
Statins Show Anti-Atherosclerotic Effects by Improving Endothelial Cell Function in a Kawasaki Disease-like Vasculitis Mouse Model

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Immunohistochemical Staining of Frozen Sections of Ascending Aortic Tunica Intima
2.2. Scoring the Intensity of eNOS Expression in Ascending Aortic Tunica Intima
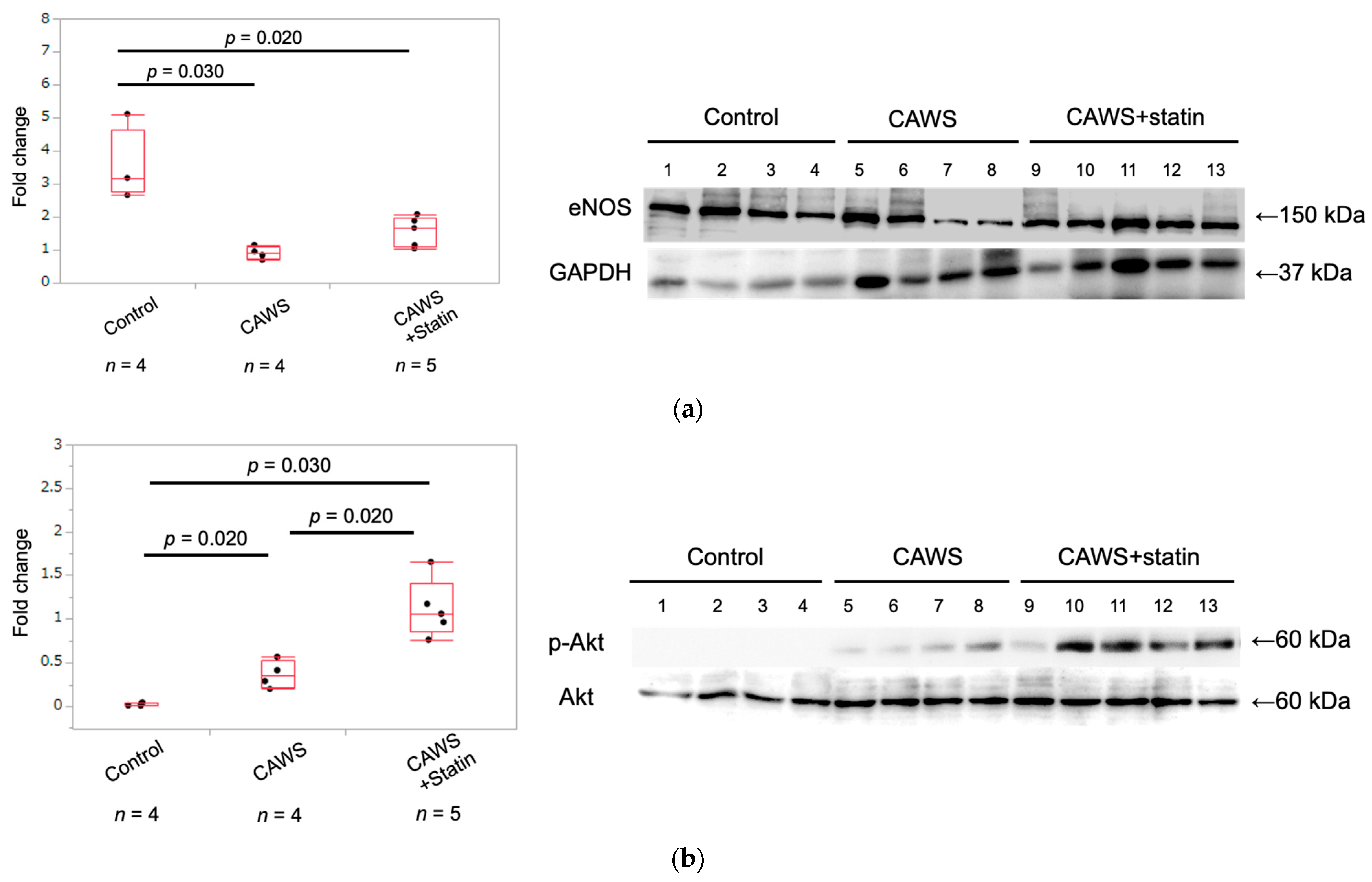
2.3. Expression of eNOS Protein in Whole Aortic Tissue
2.4. Expression of Phospho-Akt and Akt Protein in Whole Aortic Tissue
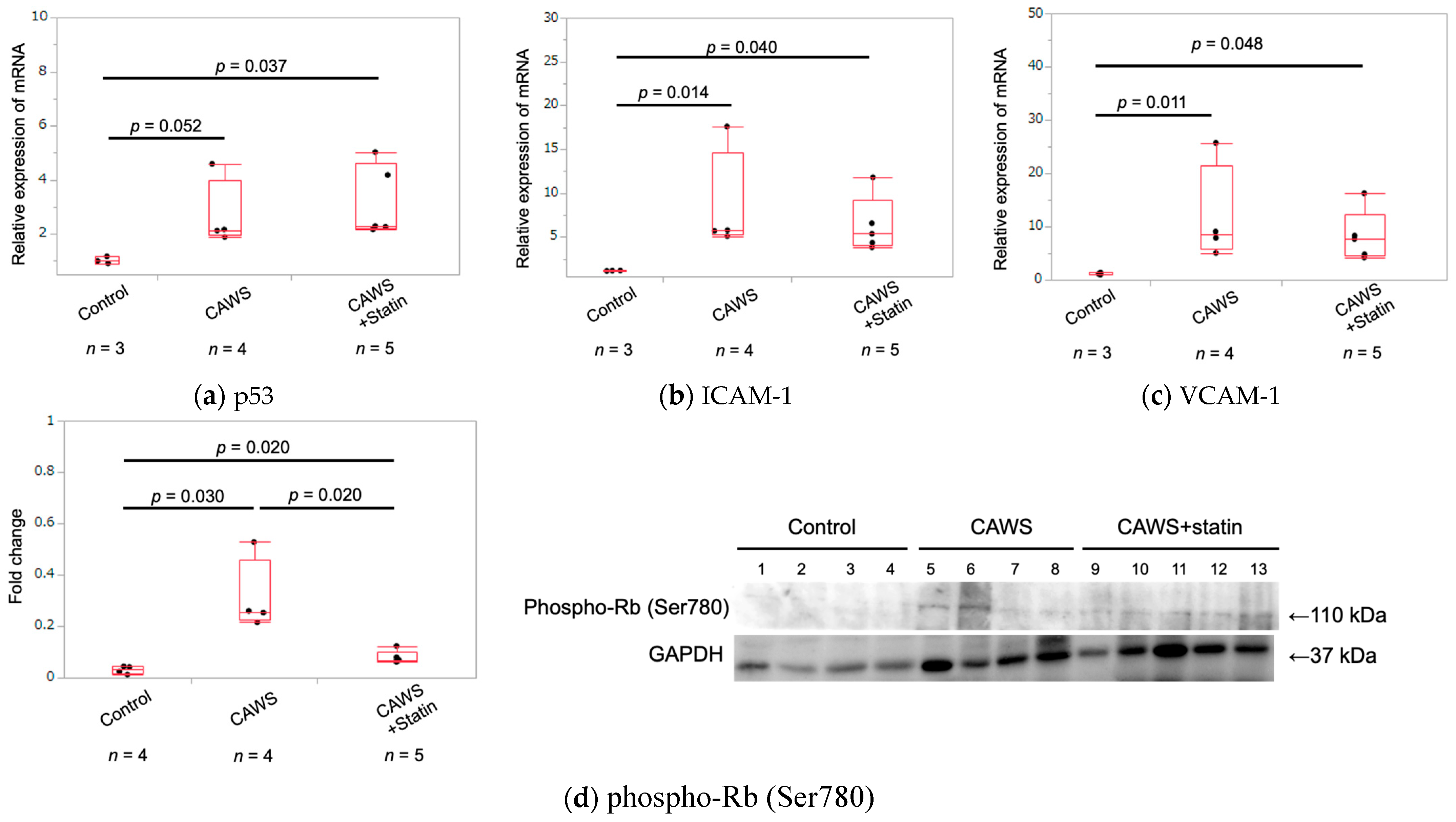
2.5. Expression of Cellular Senescence Markers in Whole Aortic Tissue
2.6. Serum hs-CRP Levels in Each Group
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Preparation of CAWS and Statin
4.3. Experimental Procedures
4.4. Assessment of Horizontal Transections of Ascending Aorta
4.5. Western Blot Analysis of Proteins Extracted from Aortic Tissue
4.6. Antibodies and Materials
4.7. RNA Isolation and Quantitative PCR
4.8. Serological Evaluation
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kawasaki, T.; Kosaki, F.; Okawa, S.; Shigematsu, I.; Yanagawa, H. A New Infantile Acute Febrile Mucocutaneous Lymph Node Syndrome (MLNS) Prevailing in Japan. Pediatrics 1974, 54, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Fukazawa, R.; Kobayashi, J.; Ayusawa, M.; Hamada, H.; Miura, M.; Mitani, Y.; Tsuda, E.; Nakajima, H.; Matsuura, H.; Ikeda, K.; et al. JCS/JSCS 2020 Guideline on Diagnosis and Management of Cardiovascular Sequelae in Kawasaki Disease. Circ. J. 2020, 84, 1348–1407. [Google Scholar] [CrossRef] [PubMed]
- Burns, J.C.; Glodé, M.P. Kawasaki Syndrome. Lancet 2004, 364, 533–544. [Google Scholar] [CrossRef] [PubMed]
- Ueno, K.; Ninomiya, Y.; Hazeki, D.; Masuda, K.; Nomura, Y.; Kawano, Y. Disruption of Endothelial Cell Homeostasis Plays a Key Role in the Early Pathogenesis of Coronary Artery Abnormalities in Kawasaki Disease. Sci. Rep. 2017, 7, 43719. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Y.-Y.; Chen, F.; Zhang, Y.; Ji, X. Are Patients Recovering from Kawasaki Disease at Increased Risk for Accelerated Atherosclerosis? A Meta-Analysis. World J. Pediatr. 2021, 17(5), 476–483. [Google Scholar] [CrossRef] [PubMed]
