

CRISPR/Cas9 Mutant Rice Ospmei12 Involved in Growth, Cell Wall Development, and Response to Phytohormone and Heavy Metal Stress


 , ,
, ,
Abstract
:1. Introduction
2. Results
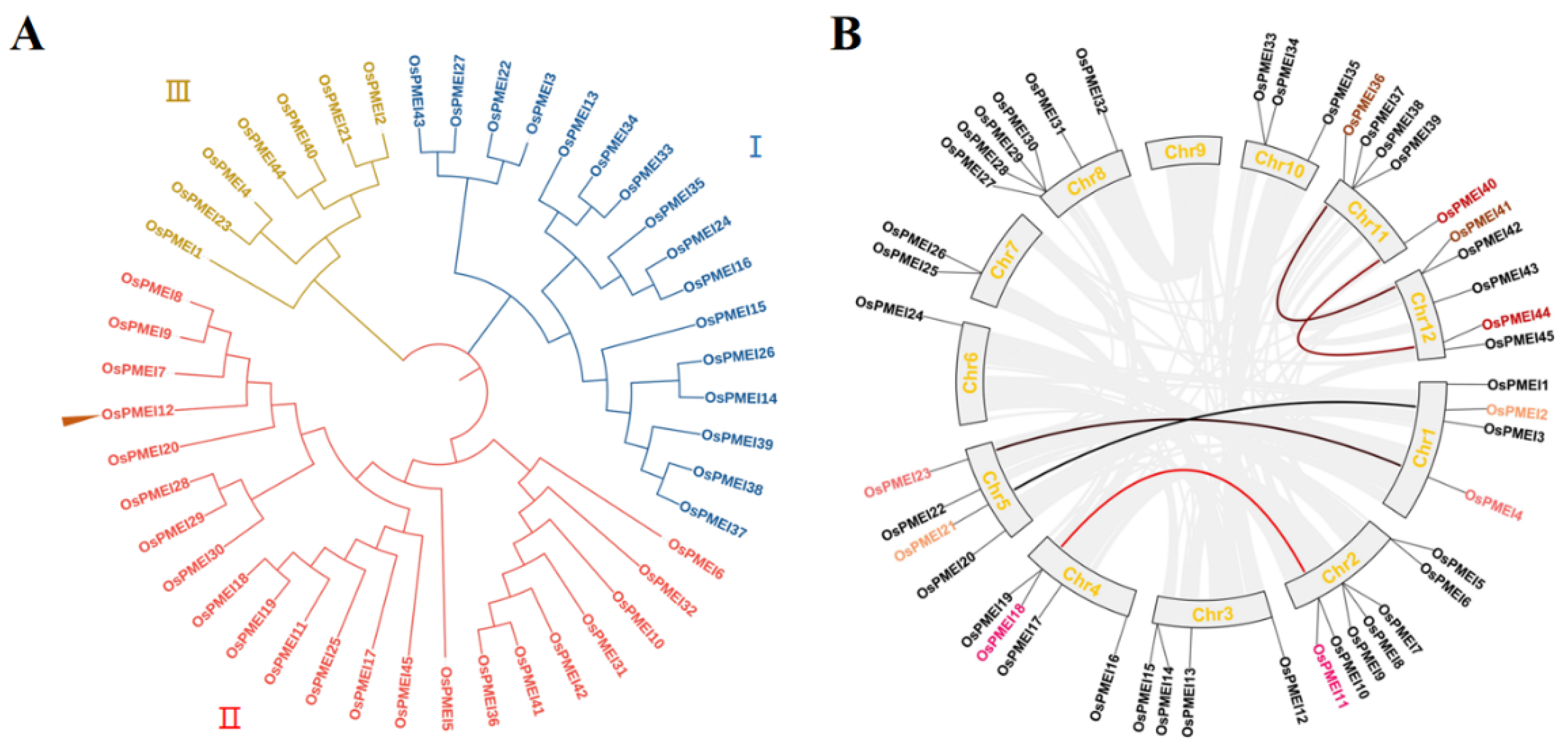
2.1. The Classification, Chromosomal Distribution, and Gene Duplication of OsPMEI Gene Family
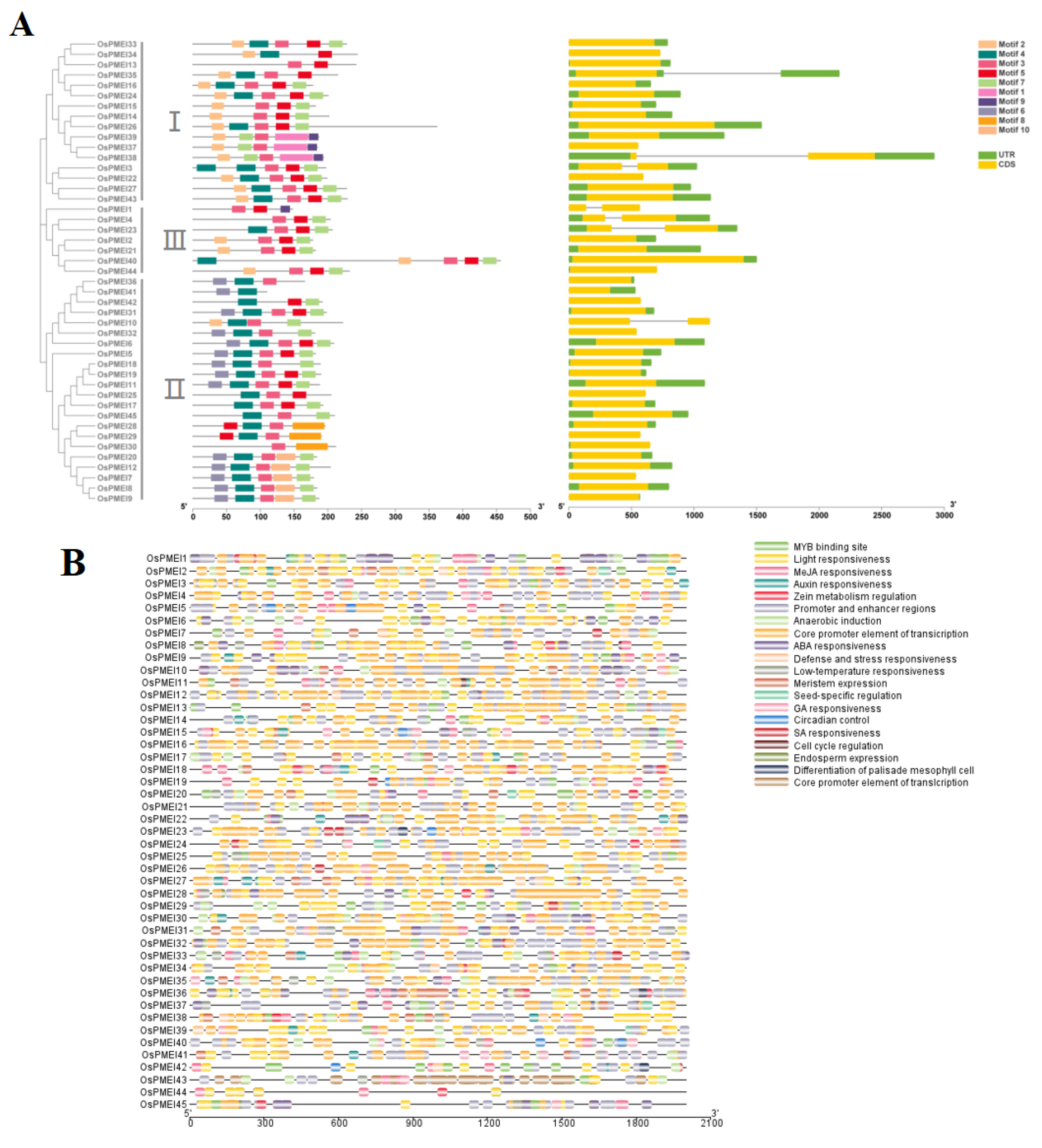
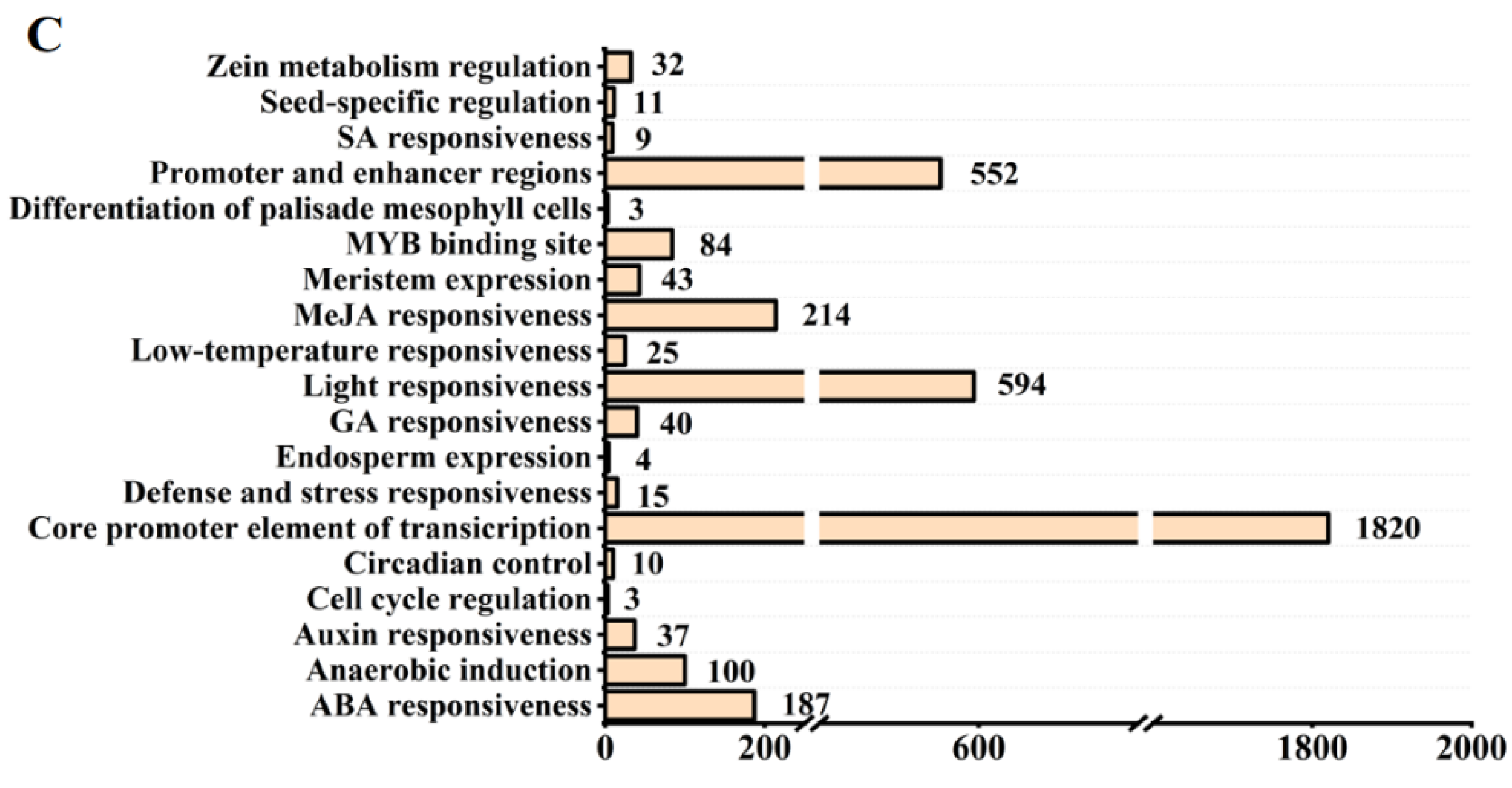
2.2. Gene Structure and Cis-Acting Elements in the Promoter Regions of OsPMEI Family
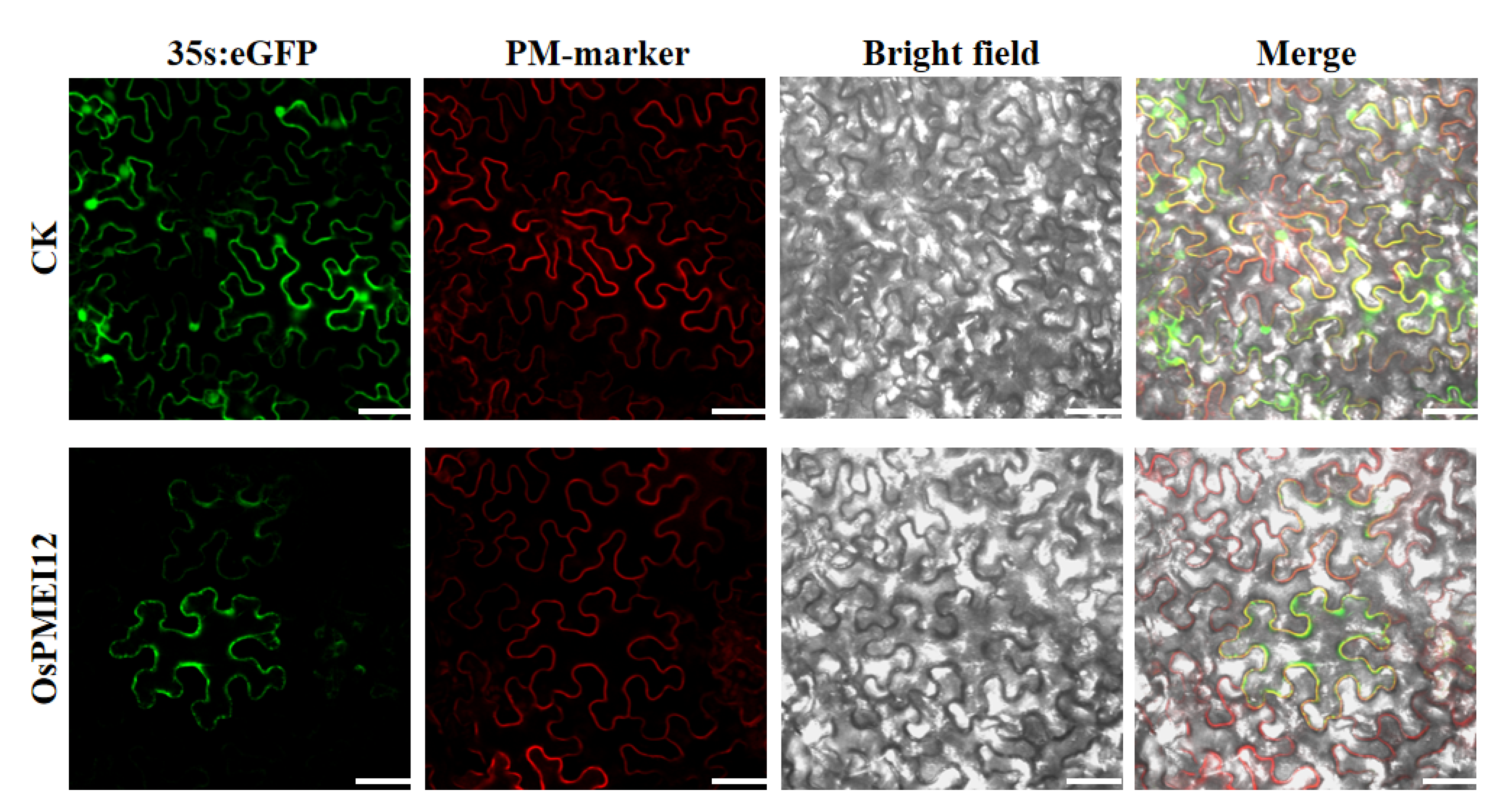
2.3. Subcellular Localization of the OsPMEI12 Protein
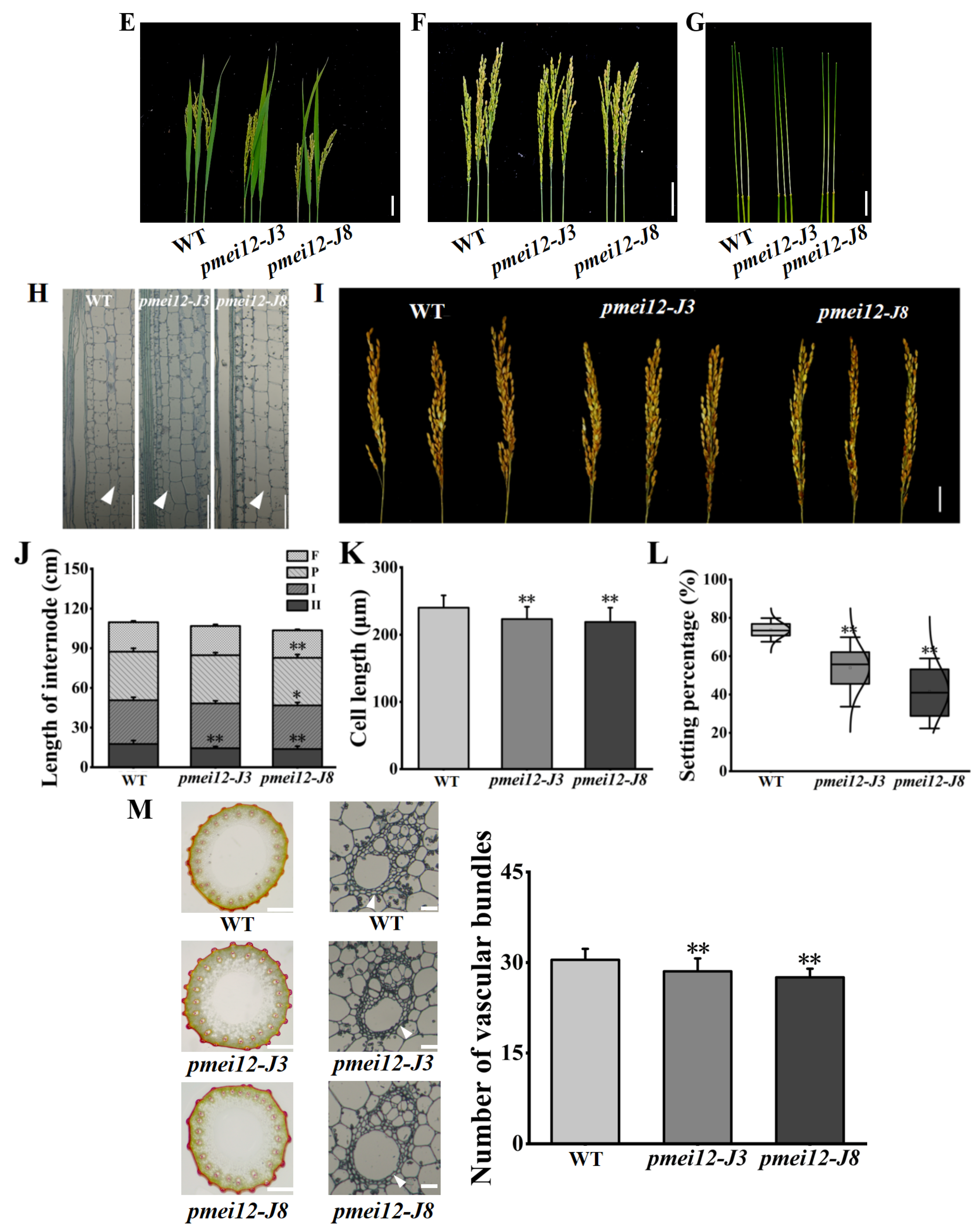
2.4. The Agronomic Traits and Vascular Bundles of OsPMEI12-Edited Lines
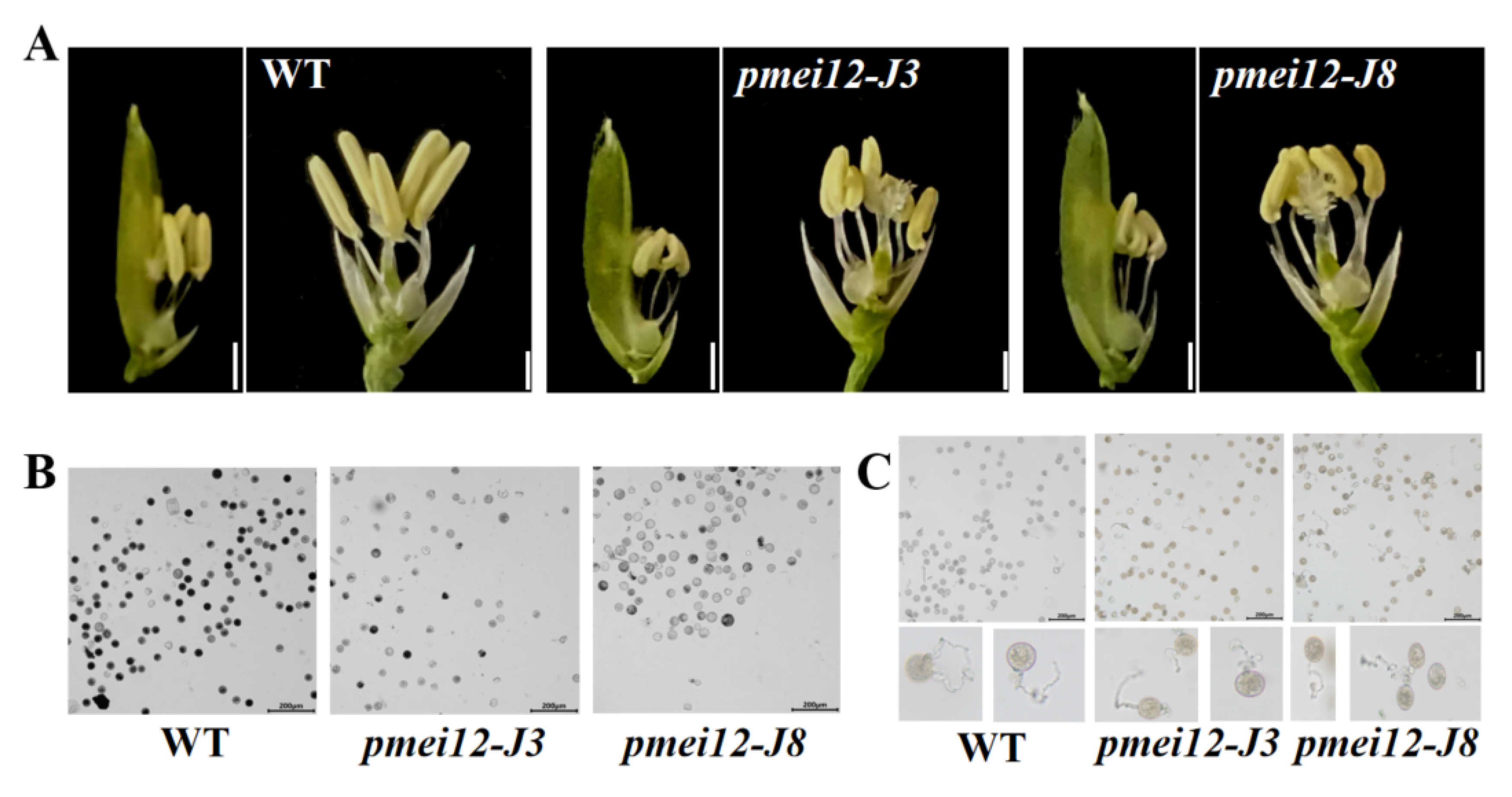
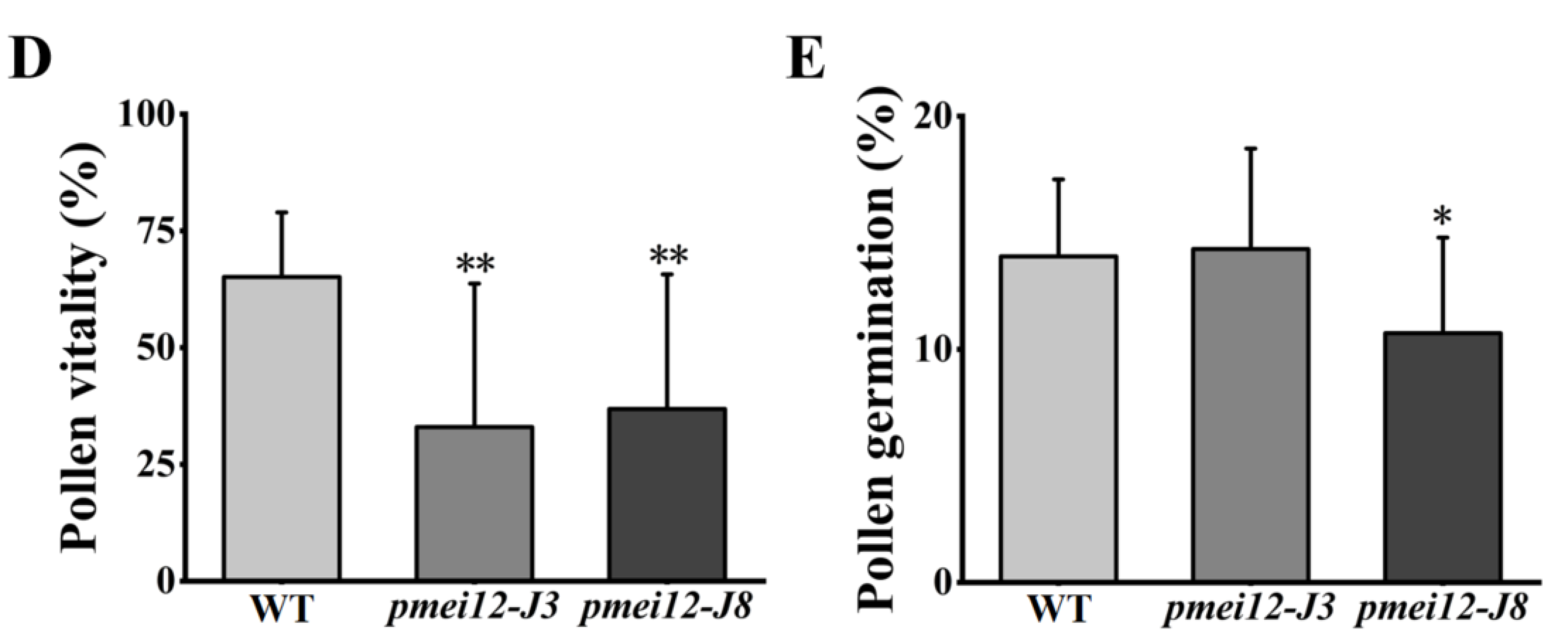
2.5. OsPMEI12 Affects Anther Growth and Pollen Fertility
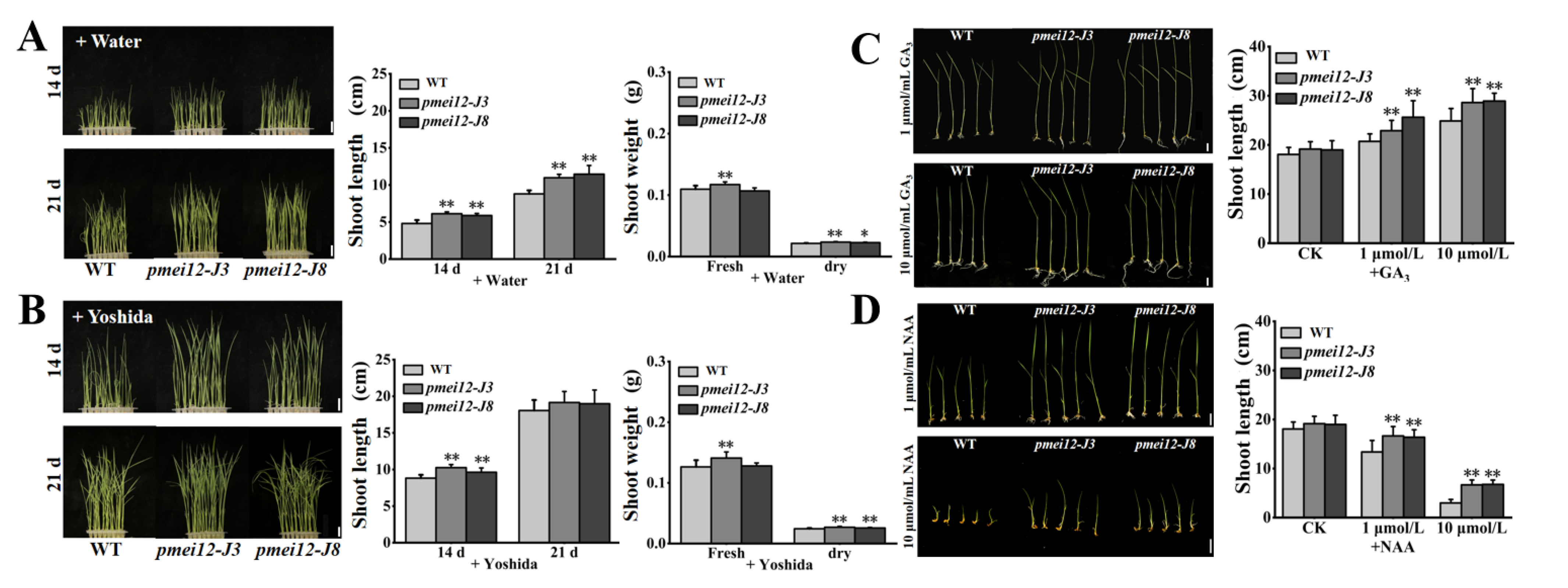
2.6. OsPMEI12-Edited Lines Showed Fast Growth and Altered Responses to Phytohormone at Seedling Stage
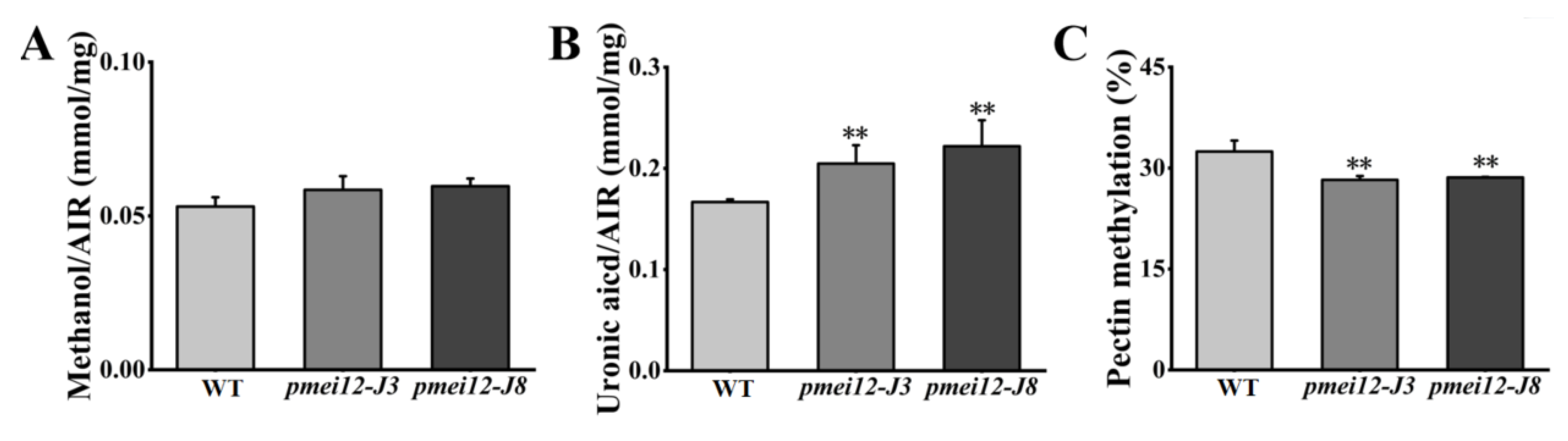
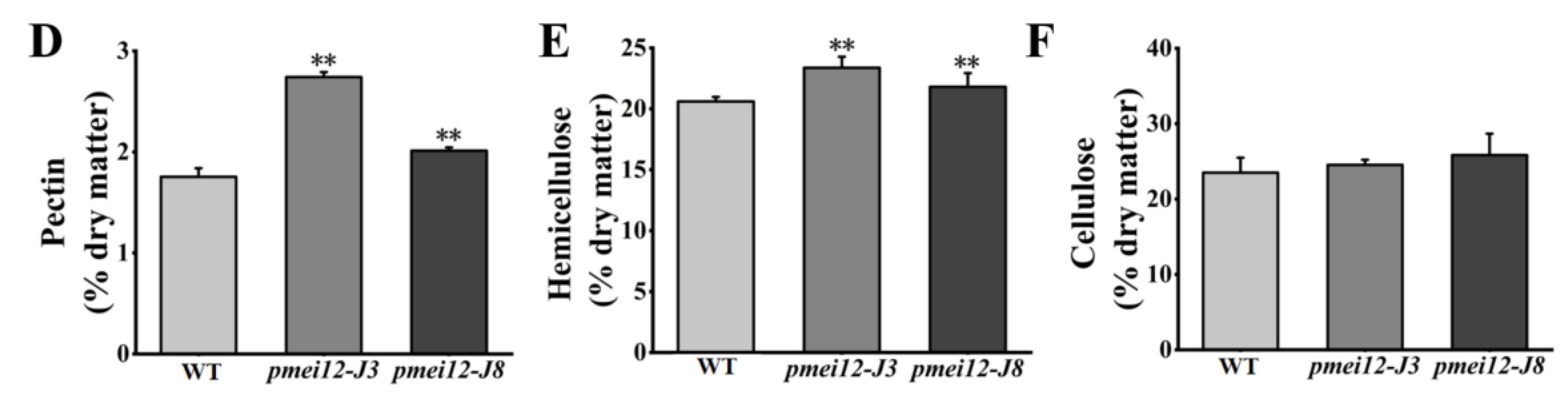
2.7. OsPMEI12 Affects the Biosynthesis of Cell Wall Components
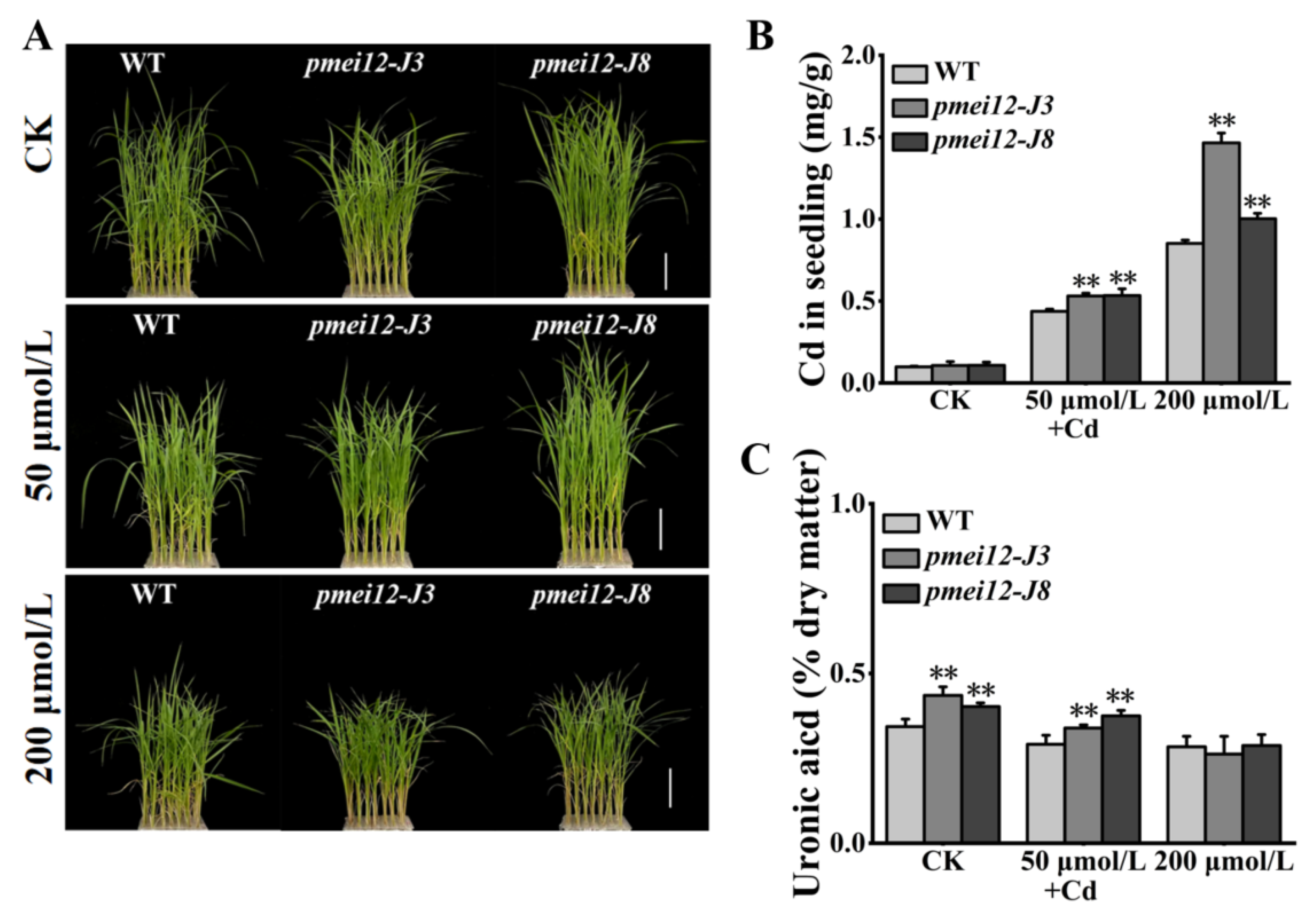
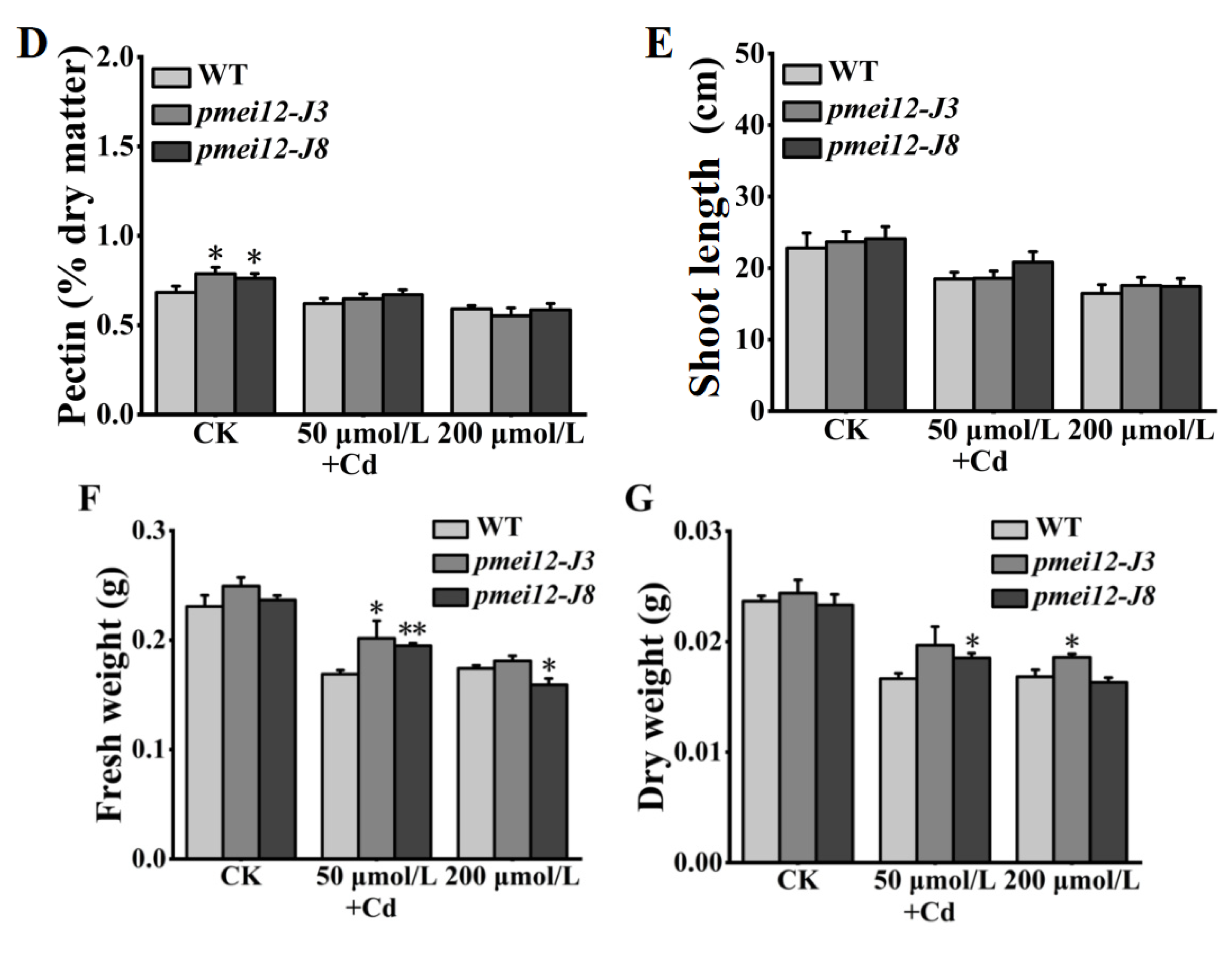
2.8. OsPMEI12 Is Involved in Cadmium Stress Response
3. Discussion
3.1. The Dynamic Effects of OsPMEI12 on the Plant Height of Rice
3.2. OsPMEI12 Involvement in the Pectin Methyl-Esterification and the Contents of the Pectin and Hemicellulose
3.3. OsPMEI12 Is Involved in the Hormone Response and Cadmium Stress
4. Materials and Methods
4.1. Experimental Materials
4.1.1. Rice Material
4.1.2. Strains and Vectors
4.1.3. Experimental Instruments and Reagents
4.2. Experimental Method
4.2.1. Bioinformatics Analysis
4.2.2. Construction of CRISPR Vector
4.2.3. PCR Detection of Transgenic Seedlings
4.2.4. Gene-Specific Primer Design
4.2.5. Investigation of Agronomic Traits
