
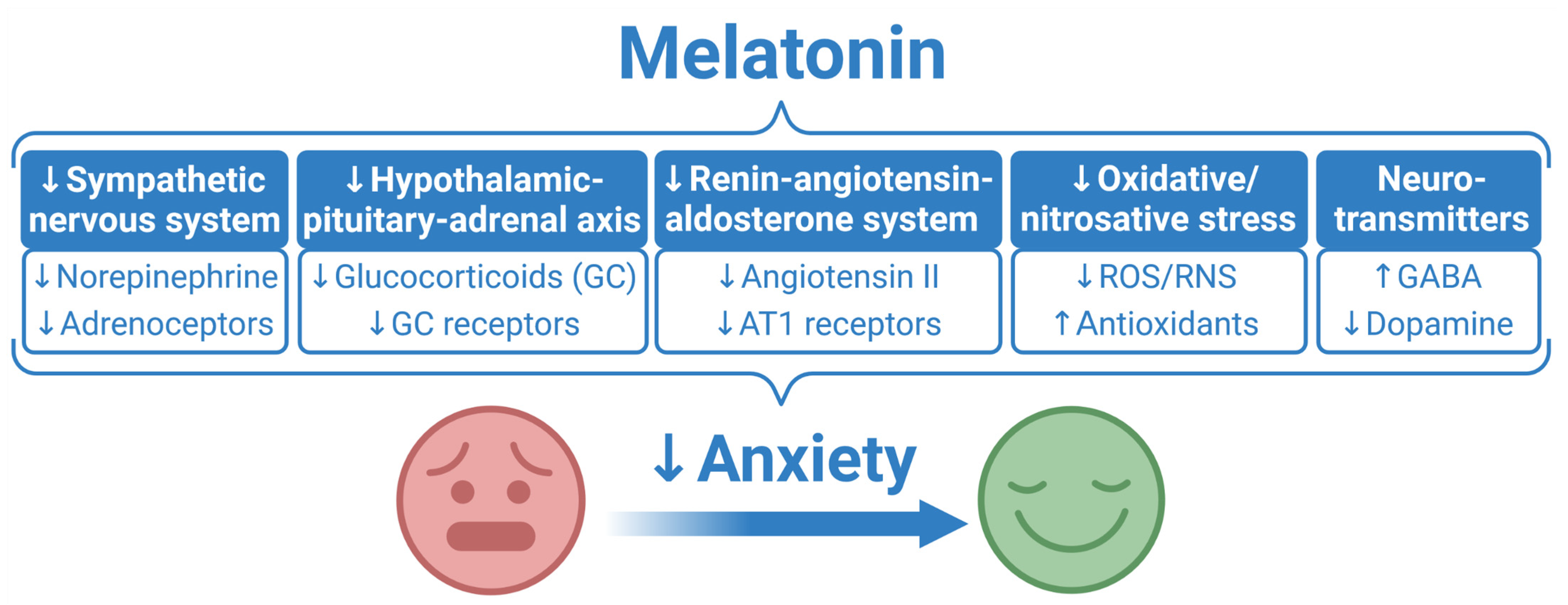
Melatonin as a Potential Approach to Anxiety Treatment
 , , ,
, , ,  and
and 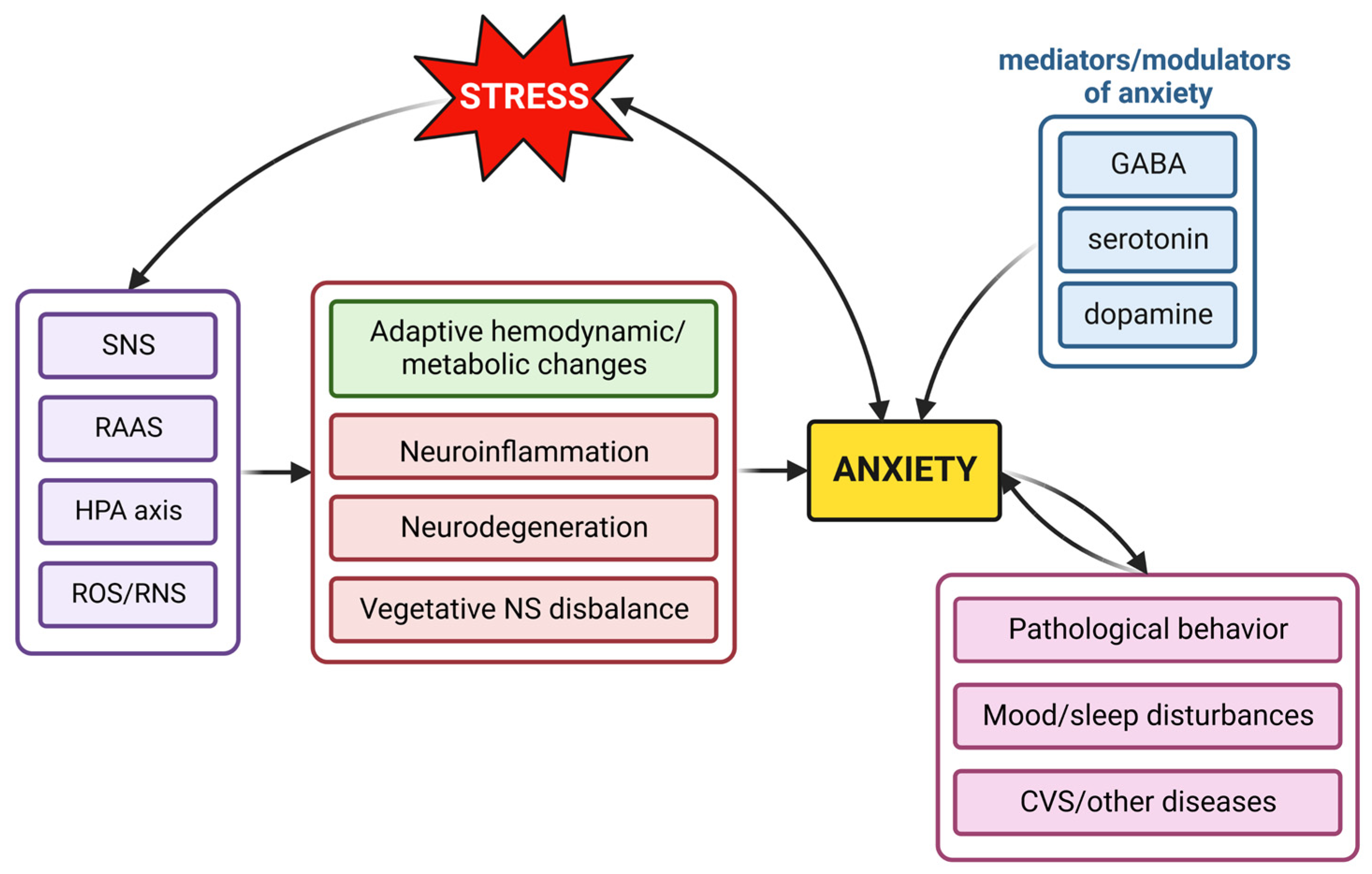{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Neurohumoral Mechanisms Contributing to Anxiety
2.1. The Role of the Sympathetic Nervous System in Anxiety
2.2. The Hypothalamus–Pituitary–Adrenal Axis in Anxiety
2.3. The Impact of the Renin–Angiotensin–Aldosterone System in Anxiety
2.4. Reactive Oxygen and Nitrogen Species in Anxiety Development
2.5. The Alterations of Neurotransmitters in Anxiety
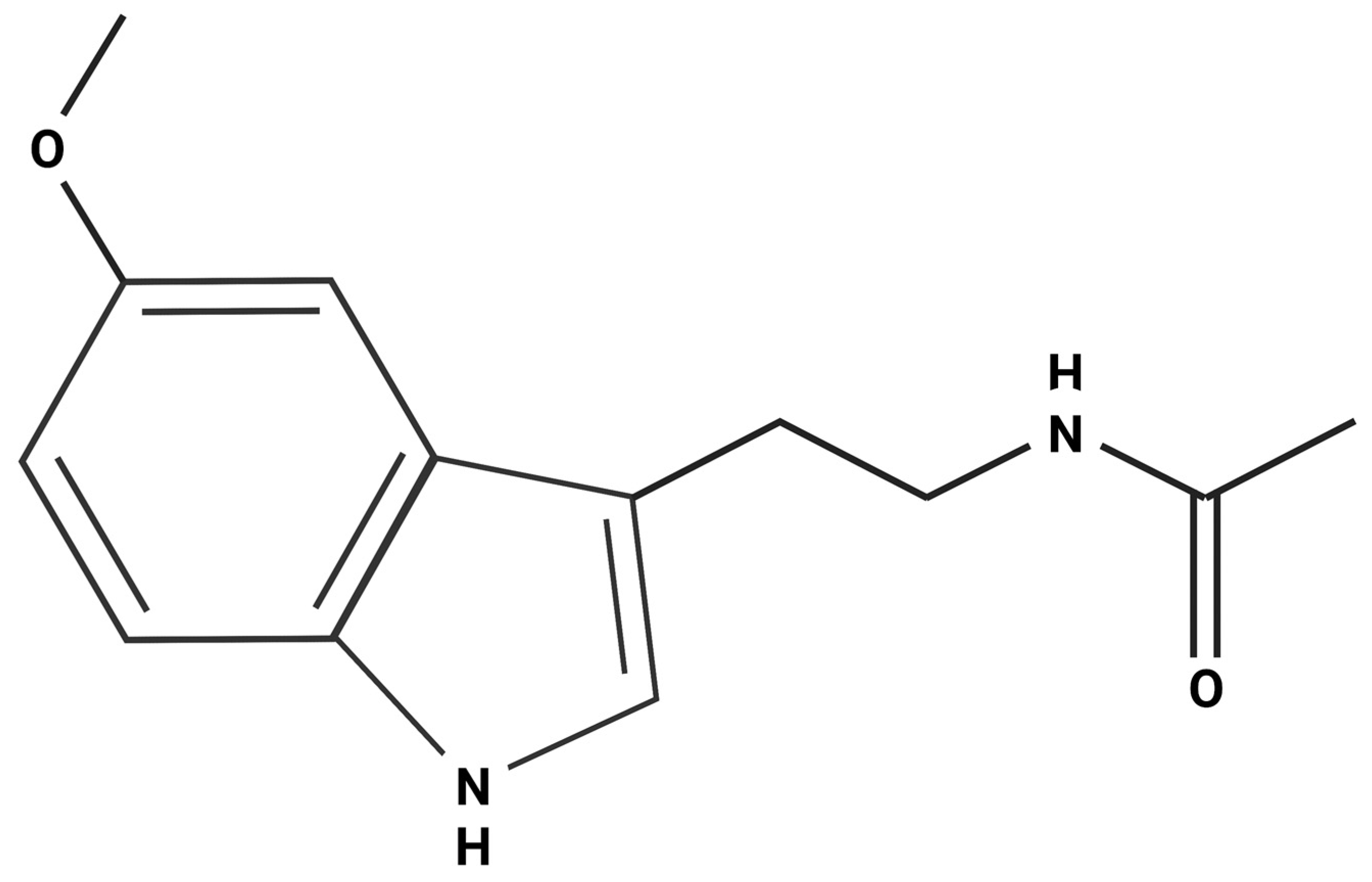
3. Melatonin: Synthesis, Phylogenetic Considerations, Signaling Pathways and Effects
3.1. Melatonin Synthesis, Evolution and Signaling Pathways
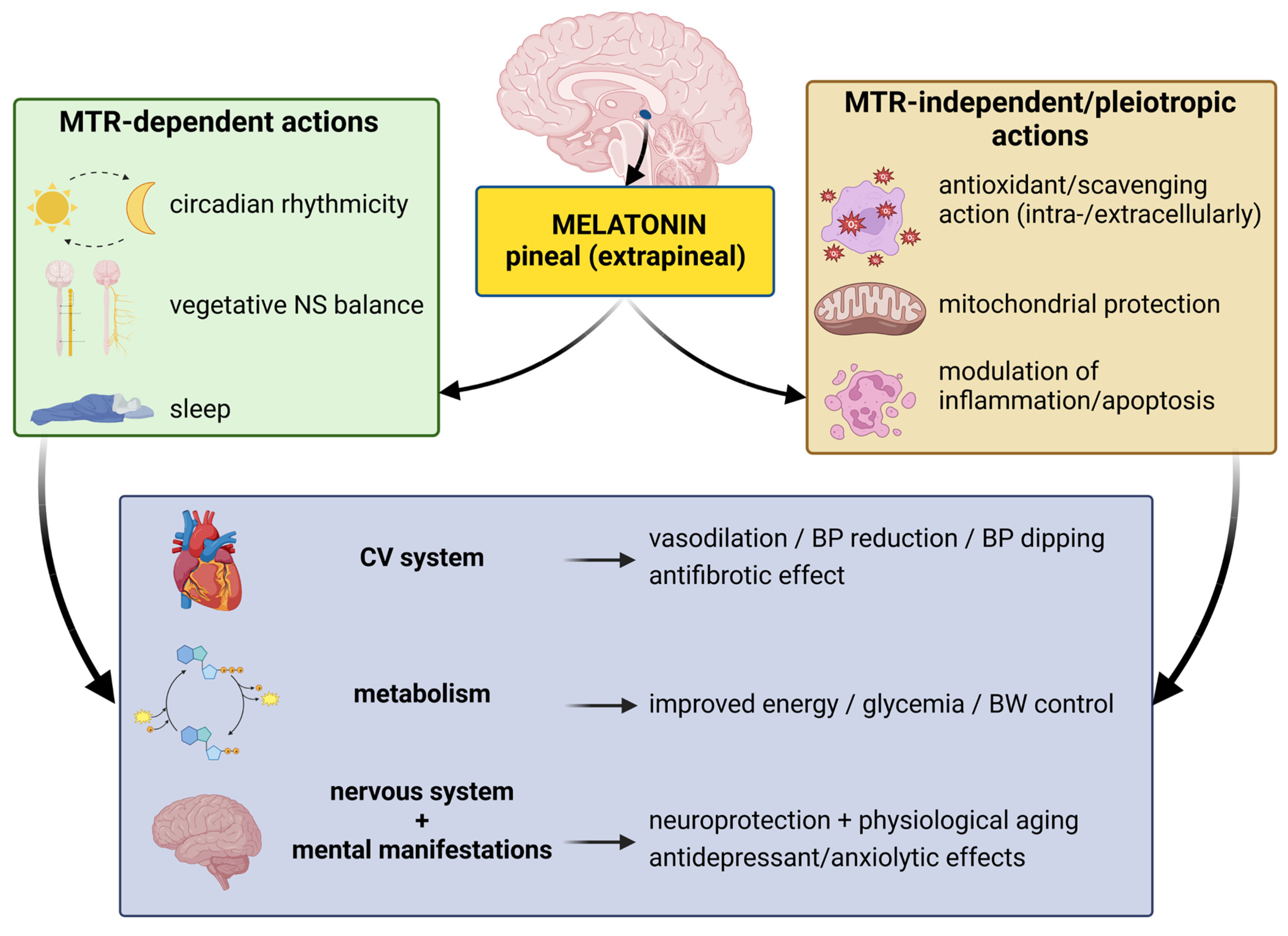
3.2. Pleiotropic Receptor-Independent Actions
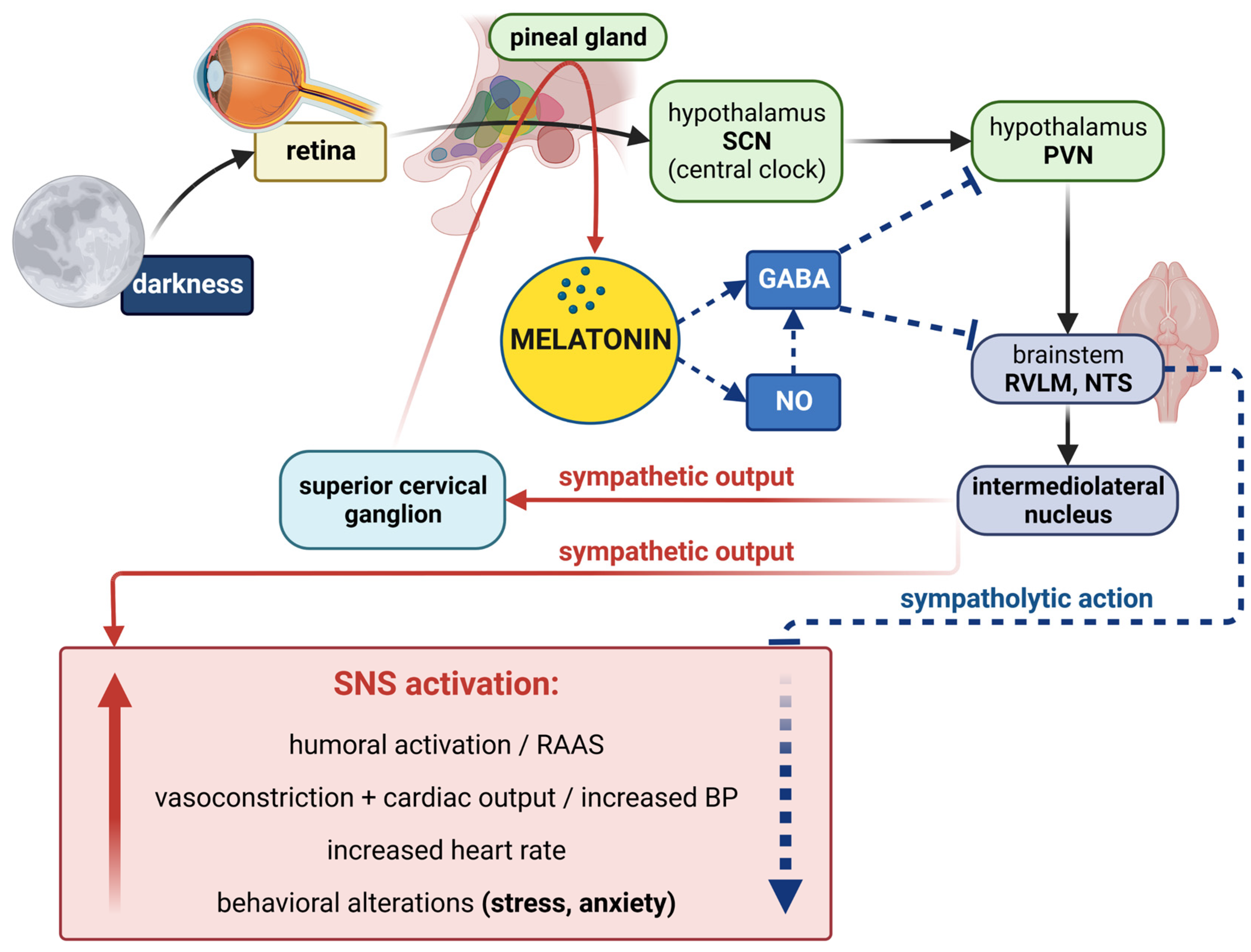
3.3. Melatonin and Circadian Rhythmicity
3.4. Anxiolytic Effect of Melatonin
3.4.1. Melatonin–Sympathetic Nervous System Interactions
3.4.2. Potential Interference of Melatonin with the Renin–Angiotensin–Aldosterone System
3.4.3. Melatonin vs. Glucocorticoids in Anxiety
3.4.4. Melatonin Interaction with Oxidative and Nitrosative Stress in Anxiety
3.4.5. Melatonin-Induced Modifications of Neurotransmission in Anxiety
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 3-OHM | 3-hydroxymelatonin |
| 5-HT | 5-hydroxytryptamine |
| 6-OHM | 6-hydroxymelatonin |
| ACE | angiotensin I-converting enzyme |
| ACTH | adrenocorticotropic hormone |
| AFMK | N1-acetyl-N2-formyl-5-methoxykynuramine |
| AMK | N1-acetyl-5-methoxykynuramine |
| Ang II | angiotensin II |
| ARBs | angiotensin receptor blockers |
| AT1 receptor | angiotensin II type 1 receptor |
