

Effects of Light Intensity on Physiological Characteristics and Expression of Genes in Coumarin Biosynthetic Pathway of Angelica dahurica

Abstract
:1. Introduction
2. Results
2.1. Effect of Light Intensity on Physiological Characteristics of Leaves
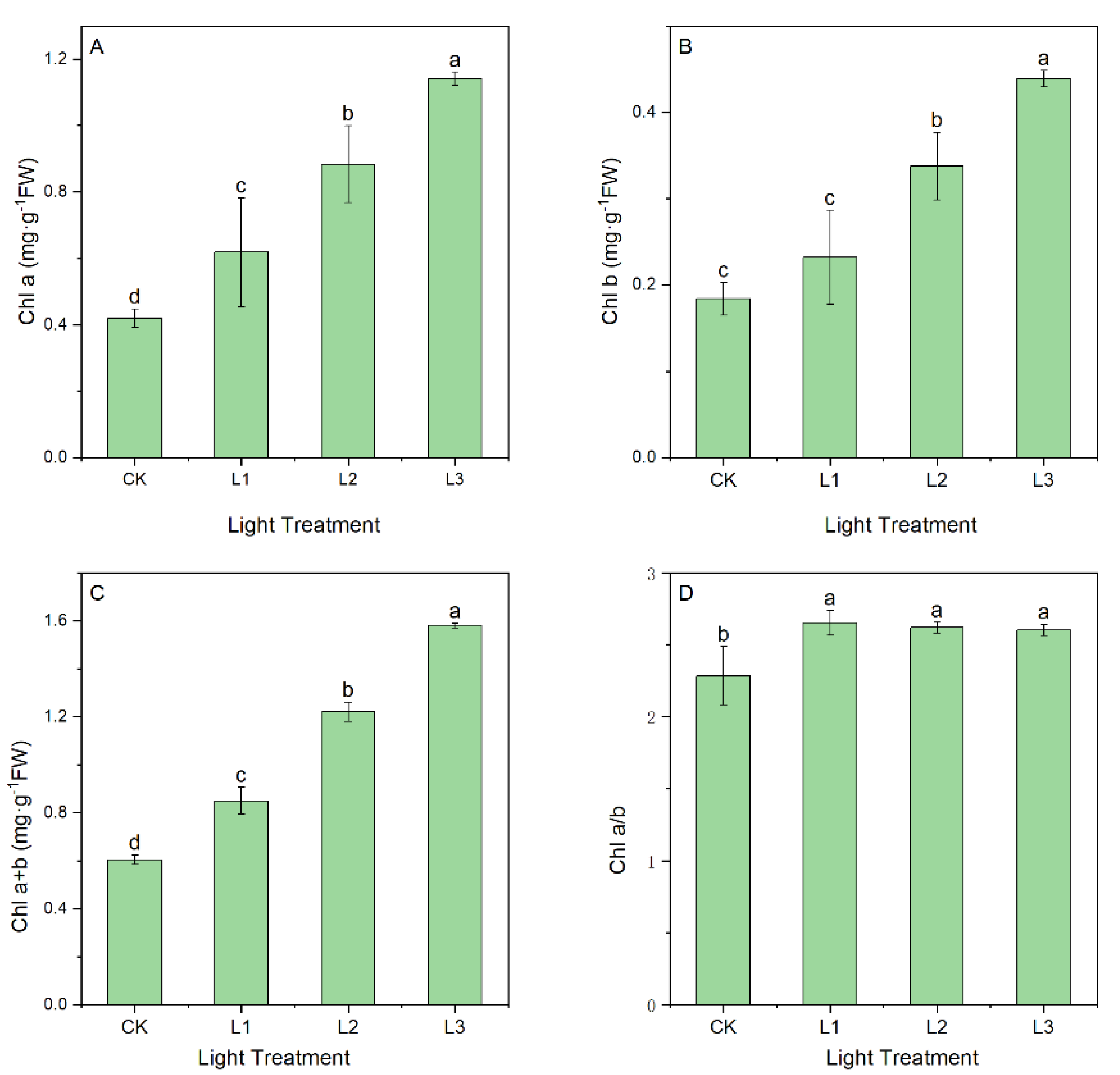
2.1.1. Effect of Light Intensity on Photosynthetic Pigments
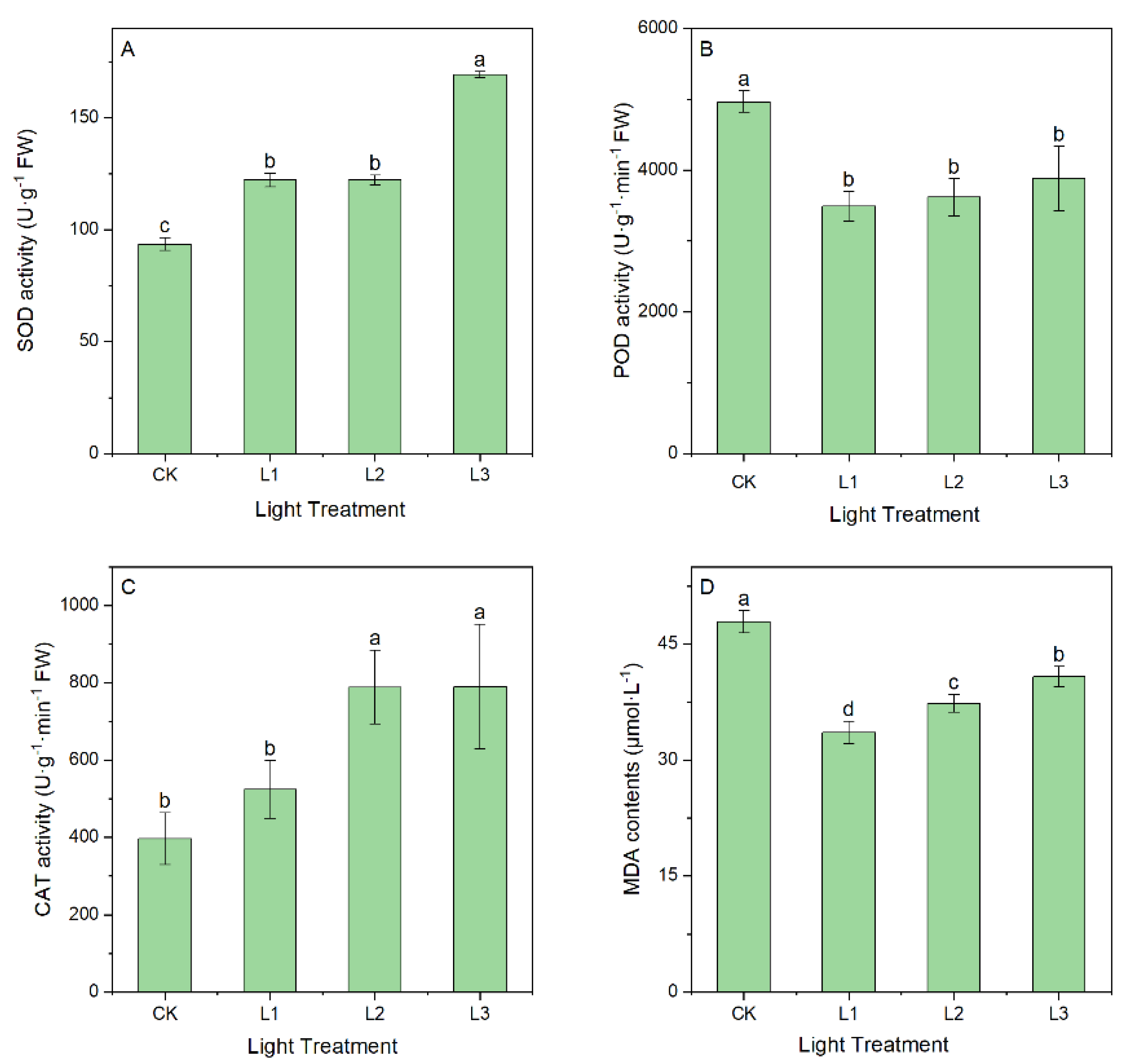
2.1.2. Effect of Light Intensity on Antioxidant Enzyme Activity and MDA Content
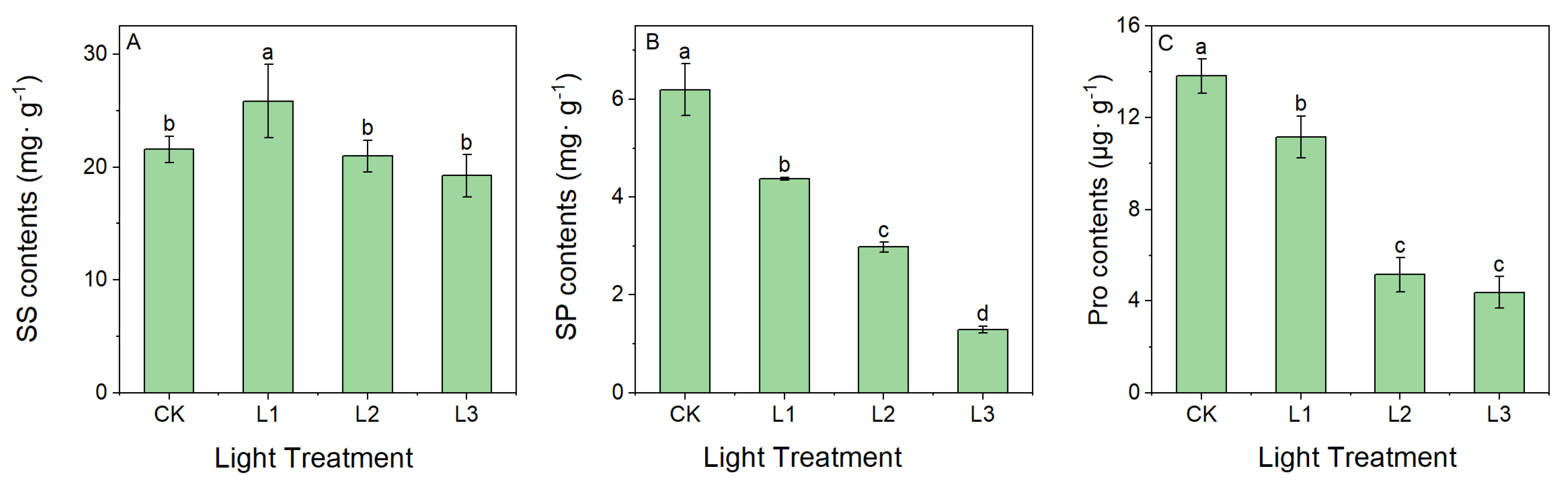
2.1.3. Effect of Light Intensity on the Content of Osmoregulatory Substances
2.2. RNA-Seq of Root Tissue under Different Light Intensities
2.2.1. De Novo Assembly
2.2.2. Functional Annotation of Unigenes
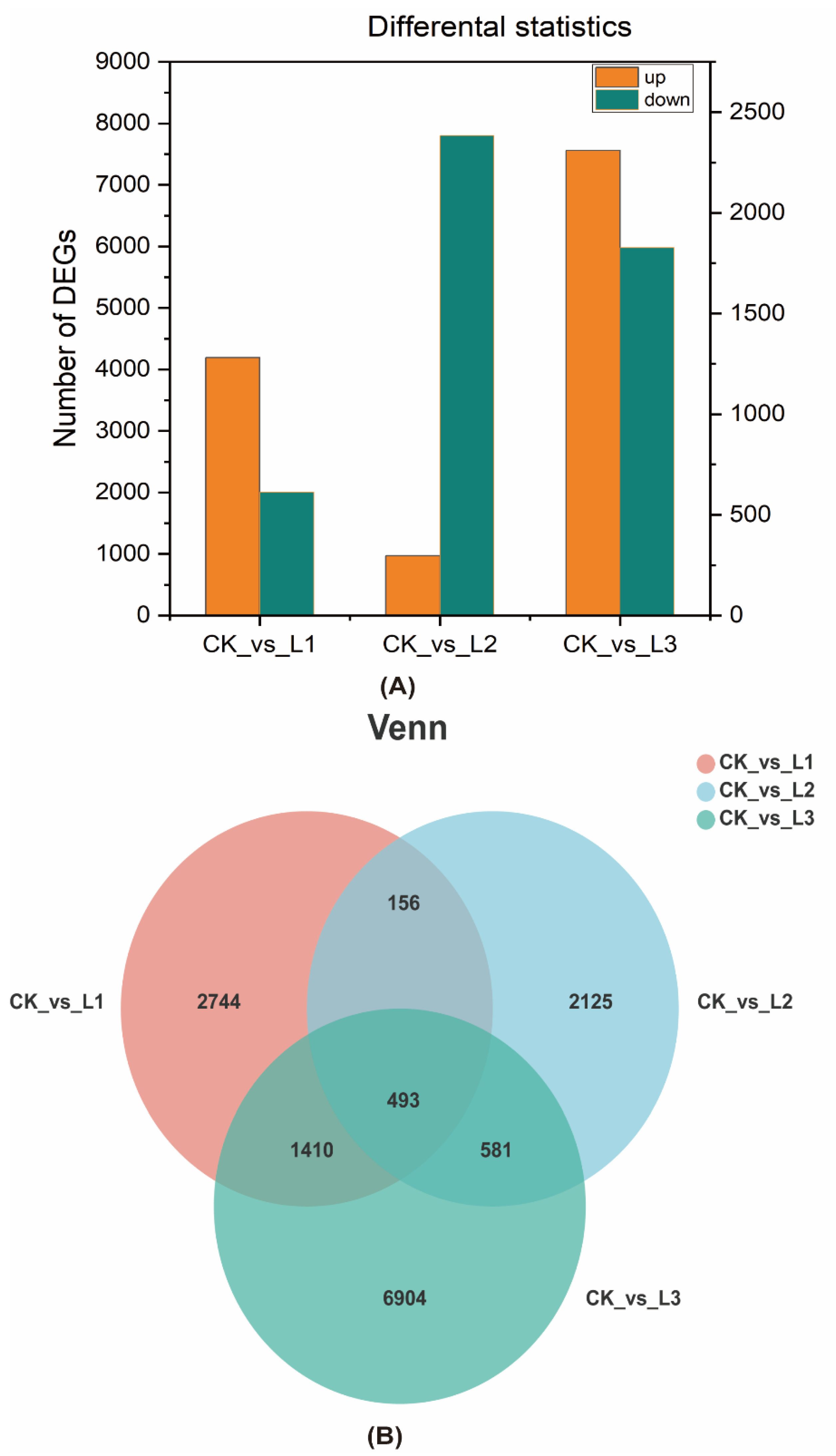
2.2.3. Analysis of Differentially Expressed Genes
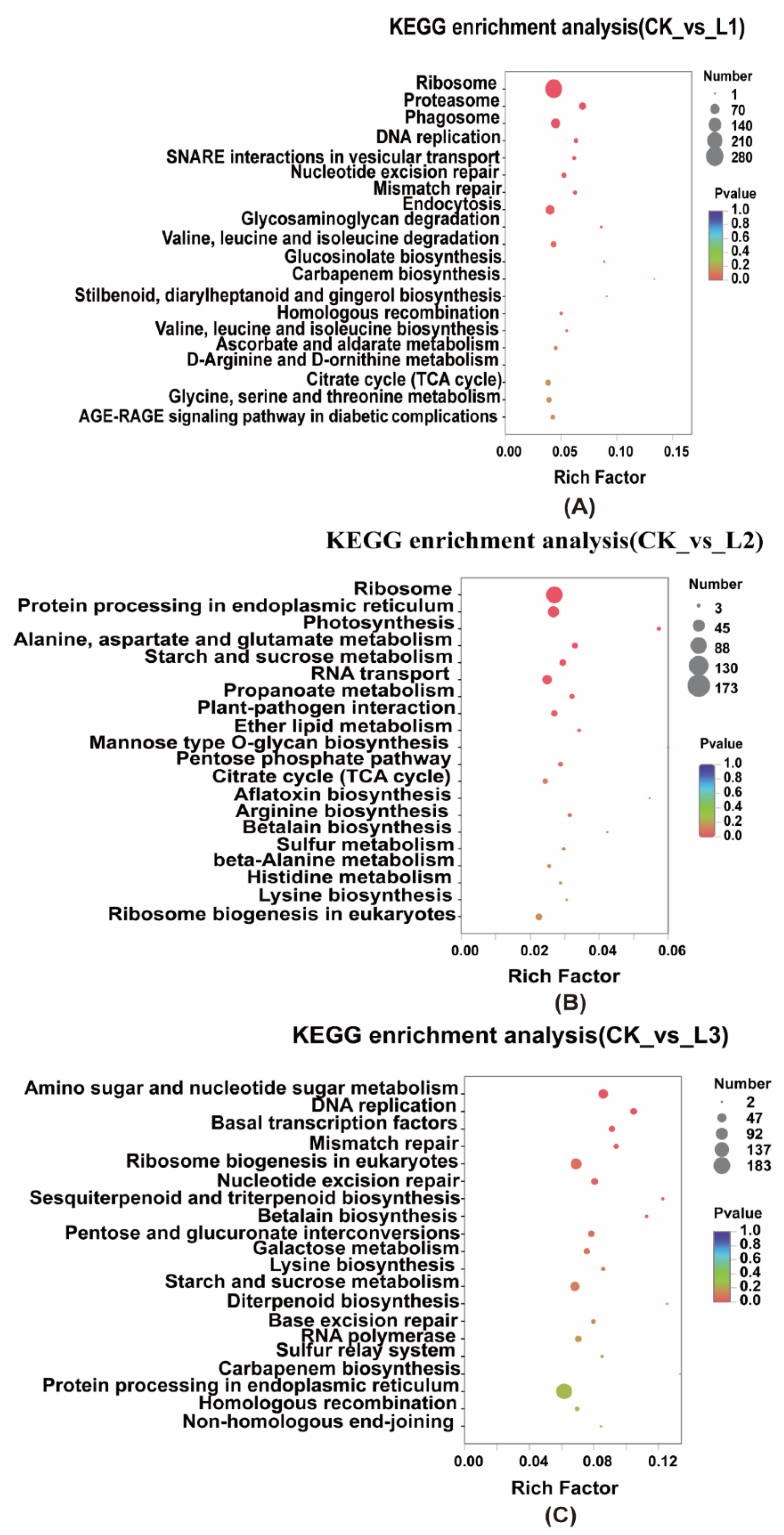
2.2.4. KEGG Enrichment of Differentially Expressed Genes
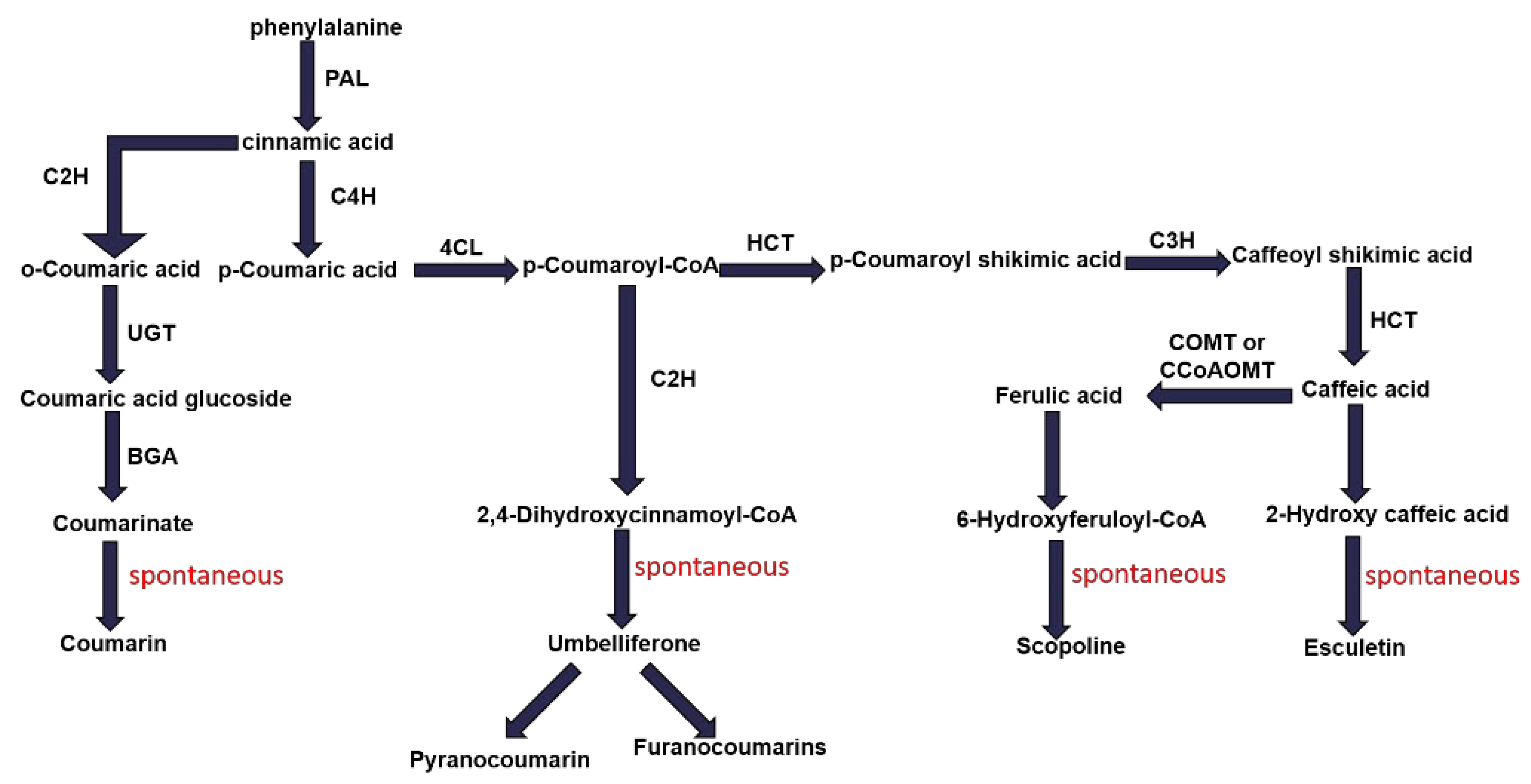
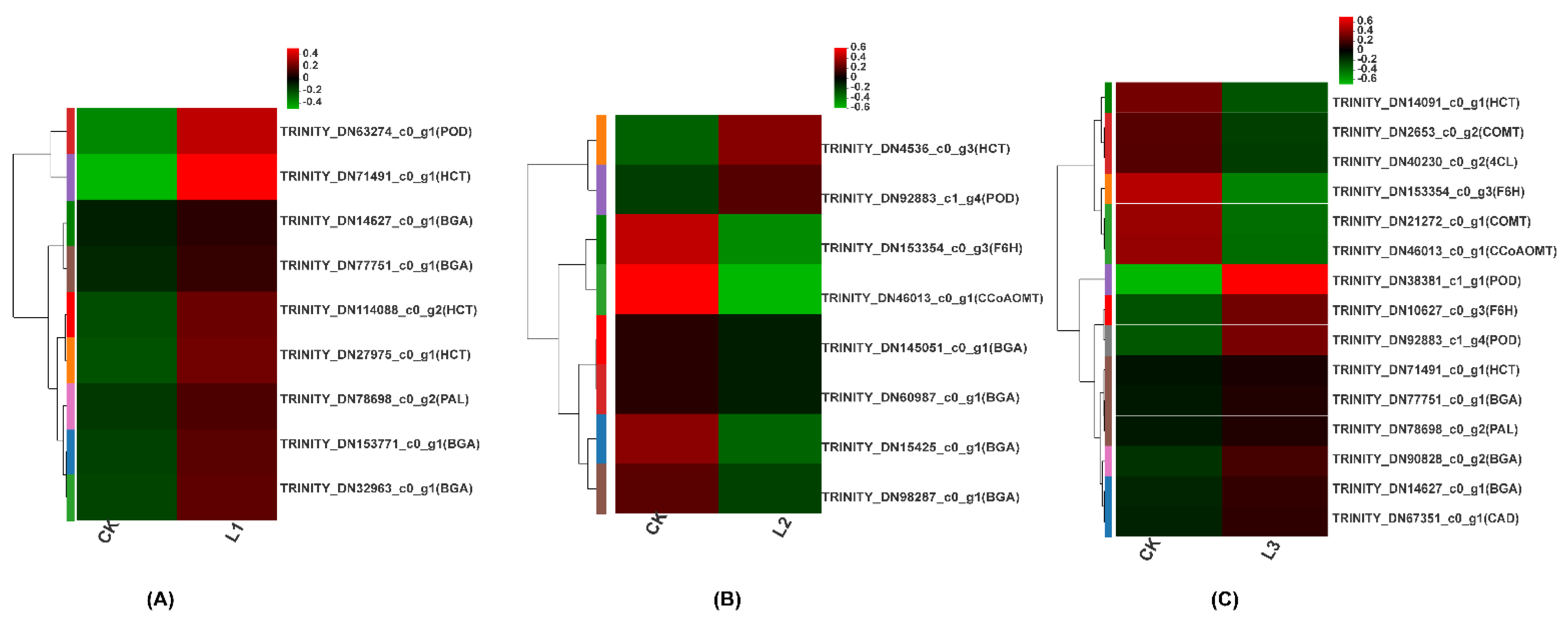
2.2.5. Coumarin Biosynthesis Pathway
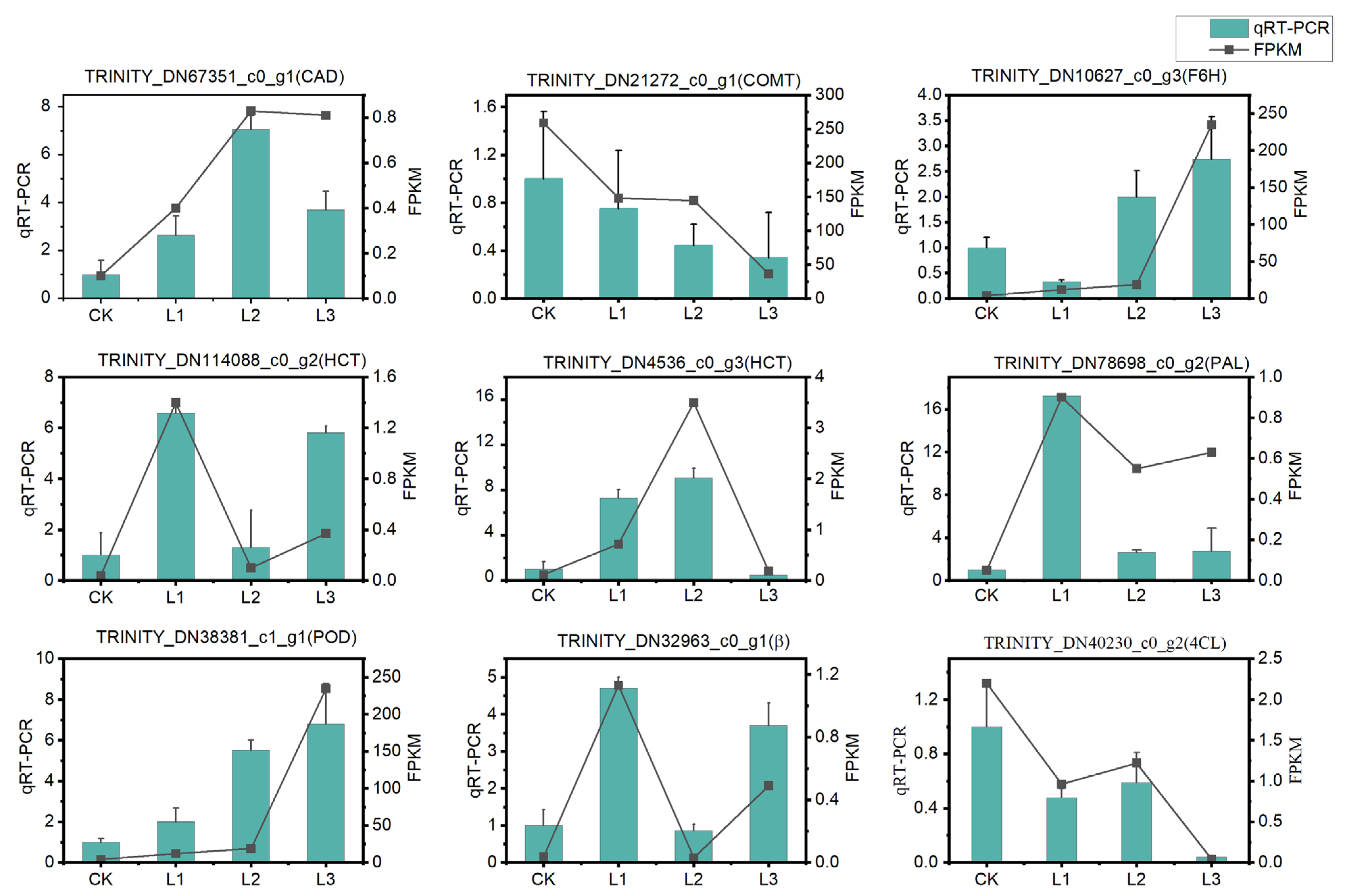
2.2.6. qRT-PCR Validation
3. Materials and Methods
3.1. Plant Materials and Experimental Design
3.2. Measurement of Physiological Indicators
3.3. cDNA Library Construction and Functional Annotation
3.4. Expression Analysis of Differentially Expressed Genes
3.5. Real-Time Quantitative PCR Validation
3.6. Statistical Analysis
4. Discussion
4.1. Effect of Different Light Intensities on Physiological Characteristics of Leaves
4.2. Effect of Different Light Intensities on the Transcriptome of Roots
4.3. Effect of Different Light Intensities on Coumarin Synthesis Pathway of Roots
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, J.J.; Zhu, L.; Zhang, X.; Zhou, J. Photosynthetic performance and growth responses of Liriope muscari (Decne.) L.H. Bailey (Asparagaceae) planted within poplar forests having different canopy densities. BMC Ecol. 2020, 20, 25. [Google Scholar] [CrossRef] [PubMed]
- Pawłowska, B.; Żupnik, M.; Szewczyk-Taranek, B.; Cioć, M. Impact of LED light sources on morphogenesis and levels of photosynthetic pigments in Gerbera jamesonii grown in vitro. Hortic. Environ. Biotechnol. 2018, 59, 115–123. [Google Scholar] [CrossRef]