4.2.6. Investigation of Agronomic Characters of Rice
4.2.7. Determination of Rice Pollen Viability
4.2.8. Rice Pollen Tube Germination
4.2.9. Extraction of Cell Wall Polysaccharides
4.2.10. Determination of Pentose, Hexose, and Uronic Acid by Colorimetry
4.2.11. Preparation of Alcohol Insoluble Residue (AIR)
4.2.12. Measurements of Methyl-Esterification
4.2.13. Subcellular Localization Analysis
4.3. Rice Hormone Treatment and Determination of Cadmium Content in Rice
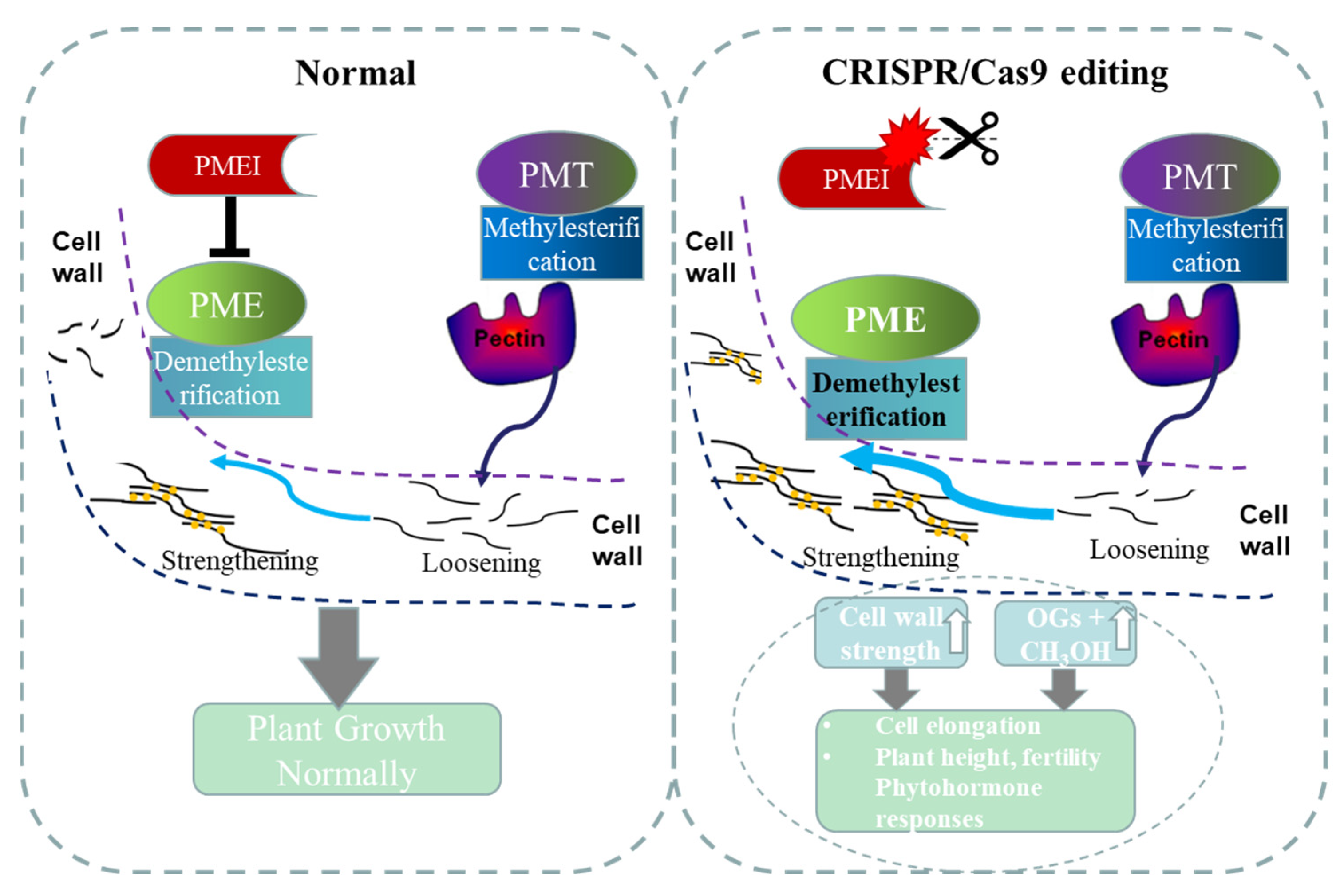
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cosgrove, D.J. Growth of the plant cell wall. Nat. Rev. Mol. Cell Biol. 2005, 6, 850–861. [Google Scholar] [CrossRef]
- Harholt, J.; Suttangkakul, A.; Vibe Scheller, H. Biosynthesis of pectin. Plant Physiol. 2010, 153, 384–395. [Google Scholar] [CrossRef] [Green Version]
- Giovane, A.; Servillo, L.; Balestrieri, C.; Raiola, A.; D’avino, R.; Tamburrini, M.; Ciardiello, M.A.; Camardella, L. Pectin methylesterase inhibitor. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2004, 1696, 245–252. [Google Scholar] [CrossRef]
- Giovane, A.; Quagliuolo, L.; Castaldo, D.; Servillo, L.; Balestrieri, C. Pectin methyl esterase from Actinidia chinensis fruits. Phytochemistry 1990, 29, 2821–2823. [Google Scholar] [CrossRef]
- Nguyen, H.P.; Jeong, H.Y.; Kim, H.; Kim, Y.C.; Lee, C. Molecular and biochemical characterization of rice pectin methylesterase inhibitors (OsPMEIs). Plant Physiol. Biochem. 2016, 101, 105–112. [Google Scholar] [CrossRef]
- Lionetti, V.; Raiola, A.; Camardella, L.; Giovane, A.; Obel, N.; Pauly, M.; Favaron, F.; Cervone, F.; Bellincampi, D. Overexpression of pectin methylesterase inhibitors in Arabidopsis restricts fungal infection by Botrytis cinerea. Plant Physiol. 2007, 143, 1871–1880. [Google Scholar] [CrossRef] [Green Version]