| ATP | adenosine triphosphate |
| BP | blood pressure |
| BSO | buthionine-S,R-sulfoximine |
| CB1 receptor | cannabinoid 1 receptor |
| CBD | cannabidiol |
| CKD | chronic kidney disease |
| CNS | central nervous system |
| CRF | corticotropin-releasing factor |
| CSF | cerebrospinal fluid |
| DRN | dorsal raphe nucleus |
| DSM | Diagnostic and Statistical Manual of Mental Disorders |
| EPM | elevated plus maze |
| GABA | γ-aminobutyric acid |
| GAD | generalized anxiety disorder |
| HPA axis | hypothalamus–pituitary–adrenal axis |
| HR | heart rate |
| iNOS | inducible nitric oxide synthase |
| LDB | light/dark box |
| L-NAME | L-NG-nitro arginine methyl ester |
| MSNA | muscle sympathetic nerve activity |
| MT1 receptor | melatonin receptor type 1 |
| MT2 receptor | melatonin receptor type 2 |
| NE | norepinephrine |
| nNOS | neuronal nitric oxide synthase |
| NO | nitric oxide |
| NSF | novelty-suppressed feeding |
| OFT | open field test |
| PD | panic disorder |
| PTSD | posttraumatic stress disorder |
| PVN | paraventricular nucleus |
| QR | quinone reductase |
| RAAS | renin–angiotensin–aldosterone system |
| RNS | reactive nitrogen species |
| ROR | retinoid orphan receptors |
| ROS | reactive oxygen species |
| RVLM | rostral ventrolateral medulla |
| RZR | retinoid Z receptors |
| sAA | salivary alpha-amylase |
| SAD | social anxiety disorder |
| SCN | suprachiasmatic nucleus |
| SHR | spontaneously hypertensive rat |
| SNRIs | serotonin norepinephrine reuptake inhibitors |
| SNS | sympathetic nervous system |
| SSRIs | selective serotonin reuptake inhibitors |
References
- Dattani, S.; Ritchie, H.; Roser, M. Mental Health. Available online: https://ourworldindata.org/mental-health (accessed on 3 October 2022).