- Gabr, A.M.M.; Ebrahim, H.S.; El-Ashry, A.A.E.-L.; El-Bahr, M.K. Importance of Artificial Environment Conditions on Plant Biotechnology, Plant Growth, and Secondary Metabolites; IGI Global: Chocolate avenue Hershey PA, USA, 2021; pp. 292–319. [Google Scholar]
- Shin, K.S.; Murthy, H.N.; Heo, J.W.; Hahn, E.J.; Paek, K.Y. The effect of light quality on the growth and development of in vitro cultured Doritaenopsis plants. Acta Physiol. Plant. 2008, 30, 339–343. [Google Scholar] [CrossRef]
- Fu, X.; Chen, Y.; Mei, X.; Katsuno, T.; Kobayashi, E.; Dong, F.; Watanabe, N.; Yang, Z. Regulation of formation of volatile compounds of tea (Camellia sinensis) leaves by single light wavelength. Sci. Rep. 2015, 5, 16858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, S.; Fu, X.; Mei, X.; Zhou, Y.; Du, B.; Watanabe, N.; Yang, Z. Regulation of biosynthesis and emission of volatile phenylpropanoids/benzenoids in petunia× hybrida flowers by multi-factors of circadian clock, light, and temperature. Plant Physiol. Biochem. 2016, 107, 1–8. [Google Scholar] [CrossRef]
- Li, Y.; Kong, D.; Fu, Y.; Sussman, M.R.; Wu, H. The effect of developmental and environmental factors on secondary metabolites in medicinal plants. Plant Physiol. Biochem. 2020, 148, 80–89. [Google Scholar] [CrossRef]
- Deng, B.; Fang, S.; Shang, X.; Fu, X.; Li, Y. Influence of provenance and shade on biomass production and triterpenoid accumulation in Cyclocarya paliurus. Agrofor. Syst. 2019, 93, 483–492. [Google Scholar] [CrossRef]
- Ghasemzadeh, A.; Jaafar, H.Z.E.; Rahmat, A.; Wahab, P.E.M.; Halim, M.R.A. Effect of Different Light Intensities on Total Phenolics and Flavonoids Synthesis and Anti-oxidant Activities in Young Ginger Varieties (Zingiber officinale Roscoe). Int. J. Mol. Sci. 2010, 11, 3885–3897. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Wang, G.; Cao, F.; Zhu, C.; Wang, G.; El-Kassaby, Y.A. Light intensity affects the growth and flavonol biosynthesis of Ginkgo (Ginkgo biloba L.). New For. 2014, 45, 765–776. [Google Scholar] [CrossRef]
- Choi, I.; Lim, H.H.; Song, Y.K.; Lee, J.W.; Kim, Y.S.; Ko, I.G.; Kim, K.J.; Shin, M.S.; Kim, K.H.; Kim, C.J. Analgesic and anti-inflammatory effect of the aqueous extract of root of Angelica Dahurica. Orient. Pharm. Exp. Med. 2008, 7, 527–533. [Google Scholar] [CrossRef]