- Fukazawa, R.; Ikegam, E.; Watanabe, M.; Hajikano, M.; Kamisago, M.; Katsube, Y.; Yamauchi, H.; Ochi, M.; Ogawa, S. Coronary Artery Aneurysm Induced by Kawasaki Disease in Children Show Features Typical Senescence. Circ. J. 2007, 71, 709–715. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Lee, Y.; Crother, T.R.; Fishbein, M.; Zhang, W.; Yilmaz, A.; Shimada, K.; Schulte, D.J.; Lehman, T.J.A.; Shah, P.K.; et al. Marked Acceleration of Atherosclerosis After Lactobacillus Casei–Induced Coronary Arteritis in a Mouse Model of Kawasaki Disease. Arterioscler. Thromb. Vasc. Biol. 2012, 32, e60–e71. [Google Scholar] [CrossRef] [Green Version]
- Motoji, Y.; Fukazawa, R.; Matsui, R.; Nagi-Miura, N.; Miyagi, Y.; Itoh, Y.; Ishii, Y. Kawasaki Disease-like Vasculitis Facilitates Atherosclerosis, and Statin Shows a Significant Antiatherosclerosis and Anti-Inflammatory Effect in a Kawasaki Disease Model Mouse. Biomedicines 2022, 10, 1794. [Google Scholar] [CrossRef]
- Xu, H.; Jiang, J.; Chen, W.; Li, W.; Chen, Z. Vascular Macrophages in Atherosclerosis. J. Immunol. Res. 2019, 2019, 4354786. [Google Scholar] [CrossRef] [Green Version]
- Shao, Y.; Saredy, J.; Yang, W.Y.; Sun, Y.; Lu, Y.; Saaoud, F.; Drummer, C., 4th; Johnson, C.; Xu, K.; Jiang, X.; et al. Vascular Endothelial Cells and Innate Immunity. Arterioscler. Thromb. Vasc. Biol. 2020, 40, e138–e152. [Google Scholar] [CrossRef]
- Checkouri, E.; Blanchard, V.; Meilhac, O. Macrophages in Atherosclerosis, First or Second Row Players? Biomedicines 2021, 9, 1214. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, Y.; Raines, E.W.; Plump, A.S.; Breslow, J.L.; Ross, R. Upregulation of VCAM-1 and ICAM-1 at Atherosclerosis-Prone Sites on the Endothelium in the ApoE-Deficient Mouse. Arterioscler. Thromb. Vasc. Biol. 1998, 18, 842–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, G.; Nystrom, F.H.; Ravichandran, L.V.; Cong, L.N.; Kirby, M.; Mostowski, H.; Quon, M.J. Roles for Insulin Receptor, PI3-Kinase, and Akt in Insulin-Signaling Pathways Related to Production of Nitric Oxide in Human Vascular Endothelial Cells. Circulation 2000, 101, 1539–1545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heiss, C.; Rodriguez-Mateos, A.; Kelm, M. Central Role of ENOS in the Maintenance of Endothelial Homeostasis. Antioxid. Redox Signal. 2015, 22, 1230–1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanda, R.; Iwasaki, K.; Yihao, P.; Umeda, M.; Fujino, T.; Kawazoe, T. Plasmalogen Attenuates LPS-Induced ICAM-1 Expression In Endothelial Cell Line. J. Osaka Dent. Univ. 2021, 55, 179–185. [Google Scholar]