- Craske, M.G.; Stein, M.B. Anxiety. Lancet 2016, 388, 3048–3059. [Google Scholar] [CrossRef] [PubMed]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Association: Washington, DC, USA, 2013; ISBN 978-0-89042-555-8. [Google Scholar]
- Garakani, A.; Murrough, J.W.; Freire, R.C.; Thom, R.P.; Larkin, K.; Buono, F.D.; Iosifescu, D.V. Pharmacotherapy of anxiety disorders: Current and emerging treatment options. Front. Psychiatry 2020, 11, 595584. [Google Scholar] [CrossRef] [PubMed]
- Bandelow, B. Current and novel psychopharmacological drugs for anxiety disorders. Adv. Exp. Med. Biol. 2020, 1191, 347–365. [Google Scholar] [CrossRef] [PubMed]
- Zareifopoulos, N.; Dylja, I. Efficacy and tolerability of vilazodone for the acute treatment of generalized anxiety disorder: A meta-analysis. Asian J. Psychiatry 2017, 26, 115–122. [Google Scholar] [CrossRef]
- Careri, J.M.; Draine, A.E.; Hanover, R.; Liebowitz, M.R. A 12-week double-blind, placebo-controlled, flexible-dose trial of vilazodone in generalized social anxiety disorder. Prim. Care Companion CNS Disord. 2015, 17, 23678. [Google Scholar] [CrossRef] [Green Version]
- Solmi, M.; Fornaro, M.; Ostinelli, E.G.; Zangani, C.; Croatto, G.; Monaco, F.; Krinitski, D.; Fusar-Poli, P.; Correll, C.U. Safety of 80 antidepressants, antipsychotics, anti-attention-deficit/hyperactivity medications and mood stabilizers in children and adolescents with psychiatric disorders: A large scale systematic meta-review of 78 adverse effects. World Psychiatry 2020, 19, 214–232. [Google Scholar] [CrossRef]
- Zarate, C.A.; Singh, J.B.; Carlson, P.J.; Brutsche, N.E.; Ameli, R.; Luckenbaugh, D.A.; Charney, D.S.; Manji, H.K. A randomized trial of an N-methyl-D-aspartate antagonist in treatment-resistant major depression. Arch. Gen. Psychiatry 2006, 63, 856–864. [Google Scholar] [CrossRef]
- Glue, P.; Neehoff, S.M.; Medlicott, N.J.; Gray, A.; Kibby, G.; McNaughton, N. Safety and efficacy of maintenance ketamine treatment in patients with treatment-refractory generalised anxiety and social anxiety disorders. J. Psychopharmacol. 2018, 32, 663–667. [Google Scholar] [CrossRef]
- Hartberg, J.; Garrett-Walcott, S.; De Gioannis, A. Impact of oral ketamine augmentation on hospital admissions in treatment-resistant depression and PTSD: A retrospective study. Psychopharmacology 2018, 235, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.P.; Bhatia, A.; Buvanendran, A.; Schwenk, E.S.; Wasan, A.D.; Hurley, R.W.; Viscusi, E.R.; Narouze, S.; Davis, F.N.; Ritchie, E.C.; et al. Consensus guidelines on the use of intravenous ketamine infusions for chronic pain from the American society of regional anesthesia and pain medicine, the American academy of pain medicine, and the American society of anesthesiologists. Reg. Anesth. Pain Med. 2018, 43, 521–546. [Google Scholar] [CrossRef] [PubMed]
- Sartori, S.B.; Singewald, N. Novel pharmacological targets in drug development for the treatment of anxiety and anxiety-related disorders. Pharmacol. Ther. 2019, 204, 107402. [Google Scholar] [CrossRef] [PubMed]
- Soares, V.P.; Campos, A.C. Evidences for the anti-panic actions of cannabidiol. Curr. Neuropharmacol. 2017, 15, 291–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narayan, A.J.; Downey, L.A.; Manning, B.; Hayley, A.C. Cannabinoid treatments for anxiety: A systematic review and consideration of the impact of sleep disturbance. Neurosci. Biobehav. Rev. 2022, 143, 104941. [Google Scholar] [CrossRef] [PubMed]
- Godoy, L.D.; Rossignoli, M.T.; Delfino-Pereira, P.; Garcia-Cairasco, N.; de Lima Umeoka, E.H. A comprehensive overview on stress neurobiology: Basic concepts and clinical implications. Front. Behav. Neurosci. 2018, 12, 127. [Google Scholar] [CrossRef] [Green Version]
- Juruena, M.F.; Eror, F.; Cleare, A.J.; Young, A.H. The role of early life stress in HPA axis and anxiety. Adv. Exp. Med. Biol. 2020, 1191, 141–153. [Google Scholar] [CrossRef]
- Nater, U.M.; Rohleder, N. Salivary alpha-amylase as a non-invasive biomarker for the sympathetic nervous system: Current state of research. Psychoneuroendocrinology 2009, 34, 486–496. [Google Scholar] [CrossRef]
- Chojnowska, S.; Ptaszyńska-Sarosiek, I.; Kępka, A.; Knaś, M.; Waszkiewicz, N. Salivary biomarkers of stress, anxiety and depression. J. Clin. Med. 2021, 10, 517. [Google Scholar] [CrossRef]
- Hughes, J.W.; Watkins, L.; Blumenthal, J.A.; Kuhn, C.; Sherwood, A. Depression and anxiety symptoms are related to increased 24-hour urinary norepinephrine excretion among healthy middle-aged women. J. Psychosom. Res. 2004, 57, 353–358. [Google Scholar] [CrossRef]
- Holwerda, S.W.; Luehrs, R.E.; Gremaud, A.L.; Wooldridge, N.A.; Stroud, A.K.; Fiedorowicz, J.G.; Abboud, F.M.; Pierce, G.L. Relative burst amplitude of muscle sympathetic nerve activity is an indicator of altered sympathetic outflow in chronic anxiety. J. Neurophysiol. 2018, 120, 11–22. [Google Scholar] [CrossRef] [Green Version]
- van Veen, J.F.; van Vliet, I.M.; Derijk, R.H.; van Pelt, J.; Mertens, B.; Zitman, F.G. Elevated alpha-amylase but not cortisol in generalized social anxiety disorder. Psychoneuroendocrinology 2008, 33, 1313–1321. [Google Scholar] [CrossRef]
- Alvarenga, M.E.; Richards, J.C.; Lambert, G.; Esler, M.D. Psychophysiological mechanisms in panic disorder: A correlative analysis of noradrenaline spillover, neuronal noradrenaline reuptake, power spectral analysis of heart rate variability, and psychological variables. Psychosom. Med. 2006, 68, 8–16. [Google Scholar] [CrossRef]
- Tanaka, Y.; Ishitobi, Y.; Maruyama, Y.; Kawano, A.; Ando, T.; Imanaga, J.; Okamoto, S.; Kanehisa, M.; Higuma, H.; Ninomiya, T.; et al. Salivary α-amylase and cortisol responsiveness following electrical stimulation stress in panic disorder patients. Neurosci. Res. 2012, 73, 80–84. [Google Scholar] [CrossRef]
- Pan, X.; Kaminga, A.C.; Wen, S.W.; Liu, A. Catecholamines in post-traumatic stress disorder: A systematic review and meta-analysis. Front. Mol. Neurosci. 2018, 11, 450. [Google Scholar] [CrossRef]
- Thoma, M.V.; Joksimovic, L.; Kirschbaum, C.; Wolf, J.M.; Rohleder, N. Altered salivary alpha-amylase awakening response in bosnian war refugees with posttraumatic stress disorder. Psychoneuroendocrinology 2012, 37, 810–817. [Google Scholar] [CrossRef]
- Park, J.; Marvar, P.J.; Liao, P.; Kankam, M.L.; Norrholm, S.D.; Downey, R.M.; McCullough, S.A.; Le, N.-A.; Rothbaum, B.O. Baroreflex dysfunction and augmented sympathetic nerve responses during mental stress in veterans with post-traumatic stress disorder. J. Physiol. 2017, 595, 4893–4908. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, D.J.; Thompson, J.M.; Lambert, G.W.; Jennings, G.L.; Schwarz, R.G.; Jefferys, D.; Turner, A.G.; Esler, M.D. Sympathetic activity in patients with panic disorder at rest, under laboratory mental stress, and during panic attacks. Arch. Gen. Psychiatry 1998, 55, 511–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaidi, S.; Atrooz, F.; Valdez, D.; Liu, H.; Kochi, C.; Bond, R.A.; Salim, S. Protective effect of propranolol and nadolol on social defeat-induced behavioral impairments in rats. Neurosci. Lett. 2020, 725, 134892. [Google Scholar] [CrossRef] [PubMed]
- Ravaris, C.L.; Friedman, M.J.; Hauri, P.J.; McHugo, G.J. A controlled study of alprazolam and propranolol in panic-disordered and agoraphobic outpatients. J. Clin. Psychopharmacol. 1991, 11, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.H.; Milgrom, P.; Fiset, L. Effect of a beta-adrenergic blocking agent on dental anxiety. J. Dent. Res. 1991, 70, 1306–1308. [Google Scholar] [CrossRef]
- Falloon, I.R.; Lloyd, G.G.; Harpin, R.E. The treatment of social phobia. real-life rehearsal with nonprofessional therapists. J. Nerv. Ment. Dis. 1981, 169, 180–184. [Google Scholar] [CrossRef]
- Giustino, T.F.; Fitzgerald, P.J.; Maren, S. Revisiting propranolol and PTSD: Memory erasure or extinction enhancement? Neurobiol. Learn Mem. 2016, 130, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Tafet, G.E.; Nemeroff, C.B. Pharmacological treatment of anxiety disorders: The role of the HPA axis. Front. Psychiatry 2020, 11, 443. [Google Scholar] [CrossRef] [PubMed]
- Wedekind, D.; Bandelow, B.; Broocks, A.; Hajak, G.; Rüther, E. Salivary, total plasma and plasma free cortisol in panic disorder. J. Neural. Transm. 2000, 107, 831–837. [Google Scholar] [CrossRef] [PubMed]
- Roy-Byrne, P.P.; Uhde, T.W.; Post, R.M.; Gallucci, W.; Chrousos, G.P.; Gold, P.W. The corticotropin-releasing hormone stimulation test in patients with panic disorder. Am. J. Psychiatry 1986, 143, 896–899. [Google Scholar] [CrossRef] [PubMed]