- Mattei, B.; Raiola, A.; Caprari, C.; Federici, L.; Bellincampi, D.; De Lorenzo, G.; Cervone, F.; Giovane, A.; Camardella, L. Studies on plant inhibitors of pectin modifying enzymes: Polygalacturonase-inhibiting protein (PGIP) and pectin methylesterase inhibtior (PMEI). Carbohydr. Bioeng. 2002. [Google Scholar] [CrossRef]
- D’Avino, R.; Camardella, L.; Christensen, T.M.I.E.; Giovane, A.; Servillo, L. Tomato pectin methylesterase: Modeling, fluorescence, and inhibitor interaction studies—Comparison with the bacterial (Erwinia chrysanthemi) enzyme. Proteins Struct. Funct. Bioinforma. 2003, 53, 830–839. [Google Scholar] [CrossRef]
- Dedeurwaerder, S.; Menu-Bouaouiche, L.; Mareck, A.; Lerouge, P.; Guerineau, F. Activity of an atypical Arabidopsis thaliana pectin methylesterase. Planta 2009, 229, 311–321. [Google Scholar] [CrossRef]
- Wormit, A.; Usadel, B. The multifaceted role of pectin methylesterase inhibitors (PMEIs). Int. J. Mol. Sci. 2018, 19, 2878. [Google Scholar] [CrossRef]
- Nguyen, H.P.; Jeong, H.Y.; Jeon, S.H.; Kim, D.; Lee, C. Rice pectin methylesterase inhibitor28 (OsPMEI28) encodes a functional PMEI and its overexpression results in a dwarf phenotype through increased pectin methylesterification levels. J. Plant Physiol. 2017, 208, 17–25. [Google Scholar] [CrossRef]
- Wolf, S.; Mravec, J.; Greiner, S.; Mouille, G.; Höfte, H. Plant cell wall homeostasis is mediated by brassinosteroid feedback signaling. Curr. Biol. 2012, 22, 1732–1737. [Google Scholar] [CrossRef] [Green Version]
- Müller, K.; Levesque-Tremblay, G.; Bartels, S.; Weitbrecht, K.; Wormit, A.; Usadel, B.; Haughn, G.; Kermode, A.R. Demethylesterification of cell wall pectins in Arabidopsis plays a role in seed germination. Plant Physiol. 2013, 161, 305–316. [Google Scholar] [CrossRef] [Green Version]
