

A Decade of CRISPR-Cas Gnome Editing in C. elegans

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
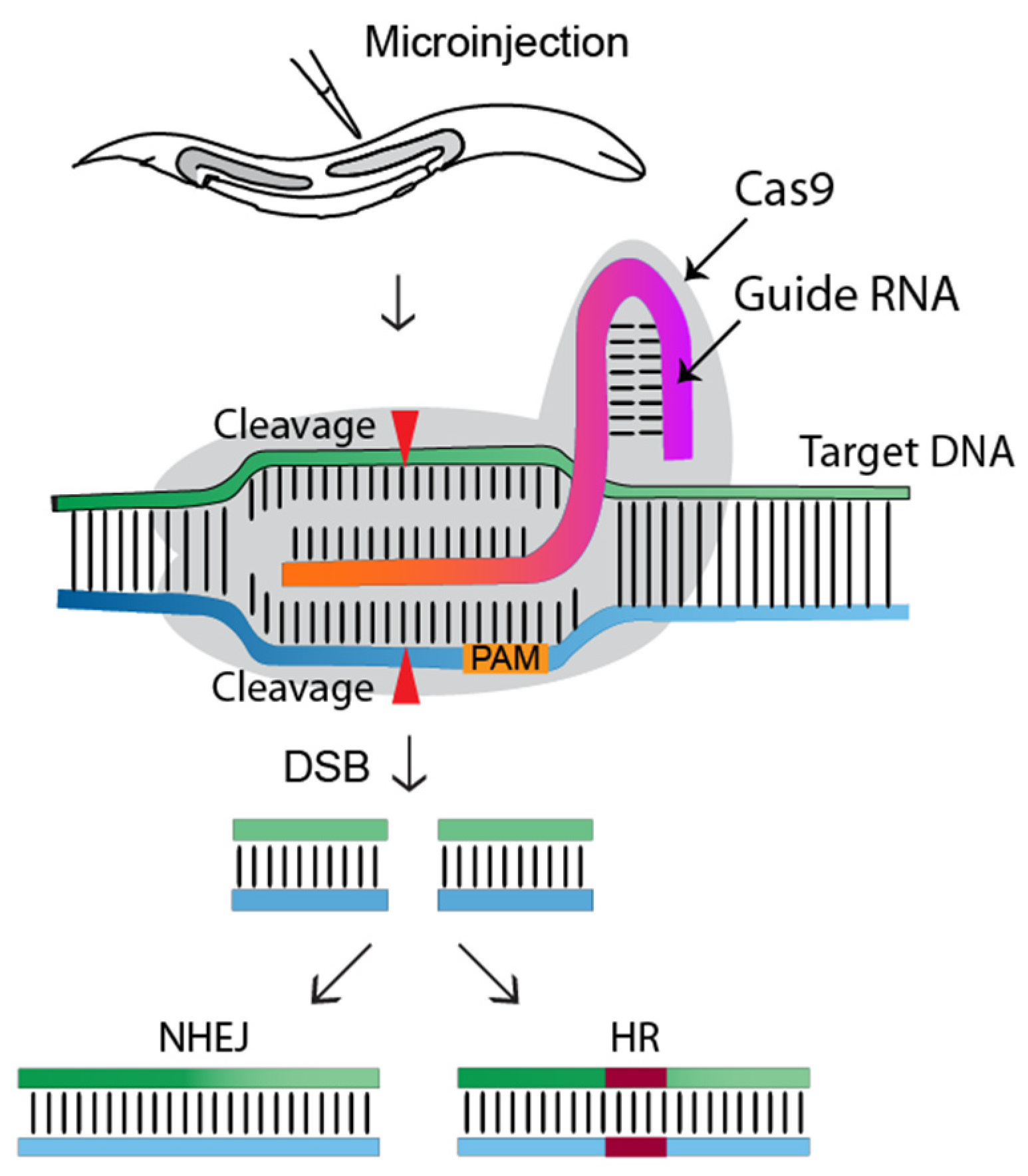
:1. Introduction
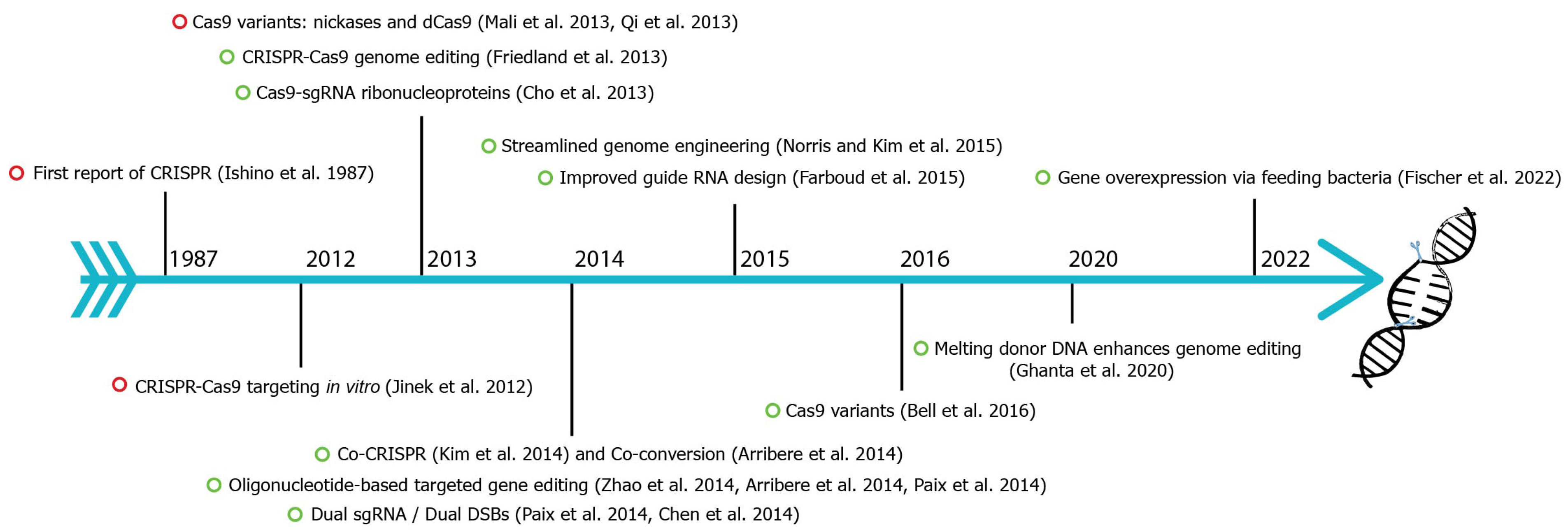
2. History of C. elegans CRISPR-Cas
3. Screening of Transgenic Worms
3.1. PCR and Drug-Based Selection
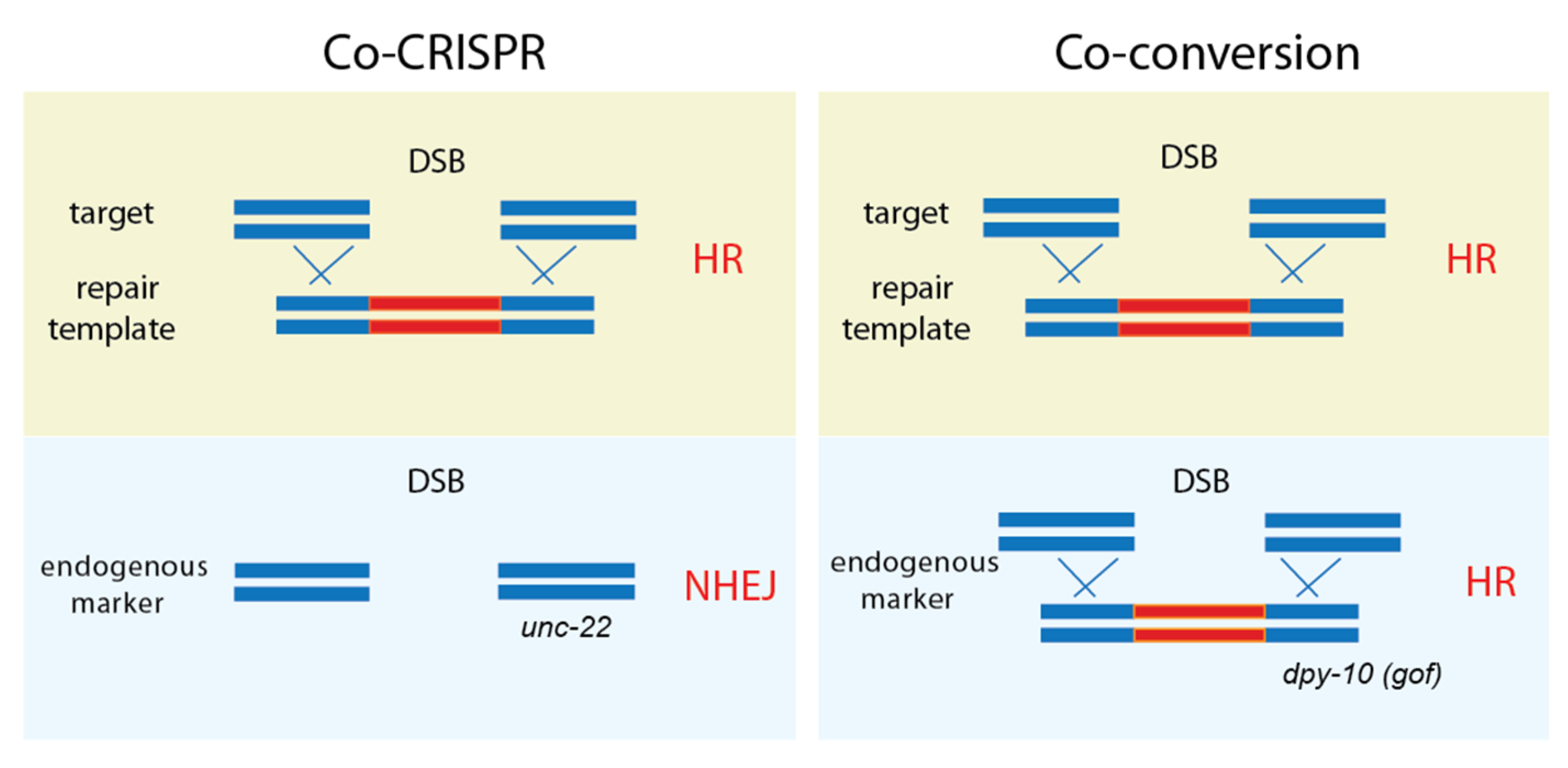
3.2. Co-CRISPR and Co-Conversion: A Mutation in an Endogenous Marker Presents A Visible Phenotype
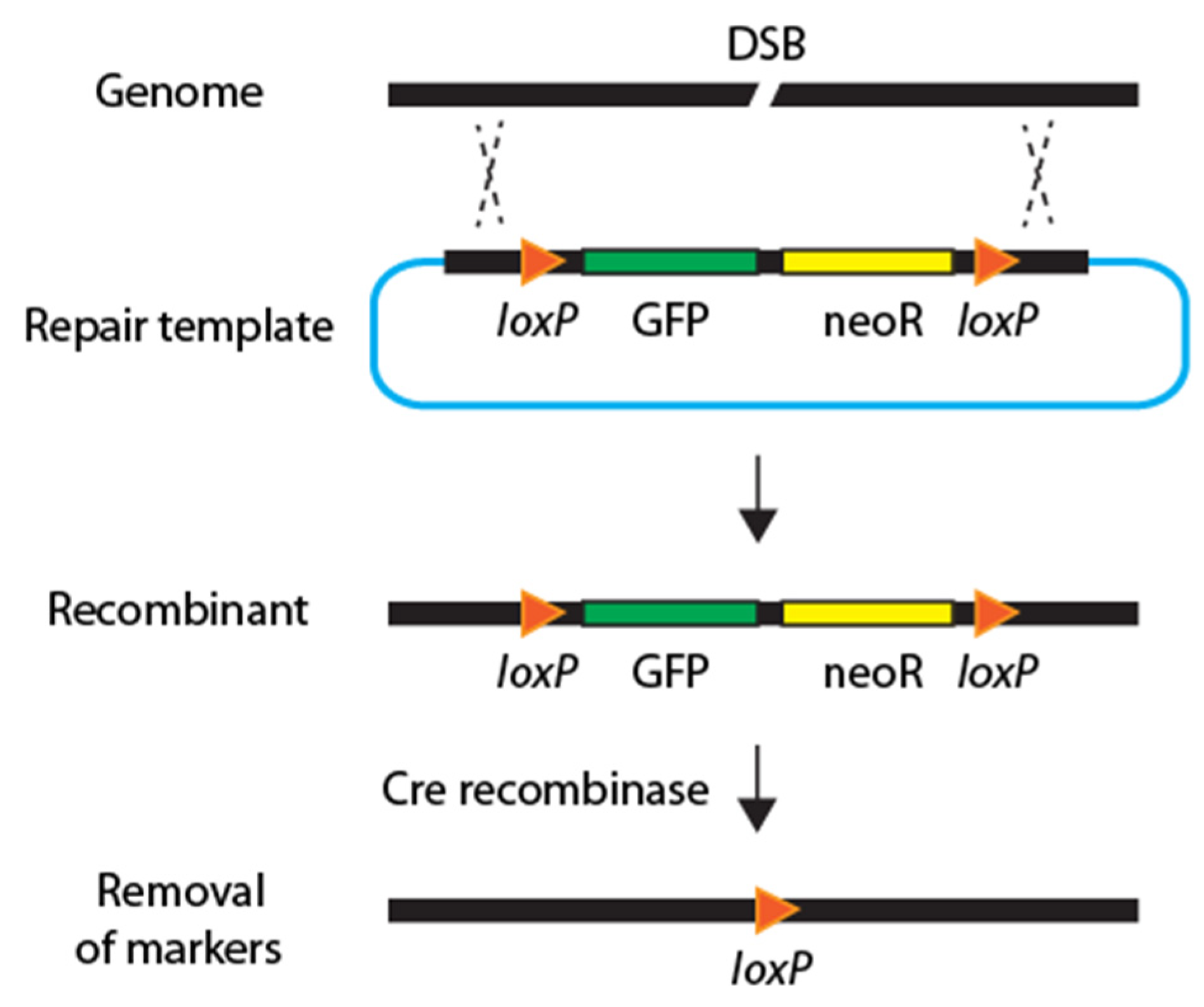
3.3. Streamlined Genome Editing
4. Advances in Genome Editing Components
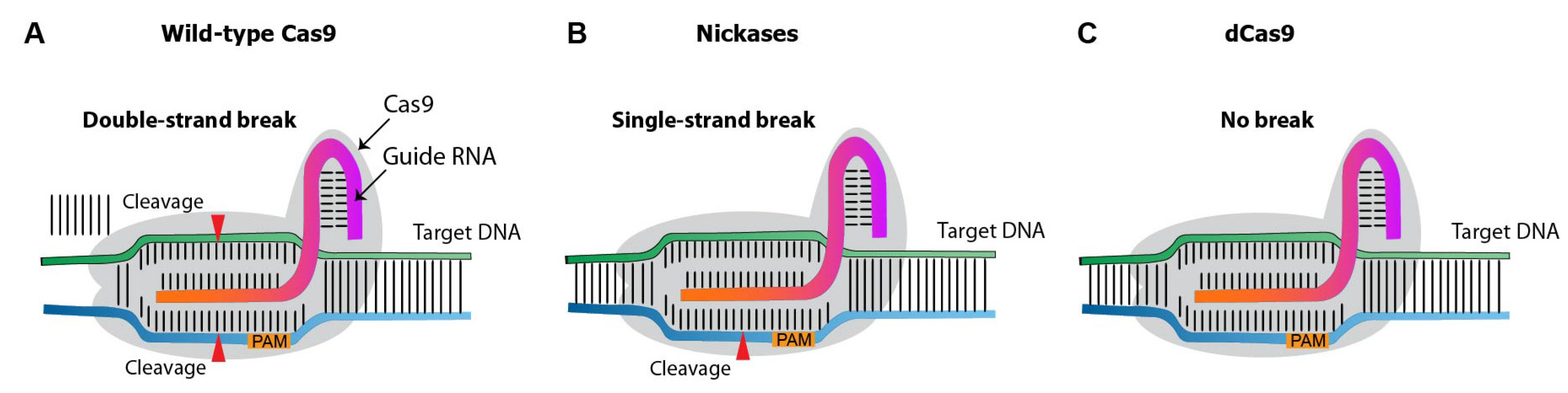
4.1. Cas9 Variants
4.2. Guide RNA
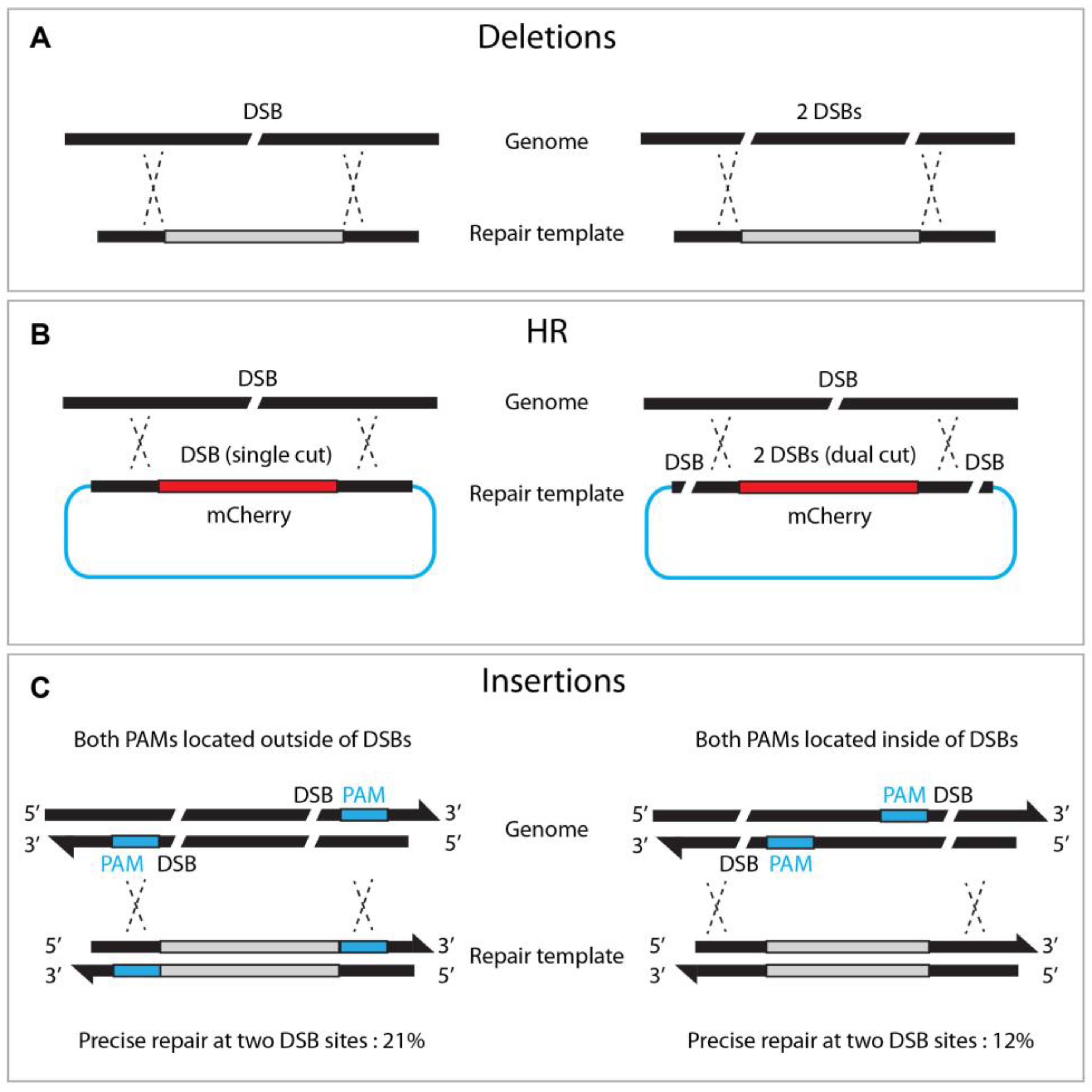
4.3. Dual sgRNA/Dual DSBs
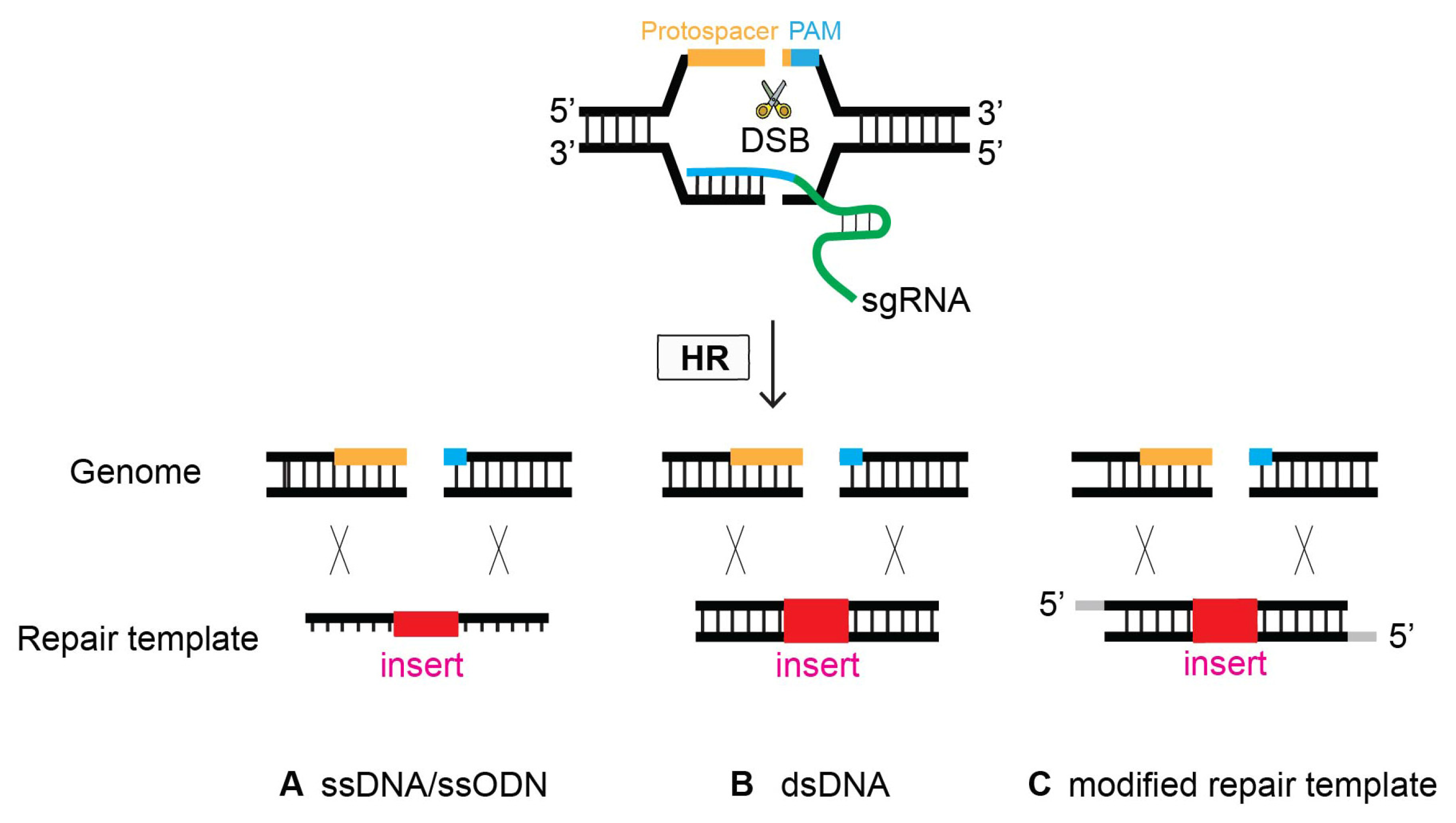
4.4. Oligonucleotide as a Repair Template for Homologous Recombination
4.5. Modified Repair Template
5. Other Developments
5.1. Off-Target Effects
5.2. CRISPR-Cas Genome Editing Protocols
- Efficient Genome Editing in Caenorhabditis elegans with a Toolkit of Dual-Marker Selection Cassettes [13].
- Precision genome editing using CRISPR-Cas9 and linear repair templates in C. elegans [43].
- An Efficient Genome Editing Strategy to Generate Putative Null Mutants in Caenorhabditis elegans Using CRISPR/Cas9 [44].
- CRISPR-Cas9-Guided Genome Engineering in Caenorhabditis elegans [1].
- An affordable plasmid miniprep suitable for proficient microinjection in Caenorhabditis elegans [45].
- Design of Repair Templates for CRISPR-Cas9-Triggered Homologous Recombination in Caenorhabditis elegans [46].
- Microinjection for precision genome editing in Caenorhabditis elegans [47].
- High-efficiency CRISPR gene editing in C. elegans using Cas9 integrated into the genome [48].
- Approaches for CRISPR/Cas9 Genome Editing in C. elegans [49].
- CRISPR/Cas9 Methodology for the Generation of Knockout Deletions in Caenorhabditis elegans [50].
- CRISPR-Based Methods for Caenorhabditis elegans Genome Engineering [51].
6. Perspectives
6.1. Pros and Impact
6.2. C. elegans for CRISPR-Cas Genome Editing
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, H.M.; Colaiacovo, M.P. CRISPR-Cas9-Guided Genome Engineering in Caenorhabditis elegans. Curr. Protoc. Mol. Biol. 2019, 129, e106. [Google Scholar] [CrossRef] [PubMed]