- Chaudieu, I.; Beluche, I.; Norton, J.; Boulenger, J.-P.; Ritchie, K.; Ancelin, M.L. Abnormal reactions to environmental stress in elderly persons with anxiety disorders: Evidence from a population study of diurnal cortisol changes. J. Affect. Disord. 2008, 106, 307–313. [Google Scholar] [CrossRef] [Green Version]
- Alpers, G.W.; Abelson, J.L.; Wilhelm, F.H.; Roth, W.T. Salivary cortisol response during exposure treatment in driving phobics. Psychosom. Med. 2003, 65, 679–687. [Google Scholar] [CrossRef]
- Reeves, J.W.; Fisher, A.J.; Newman, M.G.; Granger, D.A. Sympathetic and hypothalamic-pituitary-adrenal asymmetry in generalized anxiety disorder. Psychophysiology 2016, 53, 951–957. [Google Scholar] [CrossRef]
- Nayana, J.; Rao, B.S.S.; Srikumar, B.N. Mifepristone’s effects on depression- and anxiety-like behavior in rodents. Steroids 2022, 184, 109058. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Chen, P.; Li, Y.; Cui, K.; Noel, D.M.; Cummins, E.D.; Peterson, D.J.; Brown, R.W.; Zhu, M.-Y. Corticosterone administration up-regulated expression of norepinephrine transporter and dopamine β-hydroxylase in rat locus coeruleus and its terminal regions. J. Neurochem. 2014, 128, 445–458. [Google Scholar] [CrossRef] [Green Version]
- Ding, J.; da Silva, M.S.; Lingeman, J.; Chen, X.; Shi, Y.; Han, F.; Meijer, O.C. Late glucocorticoid receptor antagonism changes the outcome of adult life stress. Psychoneuroendocrinology 2019, 107, 169–178. [Google Scholar] [CrossRef]
- Saavedra, J.M. Brain and pituitary angiotensin. Endocr. Rev. 1992, 13, 329–380. [Google Scholar] [CrossRef]
- Bali, A.; Jaggi, A.S. Angiotensin as stress mediator: Role of its receptor and interrelationships among other stress mediators and receptors. Pharmacol. Res. 2013, 76, 49–57. [Google Scholar] [CrossRef]
- Olsson, M.; Annerbrink, K.; Westberg, L.; Melke, J.; Baghaei, F.; Rosmond, R.; Holm, G.; Andersch, S.; Allgulander, C.; Eriksson, E. Angiotensin-related genes in patients with panic disorder. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2004, 127B, 81–84. [Google Scholar] [CrossRef] [PubMed]
- Sonino, N.; Fallo, F.; Fava, G.A. Psychological aspects of primary aldosteronism. Psychother. Psychosom. 2006, 75, 327–330. [Google Scholar] [CrossRef] [PubMed]
- Gao, N.; Wang, H.; Xu, X.; Yang, Z.; Zhang, T. Angiotensin II induces cognitive decline and anxiety-like behavior via disturbing pattern of theta-gamma oscillations. Brain Res. Bull 2021, 174, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Duchemin, S.; Belanger, E.; Wu, R.; Ferland, G.; Girouard, H. Chronic perfusion of angiotensin II causes cognitive dysfunctions and anxiety in mice. Physiol. Behav. 2013, 109, 63–68. [Google Scholar] [CrossRef]
- D’Souza, M.S.; Guisinger, T.C.; Norman, H.; Seeley, S.L.; Chrissobolis, S. Regulator of G-protein signaling 5 protein protects against anxiety- and depression-like behavior. Behav. Pharmacol. 2019, 30, 712–721. [Google Scholar] [CrossRef]
- Hlavacova, N.; Jezova, D. Chronic treatment with the mineralocorticoid hormone aldosterone results in increased anxiety-like behavior. Horm. Behav. 2008, 54, 90–97. [Google Scholar] [CrossRef]
- Chrissobolis, S.; Luu, A.N.; Waldschmidt, R.A.; Yoakum, M.E.; D’Souza, M.S. Targeting the renin angiotensin system for the treatment of anxiety and depression. Pharmacol. Biochem. Behav. 2020, 199, 173063. [Google Scholar] [CrossRef]
- Repova, K.; Aziriova, S.; Krajcirovicova, K.; Simko, F. Cardiovascular therapeutics: A new potential for anxiety treatment? Med. Res. Rev. 2022, 42, 1202–1245. [Google Scholar] [CrossRef]
- Khoury, N.M.; Marvar, P.J.; Gillespie, C.F.; Wingo, A.; Schwartz, A.; Bradley, B.; Kramer, M.; Ressler, K.J. The renin-angiotensin pathway in posttraumatic stress disorder: Angiotensin-converting enzyme inhibitors and angiotensin receptor blockers are associated with fewer traumatic stress symptoms. J. Clin. Psychiatry 2012, 73, 849–855. [Google Scholar] [CrossRef] [Green Version]
- Reinecke, A.; Browning, M.; Klein Breteler, J.; Kappelmann, N.; Ressler, K.J.; Harmer, C.J.; Craske, M.G. Angiotensin regulation of amygdala response to threat in high-trait-anxiety individuals. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2018, 3, 826–835. [Google Scholar] [CrossRef]
- Shad, M.U. Is there an association between anxiety symptoms and valsartan treatment? J. Affect. Disord. 2020, 261, 111–112. [Google Scholar] [CrossRef]
- de Kloet, A.D.; Wang, L.; Pitra, S.; Hiller, H.; Smith, J.A.; Tan, Y.; Nguyen, D.; Cahill, K.M.; Sumners, C.; Stern, J.E.; et al. A unique “angiotensin-sensitive” neuronal population coordinates neuroendocrine, cardiovascular, and behavioral responses to stress. J. Neurosci. 2017, 37, 3478–3490. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Hiller, H.; Smith, J.A.; de Kloet, A.D.; Krause, E.G. Angiotensin type 1a receptors in the paraventricular nucleus of the hypothalamus control cardiovascular reactivity and anxiety-like behavior in male mice. Physiol. Genom. 2016, 48, 667–676. [Google Scholar] [CrossRef] [Green Version]
- Aziriova, S.; Bednarova, K.R.; Krajcirovicova, K.; Hrenak, J.; Rajkovicova, R.; Arendasova, K.; Kamodyova, N.; Celec, P.; Zorad, S.; Adamcova, M.; et al. Doxorubicin-induced behavioral disturbances in rats: Protective effect of melatonin and captopril. Pharmacol. Biochem. Behav. 2014, 124, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Ranjbar, H.; Aghaei, I.; Moosazadeh, M.; Shabani, M. Angiotensin II type 1 receptor blocker losartan attenuates locomotor, anxiety-like behavior, and passive avoidance learning deficits in a sub-chronic stress model. Iran. J. Basic Med. Sci. 2018, 21, 856. [Google Scholar] [CrossRef] [PubMed]
- Salmani, H.; Hosseini, M.; Baghcheghi, Y.; Moradi-Marjaneh, R.; Mokhtari-Zaer, A. Losartan modulates brain inflammation and improves mood disorders and memory impairment induced by innate immune activation: The role of PPAR-γ activation. Cytokine 2020, 125, 154860. [Google Scholar] [CrossRef] [PubMed]
- López-Rubalcava, C.; Paez-Martinez, N.; Oikawa, J. Blockade of corticosteroid receptors induces anxiolytic-like effects in streptozotocin-induced diabetic mice, and synergizes with diazepam. Behav. Pharmacol. 2013, 24, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Hlavacova, N.; Jezova, D. Effect of single treatment with the antihypertensive drug eplerenone on hormone levels and anxiety-like behaviour in rats. Endocr. Regul. 2008, 42, 147–153. [Google Scholar]
- Genaro, K.; Fabris, D.; Fachim, H.A.; Prado, W.A. Angiotensin AT1 receptors modulate the anxiogenic effects of angiotensin (5–8) injected into the rat ventrolateral periaqueductal gray. Peptides 2017, 96, 8–14. [Google Scholar] [CrossRef]
- Kumar, K.V.A.; Nagwar, S.; Thyloor, R.; Satyanarayana, S. Anti-stress and nootropic activity of drugs affecting the renin-angiotensin system in rats based on indirect biochemical evidence. J. Renin Angiotensin Aldosterone Syst. 2015, 16, 801–812. [Google Scholar] [CrossRef] [Green Version]
- Pavel, J.; Benicky, J.; Murakami, Y.; Sanchez-Lemus, E.; Saavedra, J.M. Peripherally administered angiotensin II AT1 receptor antagonists are anti-stress compounds in vivo. Ann. N. Y. Acad. Sci. 2008, 1148, 360–366. [Google Scholar] [CrossRef] [Green Version]
- Bouayed, J.; Rammal, H.; Soulimani, R. Oxidative stress and anxiety: Relationship and cellular pathways. Oxid. Med. Cell Longev. 2009, 2, 63–67. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Chanana, P. Role of nitric oxide in stress-induced anxiety: From pathophysiology to therapeutic target. Vitam. Horm. 2017, 103, 147–167. [Google Scholar] [CrossRef] [PubMed]
- Hovatta, I.; Tennant, R.S.; Helton, R.; Marr, R.A.; Singer, O.; Redwine, J.M.; Ellison, J.A.; Schadt, E.E.; Verma, I.M.; Lockhart, D.J.; et al. Glyoxalase 1 and glutathione reductase 1 regulate anxiety in mice. Nature 2005, 438, 662–666. [Google Scholar] [CrossRef] [PubMed]
- Rammal, H.; Bouayed, J.; Younos, C.; Soulimani, R. Evidence that oxidative stress is linked to anxiety-related behaviour in mice. Brain Behav. Immun. 2008, 22, 1156–1159. [Google Scholar] [CrossRef] [PubMed]
- Rammal, H.; Bouayed, J.; Younos, C.; Soulimani, R. The impact of high anxiety level on the oxidative status of mouse peripheral blood lymphocytes, granulocytes and monocytes. Eur. J. Pharmacol. 2008, 589, 173–175. [Google Scholar] [CrossRef]
