
A Novel 3-O-rhamnoside: 2″-O-xylosyltransferase Responsible for Terminal Modification of Prenylflavonol Glycosides in Epimedium pubescens Maxim.

 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
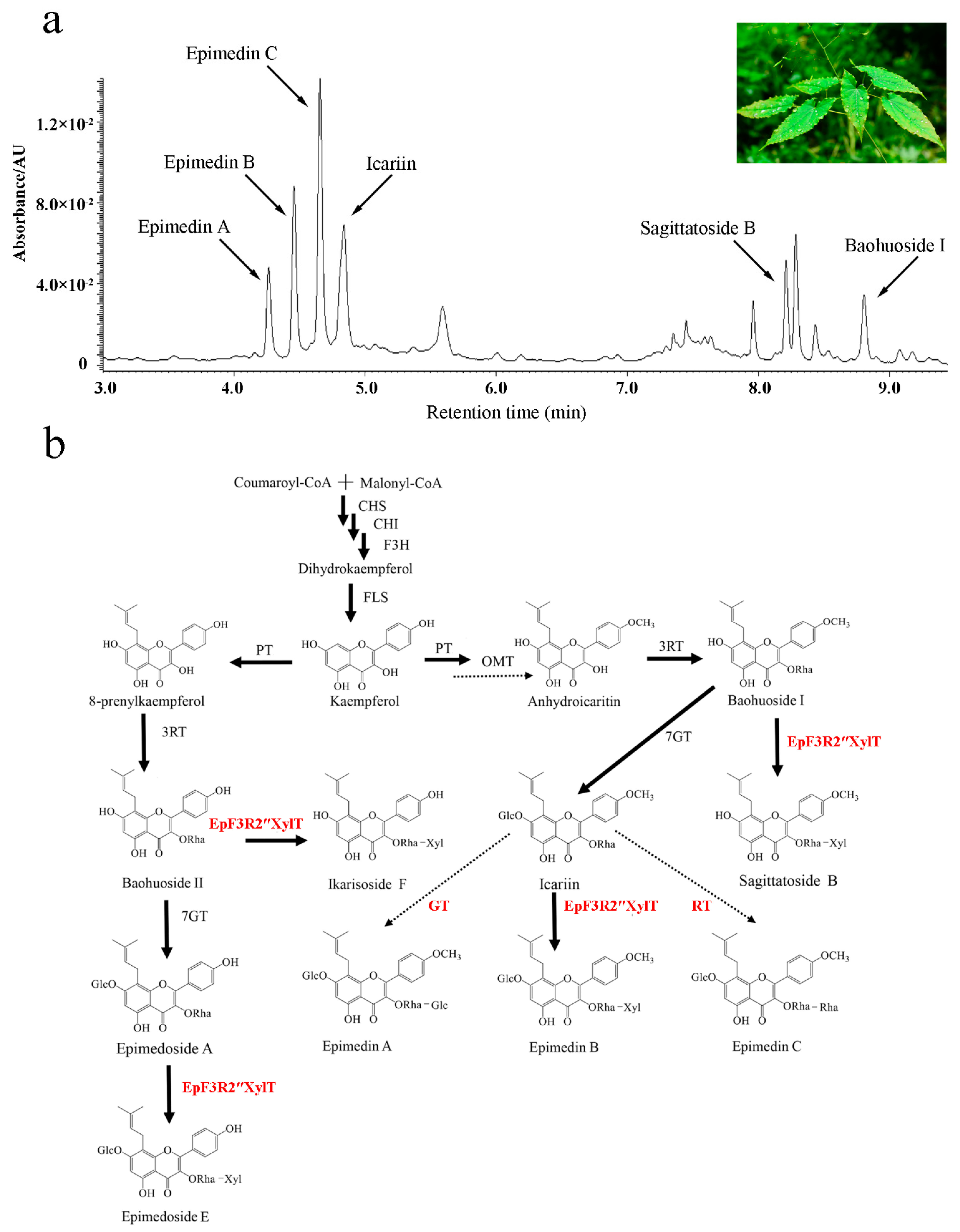
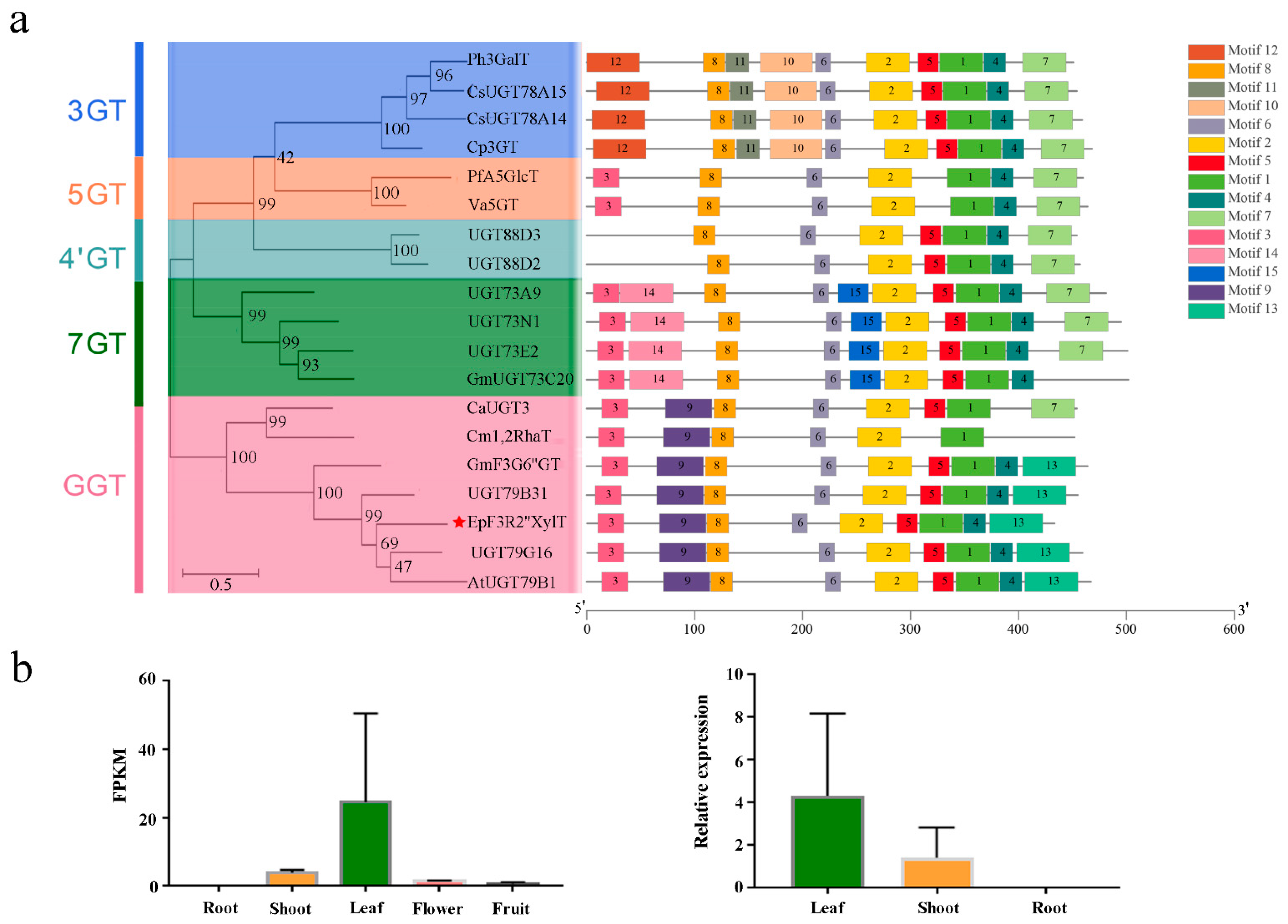
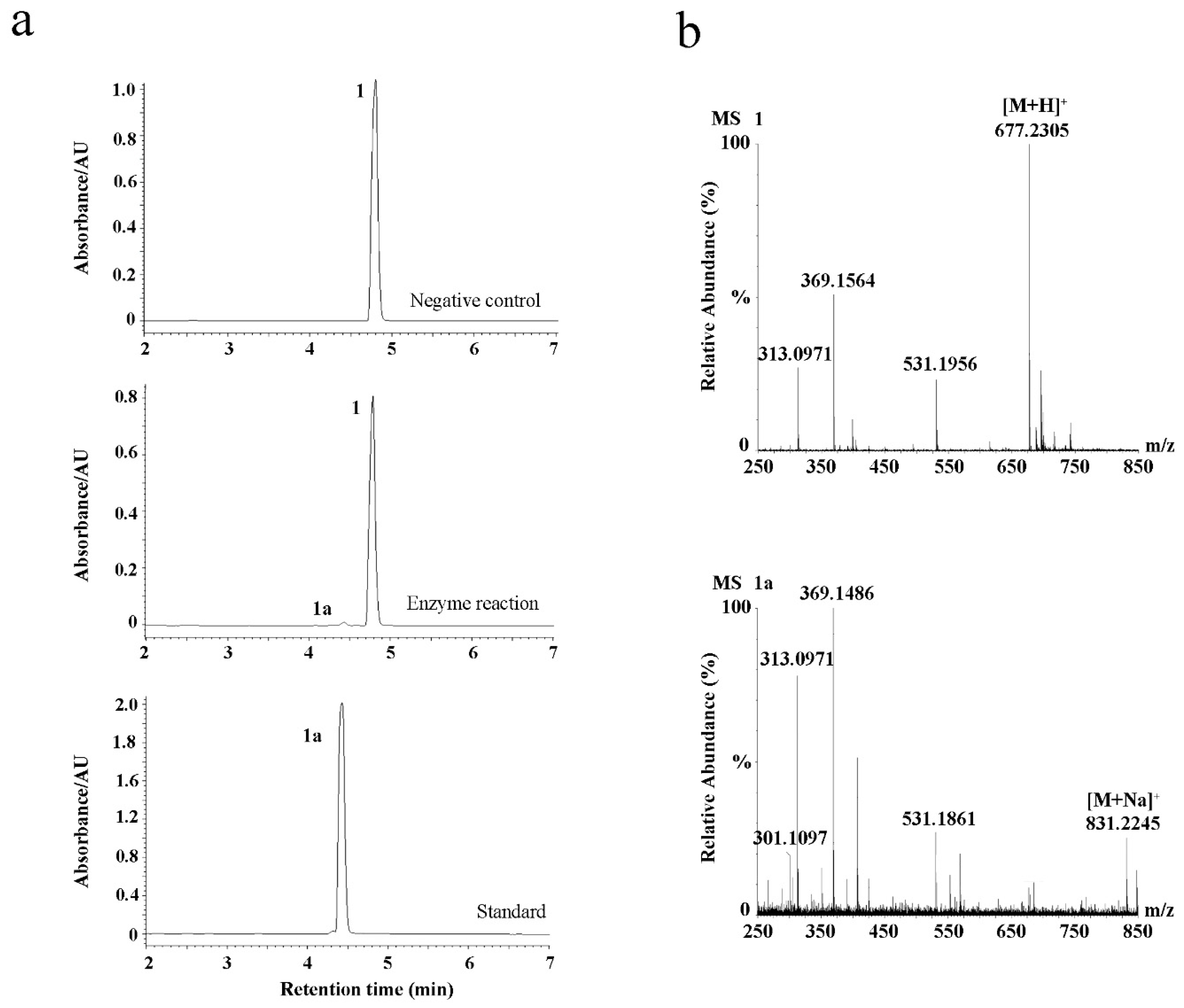
2.1. Identification of EpF3R2″XylT from E. pubescens
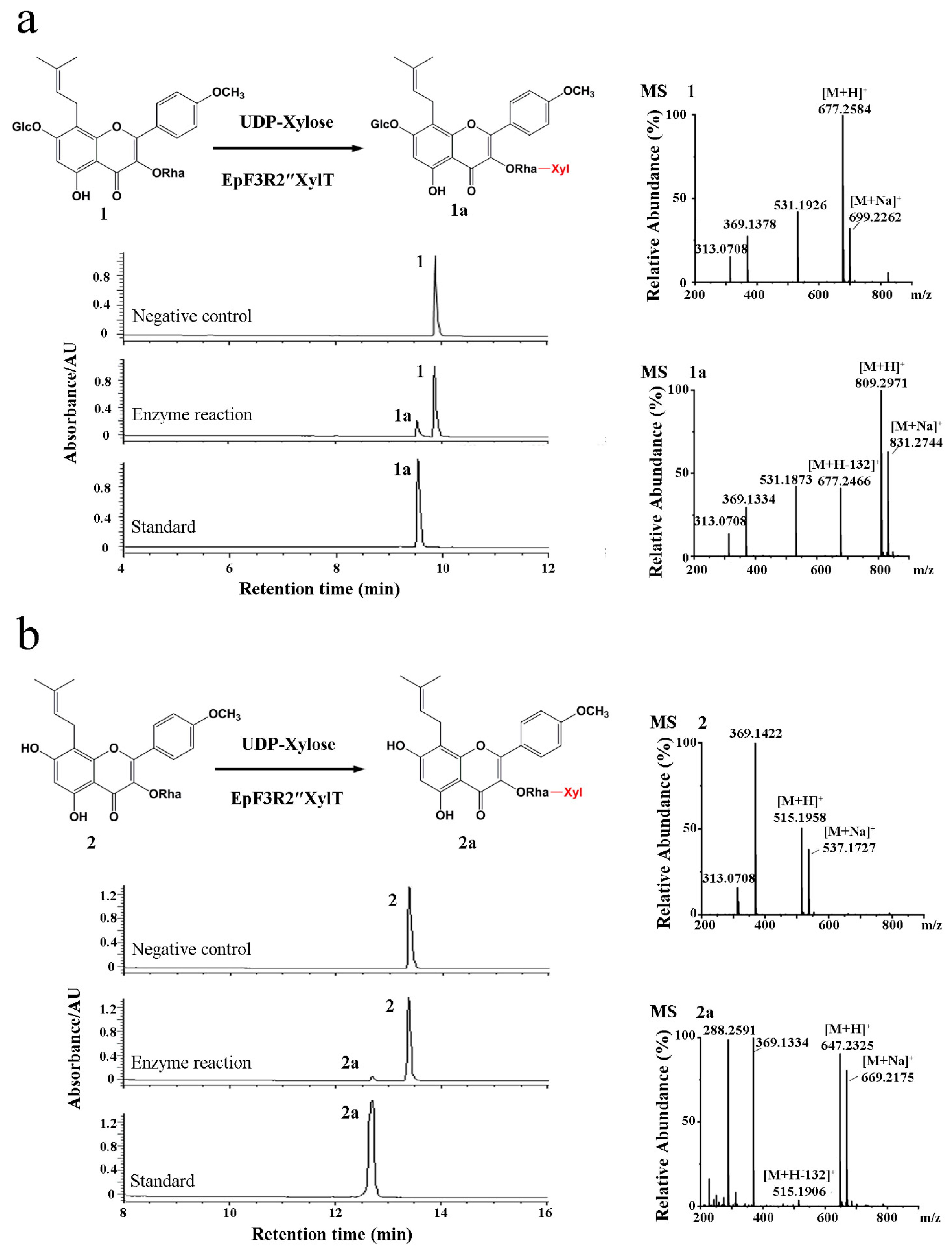
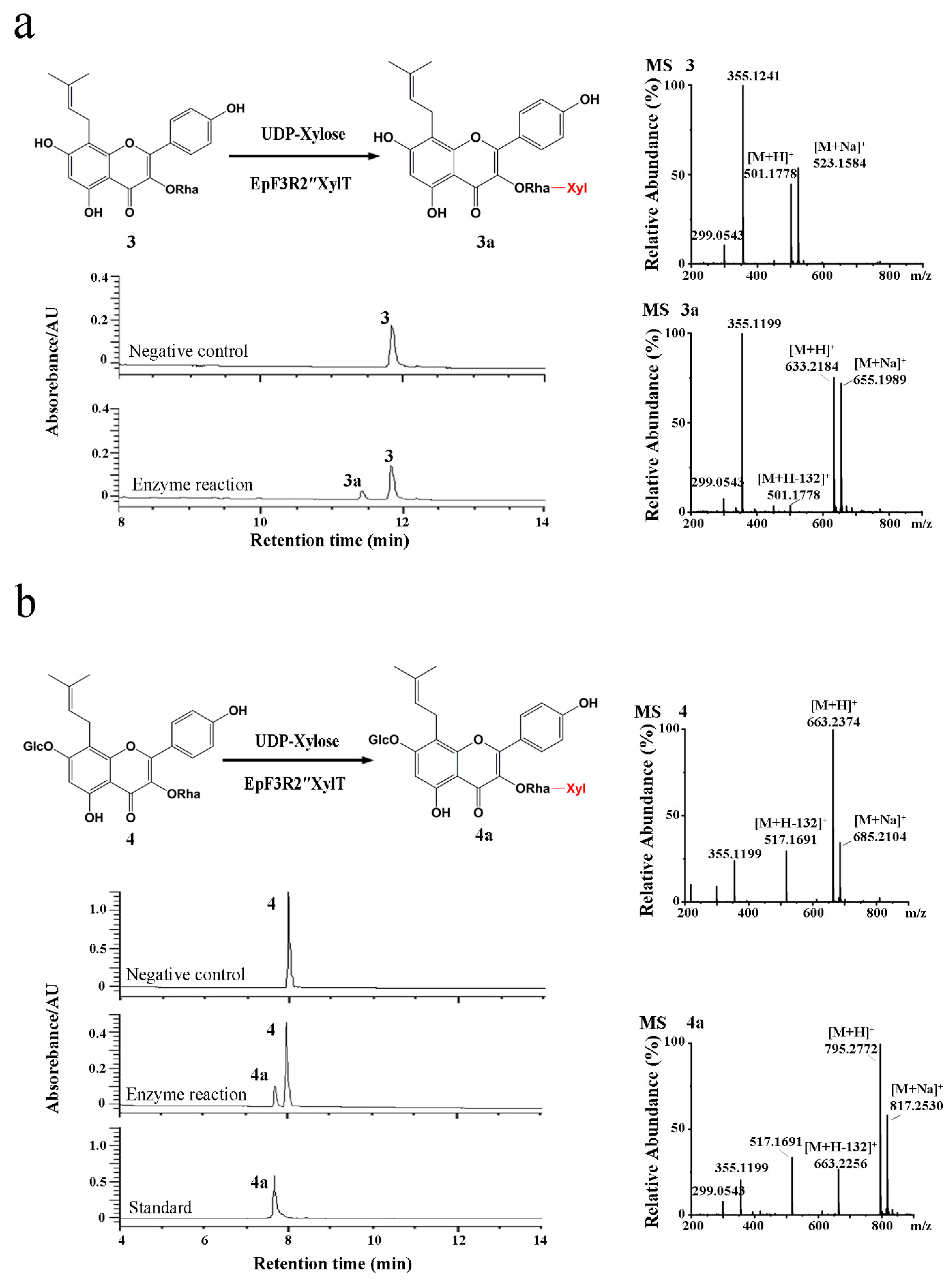
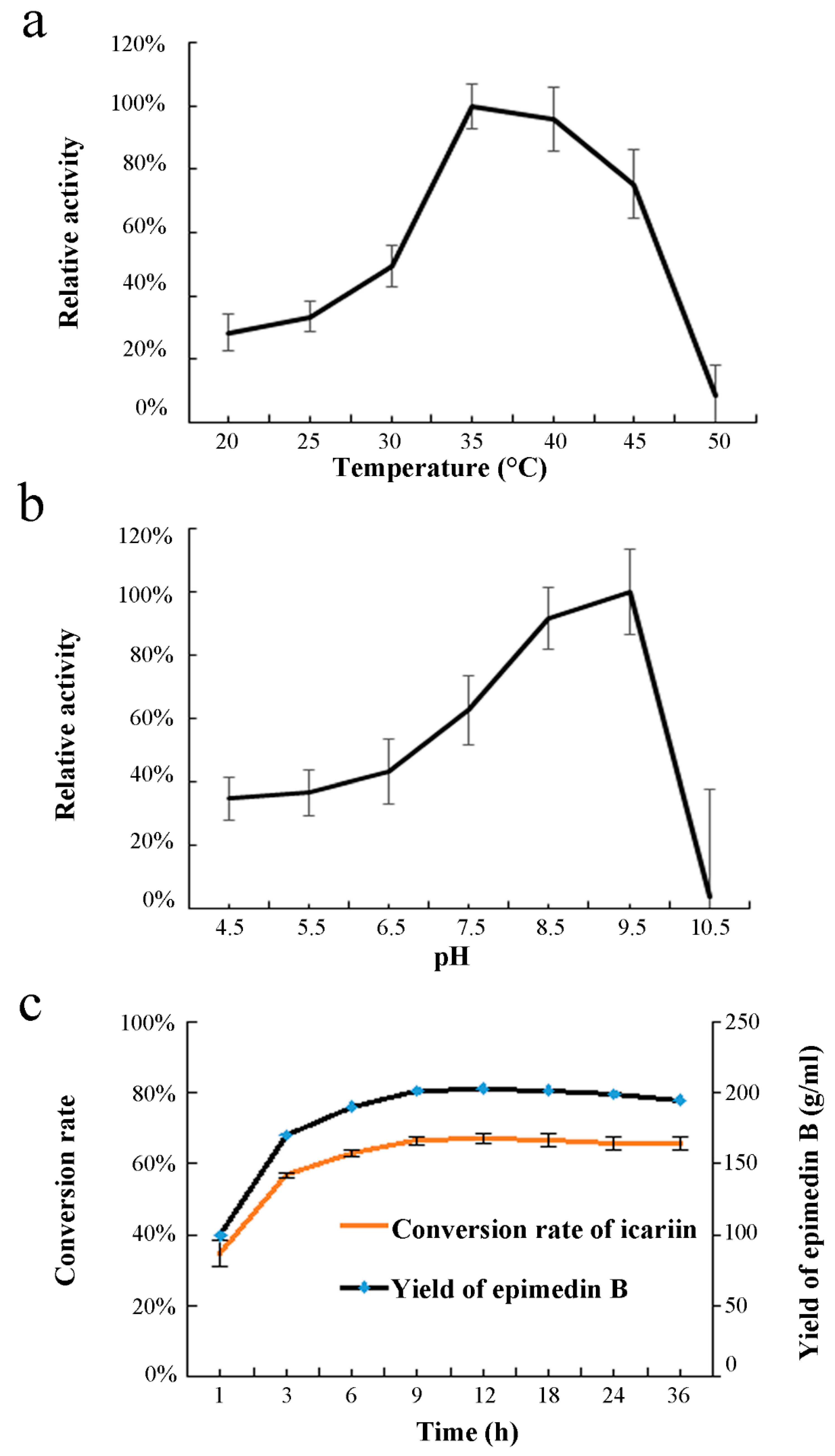
2.2. Enzyme Characterization of EpF3R2″XylT In Vitro
2.3. Enzymatic Kinetic Analysis of Recombinant EpF3R2″XylT
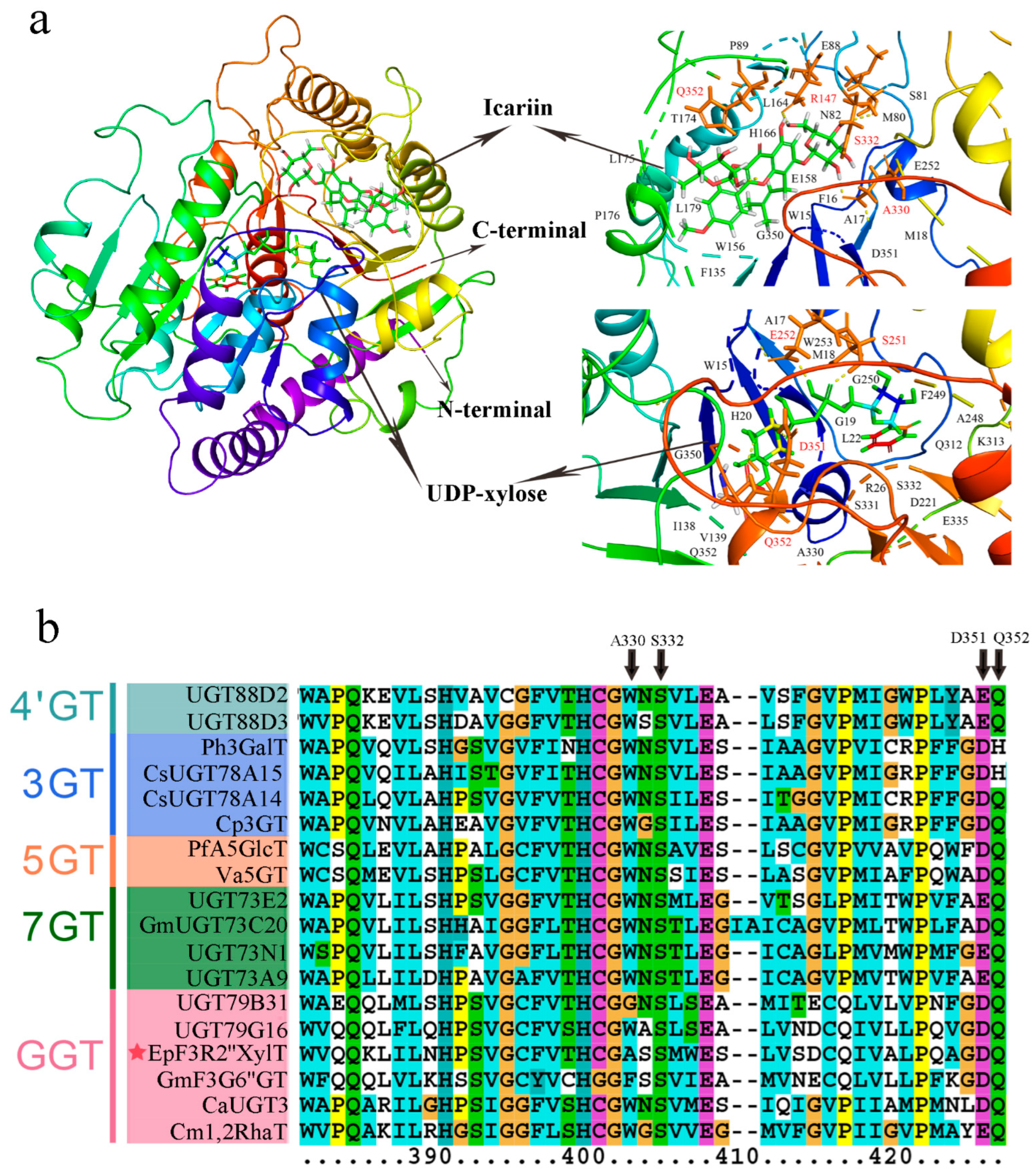
2.4. Modeling and Docking of EpF3R2″XylT Protein
2.5. Transient Expression of EpF3R2″XylT in Tobacco Leave
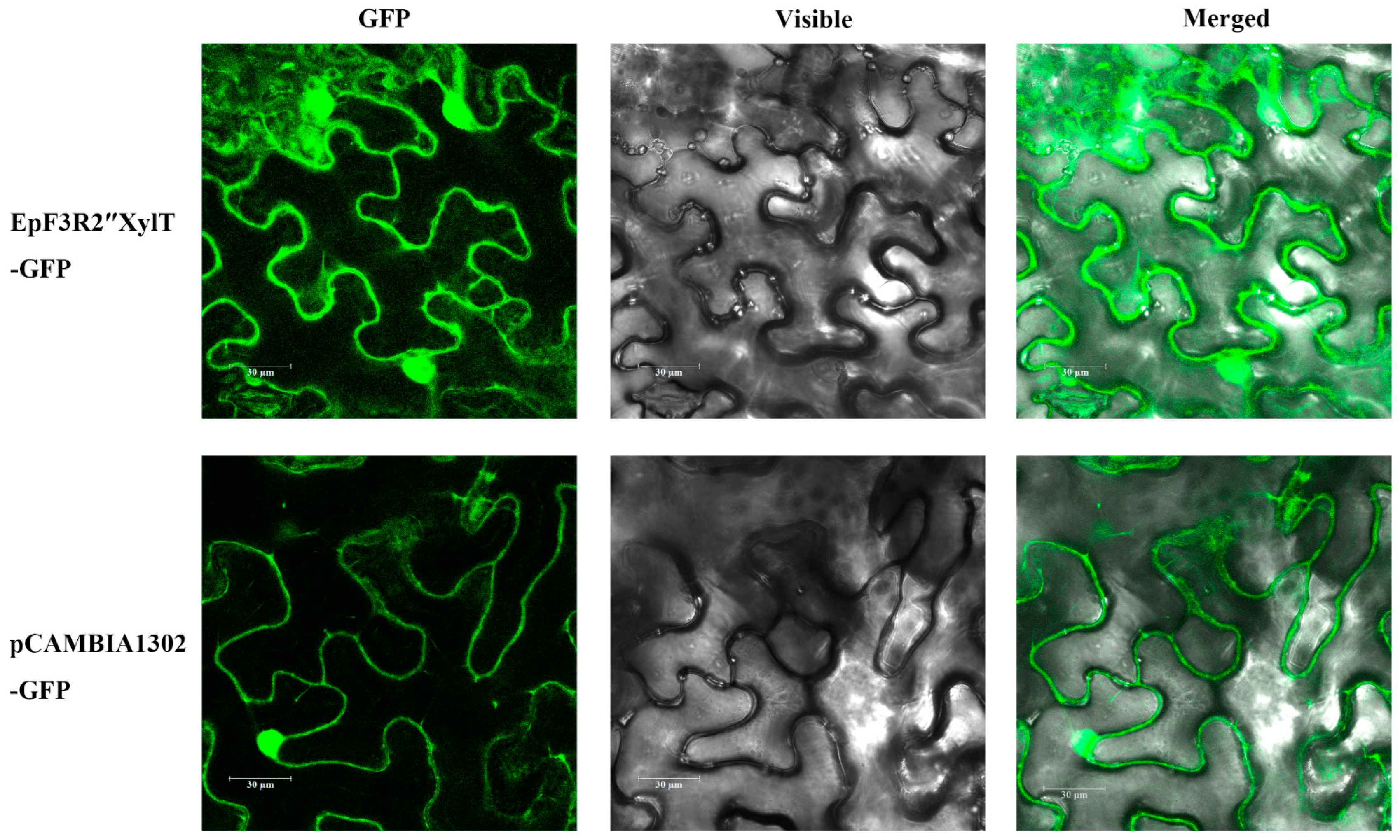
2.6. Subcellular Localization of EpF3R2″XylT
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Profiling Flavonoids in E. pubescens
4.3. Analysis of the Expression Levels of EpF3R2″XylT
4.4. Cloning and Heterologous Expression of EpF3R2″XylT in Escherichia coli
4.5. Enzymatic Assays of Recombinant EpF3R2″XylT
4.6. NMR Analysis of Enzymatic Product
4.7. Enzyme Kinetics of Recombinant EpF3R2″XylT
4.8. Bioinformatics Analyses of EpF3R2″XylT
4.9. Transient Expression of EpF3R2″XylT in Tobacco
4.10. Extraction of Crude EpF3R2″XylT Protein from Infiltrated Tobacco Leaves