- Ishino, Y.; Shinagawa, H.; Makino, K.; Amemura, M.; Nakata, A. Nucleotide sequence of the iap gene, responsible for alkaline phosphatase isozyme conversion in Escherichia coli, and identification of the gene product. J. Bacteriol. 1987, 169, 5429–5433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Mali, P.; Aach, J.; Stranges, P.B.; Esvelt, K.M.; Moosburner, M.; Kosuri, S.; Yang, L.; Church, G.M. CAS9 transcriptional activators for target specificity screening and paired nickases for cooperative genome engineering. Nat. Biotechnol. 2013, 31, 833–838. [Google Scholar] [CrossRef]
- Qi, L.S.; Larson, M.H.; Gilbert, L.A.; Doudna, J.A.; Weissman, J.S.; Arkin, A.P.; Lim, W.A. Repurposing CRISPR as an RNA-guided platform for sequence-specific control of gene expression. Cell 2013, 152, 1173–1183. [Google Scholar] [CrossRef] [Green Version]
- Friedland, A.E.; Tzur, Y.B.; Esvelt, K.M.; Colaiacovo, M.P.; Church, G.M.; Calarco, J.A. Heritable genome editing in C. elegans via a CRISPR-Cas9 system. Nat. Methods 2013, 10, 741–743. [Google Scholar] [CrossRef] [Green Version]
- Lo, T.W.; Pickle, C.S.; Lin, S.; Ralston, E.J.; Gurling, M.; Schartner, C.M.; Bian, Q.; Doudna, J.A.; Meyer, B.J. Precise and heritable genome editing in evolutionarily diverse nematodes using TALENs and CRISPR/Cas9 to engineer insertions and deletions. Genetics 2013, 195, 331–348. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Fenk, L.A.; de Bono, M. Efficient genome editing in Caenorhabditis elegans by CRISPR-targeted homologous recombination. Nucleic Acids Res. 2013, 41, e193. [Google Scholar] [CrossRef] [Green Version]
- Tzur, Y.B.; Friedland, A.E.; Nadarajan, S.; Church, G.M.; Calarco, J.A.; Colaiacovo, M.P. Heritable custom genomic modifications in Caenorhabditis elegans via a CRISPR-Cas9 system. Genetics 2013, 195, 1181–1185. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.W.; Lee, J.; Carroll, D.; Kim, J.S.; Lee, J. Heritable gene knockout in Caenorhabditis elegans by direct injection of Cas9-sgRNA ribonucleoproteins. Genetics 2013, 195, 1177–1180. [Google Scholar] [CrossRef]