- Brutus, A.; Sicilia, F.; Macone, A.; Cervone, F.; De Lorenzo, G. A domain swap approach reveals a role of the plant wall-associated kinase 1 (WAK1) as a receptor of oligogalacturonides. Proc. Natl. Acad. Sci. USA 2010, 107, 9452–9457. [Google Scholar] [CrossRef] [Green Version]
- Kohorn, B.D. Cell wall-associated kinases and pectin perception. J. Exp. Bot. 2016, 67, 489–494. [Google Scholar] [CrossRef] [Green Version]
- Yue, Z.-L.; Liu, N.; Deng, Z.-P.; Zhang, Y.; Wu, Z.-M.; Zhao, J.-L.; Sun, Y.; Wang, Z.-Y.; Zhang, S.-W. The receptor kinase OsWAK11 monitors cell wall pectin changes to fine-tune brassinosteroid signaling and regulate cell elongation in rice. Curr. Biol. 2022, 32, 2454–2466. [Google Scholar] [CrossRef]
- Aryal, B.; Jonsson, K.; Baral, A.; Sancho-Andres, G.; Routier-Kierzkowska, A.-L.; Kierzkowski, D.; Bhalerao, R.P. Interplay between cell wall and auxin mediates the control of differential cell elongation during apical hook development. Curr. Biol. 2020, 30, 1733–1739. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Chen, G.; Zhang, H.; Qian, Q.; Ding, Y. Comparative transcript profiling of alloplasmic male-sterile lines revealed altered gene expression related to pollen development in rice (Oryza sativa L.). BMC Plant Biol. 2016, 16, 175. [Google Scholar] [CrossRef] [Green Version]
- Laser, K.D.; Lersten, N.R. Anatomy and cytology of microsporogenesis in cytoplasmic male sterile angiosperms. Bot. Rev. 1972, 38, 425–454. [Google Scholar] [CrossRef]
- Kasembe, J.N.R. Phenotypic restoration of fertility in a male-sterile mutant by treatment with gibberellic acid. Nature 1967, 215, 668. [Google Scholar] [CrossRef]
- Wu, Z.; Hu, K.; Fu, J.; Qiao, A. Relationships between Cytoplasmic Male Sterility and Endogenous Hormone Content of Pepper Bud. J. South China Agric. Univ. 2010, 31, 1–4. [Google Scholar]