- Rice, M.E. Ascorbate regulation and its neuroprotective role in the brain. Trends Neurosci. 2000, 23, 209–216. [Google Scholar] [CrossRef]
- Moritz, B.; Schmitz, A.E.; Rodrigues, A.L.S.; Dafre, A.L.; Cunha, M.P. The role of vitamin C in stress-related disorders. J. Nutr. Biochem. 2020, 85, 108459. [Google Scholar] [CrossRef]
- de Almeida, A.A.C.; de Carvalho, R.B.F.; Silva, O.A.; de Sousa, D.P.; de Freitas, R.M. Potential antioxidant and anxiolytic effects of (+)-limonene epoxide in mice after marble-burying test. Pharmacol. Biochem. Behav. 2014, 118, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Walia, V.; Garg, C.; Garg, M. Nitrergic signaling modulation by ascorbic acid treatment is responsible for anxiolysis in mouse model of anxiety. Behav. Brain Res. 2019, 364, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Olajide, O.J.; Yawson, E.O.; Gbadamosi, I.T.; Arogundade, T.T.; Lambe, E.; Obasi, K.; Lawal, I.T.; Ibrahim, A.; Ogunrinola, K.Y. Ascorbic acid ameliorates behavioural deficits and neuropathological alterations in rat model of alzheimer’s disease. Environ. Toxicol. Pharmacol. 2017, 50, 200–211. [Google Scholar] [CrossRef] [PubMed]
- Koizumi, M.; Kondo, Y.; Isaka, A.; Ishigami, A.; Suzuki, E. Vitamin C impacts anxiety-like behavior and stress-induced anorexia relative to social environment in SMP30/GNL knockout mice. Nutr. Res. 2016, 36, 1379–1391. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, I.J.L.; de Souza, V.V.; Motta, V.; Da-Silva, S.L. Effects of oral vitamin C supplementation on anxiety in students: A double-blind, randomized, placebo-controlled trial. Pak J. Biol. Sci. 2015, 18, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Moritz, B.; Schwarzbold, M.L.; Guarnieri, R.; Diaz, A.P.; Rodrigues, A.L.S.; Dafre, A.L. Effects of ascorbic acid on anxiety state and affect in a non-clinical sample. Acta Neurobiol. Exp. 2017, 77, 362–372. [Google Scholar] [CrossRef] [Green Version]
- Masood, A.; Nadeem, A.; Mustafa, S.J.; O’Donnell, J.M. Reversal of oxidative stress-induced anxiety by inhibition of phosphodiesterase-2 in mice. J. Pharmacol. Exp. Ther. 2008, 326, 369–379. [Google Scholar] [CrossRef] [Green Version]
- Salim, S.; Sarraj, N.; Taneja, M.; Saha, K.; Tejada-Simon, M.V.; Chugh, G. Moderate treadmill exercise prevents oxidative stress-induced anxiety-like behavior in rats. Behav. Brain Res. 2010, 208, 545–552. [Google Scholar] [CrossRef]
- Workman, J.L.; Trainor, B.C.; Finy, M.S.; Nelson, R.J. Inhibition of neuronal nitric oxide reduces anxiety-like responses to pair housing. Behav. Brain Res. 2008, 187, 109–115. [Google Scholar] [CrossRef] [Green Version]
- Filipović, D.; Todorović, N.; Bernardi, R.E.; Gass, P. Oxidative and nitrosative stress pathways in the brain of socially isolated adult male rats demonstrating depressive- and anxiety-like symptoms. Brain Struct. Funct. 2017, 222, 1–20. [Google Scholar] [CrossRef]
- Yasunari, K.; Matsui, T.; Maeda, K.; Nakamura, M.; Watanabe, T.; Kiriike, N. Anxiety-induced plasma norepinephrine augmentation increases reactive oxygen species formation by monocytes in essential hypertension. Am. J. Hypertens. 2006, 19, 573–578. [Google Scholar] [CrossRef]
- Kuloglu, M.; Atmaca, M.; Tezcan, E.; Gecici, O.; Tunckol, H.; Ustundag, B. Antioxidant enzyme activities and malondialdehyde levels in patients with obsessive-compulsive disorder. Neuropsychobiology 2002, 46, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Kuloglu, M.; Atmaca, M.; Tezcan, E.; Ustundag, B.; Bulut, S. Antioxidant enzyme and malondialdehyde levels in patients with panic disorder. Neuropsychobiology 2002, 46, 186–189. [Google Scholar] [CrossRef]
- Marcinkiewcz, C.A.; Mazzone, C.M.; D’Agostino, G.; Halladay, L.R.; Hardaway, J.A.; DiBerto, J.F.; Navarro, M.; Burnham, N.; Cristiano, C.; Dorrier, C.E.; et al. Serotonin engages an anxiety and fear-promoting circuit in the extended amygdala. Nature 2016, 537, 97–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comai, S.; De Gregorio, D.; Posa, L.; Ochoa-Sanchez, R.; Bedini, A.; Gobbi, G. Dysfunction of serotonergic activity and emotional responses across the light-dark cycle in mice lacking melatonin MT2 receptors. J. Pineal. Res. 2020, 69, e12653. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Friedmann, D.; Xiong, J.; Liu, C.D.; Ferguson, B.R.; Weerakkody, T.; DeLoach, K.E.; Ran, C.; Pun, A.; Sun, Y.; et al. Anatomically defined and functionally distinct dorsal raphe serotonin sub-systems. Cell 2018, 175, 472.e20–487.e20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, B.M.; Merchant, K.M. Serotonin 2C receptors within the basolateral amygdala induce acute fear-like responses in an open-field environment. Brain Res. 2003, 993, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bauer, E.P. Serotonin in fear conditioning processes. Behav. Brain Res. 2015, 277, 68–77. [Google Scholar] [CrossRef]
- Kalueff, A.V.; Nutt, D.J. Role of GABA in anxiety and depression. Depress. Anxiety 2007, 24, 495–517. [Google Scholar] [CrossRef]
- Hernández-Vázquez, F.; Garduño, J.; Hernández-López, S. GABAergic modulation of serotonergic neurons in the dorsal raphe nucleus. Rev. Neurosci. 2019, 30, 289–303. [Google Scholar] [CrossRef]
- Möhler, H. The GABA system in anxiety and depression and its therapeutic potential. Neuropharmacology 2012, 62, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Manzanares, P.A.R.; Isoardi, N.A.; Carrer, H.F.; Molina, V.A. Previous stress facilitates fear memory, attenuates GABAergic inhibition, and increases synaptic plasticity in the rat basolateral amygdala. J. Neurosci. 2005, 25, 8725–8734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, T.A.; Baracz, S.J.; Everett, N.A.; Robinson, K.J.; Cornish, J.L. Differential effects of GABAA receptor activation in the prelimbic and orbitofrontal cortices on anxiety. Psychopharmacology 2020, 237, 3237–3247. [Google Scholar] [CrossRef] [PubMed]
- Zarrindast, M.-R.; Khakpai, F. The modulatory role of dopamine in anxiety-like behavior. Arch. Iran. Med. 2015, 18, 591–603. [Google Scholar] [PubMed]
- Yokoyama, M.; Suzuki, E.; Sato, T.; Maruta, S.; Watanabe, S.; Miyaoka, H. Amygdalic levels of dopamine and serotonin rise upon exposure to conditioned fear stress without elevation of glutamate. Neurosci. Lett. 2005, 379, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Quirk, G.J.; Likhtik, E.; Pelletier, J.G.; Paré, D. Stimulation of medial prefrontal cortex decreases the responsiveness of central amygdala output neurons. J. Neurosci. 2003, 23, 8800–8807. [Google Scholar] [CrossRef] [Green Version]
- Marowsky, A.; Yanagawa, Y.; Obata, K.; Vogt, K.E. A specialized subclass of interneurons mediates dopaminergic facilitation of amygdala function. Neuron 2005, 48, 1025–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez de la Mora, M.; Gallegos-Cari, A.; Crespo-Ramirez, M.; Marcellino, D.; Hansson, A.C.; Fuxe, K. Distribution of dopamine D(2)-like receptors in the rat amygdala and their role in the modulation of unconditioned fear and anxiety. Neuroscience 2012, 201, 252–266. [Google Scholar] [CrossRef]
- Simko, F.; Reiter, R.J.; Pechanova, O.; Paulis, L. Experimental models of melatonin-deficient hypertension. Front. Biosci. 2013, 18, 616–625. [Google Scholar] [CrossRef] [Green Version]
- Holmes, S.W.; Sugden, D. Proceedings: The effect of melatonin on pinealectomy-induced hypertension in the rat. Br. J. Pharmacol. 1976, 56, 360P–361P. [Google Scholar]
- Reiter, R.J.; Tan, D.-X.; Rosales-Corral, S.; Manchester, L.C. The universal nature, unequal distribution and antioxidant functions of melatonin and its derivatives. Mini Rev. Med. Chem. 2013, 13, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Bubenik, G.A. Gastrointestinal melatonin: Localization, function, and clinical relevance. Dig. Dis. Sci. 2002, 47, 2336–2348. [Google Scholar] [CrossRef]
- Carrillo-Vico, A.; Lardone, P.J.; Alvarez-Sánchez, N.; Rodríguez-Rodríguez, A.; Guerrero, J.M. Melatonin: Buffering the immune system. Int. J. Mol. Sci. 2013, 14, 8638. [Google Scholar] [CrossRef] [Green Version]
- Hardeland, R. Chronobiology of melatonin beyond the feedback to the suprachiasmatic nucleus-consequences to melatonin dysfunction. Int. J. Mol. Sci. 2013, 14, 5817. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.; Wang, H.; Chen, S.; Yu, D.; Reiter, R.J. Phytomelatonin: An emerging regulator of plant biotic stress resistance. Trends Plant Sci. 2021, 26, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Manchester, L.C.; Terron, M.P.; Flores, L.J.; Koppisepi, S. Medical implications of melatonin: Receptor-mediated and receptor-independent actions. Adv. Med. Sci. 2007, 52, 11–28. [Google Scholar] [PubMed]