- Kang, S.W.; Kim, C.Y.; Song, D.g.; Pan, C.H.; Cha, K.H.; Lee, D.-U.; Um, B.-H. Rapid Identification of Furanocoumarins in Angelica dahurica using the Online LC-MMR-MS and their Nitric Oxide Inhibitory Activity in RAW 264.7 cells. Phytochem. Anal. 2010, 21, 322–327. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, S.; Shan, C.; Shi, Y.; Wu, H.; Wu, J.; Peng, D. De novo transcriptome assembly of Angelica dahurica and characterization of coumarin biosynthesis pathway genes. Gene 2021, 791, 145713. [Google Scholar] [CrossRef] [PubMed]
- Okada, R.; Abe, H.; Okuyama, T.; Nishidono, Y.; Ishii, T.; Sato, T.; Shirako, S.; Tanaka, K.; Ikeya, Y.; Nishizawa, M. Comparison of the anti-inflammatory activities of furanocoumarins from the roots of Angelica dahurica. Bioact. Compd. Health Dis. 2021, 4, 287–300. [Google Scholar] [CrossRef]
- Wang, T.t.; Jin, H.; Li, Q.; Cheng, W.M.; Hu, Q.q.; Chen, X.h.; Bi, K.s. Isolation and simultaneous determination of coumarin compounds in Radix Angelica dahurica. Chromatographia 2007, 65, 477–481. [Google Scholar] [CrossRef]
- Zhao, H.; Feng, Y.-L.; Wang, M.; Wang, J.-J.; Liu, T.; Yu, J. The Angelica dahurica: A Review of Traditional Uses, Phytochemistry and Pharmacology. Front. Pharmacol. 2022, 13, 2367. [Google Scholar] [CrossRef]
- Xie, Y.; Zhao, W.; Zhou, T.; Fan, G.; Wu, Y. An efficient strategy based on MAE, HPLC-DAD-ESI-MS/MS and 2D-prep-HPLC-DAD for the rapid extraction, separation, identification and purification of five active coumarin components from radix angelicae dahuricae. Phytochem. Anal. 2010, 21, 473–482. [Google Scholar] [CrossRef]
- Piao, X.L.; Park, I.H.; Baek, S.H.; Kim, H.Y.; Park, M.K.; Park, J.H. Antioxidative activity of furanocoumarins isolated from Angelicae dahuricae. J. Ethnopharmacol. 2004, 93, 243–246. [Google Scholar] [CrossRef]
- Pang, Q.q.; Li, T.; Liu, L.x.; Shi, D.f.; Yao, X.s.; Li, H.b.; Yu, Y. Systematically identifying the anti-inflammatory constituents of Cimicifuga dahurica by UPLC-Q/TOF-MS combined with network pharmacology analysis. Biomed. Chromatogr. 2021, 35, 5177. [Google Scholar] [CrossRef]