- An, S.H.; Sohn, K.H.; Choi, H.W.; Hwang, I.S.; Lee, S.C.; Hwang, B.K. Pepper pectin methylesterase inhibitor protein CaPMEI1 is required for antifungal activity, basal disease resistance and abiotic stress tolerance. Planta 2008, 228, 61–78. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Sun, Y.; Pei, Y.; Zhang, X.; Wang, P.; Li, X.; Li, F.; Hou, Y. A pectin methylesterase inhibitor enhances resistance to Verticillium wilt. Plant Physiol. 2018, 176, 2202–2220. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Wang, H.; Qin, X.; Chen, K.; Zhao, J.; Zhao, Y.; Yue, B. Genome-wide identification, phylogeny and expression analysis of the PME and PMEI gene families in maize. Sci. Rep. 2019, 9, 19918. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Hu, M.; Hu, Z.; Liu, F.; Yu, H.; Yang, Q.; Gao, H.; Xu, C.; Wang, M.; Zhang, G. Insights into pectin dominated enhancements for elimination of toxic Cd and dye coupled with ethanol production in desirable lignocelluloses. Carbohydr. Polym. 2022, 286, 119298. [Google Scholar] [CrossRef]
- Douchiche, O.; Rihouey, C.; Schaumann, A.; Driouich, A.; Morvan, C. Cadmium-induced alterations of the structural features of pectins in flax hypocotyl. Planta 2007, 225, 1301–1312. [Google Scholar] [CrossRef]
- Colzi, I.; Doumett, S.; Del Bubba, M.; Fornaini, J.; Arnetoli, M.; Gabbrielli, R.; Gonnelli, C. On the role of the cell wall in the phenomenon of copper tolerance in Silene paradoxa L. Environ. Exp. Bot. 2011, 72, 77–83. [Google Scholar] [CrossRef]
- Chang, Y.; Yamamoto, Y.; Matsumoto, H. Accumulation of aluminium in the cell wall pectin in cultured tobacco (Nicotiana tabacum L.) cells treated with a combination of aluminium and iron. Plant. Cell Environ. 1999, 22, 1009–1017. [Google Scholar] [CrossRef]
- Horst, W.J.; Schmohl, N.; Kollmeier, M.; Sivaguru, M. Does aluminium affect root growth of maize through interaction with the cell wall–plasma membrane–cytoskeleton continuum? Plant Soil 1999, 215, 163–174. [Google Scholar] [CrossRef]