- Dimmeler, S.; Aicher, A.; Vasa, M.; Mildner-Rihm, C.; Adler, K.; Tiemann, M.; Rütten, H.; Fichtlscherer, S.; Martin, H.; Zeiher, A.M. HMG-CoA Reductase Inhibitors (Statins) Increase Endothelial Progenitor Cells via the PI 3-Kinase/Akt Pathway. J. Clin. Investig. 8 2001, 108, 391–397. [Google Scholar] [CrossRef]
- Muhammad, R.N.; Sallam, N.; El-Abhar, H.S. Activated ROCK/Akt/ENOS and ET-1/ERK Pathways in 5-Fluorouracil-Induced Cardiotoxicity: Modulation by Simvastatin. Sci. Rep. 2020, 10, 14693. [Google Scholar] [CrossRef]
- Heinisch, P.P.; Bello, C.; Emmert, M.Y.; Carrel, T.; Dreßen, M.; Hörer, J.; Winkler, B.; Luedi, M.M. Endothelial Progenitor Cells as Biomarkers of Cardiovascular Pathologies: A Narrative Review. Cells 2022, 11, 1678. [Google Scholar] [CrossRef]
- Donato, A.J.; Morgan, R.G.; Walker, A.E.; Lesniewski, L.A. Cellular and Molecular Biology of Aging Endothelial Cells. J. Mol. Cell. Cardiol. 2015, 89, 122–135. [Google Scholar] [CrossRef] [Green Version]
- Bui, T.M.; Wiesolek, H.L.; Sumagin, R. ICAM-1: A Master Regulator of Cellular Responses in Inflammation, Injury Resolution, and Tumorigenesis. J. Leukoc. Biol. 2020, 108, 787–799. [Google Scholar] [CrossRef]
- Di Micco, R.; Krizhanovsky, V.; Baker, D.; d’Adda di Fagagna, F. Cellular Senescence in Ageing: From Mechanisms to Therapeutic Opportunities. Nat. Rev. Mol. Cell Biol. 2020, 22, 75–95. [Google Scholar] [CrossRef]
- Nachtigal, P.; Pospisilova, N.; Jamborova, G.; Pospechova, K.; Solichova, D.; Andrys, C.; Zdansky, P.; Micuda, S.; Semecky, V. Atorvastatin Has Hypolipidemic and Anti-Inflammatory Effects in ApoE/LDL Receptor-Double-Knockout Mice. Life Sci. 2008, 82, 708–717. [Google Scholar] [CrossRef]
- Oesterle, A.; Laufs, U.; Liao, J.K. Pleiotropic Effects of Statins on the Cardiovascular System. Circ. Res. 2017, 120, 229–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiojima, I.; Walsh, K. Role of Akt Signaling in Vascular Homeostasis and Angiogenesis. Circ. Res. 2002, 90, 1243–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eto, M.; Kozai, T.; Cosentino, F.; Joch, H.; Lüscher, T.F. Statin Prevents Tissue Factor Expression in Human Endothelial Cells: Role of Rho/Rho-Kinase and Akt Pathways. Circulation 2002, 105, 1756–1759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, N.; Hayashi, Y.; Takahashi, Y.; Hattori, Y. Phosphorylation of Endothelial Nitric-Oxide Synthase Is Diminished in Mesenteric Arteries from Septic Rabbits Depending on the Altered Phosphatidylinositol 3-Kinase/Akt Pathway: Reversal Effect of Fluvastatin Therapy. J. Pharmacol. Exp. Ther. 2006, 319, 1348–1354. [Google Scholar] [CrossRef] [Green Version]