4.11. Accession Number
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ma, H.; He, X.; Yang, Y.; Li, M.; Hao, D.; Jia, Z. The genus Epimedium: An ethnopharmacological and phytochemical review. J. Ethnopharmacol. 2011, 134, 519–541. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.K.; Choi, Y.H.; Kwon, H.; Lee, S.B.; Kim, D.H.; Sung, C.K.; Park, Y.I.; Dong, M.S. Estrogenic/antiestrogenic activities of a Epimedium koreanum extract and its major components: In vitro and in vivo studies. Food Chem. Toxicol. 2012, 50, 2751–2759. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Lien, E.J.; Lien, L.L. Chemical and pharmacological investigations of Epimedium species: A survey. Prog. Drug Res. 2003, 60, 1–57. [Google Scholar] [PubMed]
- Shen, P.; Guo, B.L.; Gong, Y.; Hong, D.Y.Q.; Hong, Y.; Yong, E.L. Taxonomic, genetic, chemical and estrogenic characteristics of Epimedium species. Phytochemistry 2007, 68, 1448–1458. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.F.; Yang, T.S.; Li, Z.Z.; Wang, Y. Simultaneous extraction of epimedin A, B, C and icariin from Herba Epimedii by ultrasonic technique. Ultrason. Sonochem. 2008, 15, 376–385. [Google Scholar] [CrossRef] [PubMed]
- Feng, K.; Chen, R.; Xie, K.; Chen, D.; Liu, J.; Du, W.; Yang, L.; Dai, J. Ep7GT, a glycosyltransferase with sugar donor flexibility from Epimedium pseudowushanense, catalyzes the 7-O-glycosylation of baohuoside. Org. Biomol. Chem. 2019, 17, 8106–8114. [Google Scholar] [CrossRef]