- Dong, X.d.; Liu, Y.n.; Zhao, Y.; Liu, A.j.; Ji, H.y.; Yu, J. Structural characterization of a water-soluble polysaccharide from Angelica dahurica and its antitumor activity in H22 tumor-bearing mice. Int. J. Biol. Macromol. 2021, 193, 219–227. [Google Scholar] [CrossRef]
- Wang, Z.; Xiao, S.; Wang, Y.; Liu, J.; Ma, H.; Wang, Y.; Tian, Y.; Hou, W. Effects of light irradiation on essential oil biosynthesis in the medicinal plant Asarum heterotropoides Fr. Schmidt var. mandshuricum (Maxim) Kitag. PLoS ONE 2020, 15, e0237952. [Google Scholar] [CrossRef]
- Vogt, T. Phenylpropanoid biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Huang, J.; Hua, S.; Zhang, Y.; Zhang, Y.; Li, T.; Dong, L.; Gao, Q.; Fu, X. The ethnopharmacology, phytochemistry and pharmacology of Angelica Biserrata—A review. J. Ethnopharmacol. 2019, 231, 152–169. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.-L.; Zeng, R.; Gu, C.-M.; Qu, Y.; Huang, L.-F. Angelica sinensis in China-A review of botanical profile, ethnopharmacology, phytochemistry and chemical analysis. J. Ethnopharmacol. 2016, 190, 116–141. [Google Scholar] [CrossRef] [PubMed]
- Li, H. Principles and Experimental Techniques of Plant Physiology and Biochemistry; Higher Education Press: Beijing, China, 2000; Volume 5. [Google Scholar]
- Xu, L.; Yang, P.; Feng, Y.; Xu, H.; Cao, Y.; Tang, Y.; Yuan, S.; Liu, X.; Ming, J. Spatiotemporal Transcriptome Analysis Provides Insights into Bicolor Tepal Development in Lilium “Tiny Padhye”. Front. Plant Sci. 2017, 8, 398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Smith-Unna, R.; Boursnell, C.; Patro, R.; Hibberd, J.M.; Kelly, S. TransRate: Reference-free quality assessment of de novo transcriptome assemblies. Genome Res. 2016, 26, 1134–1144. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Boratyn, G.M.; Thierry-Mieg, J.; Thierry-Mieg, D.; Busby, B.; Madden, T.L. Magic-BLAST, an accurate RNA-seq aligner for long and short reads. BMC Bioinform. 2019, 20, 405. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Lin, Y.; Xue, Y.; Mao, M.; Zhou, X.; Hu, H.; Liu, J.; Feng, L.; Zhang, H.; Luo, J.; et al. Light Intensity Affects the Coloration and Structure of Chimeric Leaves of Ananas comosus var. bracteatus. Phyton Int. J. Exp. Bot. 2022, 91, 333–348. [Google Scholar] [CrossRef]