- Reiter, R.J.; Tan, D.-X.; Fuentes-Broto, L. Melatonin: A multitasking molecule. Prog. Brain Res. 2010, 181, 127–151. [Google Scholar] [CrossRef]
- Rezzani, R.; Franco, C.; Hardeland, R.; Rodella, L.F. Thymus-pineal gland axis: Revisiting its role in human life and ageing. Int. J. Mol. Sci. 2020, 21, 8806. [Google Scholar] [CrossRef] [PubMed]
- Tengattini, S.; Reiter, R.J.; Tan, D.-X.; Terron, M.P.; Rodella, L.F.; Rezzani, R. Cardiovascular diseases: Protective effects of melatonin. J. Pineal. Res. 2008, 44, 16–25. [Google Scholar] [CrossRef]
- Simko, F.; Paulis, L. Melatonin as a potential antihypertensive treatment. J. Pineal. Res. 2007, 42, 319–322. [Google Scholar] [CrossRef]
- Korkmaz, A.; Topal, T.; Tan, D.-X.; Reiter, R.J. Role of melatonin in metabolic regulation. Rev. Endocr. Metab. Disord. 2009, 10, 261–270. [Google Scholar] [CrossRef]
- Stankov, B.; Reiter, R.J. Melatonin receptors: Current status, facts, and hypotheses. Life Sci. 1990, 46, 971–982. [Google Scholar] [CrossRef] [PubMed]
- Paulis, L.; Simko, F.; Laudon, M. Cardiovascular effects of melatonin receptor agonists. Expert. Opin. Investig. Drugs. 2012, 21, 1661–1678. [Google Scholar] [CrossRef] [PubMed]
- Nosjean, O.; Ferro, M.; Coge, F.; Beauverger, P.; Henlin, J.M.; Lefoulon, F.; Fauchere, J.L.; Delagrange, P.; Canet, E.; Boutin, J.A. Identification of the melatonin-binding site MT3 as the quinone reductase 2. J. Biol. Chem. 2000, 275, 31311–31317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slominski, R.M.; Reiter, R.J.; Schlabritz-Loutsevitch, N.; Ostrom, R.S.; Slominski, A.T. Melatonin membrane receptors in peripheral tissues: Distribution and functions. Mol. Cell Endocrinol. 2012, 351, 152–166. [Google Scholar] [CrossRef] [Green Version]
- Benítez-King, G. Melatonin as a cytoskeletal modulator: Implications for cell physiology and disease. J. Pineal. Res. 2006, 40, 1–9. [Google Scholar] [CrossRef]
- Tan, D.; Reiter, R.J.; Manchester, L.C.; Yan, M.; El-Sawi, M.; Sainz, R.M.; Mayo, J.C.; Kohen, R.; Allegra, M.; Hardeland, R. Chemical and physical properties and potential mechanisms: Melatonin as a broad spectrum antioxidant and free radical scavenger. Curr. Top Med. Chem. 2002, 2, 181–197. [Google Scholar] [CrossRef] [Green Version]
- Reiter, R.J.; Manchester, L.C.; Fuentes-Broto, L.; Tan, D.-X. Cardiac hypertrophy and remodelling: Pathophysiological consequences and protective effects of melatonin. J. Hypertens. 2010, 28 (Suppl. S1), S7–S12. [Google Scholar] [CrossRef]
- Rodriguez, C.; Mayo, J.C.; Sainz, R.M.; Antolín, I.; Herrera, F.; Martín, V.; Reiter, R.J. Regulation of antioxidant enzymes: A significant role for melatonin. J. Pineal. Res. 2004, 36, 1–9. [Google Scholar] [CrossRef]
- Limón-Pacheco, J.H.; Gonsebatt, M.E. The glutathione system and its regulation by neurohormone melatonin in the central nervous system. Cent. Nerv. Syst. Agents Med. Chem. 2010, 10, 287–297. [Google Scholar] [CrossRef]
- Hardeland, R.; Cardinali, D.P.; Srinivasan, V.; Spence, D.W.; Brown, G.M.; Pandi-Perumal, S.R. Melatonin—A pleiotropic, orchestrating regulator molecule. Prog. Neurobiol. 2011, 93, 350–384. [Google Scholar] [CrossRef] [Green Version]
- Gitto, E.; Tan, D.X.; Reiter, R.J.; Karbownik, M.; Manchester, L.C.; Cuzzocrea, S.; Fulia, F.; Barberi, I. Individual and synergistic antioxidative actions of melatonin: Studies with vitamin E, vitamin C, glutathione and desferrioxamine (desferoxamine) in rat liver homogenates. J. Pharm. Pharmacol. 2001, 53, 1393–1401. [Google Scholar] [CrossRef]
- López-Burillo, S.; Tan, D.-X.; Mayo, J.C.; Sainz, R.M.; Manchester, L.C.; Reiter, R.J. Melatonin, xanthurenic acid, resveratrol, EGCG, vitamin C and alpha-lipoic acid differentially reduce oxidative DNA damage induced by fenton reagents: A study of their individual and synergistic actions. J. Pineal. Res. 2003, 34, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Galano, A.; Tan, D.X.; Reiter, R.J. On the free radical scavenging activities of melatonin’s metabolites, AFMK and AMK. J. Pineal. Res. 2013, 54, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Galano, A.; Medina, M.E.; Tan, D.X.; Reiter, R.J. Melatonin and its metabolites as copper chelating agents and their role in inhibiting oxidative stress: A physicochemical analysis. J. Pineal. Res. 2015, 58, 107–116. [Google Scholar] [CrossRef]
- Maldonado, M.D.; Mora-Santos, M.; Naji, L.; Carrascosa-Salmoral, M.P.; Naranjo, M.C.; Calvo, J.R. Evidence of melatonin synthesis and release by mast cells. Possible modulatory role on inflammation. Pharmacol. Res. 2010, 62, 282–287. [Google Scholar] [CrossRef]
- Hardeland, R. Melatonin and inflammation-story of a double-edged blade. J. Pineal. Res. 2018, 65, e12525. [Google Scholar] [CrossRef] [Green Version]
- Simko, F.; Reiter, R.J. Is melatonin deficiency a unifying pathomechanism of high risk patients with COVID-19? Life Sci. 2020, 256, 117902. [Google Scholar] [CrossRef] [PubMed]
- Simko, F.; Hrenak, J.; Dominguez-Rodriguez, A.; Reiter, R.J. Melatonin as a putative protection against myocardial injury in COVID-19 infection. Expert. Rev. Clin. Pharm. 2020, 13, 921–924. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, X.; Ni, L.; Di, X.; Ma, B.; Niu, S.; Liu, C.; Reiter, R.J. COVID-19: Melatonin as a potential adjuvant treatment. Life Sci. 2020, 250, 117583. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Sharma, R.; Simko, F.; Dominguez-Rodriguez, A.; Tesarik, J.; Neel, R.L.; Slominski, A.T.; Kleszczynski, K.; Martin-Gimenez, V.M.; Manucha, W.; et al. Melatonin: Highlighting its use as a potential treatment for SARS-CoV-2 infection. Cell Mol. Life Sci. 2022, 79, 143. [Google Scholar] [CrossRef]
- Uguz, A.C.; Cig, B.; Espino, J.; Bejarano, I.; Naziroglu, M.; Rodríguez, A.B.; Pariente, J.A. Melatonin potentiates chemotherapy-induced cytotoxicity and apoptosis in rat pancreatic tumor cells. J. Pineal. Res. 2012, 53, 91–98. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.X.; Rosales-Corral, S.; Galano, A.; Zhou, X.J.; Xu, B. Mitochondria: Central organelles for melatonin’s antioxidant and anti-aging actions. Molecules 2018, 23, 509. [Google Scholar] [CrossRef] [Green Version]
- Reiter, R.J.; Ma, Q.; Sharma, R. Melatonin in mitochondria: Mitigating clear and present dangers. Physiology 2020, 35, 86–95. [Google Scholar] [CrossRef]
- Reiter, R.J.; Sharma, R.; Rosales-Corral, S.; de Campos Zuccari, D.A.P.; de Almeida Chuffa, L.G. Melatonin: A mitochondrial resident with a diverse skill set. Life Sci. 2022, 301, 120612. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Rosales-Corral, S.; Sharma, R.F. Perturbations and their contributions to chaotic physiology. Adv. Med. Sci. 2020, 65, 394–402. [Google Scholar] [CrossRef]
- Vanĕcek, J.; Janský, L. Short days induce changes in specific melatonin binding in hamster median eminence and anterior pituitary. Brain Res. 1989, 477, 387–390. [Google Scholar] [CrossRef] [PubMed]
- Malpaux, B.; Migaud, M.; Tricoire, H.; Chemineau, P. Biology of mammalian photoperiodism and the critical role of the pineal gland and melatonin. J. Biol. Rhythm. 2001, 16, 336–347. [Google Scholar] [CrossRef] [PubMed]
- Duncan, M.J.; Takahashi, J.S.; Dubocovich, M.L. Characteristics and autoradiographic localization of 2-[125I]iodomelatonin binding sites in djungarian hamster brain. Endocrinology 1989, 125, 1011–1018. [Google Scholar] [CrossRef]
- Molcan, L.; Sutovska, H.; Okuliarova, M.; Senko, T.; Krskova, L.; Zeman, M. Dim light at night attenuates circadian rhythms in the cardiovascular system and suppresses melatonin in rats. Life Sci. 2019, 231, 116568. [Google Scholar] [CrossRef] [PubMed]
- Golus, P.; King, M.G. The effects of melatonin on open field behavior. Pharmacol. Biochem. Behav. 1981, 15, 883–885. [Google Scholar] [CrossRef] [PubMed]
- Golombek, D.A.; Martini, M.; Cardinali, D.P. Melatonin as an anxiolytic in rats: Time dependence and interaction with the central GABAergic system. Eur. J. Pharmacol. 1993, 237, 231–236. [Google Scholar] [CrossRef]