- Feng, K.; Chen, R.; Xie, K.; Chen, D.; Guo, B.; Liu, X.; Liu, J.; Zhang, M.; Dai, J. A regiospecific rhamnosyltransferase from Epimedium pseudowushanense catalyzes the 3-O-rhamnosylation of prenylflavonols. Org. Biomol. Chem. 2018, 16, 452–458. [Google Scholar] [CrossRef]
- Yao, Y.; Gu, J.; Luo, Y.; Wang, Y.; Pang, Y.; Shen, G.; Guo, B. Genome-wide analysis of UGT gene family identified key gene for the biosynthesis of bioactive flavonol glycosides in Epimedium pubescens Maxim. Synth. Syst. Biotechnol. 2022, 7, 1095–1107. [Google Scholar] [CrossRef]
- Yang, X.; Chen, J.; Huang, W.; Zhang, Y.; Yan, X.; Zhou, Z.; Wang, Y. Synthesis of icariin in tobacco leaf by overexpression of a glucosyltransferase gene from Epimedium sagittatum. Ind. Crops Prod. 2020, 156, 112841. [Google Scholar] [CrossRef]
- Lyu, Y.; Liu, S.; Gao, S.; Zhou, J. Identification and characterization of three flavonoid 3-O-glycosyltransferases from Epimedium koreanum Nakai. Biochem. Eng. J. 2020, 163, 107759. [Google Scholar] [CrossRef]
- Wang, P.; Li, C.; Li, X.; Huang, W.; Wang, Y.; Wang, J.; Zhang, Y.; Yang, X.; Yan, X.; Wang, Y.; et al. Complete biosynthesis of the potential medicine icaritin by engineered Saccharomyces cerevisiae and Escherichia coli. Sci. Bull. 2021, 66, 1906–1916. [Google Scholar] [CrossRef]