- Fruman, D.A.; Chiu, H.; Hopkins, B.D.; Bagrodia, S.; Cantley, L.C.; Abraham, R.T. The PI3K Pathway in Human Disease. Cell 2017, 170, 605–635. [Google Scholar] [CrossRef] [Green Version]
- Kureishi, Y.; Luo, Z.; Shiojima, I.; Bialik, A.; Fulton, D.; Lefer, D.J.; Sessa, W.C.; Walsh, K. The HMG-CoA Reductase Inhibitor Simvastatin Activates the Protein Kinase Akt and Promotes Angiogenesis in Normocholesterolemic Animals. Nat. Med. 2000, 6, 1004–1010. [Google Scholar] [CrossRef]
- De Oliveira, T.S.; de Oliveira, L.M.; de Oliveira, L.P.; da Costa, R.M.; Tostes, R.d.C.; Georg, R.d.C.; Costa, E.A.; Lobato, N.d.S.; Filgueira, F.P.; Ghedini, P.C. Activation of PI3K/Akt Pathway Mediated by Estrogen Receptors Accounts for Estrone-Induced Vascular Activation of CGMP Signaling. Vasc. Pharmacol. 2018, 110, 42–48. [Google Scholar] [CrossRef]
- Huang, W.-P.; Chen, C.-Y.; Lin, T.-W.; Kuo, C.-S.; Huang, H.-L.; Huang, P.-H.; Lin, S.-J. Fibroblast Growth Factor 21 Reverses High-Fat Diet-Induced Impairment of Vascular Function via the Anti-Oxidative Pathway in ApoE Knockout Mice. J. Cell. Mol. Med. 2022, 26, 2451–2461. [Google Scholar] [CrossRef]
- Troutman, T.D.; Bazan, J.F.; Pasare, C. Toll-like Receptors, Signaling Adapters and Regulation of the pro-Inflammatory Response by PI3K. Cell Cycle 2012, 11, 3559–3567. [Google Scholar] [CrossRef] [PubMed]
- Covarrubias, A.J.; Aksoylar, H.I.; Horng, T. Control of Macrophage Metabolism and Activation by MTOR and Akt Signaling. Semin. Immunol. 2015, 27, 286–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cianciulli, A.; Calvello, R.; Porro, C.; Trotta, T.; Salvatore, R.; Panaro, M.A. PI3k/Akt Signalling Pathway Plays a Crucial Role in the Anti-Inflammatory Effects of Curcumin in LPS-Activated Microglia. Int. Immunopharmacol. 2016, 36, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.M.; Shimizu, C.; Oharaseki, T.; Takahashi, K.; Daniels, L.B.; Kahn, A.; Adamson, R.; Dembitsky, W.; Gordon, J.B.; Burns, J.C. Role of TGF-β Signaling in Remodeling of Noncoronary Artery Aneurysms in Kawasaki Disease. Pediatr. Dev. Pathol. 2015, 18, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; Zhao, X.; Zhang, L.; Zhang, J.; L’Huillier, A.; Ling, W.; Roberts, A.I.; Le, A.D.; Shi, S.; Shao, C.; et al. Inflammatory Cytokine-Induced Intercellular Adhesion Molecule-1 and Vascular Cell Adhesion Molecule-1 in Mesenchymal Stem Cells Are Critical for Immunosuppression. J. Immunol. 2010, 184, 2321–2328. [Google Scholar] [CrossRef] [Green Version]