- Kim, H.; Ishidate, T.; Ghanta, K.S.; Seth, M.; Conte, D., Jr.; Shirayama, M.; Mello, C.C. A co-CRISPR strategy for efficient genome editing in Caenorhabditis elegans. Genetics 2014, 197, 1069–1080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arribere, J.A.; Bell, R.T.; Fu, B.X.; Artiles, K.L.; Hartman, P.S.; Fire, A.Z. Efficient marker-free recovery of custom genetic modifications with CRISPR/Cas9 in Caenorhabditis elegans. Genetics 2014, 198, 837–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norris, A.D.; Kim, H.M.; Colaiacovo, M.P.; Calarco, J.A. Efficient Genome Editing in Caenorhabditis elegans with a Toolkit of Dual-Marker Selection Cassettes. Genetics 2015, 201, 449–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dickinson, D.J.; Pani, A.M.; Heppert, J.K.; Higgins, C.D.; Goldstein, B. Streamlined Genome Engineering with a Self-Excising Drug Selection Cassette. Genetics 2015, 200, 1035–1049. [Google Scholar] [CrossRef] [Green Version]
- Bell, R.T.; Fu, B.X.; Fire, A.Z. Cas9 Variants Expand the Target Repertoire in Caenorhabditis elegans. Genetics 2016, 202, 381–388. [Google Scholar] [CrossRef] [Green Version]
- Farboud, B.; Meyer, B.J. Dramatic enhancement of genome editing by CRISPR/Cas9 through improved guide RNA design. Genetics 2015, 199, 959–971. [Google Scholar] [CrossRef] [Green Version]
- Paix, A.; Wang, Y.; Smith, H.E.; Lee, C.Y.; Calidas, D.; Lu, T.; Smith, J.; Schmidt, H.; Krause, M.W.; Seydoux, G. Scalable and versatile genome editing using linear DNAs with microhomology to Cas9 Sites in Caenorhabditis elegans. Genetics 2014, 198, 1347–1356. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Xu, F.; Zhu, C.; Ji, J.; Zhou, X.; Feng, X.; Guang, S. Dual sgRNA-directed gene knockout using CRISPR/Cas9 technology in Caenorhabditis elegans. Sci. Rep. 2014, 4, 7581. [Google Scholar] [CrossRef] [Green Version]
- Zhao, P.; Zhang, Z.; Ke, H.; Yue, Y.; Xue, D. Oligonucleotide-based targeted gene editing in C. elegans via the CRISPR/Cas9 system. Cell Res. 2014, 24, 247–250. [Google Scholar] [CrossRef] [Green Version]
- Ghanta, K.S.; Mello, C.C. Melting dsDNA Donor Molecules Greatly Improves Precision Genome Editing in Caenorhabditis elegans. Genetics 2020, 216, 643–650. [Google Scholar] [CrossRef]
- Fischer, F.; Benner, C.; Goyala, A.; Grigolon, G.; Vitiello, D.; Wu, J.; Zarse, K.; Ewald, C.Y.; Ristow, M. Ingestion of single guide RNAs induces gene overexpression and extends lifespan in Caenorhabditis elegans via CRISPR activation. J. Biol. Chem. 2022, 298, 102085. [Google Scholar] [CrossRef] [PubMed]