- Vogt, T.; Jones, P. Glycosyltransferases in plant natural product synthesis: Characterization of a supergene family. Trends Plant Sci. 2000, 5, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.; Vogt, T. Glycosyltransferases in secondary plant metabolism: Tranquilizers and stimulant controllers. Planta 2001, 213, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Yonekura-Sakakibara, K.; Nakabayashi, R.; Sugawara, S.; Tohge, T.; Ito, T.; Koyanagi, M.; Kitajima, M.; Takayama, H.; Saito, K. A flavonoid 3-O-glucoside:2″-O-glucosyltransferase responsible for terminal modification of pollen-specific flavonols in Arabidopsis thaliana. Plant J. 2014, 79, 769–782. [Google Scholar] [CrossRef] [Green Version]
- Rojas Rodas, F.; Di, S.; Murai, Y.; Iwashina, T.; Sugawara, S.; Mori, T.; Nakabayashi, R.; Yonekura-Sakakibara, K.; Saito, K.; Takahashi, R. Cloning and characterization of soybean gene Fg1 encoding flavonol 3-O-glucoside/galactoside (1→6) glucosyltransferase. Plant Mol. Biol. 2016, 92, 445–456. [Google Scholar] [CrossRef]
- Knoch, E.; Sugawara, S.; Mori, T.; Nakabayashi, R.; Saito, K.; Yonekura-Sakakibara, K. UGT79B31 is responsible for the final modification step of pollen-specific flavonoid biosynthesis in Petunia hybrida. Planta 2018, 247, 779–790. [Google Scholar] [CrossRef] [Green Version]