- Ma, X.; Song, L.; Yu, W.; Hu, Y.; Liu, Y.; Wu, J.; Ying, Y. Growth, physiological, and biochemical responses of Camptotheca acuminata seedlings to different light environments. Front. Plant Sci. 2015, 6, 321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertamini, M.; Muthuchelian, K.; Nedunchezhian, N. Shade effect alters leaf pigments and photosynthetic responses in Norway spruce (Picea abies L.) grown under field conditions. Photosynthetica 2006, 44, 227–234. [Google Scholar] [CrossRef]
- Lee, K.C.; Park, W.G. Physiological Response and Growth Performance of Parasenecio firmus under Different Shading Treatments. Korean J. Agric. For. Meteorol. 2012, 14, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Alscher, R.G.; Erturk, N.; Heath, L.S. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J. Exp. Bot. 2002, 53, 1331–1341. [Google Scholar] [CrossRef]
- Bowler, C.; Montagu, M.V.; Inze, D. Superoxide Dismutase and Stress Tolerance. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1992, 43, 83–116. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Zhu, H.; Li, X.; Zhai, W.; Liu, Y.; Gao, Q.; Liu, J.; Ren, L.; Chen, H.; Zhu, Y. Effects of low light on photosynthetic properties, antioxidant enzyme activity, and anthocyanin accumulation in purple pak-choi (Brassica campestris ssp. Chinensis Makino). PLoS ONE 2017, 12, e0179305. [Google Scholar] [CrossRef]
- Deng, Y.; Jia, X.; Sun, X.; Liang, L.; Su, J. Comparison of jasmine antioxidant system responses to different degrees and durations of shade. Acta Physiol. Plant. 2018, 40, 41. [Google Scholar] [CrossRef]
- Liu, S.-l.; Yang, R.-j.; Ren, B.; Wang, M.-h.; Ma, M.-d. Differences in photosynthetic capacity, chlorophyll fluorescence, and antioxidant system between invasive Alnus formosana and its native congener in response to different irradiance levels. Botany 2016, 94, 1087–1101. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.; Khalid, M.; Jan, F.; Saeed ur, R.; Jiang, X.; Yu, X. Effects of light-regulation and intensity on the growth, physiological and biochemical properties of Aralia elata (miq.) seedlings. S. Afr. J. Bot. 2019, 121, 456–462. [Google Scholar] [CrossRef]