- Plasterk, R.H.; Groenen, J.T. Targeted alterations of the Caenorhabditis elegans genome by transgene instructed DNA double strand break repair following Tc1 excision. EMBO J. 1992, 11, 287–290. [Google Scholar] [CrossRef]
- Berezikov, E.; Bargmann, C.I.; Plasterk, R.H. Homologous gene targeting in Caenorhabditis elegans by biolistic transformation. Nucleic Acids Res. 2004, 32, e40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleinstiver, B.P.; Prew, M.S.; Tsai, S.Q.; Topkar, V.V.; Nguyen, N.T.; Zheng, Z.; Gonzales, A.P.; Li, Z.; Peterson, R.T.; Yeh, J.R.; et al. Engineered CRISPR-Cas9 nucleases with altered PAM specificities. Nature 2015, 523, 481–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vicencio, J.; Sánchez-Bolaños, C.; Moreno-Sánchez, I.; Brena, D.; Vejnar, C.E.; Kukhtar, D.; Ruiz-López, M.; Cots-Ponjoan, M.; Rubio, A.; Melero, N.R.; et al. Genome editing in animals with minimal PAM CRISPR-Cas9 enzymes. Nat. Commun. 2022, 13, 2601. [Google Scholar] [CrossRef]
- Waaijers, S.; Portegijs, V.; Kerver, J.; Lemmens, B.B.; Tijsterman, M.; van den Heuvel, S.; Boxem, M. CRISPR/Cas9-targeted mutagenesis in Caenorhabditis elegans. Genetics 2013, 195, 1187–1191. [Google Scholar] [CrossRef] [Green Version]
- Farboud, B.; Severson, A.F.; Meyer, B.J. Strategies for Efficient Genome Editing Using CRISPR-Cas9. Genetics 2019, 211, 431–457. [Google Scholar] [CrossRef] [Green Version]
- Mali, P.; Yang, L.; Esvelt, K.M.; Aach, J.; Guell, M.; DiCarlo, J.E.; Norville, J.E.; Church, G.M. RNA-guided human genome engineering via Cas9. Science 2013, 339, 823–826. [Google Scholar] [CrossRef] [Green Version]
- Richardson, C.D.; Kazane, K.R.; Feng, S.J.; Zelin, E.; Bray, N.L.; Schafer, A.J.; Floor, S.N.; Corn, J.E. CRISPR-Cas9 genome editing in human cells occurs via the Fanconi anemia pathway. Nat. Genet. 2018, 50, 1132–1139. [Google Scholar] [CrossRef]
- Moreno-Mateos, M.A.; Vejnar, C.E.; Beaudoin, J.D.; Fernandez, J.P.; Mis, E.K.; Khokha, M.K.; Giraldez, A.J. CRISPRscan: Designing highly efficient sgRNAs for CRISPR-Cas9 targeting in vivo. Nat. Methods 2015, 12, 982–988. [Google Scholar] [CrossRef]