- Montefiori, M.; Espley, R.V.; Stevenson, D.; Cooney, J.; Datson, P.M.; Saiz, A.; Atkinson, R.G.; Hellens, R.P.; Allan, A.C. Identification and characterisation of F3GT1 and F3GGT1, two glycosyltransferases responsible for anthocyanin biosynthesis in red-fleshed kiwifruit (Actinidia chinensis). Plant J. 2011, 65, 106–118. [Google Scholar] [CrossRef]
- Yonekura-Sakakibara, K.; Fukushima, A.; Nakabayashi, R.; Hanada, K.; Matsuda, F.; Sugawara, S.; Inoue, E.; Kuromori, T.; Ito, T.; Shinozaki, K.; et al. Two glycosyltransferases involved in anthocyanin modification delineated by transcriptome independent component analysis in Arabidopsis thaliana. Plant J. 2012, 69, 154–167. [Google Scholar] [CrossRef] [Green Version]
- Yonekura-Sakakibara, K.; Saito, K. Function, structure, and evolution of flavonoid glycosyltransferases in plants. Recent Adv. Polyphen. Res. 2014, 4, 61–82. [Google Scholar] [CrossRef]
- Morita, Y.; Hoshino, A.; Kikuchi, Y.; Okuhara, H.; Ono, E.; Tanaka, Y.; Fukui, Y.; Saito, N.; Nitasaka, E.; Noguchi, H.; et al. Japanese morning glory dusky mutants displaying reddish-brown or purplish-gray flowers are deficient in a novel glycosylation enzyme for anthocyanin biosynthesis, UDP-glucose: Anthocyanidin 3-O-glucoside-2″-O-glucosyltransferase, due to 4-bp insertions in the gene. Plant J. 2005, 42, 353–363. [Google Scholar] [CrossRef]