- Eticha, D.; Stass, A.; Horst, W.J. Cell-wall pectin and its degree of methylation in the maize root-apex: Significance for genotypic differences in aluminium resistance. Plant. Cell Environ. 2005, 28, 1410–1420. [Google Scholar] [CrossRef]
- Konno, H.; Nakato, T.; Nakashima, S.; Katoh, K. Lygodium japonicum fern accumulates copper in the cell wall pectin. J. Exp. Bot. 2005, 56, 1923–1931. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.J.; Wang, L. CRISPR/Cas9 technology for improving agronomic traits and future prospective in agriculture. Planta 2021, 254, 68. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.J.; Zuo, H.; Xu, Q. Genomic insights into citrus domestication and its important agronomic traits. Plant Commun. 2021, 2, 100138. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-J.; Jeong, H.Y.; Kang, S.-Y.; Silva, J.; Kim, E.-J.; Park, S.K.; Jung, K.-H.; Lee, C. Physiological importance of pectin modifying genes during rice pollen development. Int. J. Mol. Sci. 2020, 21, 4840. [Google Scholar] [CrossRef]
- Du, J.; Kirui, A.; Huang, S.; Wang, L.; Barnes, W.J.; Kiemle, S.N.; Zheng, Y.; Rui, Y.; Ruan, M.; Qi, S. Mutations in the pectin methyltransferase QUASIMODO2 influence cellulose biosynthesis and wall integrity in Arabidopsis. Plant Cell 2020, 32, 3576–3597. [Google Scholar] [CrossRef]
- Li, F.; Xie, G.; Huang, J.; Zhang, R.; Li, Y.; Zhang, M.; Wang, Y.; Li, A.; Li, X.; Xia, T. Os CESA 9 conserved-site mutation leads to largely enhanced plant lodging resistance and biomass enzymatic saccharification by reducing cellulose DP and crystallinity in rice. Plant Biotechnol. J. 2017, 15, 1093–1104. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Yuan, D.; Gao, W.; Li, Y.; Tan, J.; Zhang, X. A comparative genome analysis of PME and PMEI families reveals the evolution of pectin metabolism in plant cell walls. PLoS ONE 2013, 8, e72082. [Google Scholar] [CrossRef] [Green Version]