- Naito, Y.; Hino, K.; Bono, H.; Ui-Tei, K. CRISPRdirect: Software for designing CRISPR/Cas guide RNA with reduced off-target sites. Bioinformatics 2015, 31, 1120–1123. [Google Scholar] [CrossRef] [Green Version]
- Concordet, P.J.; Haeussler, M. CRISPOR: Intuitive guide selection for CRISPR/Cas9 genome editing experiments and screens. Nucleic Acids Res. 2018, 46, W242–W245. [Google Scholar] [CrossRef] [Green Version]
- Hutter, H.; Ng, M.P.; Chen, N. GExplore: A web server for integrated queries of protein domains, gene expression and mutant phenotypes. BMC Genom. 2009, 10, 529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Li, M.; Feng, X.; Guang, S. Targeted Chromosomal Translocations and Essential Gene Knockout Using CRISPR/Cas9 Technology in Caenorhabditis elegans. Genetics 2015, 201, 1295–1306. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.P.; Li, X.L.; Li, G.H.; Chen, W.; Arakaki, C.; Botimer, G.D.; Baylink, D.; Zhang, L.; Wen, W.; Fu, Y.W.; et al. Efficient precise knockin with a double cut HDR donor after CRISPR/Cas9-mediated double-stranded DNA cleavage. Genome Biol. 2017, 18, 35. [Google Scholar] [CrossRef] [Green Version]
- Dokshin, G.A.; Ghanta, K.S.; Piscopo, K.M.; Mello, C.C. Robust Genome Editing with Short Single-Stranded and Long, Partially Single-Stranded DNA Donors in Caenorhabditis elegans. Genetics 2018, 210, 781–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renaud, J.B.; Boix, C.; Charpentier, M.; De Cian, A.; Cochennec, J.; Duvernois-Berthet, E.; Perrouault, L.; Tesson, L.; Edouard, J.; Thinard, R.; et al. Improved Genome Editing Efficiency and Flexibility Using Modified Oligonucleotides with TALEN and CRISPR-Cas9 Nucleases. Cell. Rep. 2016, 14, 2263–2272. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Mackley, V.A.; Rao, A.; Chong, A.T.; Dewitt, M.A.; Corn, J.E.; Murthy, N. Synthetically modified guide RNA and donor DNA are a versatile platform for CRISPR-Cas9 engineering. Elife 2017, 6, e25312. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Triana, J.A.; Tavhelidse, T.; Thumberger, T.; Thomas, I.; Wittbrodt, B.; Kellner, T.; Anlas, K.; Tsingos, E.; Wittbrodt, J. Efficient single-copy HDR by 5′ modified long dsDNA donors. Elife 2018, 7, e39468. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Guo, Y.; Tian, Q.; Lan, Y.; Yeh, H.; Zhang, M.; Tasan, I.; Jain, S.; Zhao, H. An efficient gene knock-in strategy using 5’-modified double-stranded DNA donors with short homology arms. Nat. Chem. Biol. 2020, 16, 387–390. [Google Scholar] [CrossRef]