- Xia, Q.; Xu, D.; Huang, Z.; Liu, J.; Wang, X.; Wang, X.; Liu, S. Preparation of icariside II from icariin by enzymatic hydrolysis method. Fitoterapia 2010, 8, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Oshima, Y.; Okamoto, M.; Hikino, H. Epimedins A, B and C, flavonoid glycosides of Epimedium koreanum herbs. Heterocycles 1987, 26, 935–938. [Google Scholar] [CrossRef]
- Zhang, J.; Tang, M.; Chen, Y.; Ke, D.; Zhou, J.; Xu, X.; Yang, W.; He, J.; Dong, H.; Wei, Y.; et al. Catalytic flexibility of rice glycosyltransferase OsUGT91C1 for the production of palatable steviol glycosides. Nat. Commun. 2021, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Bowles, D.; Lim, E.K.; Poppenberger, B.; Vaistij, F.E. Glycosyltransferases of lipophilic small molecules. Annu. Rev. Plant Biol. 2006, 57, 567–597. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Zheng, W.; Sun, X.; Yuan, M.; Zhang, J.; Chen, X.; Yu, K.; Guo, B.; Ma, B. Comparative analysis of chemical components in different parts of Epimedium Herb. J. Pharm. Biomed. Anal. 2021, 198, 113984. [Google Scholar] [CrossRef]
- Masada, S.; Terasaka, K.; Oguchi, Y.; Okazaki, S.; Mizushima, T.; Mizukami, H. Functional and structural characterization of a flavonoid glucoside 1,6-glucosyltransferase from Catharanthus roseus. Plant Cell Physiol. 2009, 50, 1401–1415. [Google Scholar] [CrossRef] [Green Version]
