J.S.; Park, I.C.; Hwang, S.G. CIP2A modulates cell-cycle progression in human cancer cells by regulating the stability and activity of Plk1. Cancer Res. 2013, 73, 6667–6678. [Google Scholar] [CrossRef] [Green Version]
- Puustinen, P.; Rytter, A.; Mortensen, M.; Kohonen, P.; Moreira, J.M.; Jaattela, M. CIP2A oncoprotein controls cell growth and autophagy through mTORC1 activation. J. Cell Biol. 2014, 204, 713–727. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, K. miR-383 Down-Regulates the Oncogene CIP2A to Influence Glioma Proliferation and Invasion. Onco-Targets Ther 2020, 13, 4063–4074. [Google Scholar] [CrossRef]
- Xu, Z.; Chen, X.; Hegazy, A.M.; Zhang, S.; Liu, X.; Tian, J.; Wu, J.; Shi, F.; Li, L.; Niu, X.; et al. Depletion of CIP2A inhibits the proliferation, migration, invasion and epithelial-mesenchymal transition of glioma cells. Brain Res. Bull. 2021, 173, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Khanna, A.; Thoms, J.A.I.; Stringer, B.W.; Chung, S.A.; Ensbey, K.S.; Jue, T.R.; Jahan, Z.; Subramanian, S.; Anande, G.; Shen, H.; et al. Constitutive CHK1 Expression Drives a pSTAT3-CIP2A Circuit that Promotes Glioblastoma Cell Survival and Growth. Mol. Cancer Res. 2020, 18, 709–722. [Google Scholar] [CrossRef]
- Qin, S.; Li, J.; Si, Y.; He, Z.; Zhang, T.; Wang, D.; Liu, X.; Guo, Y.; Zhang, L.; Li, S.; et al. Cucurbitacin B induces inhibitory effects via CIP2A/PP2A/Akt pathway in glioblastoma multiforme. Mol. Carcinog 2018, 57, 687–699. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Inhibitor | Mode of Inhibition | References |
|---|---|---|
| LB100 | Binds to PP2A-C and reduces its catalytic activity | [30,31,32,33,34,35,36] |
| PME-1 | Demethylation of L309 on the subunit C of PP2A | [27] |
| SET | Binds to and inactivates PP2A | [37,38] |
| CIP2A | Binds to PP2A, preventing the dephosphorylation of c-MYC and Akt | [29] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cucinotta, L.; Filippone, A.; Casili, G.; Lanza, M.; Bova, V.; Capra, A.P.; Giuffrida, R.; Colarossi, C.; Sciacca, D.; Paterniti, I.; et al. The Pivotal Role of Protein Phosphatase 2A (PP2A) in Brain Tumors. Int. J. Mol. Sci. 2022, 23, 15717. https://doi.org/10.3390/ijms232415717
Cucinotta L, Filippone A, Casili G, Lanza M, Bova V, Capra AP, Giuffrida R, Colarossi C, Sciacca D, Paterniti I, et al. The Pivotal Role of Protein Phosphatase 2A (PP2A) in Brain Tumors. International Journal of Molecular Sciences. 2022; 23(24):15717. https://doi.org/10.3390/ijms232415717
Chicago/Turabian StyleCucinotta, Laura, Alessia Filippone, Giovanna Casili, Marika Lanza, Valentina Bova, Anna Paola Capra, Raffaella Giuffrida, Cristina Colarossi, Dorotea Sciacca, Irene Paterniti, and et al. 2022. "The Pivotal Role of Protein Phosphatase 2A (PP2A) in Brain Tumors" International Journal of Molecular Sciences 23, no. 24: 15717. https://doi.org/10.3390/ijms232415717
APA StyleCucinotta, L., Filippone, A., Casili, G., Lanza, M., Bova, V., Capra, A. P., Giuffrida, R., Colarossi, C., Sciacca, D., Paterniti, I., Cuzzocrea, S., Campolo, M., & Esposito, E. (2022). The Pivotal Role of Protein Phosphatase 2A (PP2A) in Brain Tumors. International Journal of Molecular Sciences, 23(24), 15717. https://doi.org/10.3390/ijms232415717