- Morsy, M.R.; Jouve, L.; Hausman, J.-F.; Hoffmann, L.; Stewart, J.M. Alteration of oxidative and carbohydrate metabolism under abiotic stress in two rice (Oryza sativa L.) genotypes contrasting in chilling tolerance. J. Plant Physiol. 2007, 164, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Iqbal, N.; Rahman, T.; Liu, T.; Brestic, M.; Safdar, M.E.; Asghar, M.A.; Farooq, M.U.; Shafiq, I.; Ali, A.; et al. Shade effect on carbohydrates dynamics and stem strength of soybean genotypes. Environ. Exp. Bot. 2019, 162, 374–382. [Google Scholar] [CrossRef]
- Verheul, M. Effects of plant density, leaf removal and light intensity on tomato quality and yield. Acta Hortic. 2012, 956, 365–372. [Google Scholar] [CrossRef]
- Liu, L.; Zuo, Z.T.; Xu, F.R.; Wang, Y.Z. Study on Quality Response to Environmental Factors and Geographical Traceability of Wild Gentiana rigescens Franch. Front. Plant Sci. 2020, 11, 1128. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, T.; Luo, J.; Zhang, Q.; Xu, S.; Han, C.; Xu, J.; Chen, M.; Chen, Y.; Kong, L. Integration of a Decrescent Transcriptome and Metabolomics Dataset of Peucedanum praeruptorum to Investigate the CYP450 and MDR Genes Involved in Coumarins Biosynthesis and Transport. Front. Plant Sci. 2015, 6, 996. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Zhang, Z.; Chen, S.; Ma, L.; Wang, H.; Dong, R.; Wang, Y.; Liu, Z. Global transcriptome profiling analysis reveals insight into saliva-responsive genes in alfalfa. Plant Cell Rep. 2016, 35, 561–571. [Google Scholar] [CrossRef]
- Luo, K.; Wu, F.; Zhang, D.; Dong, R.; Fan, Z.; Zhang, R.; Yan, Z.; Wang, Y.; Zhang, J. Transcriptomic profiling of Melilotus albus near-isogenic lines contrasting for coumarin content. Sci. Rep. 2017, 7, 4577. [Google Scholar] [CrossRef]
- Naoumkina, M.A.; Zhao, Q.; Gallego-Giraldo, L.; Dai, X.; Zhao, P.X.; Dixon, R.A. Genome-wide analysis of phenylpropanoid defence pathways. Mol. Plant Pathol. 2010, 11, 829–846. [Google Scholar] [CrossRef] [PubMed]