- Ghanta, K.S.; Chen, Z.; Mir, A.; Dokshin, G.A.; Krishnamurthy, P.M.; Yoon, Y.; Gallant, J.; Xu, P.; Zhang, X.-O.; Ozturk, A.R.; et al. 5′-Modifications improve potency and efficacy of DNA donors for precision genome editing. eLife 2021, 10, e72216. [Google Scholar] [CrossRef] [PubMed]
- CChiu, H.; Schwartz, H.T.; Antoshechkin, I.; Sternberg, P.W. Transgene-free genome editing in Caenorhabditis elegans using CRISPR-Cas. Genetics 2013, 195, 1167–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paix, A.; Folkmann, A.; Seydoux, G. Precision genome editing using CRISPR-Cas9 and linear repair templates in C. elegans. Methods 2017, 121–122, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Park, H.; Liu, J.; Sternberg, P.W. An Efficient Genome Editing Strategy To Generate Putative Null Mutants in Caenorhabditis elegans Using CRISPR/Cas9. G3 (Bethesda) 2018, 8, 3607–3616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.M.; Tian, S.; Wang, S. An affordable plasmid miniprep suitable for proficient microinjection in Caenorhabditis elegans. 3 Biotech 2020, 10, 350. [Google Scholar] [CrossRef]
- Kim, H.M.; Zhang, X. Design of Repair Templates for CRISPR-Cas9-Triggered Homologous Recombination in Caenorhabditis elegans. In Springer Protocols Handbooks; Humana: New York, NY, USA, 2021. [Google Scholar]
- Ghanta, K.S.; Ishidate, T.; Mello, C.C. Microinjection for precision genome editing in Caenorhabditis elegans. STAR Protoc. 2021, 2, 100748. [Google Scholar] [CrossRef]
- Schwartz, M.L.; Davis, M.W.; Rich, M.S.; Jorgensen, E.M. High-efficiency CRISPR gene editing in C. elegans using Cas9 integrated into the genome. PLoS Genet. 2021, 17, e1009755. [Google Scholar] [CrossRef]
- Martin, C.J.; Calarco, J.A. Approaches for CRISPR/Cas9 Genome Editing in C. elegans. Methods Mol. Biol. 2022, 2468, 215–237. [Google Scholar] [CrossRef]