- Khaidakov, M.; Wang, X.; Mehta, J.L. Potential Involvement of LOX-1 in Functional Consequences of Endothelial Senescence. PLoS ONE 2011, 6, e20964. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Shen, H.; Schenten, D.; Shan, P.; Lee, P.J.; Goldstein, D.R. Aging Enhances the Basal Production of IL-6 and CCL2 in Vascular Smooth Muscle Cells. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 103–109. [Google Scholar] [CrossRef] [Green Version]
- Warboys, C.M.; de Luca, A.; Amini, N.; Luong, L.; Duckles, H.; Hsiao, S.; White, A.; Biswas, S.; Khamis, R.; Chong, C.K.; et al. Disturbed Flow Promotes Endothelial Senescence via a P53-Dependent Pathway. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 985–995. [Google Scholar] [CrossRef] [Green Version]
- Merdji, H.; Kassem, M.; Chomel, L.; Clere-Jehl, R.; Helms, J.; Kurihara, K.; Chaker, A.B.; Auger, C.; Schini-Kerth, V.; Toti, F.; et al. Septic Shock as a Trigger of Arterial Stress-Induced Premature Senescence: A New Pathway Involved in the Post Sepsis Long-Term Cardiovascular Complications. Vascul. Pharmacol. 2021, 141, 106922. [Google Scholar] [CrossRef]
- Hayflick, L.; Moorhead, P.S. The Serial Cultivation of Human Diploid Cell Strains. Exp. Cell Res. 1961, 25, 585–621. [Google Scholar] [CrossRef]
- Baker, D.J.; Childs, B.G.; Durik, M.; Wijers, M.E.; Sieben, C.J.; Zhong, J.; Saltness, R.A.; Jeganathan, K.B.; Verzosa, G.C.; Pezeshki, A.; et al. Naturally Occurring P16Ink4a-Positive Cells Shorten Healthy Lifespan. Nature 2016, 530, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Hara, E.; Smith, R.; Parry, D.; Tahara, H.; Stone, S.; Peters, G. Regulation of P16CDKN2 Expression and Its Implications for Cell Immortalization and Senescence. Mol. Cell. Biol. 1996, 16, 859–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viallard, J.F.; Lacombe, F.; Belloc, F.; Pellegrin, J.L.; Reiffers, J. Molecular mechanisms controlling the cell cycle: Fundamental aspects and implications for oncology. Cancer Radiother. 2001, 5, 109–129. [Google Scholar] [CrossRef]
- Levine, A.J.; Oren, M. The First 30 Years of P53: Growing Ever More Complex. Nat. Rev. Cancer 2009, 9, 749–758. [Google Scholar] [CrossRef] [Green Version]
- Meek, D.W.; Anderson, C.W. Posttranslational Modification of P53: Cooperative Integrators of Function. Cold Spring Harb. Perspect. Biol. 2009, 1, a000950. [Google Scholar] [CrossRef] [Green Version]