- Shen, G.; Luo, Y.; Yao, Y.; Meng, G.; Zhang, Y.; Wang, Y.; Xu, C.; Liu, X.; Zheng, C.; Ding, G.; et al. The discovery of a key prenyltransferase gene assisted by a chromosome-level Epimedium pubescens genome. Front. Plant Sci. 2022, 13, 1–14. [Google Scholar] [CrossRef]
- Khramtsov, P.; Kalashnikova, T.; Bochkova, M.; Kropaneva, M.; Timganova, V.; Zamorina, S.; Rayev, M. Measuring the concentration of protein nanoparticles synthesized by desolvation method: Comparison of bradford assay, BCA assay, hydrolysis/UV spectroscopy and gravimetric analysis. Int. J. Pharm. 2021, 599, 1–15. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Multiple sequence alignment using ClustalW and ClustalX. Curr. Protoc. Bioinform. 2002, 2, 1–22. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Bailey, T.; Bodén, M.; Buske, F.; Frith, M.; Grant, C.; Clementi, L.; Ren, J.; Li, W.; Noble, W. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, 296–303. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GTs Substrates | Km (Μm) | Kcat (S−1) | Kcat/Km (S–1 Μm–1) |
|---|---|---|---|
| Icariin | 75.96 ± 11.91 | 37.31 | 0.49 |
| Baohuoside I | 113.15 ± 37.60 | 16.64 | 0.15 |