- Müller, K.; Levesque-Tremblay, G.; Fernandes, A.; Wormit, A.; Bartels, S.; Usadel, B.; Kermode, A. Overexpression of a pectin methylesterase inhibitor in Arabidopsis thaliana leads to altered growth morphology of the stem and defective organ separation. Plant Signal. Behav. 2013, 8, e26464. [Google Scholar] [CrossRef] [Green Version]
- Klavons, J.A.; Bennett, R.D. Determination of methanol using alcohol oxidase and its application to methyl ester content of pectins. J. Agric. Food Chem. 1986, 34, 597–599. [Google Scholar] [CrossRef]
- Hong, M.J.; Kim, D.Y.; Lee, T.G.; Jeon, W.B.; Seo, Y.W. Functional characterization of pectin methylesterase inhibitor (PMEI) in wheat. Genes Genet. Syst. 2010, 85, 97–106. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, S.; Gupta, S.M.; Sane, A.P.; Nath, P. Isolation and characterization of ripening related pectin methylesterase inhibitor gene from banana fruit. Physiol. Mol. Biol. Plants 2012, 18, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Yu, H.; Xiong, X.; Yue, X.; Yu, Y.; Huang, L.; Cao, J. Genome-wide identification, molecular evolution, and expression profiling analysis of pectin methylesterase inhibitor genes in Brassica campestris ssp. chinensis. Int. J. Mol. Sci. 2018, 19, 1338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, C.; Ming, H.; Lin, C.; Naidu, R.; Bolan, N. Phytoextraction of heavy metal from tailing waste using Napier grass. CATENA 2016, 136, 74–83. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Filisetti-Cozzi, T.M.C.C.; Carpita, N.C. Measurement of uronic acids without interference from neutral sugars. Anal. Biochem. 1991, 197, 157–162. [Google Scholar] [CrossRef]