- Fraser, C.M.; Chapple, C. The phenylpropanoid pathway in Arabidopsis. Arab. Book 2011, 9, e0152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Q.; Chen, S.; Wu, Y.; He, Y.; Feng, J.; Yang, Z.; Lin, W.; Zheng, G.; Li, Y.; Chen, H. Comparative Transcriptome Analyses of Gene Response to Different Light Conditions of Camellia oleifera Leaf Using Illumina and Single-Molecule Real-Time-Based RNA-Sequencing. Forests 2020, 11, 91. [Google Scholar] [CrossRef] [Green Version]
- Turgut-Kara, N. Comparative phylogenetic analysis of phenylpropanoid metabolism genes of legume plants. Plant Omics 2015, 8, 55–61. [Google Scholar]
- Deluc, L.; Barrieu, F.o.; Marchive, C.; Lauvergeat, V.; Decendit, A.; Richard, T.; Carde, J.-P.; Mérillon, J.-M.; Hamdi, S.d. Characterization of a Grapevine R2R3-MYB Transcription Factor That Regulates the Phenylpropanoid Pathway. Plant Physiol. 2005, 140, 499–511. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Pan, D.; Liang, M.; Abubakar, Y.S.; Li, J.; Lin, J.; Chen, S.; Chen, W. Regulation of Anthocyanin Biosynthesis in Purple Leaves of Zijuan Tea (Camellia sinensis var. kitamura). Int. J. Mol. Sci. 2017, 18, 833. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Sun, X.; Yuan, Q.; Yan, Y. Combinatorial biosynthesis of plant-specific coumarins in bacteria. Metab. Eng. 2013, 18, 69–77. [Google Scholar] [CrossRef]
- Yang, S.M.; Shim, G.Y.; Kim, B.G.; Ahn, J.H. Biological synthesis of coumarins in Escherichia coli. Microb. Cell Factories 2015, 14, 65. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Wang, N.; Sui, Z.; Huang, C.; Zeng, Z.; Kong, L. The Molecular and Structural Basis of O-methylation Reaction in Coumarin Biosynthesis in Peucedanum praeruptorum Dunn. Int. J. Mol. Sci. 2019, 20, 1533. [Google Scholar] [CrossRef] [Green Version]
- Kai, K.; Shimizu, B.-i.; Mizutani, M.; Sakata, K. Analysis of Coumarin Biosynthesis Pathway in Arabidopsis thaliana. Plant Cell Physiol. Suppl. 2005, 2005, 864. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviations | Terminology | Abbreviations | Terminology |
|---|---|---|---|
| A. dahurica | Angelica dahurica (Fisch.ex Hoffm.) Benth. et Hook. f. | Chl b | Chlorophyll b |
| PAL | Phenylalanine ammonia-lyase | Chl a + b | Chlorophyll a + b |
| C4H | Cinnamic acid-4-hydroxylase | Chl a/b | Chlorophyll a/b |