| Baohuoside II | 123.97 ± 16.45 | 15.71 | 0.13 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, Y.; Gu, J.; Luo, Y.; Zhang, Y.; Wang, Y.; Pang, Y.; Jia, S.; Xu, C.; Li, D.; Suo, F.; et al. A Novel 3-O-rhamnoside: 2″-O-xylosyltransferase Responsible for Terminal Modification of Prenylflavonol Glycosides in Epimedium pubescens Maxim. Int. J. Mol. Sci. 2022, 23, 16050. https://doi.org/10.3390/ijms232416050
Yao Y, Gu J, Luo Y, Zhang Y, Wang Y, Pang Y, Jia S, Xu C, Li D, Suo F, et al. A Novel 3-O-rhamnoside: 2″-O-xylosyltransferase Responsible for Terminal Modification of Prenylflavonol Glycosides in Epimedium pubescens Maxim. International Journal of Molecular Sciences. 2022; 23(24):16050. https://doi.org/10.3390/ijms232416050
Chicago/Turabian StyleYao, Yu, Jiajun Gu, Yanjiao Luo, Yixin Zhang, Yuanyue Wang, Yongzhen Pang, Shangang Jia, Chaoqun Xu, Doudou Li, Fengmei Suo, and et al. 2022. "A Novel 3-O-rhamnoside: 2″-O-xylosyltransferase Responsible for Terminal Modification of Prenylflavonol Glycosides in Epimedium pubescens Maxim." International Journal of Molecular Sciences 23, no. 24: 16050. https://doi.org/10.3390/ijms232416050
APA StyleYao, Y., Gu, J., Luo, Y., Zhang, Y., Wang, Y., Pang, Y., Jia, S., Xu, C., Li, D., Suo, F., Shen, G., & Guo, B. (2022). A Novel 3-O-rhamnoside: 2″-O-xylosyltransferase Responsible for Terminal Modification of Prenylflavonol Glycosides in Epimedium pubescens Maxim. International Journal of Molecular Sciences, 23(24), 16050. https://doi.org/10.3390/ijms232416050