- Batistic, O.; Sorek, N.; Schultke, S. Dual fatty acyl modification determines the localization and plasma membrane targeting of CBL/CIPK Ca2+ signaling complexes in Arabidopsis. Plant Cell 2008, 20, 1346–1362. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tiller Number | Total Number of Panicles | Effective Panicles | 1000 Grain Weight (g) | Plant Height (cm) | |
|---|---|---|---|---|---|
| WT | 48 ± 12 | 38 ± 3 | 23 ± 4 | 21.23 ± 0.32 | 72.96 ± 3.29 |
| pmei12-J3 | 48 ± 13 | 43 ± 15 | 23 ± 9 | 21.2 ± 0.043 | 68.37 ± 2.75 ** |
| pmei12-J8 | 50 ± 13 | 36 ± 10 | 22 ± 6 | 21.2 ± 0.16 | 67.57 ± 3.29 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Rao, M.J.; Li, J.; Wang, Y.; Chen, P.; Yu, H.; Ma, C.; Wang, L. CRISPR/Cas9 Mutant Rice Ospmei12 Involved in Growth, Cell Wall Development, and Response to Phytohormone and Heavy Metal Stress. Int. J. Mol. Sci. 2022, 23, 16082. https://doi.org/10.3390/ijms232416082
Li Z, Rao MJ, Li J, Wang Y, Chen P, Yu H, Ma C, Wang L. CRISPR/Cas9 Mutant Rice Ospmei12 Involved in Growth, Cell Wall Development, and Response to Phytohormone and Heavy Metal Stress. International Journal of Molecular Sciences. 2022; 23(24):16082. https://doi.org/10.3390/ijms232416082
Chicago/Turabian StyleLi, Zhaoyang, Muhammad Junaid Rao, Jiaying Li, Yanting Wang, Peng Chen, Hua Yu, Chongjian Ma, and Lingqiang Wang. 2022. "CRISPR/Cas9 Mutant Rice Ospmei12 Involved in Growth, Cell Wall Development, and Response to Phytohormone and Heavy Metal Stress" International Journal of Molecular Sciences 23, no. 24: 16082. https://doi.org/10.3390/ijms232416082
APA StyleLi, Z., Rao, M. J., Li, J., Wang, Y., Chen, P., Yu, H., Ma, C., & Wang, L. (2022). CRISPR/Cas9 Mutant Rice Ospmei12 Involved in Growth, Cell Wall Development, and Response to Phytohormone and Heavy Metal Stress. International Journal of Molecular Sciences, 23(24), 16082. https://doi.org/10.3390/ijms232416082