- Ochoa-Sanchez, R.; Rainer, Q.; Comai, S.; Spadoni, G.; Bedini, A.; Rivara, S.; Fraschini, F.; Mor, M.; Tarzia, G.; Gobbi, G. Anxiolytic effects of the melatonin MT2 receptor partial agonist UCM765: Comparison with melatonin and diazepam. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2012, 39, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Bustamante-García, R.; Lira-Rocha, A.S.; Espejo-González, O.; Gómez-Martínez, A.E.; Picazo, O. Anxiolytic-like effects of a new 1-N substituted analog of melatonin in pinealectomized rats. Prog. Neuropsychopharmacol. Biol. Psychiatry 2014, 51, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.A.E.; Ahmed, H.I.; El-Morsy, E.M. Melatonin protects against diazinon-induced neurobehavioral changes in rats. Neurochem. Res. 2013, 38, 2227–2236. [Google Scholar] [CrossRef]
- Krsková, L.; Vrabcová, M.; Zeman, M. Effect of melatonin on exploration and anxiety in normotensive and hypertensive rats with high activity of renin-angiotensin system. Neuro Endocrinol. Lett. 2007, 28, 295–301. [Google Scholar]
- Xie, L.-L.; Li, S.-S.; Fan, Y.-J.; Qi, M.-M.; Li, Z.-Z. Melatonin alleviates traumatic brain injury-induced anxiety-like behaviors in rats: Roles of the protein kinase A/CAMP-response element binding signaling pathway. Exp. Ther. Med. 2022, 23, 248. [Google Scholar] [CrossRef]
- Pierrefiche, G.; Zerbib, R.; Laborit, H. Anxiolytic activity of melatonin in mice: Involvement of benzodiazepine receptors. Res. Commun. Chem. Pathol. Pharmacol. 1993, 82, 131–142. [Google Scholar]
- Kopp, C.; Vogel, E.; Rettori, M.; Delagrange, P.; Misslin, R. Anxiolytic-like properties of melatonin receptor agonists in mice: Involvement of Mt1 and/or MT2 receptors in the regulation of emotional responsiveness. Neuropharmacology 2000, 39, 1865–1871. [Google Scholar] [CrossRef]
- Tian, S.; Laudon, M.; Han, L.; Gao, J.; Huang, F.; Yang, Y.; Deng, H. Antidepressant- and anxiolytic effects of the novel melatonin agonist neu-P11 in rodent models. Acta Pharmacol. Sin. 2010, 31, 775–783. [Google Scholar] [CrossRef]
- Caumo, W.; Levandovski, R.; Hidalgo, M.P.L. Preoperative anxiolytic effect of melatonin and clonidine on postoperative pain and morphine consumption in patients undergoing abdominal hysterectomy: A double-blind, randomized, placebo-controlled study. J. Pain. 2009, 10, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Abbasivash, R.; Salimi, S.; Ahsan, B.; Moallemi, N.; Sane, S. The effect of melatonin on anxiety and pain of tourniquet in intravenous regional anesthesia. Adv. Biomed. Res. 2019, 8, 67. [Google Scholar] [CrossRef] [PubMed]
- Patel, T.; Kurdi, M.S. A comparative study between oral melatonin and oral midazolam on preoperative anxiety, cognitive, and psychomotor functions. J. Anaesthesiol. Clin. Pharmacol. 2015, 31, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.V.; Halladin, N.L.; Rosenberg, J.; Gögenur, I.; Møller, A.M. Melatonin for pre- and postoperative anxiety in adults. Cochrane Database Syst. Rev. 2015, 2015, CD009861. [Google Scholar] [CrossRef]
- Madsen, B.K.; Zetner, D.; Møller, A.M.; Rosenberg, J. Melatonin for preoperative and postoperative anxiety in adults. Cochrane Database Syst. Rev. 2020, 12, CD009861. [Google Scholar] [CrossRef]
- Samarkandi, A.; Naguib, M.; Riad, W.; Thalaj, A.; Alotibi, W.; Aldammas, F.; Albassam, A. Melatonin vs. Midazolam premedication in children: A double-blind, placebo-controlled study. Eur. J. Anaesthesiol. 2005, 22, 189–196. [Google Scholar] [CrossRef]
- Impellizzeri, P.; Vinci, E.; Gugliandolo, M.C.; Cuzzocrea, F.; Larcan, R.; Russo, T.; Gravina, M.R.; Arena, S.; D’Angelo, G.; Gitto, E.; et al. Premedication with Melatonin vs. Midazolam: Efficacy on anxiety and compliance in paediatric surgical patients. Eur. J. Pediatr. 2017, 176, 947–953. [Google Scholar] [CrossRef]
- Jouybar, R.; Kazemifar, S.; Asmarian, N.; Karami, A.; Khademi, S. Comparison of the effect of melatonin, dexmedetomidine, and gabapentin on reduction of postoperative pain and anxiety following laminectomy: A randomized clinical trial. BMC Anesthesiol. 2022, 22, 318. [Google Scholar] [CrossRef]
- SaberMoghaddam, M.; Sheybani, S.; Bakhtiari, E.; Shakiba, M. The effect of preoperative sublingual melatonin on postoperative pain severity in patients undergoing colorectal surgery: A triple-blinded randomized trial. Med. J. Islam. Repub. Iran 2022, 36, 90. [Google Scholar] [CrossRef]
- Acikgoz, B.; Dalkiran, B.; Dayi, A. An overview of the currency and usefulness of behavioral tests used from past to present to assess anxiety, social behavior and depression in rats and mice. Behav. Processes 2022, 200, 104670. [Google Scholar] [CrossRef]
- Mohammadi, S.; Zahmatkesh, M. A surgical modification in the technique of rat pinealectomy. Anat. Sci. Int. 2022, 1–12. [Google Scholar] [CrossRef]
- Ali, S.T.; Asthana, V.; Gupta, D.; Singh, S.K. A comparative evaluation of oral clonidine, dexmedetomidine, and melatonin as premedicants in pediatric patients undergoing subumbilical surgeries. Rom. J. Anaesth. Intensive Care 2020, 27, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Singla, L.; Mathew, P.J.; Jain, A.; Yaddanapudi, S.; Peters, N.J. Oral melatonin as part of multimodal anxiolysis decreases emergence delirium in children whereas midazolam does not: A randomised, double-blind, placebo-controlled study. Eur. J. Anaesthesiol. 2021, 38, 1130–1137. [Google Scholar] [CrossRef]
- Besag, F.M.C.; Vasey, M.J.; Lao, K.S.J.; Wong, I.C.K. Adverse events associated with melatonin for the treatment of primary or secondary sleep disorders: A systematic review. CNS Drugs 2019, 33, 1167–1186. [Google Scholar] [CrossRef]
- Wang, H.R.; Woo, Y.S.; Bahk, W.-M. The role of melatonin and melatonin agonists in counteracting antipsychotic-induced metabolic side effects: A systematic review. Int. Clin. Psychopharmacol. 2016, 31, 301–306. [Google Scholar] [CrossRef] [Green Version]
- Huang, F.; Yang, Z.; Li, C.-Q. The melatonergic system in anxiety disorders and the role of melatonin in conditional fear. Vitam. Horm. 2017, 103, 281–294. [Google Scholar] [CrossRef]
- Comai, S.; Ochoa-Sanchez, R.; Dominguez-Lopez, S.; Bambico, F.R.; Gobbi, G. Melancholic-like behaviors and circadian neurobiological abnormalities in melatonin MT1 receptor knockout mice. Int. J. Neuropsychopharmacol. 2015, 18, pyu075. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Clough, S.J.; Dubocovich, M.L. Role of the MT1 and MT2 melatonin receptors in mediating depressive- and anxiety-like behaviors in C3H/HeN mice. Genes Brain Behav. 2017, 16, 546–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomson, D.M.; Mitchell, E.J.; Openshaw, R.L.; Pratt, J.A.; Morris, B.J. Mice lacking melatonin MT2 receptors exhibit attentional deficits, anxiety and enhanced social interaction. J. Psychopharmacol. 2021, 35, 1265–1276. [Google Scholar] [CrossRef] [PubMed]
- Becker, C.; Andre, J.; Zeau, B.; Rettori, M.-C.; Guardiola-Lemaitre, B.; Hamon, M.; Benoliel, J.-J. Melatonin MT(1/2) receptor stimulation reduces cortical overflow of cholecystokinin-like material in a model of anticipation of social defeat in the rat. Neuropharmacology 2004, 46, 1158–1167. [Google Scholar] [CrossRef] [PubMed]
- Paulis, L.; Simko, F. Blood pressure modulation and cardiovascular protection by melatonin: Potential mechanisms behind. Physiol. Res. 2007, 56, 671–684. [Google Scholar] [CrossRef] [PubMed]
- Simko, F.; Reiter, R.J.; Paulis, L. Melatonin as a rational alternative in the conservative treatment of resistant hypertension. Hypertens. Res. 2019, 42, 1828–1831. [Google Scholar] [CrossRef] [Green Version]
- Kalsbeek, A.; Garidou, M.L.; Palm, I.F.; Van Der Vliet, J.; Simonneaux, V.; Pévet, P.; Buijs, R.M. Melatonin sees the light: Blocking GABA-ergic transmission in the paraventricular nucleus induces daytime secretion of melatonin. Eur. J. Neurosci. 2000, 12, 3146–3154. [Google Scholar] [CrossRef]
- Wang, F.; Li, J.; Wu, C.; Yang, J.; Xu, F.; Zhao, Q. The GABA(A) receptor mediates the hypnotic activity of melatonin in rats. Pharmacol. Biochem. Behav. 2003, 74, 573–578. [Google Scholar] [CrossRef]
- Patel, K.P.; Li, Y.F.; Hirooka, Y. Role of nitric oxide in central sympathetic outflow. Exp. Biol. Med. 2001, 226, 814–824. [Google Scholar] [CrossRef]
- Rossi, N.F.; Black, S.M.; Telemaque-Potts, S.; Chen, H. Neuronal nitric oxide synthase activity in the paraventricular nucleus buffers central endothelin-1- induced pressor response and vasopressin secretion. J. Cardiovasc. Pharmacol. 2004, 44 (Suppl. S1), S283–S288. [Google Scholar] [CrossRef]
- Benova, M.; Herichova, I.; Stebelova, K.; Paulis, L.; Krajcirovicova, K.; Simko, F.; Zeman, M. Effect of L-NAME-induced hypertension on melatonin receptors and melatonin levels in the pineal gland and the peripheral organs of rats. Hypertens. Res. 2009, 32, 242–247. [Google Scholar] [CrossRef] [Green Version]