- Au, V.; Li-Leger, E.; Raymant, G.; Flibotte, S.; Chen, G.; Martin, K.; Fernando, L.; Doell, C.; Rosell, F.I.; Wang, S.; et al. CRISPR/Cas9 Methodology for the Generation of Knockout Deletions in Caenorhabditis elegans. G3 (Bethesda) 2019, 9, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Dickinson, D.J.; Goldstein, B. CRISPR-Based Methods for Caenorhabditis elegans Genome Engineering. Genetics 2016, 202, 885–901. [Google Scholar] [CrossRef]
- Lim, Y.H.; Wong, F.T.; Yeo, W.L.; Ching, K.C.; Lim, Y.W.; Heng, E.; Chen, S.; Tsai, D.J.; Lauderdale, T.L.; Shia, K.S.; et al. Auroramycin: A Potent Antibiotic from Streptomyces roseosporus by CRISPR-Cas9 Activation. Chembiochem 2018, 19, 1716–1719. [Google Scholar] [CrossRef]
- Meneely, P.M.; Dahlberg, C.L.; Rose, J.K. Working with Worms: Caenorhabditis elegans as a Model Organism. Curr. Protoc. Essent. Lab. Tech. 2019, 19, e35. [Google Scholar] [CrossRef] [Green Version]
- Corsi, A.K. A biochemist’s guide to Caenorhabditis elegans. Anal. Biochem. 2006, 359, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, D.; Schwarz, E.M.; Thomas, C.G.; Felde, R.L.; Korf, I.F.; Cutter, A.D.; Schartner, C.M.; Ralston, E.J.; Meyer, B.J.; Haag, E.S. Rapid genome shrinkage in a self-fertile nematode reveals sperm competition proteins. Science 2018, 359, 55–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LLok, J.B. CRISPR/Cas9 Mutagenesis and Expression of Dominant Mutant Transgenes as Functional Genomic Approaches in Parasitic Nematodes. Front. Genet. 2019, 10, 656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gang, S.S.; Castelletto, M.L.; Bryant, A.S.; Yang, E.; Mancuso, N.; Lopez, J.B.; Pellegrini, M.; Hallem, E.A. Targeted mutagenesis in a human-parasitic nematode. PLoS Pathog. 2017, 13, e1006675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, S.; Pathak, P.; Shao, H.; Lok, J.B.; Pires-daSilva, A. Liposome-based transfection enhances RNAi and CRISPR-mediated mutagenesis in non-model nematode systems. Sci. Rep. 2019, 9, 483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Halloran, D.M. Genome aware CRISPR gRNA target prediction for parasitic nematodes. Mol. Biochem. Parasitol. 2019, 227, 25–28. [Google Scholar] [CrossRef]
- Cohen, S.; Sternberg, P. Genome editing of Caenorhabditis briggsae using CRISPR/Cas9 co-conversion marker dpy-10. MicroPubl. Biol. 2019, 2019. [Google Scholar] [CrossRef]