- Nag, S.; Qin, J.; Srivenugopal, K.S.; Wang, M.; Zhang, R. The MDM2-P53 Pathway Revisited. J. Biomed. Res. 2013, 27, 254–271. [Google Scholar] [CrossRef]
- Dustin, M.L.; Rothlein, R.; Bhan, A.K.; Dinarello, C.A.; Springer, T.A. Induction by IL 1 and Interferon-Gamma: Tissue Distribution, Biochemistry, and Function of a Natural Adherence Molecule (ICAM-1). J. Immunol. 1986, 137, 245–254. [Google Scholar]
- Troncoso, M.F.; Ortiz-Quintero, J.; Garrido-Moreno, V.; Sanhueza-Olivares, F.; Guerrero-Moncayo, A.; Chiong, M.; Castro, P.F.; García, L.; Gabrielli, L.; Corbalán, R.; et al. VCAM-1 as a Predictor Biomarker in Cardiovascular Disease. Biochim. Biophys. Acta Mol. Basis Dis. 2021, 1867, 166170. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Fukazawa, R.; Nagi-Miura, N.; Ohno, N.; Suzuki, N.; Katsube, Y.; Kamisago, M.; Akao, M.; Watanabe, M.; Hashimoto, K.; et al. Interleukin-1beta Inhibition Attenuates Vasculitis in a Mouse Model of Kawasaki Disease. J. Nippon Med. Sch. 2019, 86, 108–116. [Google Scholar] [CrossRef] [Green Version]
- Gibson, E.M.; Henson, E.S.; Haney, N.; Villanueva, J.; Gibson, S.B. Epidermal Growth Factor Protects Epithelial-Derived Cells from Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand-Induced Apoptosis by Inhibiting Cytochrome c Release. Cancer Res. 2002, 62, 488–496. [Google Scholar]
- Kim, E.-C.; Yun, B.-S.; Ryoo, I.-J.; Min, J.-K.; Won, M.-H.; Lee, K.-S.; Kim, Y.-M.; Yoo, I.-D.; Kwon, Y.-G. Complestatin Prevents Apoptotic Cell Death: Inhibition of a Mitochondrial Caspase Pathway through AKT/PKB Activation. Biochem. Biophys. Res. Commun. 2004, 313, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Ouyang, G.; Bao, S. The Activation of Akt/PKB Signaling Pathway and Cell Survival. J. Cell. Mol. Med. 2005, 9, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Yanae, M.; Tsubaki, M.; Satou, T.; Itoh, T.; Imano, M.; Yamazoe, Y.; Nishida, S. Statin-Induced Apoptosis via the Suppression of ERK1/2 and Akt Activation by Inhibition of the Geranylgeranyl-Pyrophosphate Biosynthesis in Glioblastoma. J. Exp. Clin. Cancer Res. 2011, 30, 74. [Google Scholar] [CrossRef] [PubMed]
- NC3Rs Reporting Guidelines Working Group Animal Research: Reporting in Vivo Experiments: The ARRIVE Guidelines. Exp. Physiol. 2010, 95, 842–844. [CrossRef] [Green Version]
- Uchiyama, M.; Ohno, N.; Miura, N.N.; Adachi, Y.; Aizawa, M.W.; Tamura, H.; Tanaka, S.; Yadomae, T. Chemical and Immunochemical Characterization of Limulus Factor G-Activating Substance OfCandidaspp. FEMS Immunol. Med. Microbiol. 1999, 24, 411–420. [Google Scholar] [CrossRef]