| 4CL | 4-coumaric acid-CoA ligase | MDA | Malondialdehyde concentration |
| HCT | Hydroxycinnamic acid acyltransferase | SOD | Superoxide dismutase |
| C3H | P-coumaric acid 3-hydroxylase | POD | peroxidase |
| COMT | Caffeic acid O-methyltransferase | CAT | Catalase |
| CCoAOMT | Caffeoyl CoA O-methyltransferase | Pro | Free proline |
| BGA | β-glucosidase | SP | Soluble protein |
| F6H | Feruloyl-CoA 6-hydroxylase | SS | Soluble sugar |
| CAD | Coniferyl-aldehyde dehydrogenase | GO | Gene Ontology |
| C2H | Cinnamic acid 2-hydroxylase | KEGG | Kyoto Encyclopedia of Genes and Genomes |
| qRT-PCR | Real-time quantitative Polymerase Chain Reaction | eggNOG | Non-supervised orthologous groups |
| RNA-seq | RNA-sequencing | NR | Non-redundant protein sequence |
| RSEM | RNA-Seq by Expectation-Maximization | Swiss-Prot | RefSeq non-redundant proteins |
| Chl a | Chlorophyll a | Pfam | Protein families |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.; Zhai, Y.; Huang, Y.; Huang, Y.; Liu, K.; Zhang, J.; Zhou, J. Effects of Light Intensity on Physiological Characteristics and Expression of Genes in Coumarin Biosynthetic Pathway of Angelica dahurica. Int. J. Mol. Sci. 2022, 23, 15912. https://doi.org/10.3390/ijms232415912
Huang Y, Zhai Y, Huang Y, Huang Y, Liu K, Zhang J, Zhou J. Effects of Light Intensity on Physiological Characteristics and Expression of Genes in Coumarin Biosynthetic Pathway of Angelica dahurica. International Journal of Molecular Sciences. 2022; 23(24):15912. https://doi.org/10.3390/ijms232415912
Chicago/Turabian StyleHuang, Yongjie, Yongnan Zhai, Ya Huang, Yufeng Huang, Kun Liu, Jie Zhang, and Jihai Zhou. 2022. "Effects of Light Intensity on Physiological Characteristics and Expression of Genes in Coumarin Biosynthetic Pathway of Angelica dahurica" International Journal of Molecular Sciences 23, no. 24: 15912. https://doi.org/10.3390/ijms232415912
APA StyleHuang, Y., Zhai, Y., Huang, Y., Huang, Y., Liu, K., Zhang, J., & Zhou, J. (2022). Effects of Light Intensity on Physiological Characteristics and Expression of Genes in Coumarin Biosynthetic Pathway of Angelica dahurica. International Journal of Molecular Sciences, 23(24), 15912. https://doi.org/10.3390/ijms232415912