- Culp, E.; Richman, C.; Sharanya, D.; Jhaveri, N.; van den Berg, W.; Gupta, B.P. Genome editing in the nematode Caenorhabditis briggsae using the CRISPR/Cas9 system. Biol. Methods Protoc. 2020, 5, bpaa003. [Google Scholar] [CrossRef]
- Yan, W.X.; Hunnewell, P.; Alfonse, L.E.; Carte, J.M.; Keston-Smith, E.; Sothiselvam, S.; Garrity, A.J.; Chong, S.; Makarova, K.S.; Koonin, E.V.; et al. Functionally diverse type V CRISPR-Cas systems. Science 2019, 363, 88–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abudayyeh, O.O.; Gootenberg, J.S.; Essletzbichler, P.; Han, S.; Joung, J.; Belanto, J.J.; Verdine, V.; Cox, D.B.T.; Kellner, M.J.; Regev, A.; et al. RNA targeting with CRISPR-Cas13. Nature 2017, 550, 280–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, J.; Kang, D.D.; Dong, Y. Harnessing lipid nanoparticles for efficient CRISPR delivery. Biomater. Sci. 2021, 9, 6001–6011. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Wu, Q.; Wei, Y.; Gong, C. CRISPR-Cas9 Delivery by Artificial Virus (RRPHC). Methods Mol. Biol. 2019, 1961, 81–91. [Google Scholar] [CrossRef]
- Kaletta, T.; Hengartner, M.O. Finding function in novel targets: C. elegans as a model organism. Nat. Rev. Drug Discov. 2006, 5, 387–398. [Google Scholar] [CrossRef]
- Leung, M.C.; Williams, P.L.; Benedetto, A.; Au, C.; Helmcke, K.J.; Aschner, M.; Meyer, J.N. Caenorhabditis elegans: An emerging model in biomedical and environmental toxicology. Toxicol. Sci. 2008, 106, 5–28. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.-M.; Hong, Y.; Chen, J. A Decade of CRISPR-Cas Gnome Editing in C. elegans. Int. J. Mol. Sci. 2022, 23, 15863. https://doi.org/10.3390/ijms232415863
Kim H-M, Hong Y, Chen J. A Decade of CRISPR-Cas Gnome Editing in C. elegans. International Journal of Molecular Sciences. 2022; 23(24):15863. https://doi.org/10.3390/ijms232415863
Chicago/Turabian StyleKim, Hyun-Min, Yebin Hong, and Jiani Chen. 2022. "A Decade of CRISPR-Cas Gnome Editing in C. elegans" International Journal of Molecular Sciences 23, no. 24: 15863. https://doi.org/10.3390/ijms232415863
APA StyleKim, H. -M., Hong, Y., & Chen, J. (2022). A Decade of CRISPR-Cas Gnome Editing in C. elegans. International Journal of Molecular Sciences, 23(24), 15863. https://doi.org/10.3390/ijms232415863