- Flurkey, K.; Mcurrer, J.; Harrison, D. Mouse Models in Aging Research. In The Mouse in Biomedical Research; Elsevier: Amsterdam, The Netherlands, 2007; pp. 637–672. [Google Scholar]
- Hirata, N.; Ishibashi, K.-I.; Usui, T.; Yoshioka, J.; Hata, S.; Adachi, Y.; Nagi-Miura, N.; Ohta, S.; Ohno, N. A Model of Left Ventricular Dysfunction Complicated by CAWS Arteritis in DBA/2 Mice. Int. J. Vasc. Med. 2012, 2012, 570297. [Google Scholar] [CrossRef]
- Palliyaguru, D.L.; Vieira Ligo Teixeira, C.; Duregon, E.; di Germanio, C.; Alfaras, I.; Mitchell, S.J.; Navas-Enamorado, I.; Shiroma, E.J.; Studenski, S.; Bernier, M.; et al. Study of Longitudinal Aging in Mice: Presentation of Experimental Techniques. J. Gerontol. A Biol. Sci. Med. Sci. 2021, 76, 552–560. [Google Scholar] [CrossRef]
- Takahashi, K.; Oharaseki, T.; Yokouchi, Y.; Miura, N.N.; Ohno, N.; Okawara, A.I.; Murata, H.; Naoe, S.; Suzuki, K. Administration of Human Immunoglobulin Suppresses Development of Murine Systemic Vasculitis Induced with Candida Albicans Water-Soluble Fraction: An Animal Model of Kawasaki Disease. Mod. Rheumatol. 2010, 20, 160–167. [Google Scholar] [CrossRef]
- Abe, Y.; Suzuki, Y.; Kawamura, K.; Tanaka, N. MEP50/PRMT5-Mediated Methylation Activates GLI1 in Hedgehog Signalling through Inhibition of Ubiquitination by the ITCH/NUMB Complex. Commun. Biol. 2019, 2, 23. [Google Scholar] [CrossRef] [Green Version]
- Uehara, I.; Kajita, M.; Tanimura, A.; Hida, S.; Onda, M.; Naito, Z.; Taki, S.; Tanaka, N. 2-Deoxy-d-Glucose Induces Deglycosylation of Proinflammatory Cytokine Receptors and Strongly Reduces Immunological Responses in Mouse Models of Inflammation. Pharmacol. Res. Perspect. 2022, 10, e00940. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 10 Weeks after CAWS Administration | ||||||
|---|---|---|---|---|---|---|
| Control | CAWS | CAWS+Statin | ||||
| Body weight (g) | 32.6 ± 2.9 | 29.1 ± 2.7 | 25.6 ± 2.5 * | |||
| (n = 5) | (n = 5) | (n = 5) | ||||
| Spleen weight (g) | 0.14 ± 0.01 | 0.29 ± 0.05 * | 0.22 ± 0.04 ** | |||
| (n = 5) | (n = 5) | (n = 5) | ||||
| Spleen/Body weight ratio(%) | 0.44 ± 0.04 | 0.98 ± 0.11 * | 0.87 ± 0.11 * | |||
| (n = 5) | (n = 5) | (n = 5) | ||||
| Serological Evaluation | ||||||
| hs-CRP (pg/mL) | 86.8 ± 15.3 | 1127.2 ± 969.6 * | 840.4 ± 776.6 * | |||
| (n = 5) | (n = 5) | (n = 5) | ||||
| Immunohistochemical staining | ||||||
| eNOS scoring in ascending aorta | 25.0 | [27.7, 21] | 2.3 | [9.2, 1.7] * | 20.0 | [22.5, 17.5] ** |
| (n = 5) | (n = 5) | (n = 5) | ||||
| Western blot analysis for Aortic tissue | ||||||
| Expression of eNOS protein | 3.5 ± 1.1 | 0.9 ± 0.2 * | 1.6 ± 0.5 * | |||
| (n = 4) | (n = 4) | (n = 5) | ||||
| Phospho-Akt/Akt ratio | 0.02 ± 0.01 | 0.4 ± 0.16 * | 1.12 ± 0.33 *, ** | |||
| (n = 4) | (n = 4) | (n = 5) | ||||
| Expression of phosoho-Rb(Ser780) | 0.03 ± 0.02 | 0.31 ± 0.14 * | 0.08 ± 0.03 *, ** | |||
| (n = 4) | (n = 4) | (n = 5) | ||||
| Quantitative PCR analysis for Aortic tissue | ||||||
| p53 | 1.1 ± 0.1 | 2.9 ± 1.4 * | 3.4 ± 1.4 * | |||
| (n = 3) | (n = 4) | (n = 5) | ||||
| ICAM-1 | 1.2 ± 0.02 | 8.5 ± 6.0 * | 1.4 ± 3.2 * | |||
| (n = 3) | (n = 4) | (n = 5) | ||||
| VCAM-1 | 1.2 ± 0.2 | 11.9 ± 9.4 * | 8.2 ± 4.8 * | |||
| (n = 3) | (n = 4) | (n = 5) | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Motoji, Y.; Fukazawa, R.; Matsui, R.; Abe, Y.; Uehara, I.; Watanabe, M.; Hashimoto, Y.; Miyagi, Y.; Nagi-Miura, N.; Tanaka, N.; et al. Statins Show Anti-Atherosclerotic Effects by Improving Endothelial Cell Function in a Kawasaki Disease-like Vasculitis Mouse Model. Int. J. Mol. Sci. 2022, 23, 16108. https://doi.org/10.3390/ijms232416108
Motoji Y, Fukazawa R, Matsui R, Abe Y, Uehara I, Watanabe M, Hashimoto Y, Miyagi Y, Nagi-Miura N, Tanaka N, et al. Statins Show Anti-Atherosclerotic Effects by Improving Endothelial Cell Function in a Kawasaki Disease-like Vasculitis Mouse Model. International Journal of Molecular Sciences. 2022; 23(24):16108. https://doi.org/10.3390/ijms232416108
Chicago/Turabian StyleMotoji, Yusuke, Ryuji Fukazawa, Ryosuke Matsui, Yoshinori Abe, Ikuno Uehara, Makoto Watanabe, Yoshiaki Hashimoto, Yasuo Miyagi, Noriko Nagi-Miura, Nobuyuki Tanaka, and et al. 2022. "Statins Show Anti-Atherosclerotic Effects by Improving Endothelial Cell Function in a Kawasaki Disease-like Vasculitis Mouse Model" International Journal of Molecular Sciences 23, no. 24: 16108. https://doi.org/10.3390/ijms232416108
APA StyleMotoji, Y., Fukazawa, R., Matsui, R., Abe, Y., Uehara, I., Watanabe, M., Hashimoto, Y., Miyagi, Y., Nagi-Miura, N., Tanaka, N., & Ishii, Y. (2022). Statins Show Anti-Atherosclerotic Effects by Improving Endothelial Cell Function in a Kawasaki Disease-like Vasculitis Mouse Model. International Journal of Molecular Sciences, 23(24), 16108. https://doi.org/10.3390/ijms232416108