- Pechanova, O.; Paulis, L.; Simko, F. Peripheral and central effects of melatonin on blood pressure regulation. Int. J. Mol. Sci. 2014, 15, 17920. [Google Scholar] [CrossRef] [Green Version]
- Laflamme, A.-K.; Wu, L.; Foucart, S.; de Champlain, J. Impaired basal sympathetic tone and alpha1-adrenergic responsiveness in association with the hypotensive effect of melatonin in spontaneously hypertensive rats. Am. J. Hypertens. 1998, 11, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Girouard, H.; Chulak, C.; LeJossec, M.; Lamontagne, D.; de Champlain, J. Chronic antioxidant treatment improves sympathetic functions and beta-adrenergic pathway in the spontaneously hypertensive rats. J. Hypertens. 2003, 21, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Girouard, H.; Denault, C.; Chulak, C.; de Champlain, J. Treatment by N-acetylcysteine and melatonin increases cardiac baroreflex and improves antioxidant reserve. Am. J. Hypertens. 2004, 17, 947–954. [Google Scholar] [CrossRef] [Green Version]
- Nishi, E.E.; Almeida, V.R.; Amaral, F.G.; Simon, K.A.; Futuro-Neto, H.A.; Pontes, R.B.; Cespedes, J.G.; Campos, R.R.; Bergamaschi, C.T. Melatonin attenuates renal sympathetic overactivity and reactive oxygen species in the brain in neurogenic hypertension. Hypertens. Res. 2019, 42, 1683–1691. [Google Scholar] [CrossRef]
- Jiao, L.; Wang, Y.; Zhang, S.; Wang, Y.; Liu, Z.; Liu, Z.; Zhou, Y.; Zhou, H.; Xu, X.; Li, Z.; et al. Melatonin improves cardiac remodeling and brain-heart sympathetic hyperactivation aggravated by light disruption after myocardial infarction. J. Pineal. Res. 2022, 73, e12829. [Google Scholar] [CrossRef]
- Campos, L.A.; Bueno, C.; Barcelos, I.P.; Halpern, B.; Brito, L.C.; Amaral, F.G.; Baltatu, O.C.; Cipolla-Neto, J. Melatonin therapy improves cardiac autonomic modulation in pinealectomized patients. Front. Endocrinol. 2020, 11, 239. [Google Scholar] [CrossRef]
- Simko, F.; Baka, T.; Paulis, L.; Reiter, R.J. Elevated heart rate and nondipping heart rate as potential targets for melatonin: A review. J. Pineal. Res. 2016, 61, 127–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simko, F.; Hrenak, J.; Adamcova, M.; Paulis, L. Renin-angiotensin-aldosterone system: Friend or foe-the matter of balance. Insight on history, therapeutic implications and COVID-19 interactions. Int. J. Mol. Sci. 2021, 22, 3217. [Google Scholar] [CrossRef] [PubMed]
- Grossman, E.; Laudon, M.; Zisapel, N. Effect of melatonin on nocturnal blood pressure: Meta-analysis of randomized controlled trials. Vasc. Health Risk Manag. 2011, 7, 577–584. [Google Scholar] [CrossRef] [Green Version]
- Simko, F. Chronobiology of blood pressure: Emerging implications of melatonin. Eur. J. Clin. Invest. 2012, 42, 1252–1254. [Google Scholar] [CrossRef] [PubMed]
- Simko, F.; Pechanova, O. Potential roles of melatonin and chronotherapy among the new trends in hypertension treatment. J. Pineal. Res. 2009, 47, 127–133. [Google Scholar] [CrossRef]
- Cvikova, D.; Sutovska, H.; Babarikova, K.; Molcan, L. Hypotensive effects of melatonin in rats: Focus on the model, measurement, application, and main mechanisms. Hypertens. Res. 2022, 45, 1929–1944. [Google Scholar] [CrossRef]
- Simko, F.; Pechanova, O.; Pelouch, V.; Krajcirovicova, K.; Mullerova, M.; Bednarova, K.; Adamcova, M.; Paulis, L. Effect of melatonin, captopril, spironolactone and simvastatin on blood pressure and left ventricular remodelling in spontaneously hypertensive rats. J. Hypertens. 2009, 27, S5–S10. [Google Scholar] [CrossRef]
- Simko, F.; Pechanova, O.; Pelouch, V.; Krajcirovicova, K.; Celec, P.; Palffy, R.; Bednarova, K.; Vrankova, S.; Adamcova, M.; Paulis, L. Continuous light and L-NAME-induced left ventricular remodelling: Different protection with melatonin and captopril. J. Hypertens. 2010, 28 (Suppl. S1), S13–S18. [Google Scholar] [CrossRef] [PubMed]
- Simko, F.; Pechanova, O.; Bednarova, K.R.; Krajcirovicova, K.; Celec, P.; Kamodyova, N.; Zorad, S.; Kucharska, J.; Gvozdjakova, A.; Adamcova, M.; et al. Hypertension and cardiovascular remodelling in rats exposed to continuous light: Protection by ACE-inhibition and melatonin. Mediat. Inflamm. 2014, 2014, 703175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simko, F.; Pechanova, O.; Repova, K.; Aziriova, S.; Krajcirovicova, K.; Celec, P.; Tothova, L.; Vrankova, S.; Balazova, L.; Zorad, S.; et al. Lactacystin-induced model of hypertension in rats: Effects of melatonin and captopril. Int. J. Mol. Sci. 2017, 18, 1612. [Google Scholar] [CrossRef] [PubMed]
- Simko, F.; Paulis, L. Antifibrotic effect of melatonin—Perspective protection in hypertensive heart disease. Int. J. Cardiol. 2013, 168, 2876–2877. [Google Scholar] [CrossRef] [PubMed]
- Simko, F.; Bednarova, K.R.; Krajcirovicova, K.; Hrenak, J.; Celec, P.; Kamodyova, N.; Gajdosechova, L.; Zorad, S.; Adamcova, M. Melatonin reduces cardiac remodeling and improves survival in rats with isoproterenol-induced heart failure. J. Pineal. Res. 2014, 57, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Repová-Bednárová, K.; Aziriová, S.; Hrenák, J.; Krajčírovičová, K.; Adamcová, M.; Paulis, L.; Simko, F. Effect of captopril and melatonin on fibrotic rebuilding of the aorta in 24 hour light-induced hypertension. Physiol. Res. 2013, 62, S135–S141. [Google Scholar] [CrossRef]
- Jonas, M.; Garfinkel, D.; Zisapel, N.; Laudon, M.; Grossman, E. Impaired nocturnal melatonin secretion in non-dipper hypertensive patients. Blood Press 2003, 12, 19–24. [Google Scholar]
- Baltatu, O.; Janssen, B.J.; Bricca, G.; Plehm, R.; Monti, J.; Ganten, D.; Bader, M. Alterations in blood pressure and heart rate variability in transgenic rats with low brain angiotensinogen. Hypertension 2001, 37, 408–413. [Google Scholar] [CrossRef] [Green Version]
- Baltatu, O.; Afeche, S.C.; José dos Santos, S.H.; Campos, L.A.; Barbosa, R.; Michelini, L.C.; Bader, M.; Cipolla-Neto, J. Locally synthesized angiotensin modulates pineal melatonin generation. J. Neurochem. 2002, 80, 328–334. [Google Scholar] [CrossRef] [Green Version]
- Abrahão, M.V.; Dos Santos, N.F.T.; Kuwabara, W.M.T.; do Amaral, F.G.; do Carmo Buonfiglio, D.; Peres, R.; Vendrame, R.F.A.; Flávio da Silveira, P.; Cipolla-Neto, J.; Baltatu, O.C.; et al. Identification of insulin-regulated aminopeptidase (IRAP) in the rat pineal gland and the modulation of melatonin synthesis by angiotensin IV. Brain Res. 2019, 1704, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Campos, L.A.; Cipolla-Neto, J.; Amaral, F.G.; Michelini, L.C.; Bader, M.; Baltatu, O.C. The angiotensin-melatonin axis. Int. J. Hypertens. 2013, 2013, 521783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishigaki, S.; Ohashi, N.; Isobe, S.; Tsuji, N.; Iwakura, T.; Ono, M.; Sakao, Y.; Tsuji, T.; Kato, A.; Miyajima, H.; et al. Impaired endogenous nighttime melatonin secretion relates to intrarenal renin-angiotensin system activation and renal damage in patients with chronic kidney disease. Clin. Exp. Nephrol. 2016, 20, 878–884. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, A.; Kobori, H. Independent regulation of renin-angiotensin-aldosterone system in the kidney. Clin. Exp. Nephrol. 2018, 22, 1231–1239. [Google Scholar] [CrossRef] [Green Version]
- Ishigaki, S.; Ohashi, N.; Matsuyama, T.; Isobe, S.; Tsuji, N.; Iwakura, T.; Fujikura, T.; Tsuji, T.; Kato, A.; Miyajima, H.; et al. Melatonin ameliorates intrarenal renin-angiotensin system in a 5/6 nephrectomy rat model. Clin. Exp. Nephrol. 2018, 22, 539–549. [Google Scholar] [CrossRef]
- Ohashi, N.; Ishigaki, S.; Isobe, S. The pivotal role of melatonin in ameliorating chronic kidney disease by suppression of the renin-angiotensin system in the kidney. Hypertens. Res. 2019, 42, 761–768. [Google Scholar] [CrossRef]
- Simko, F.; Baka, T.; Krajcirovicova, K.; Repova, K.; Aziriova, S.; Zorad, S.; Poglitsch, M.; Adamcova, M.; Reiter, R.J.; Paulis, L. Effect of melatonin on the renin-angiotensin-aldosterone system in l-NAME-induced hypertension. Molecules 2018, 23, 265. [Google Scholar] [CrossRef] [Green Version]
- Marinova, C.; Persengiev, S.; Konakchieva, R.; Ilieva, A.; Patchev, V. Melatonin effects on glucocorticoid receptors in rat brain and pituitary: Significance in adrenocortical regulation. Int. J. Biochem. 1991, 23, 479–481. [Google Scholar] [CrossRef]
- Quiros, I.; Mayo, J.C.; Garcia-Suarez, O.; Hevia, D.; Martin, V.; Rodríguez, C.; Sainz, R.M. Melatonin prevents glucocorticoid inhibition of cell proliferation and toxicity in hippocampal cells by reducing glucocorticoid receptor nuclear translocation. J. Steroid Biochem. Mol. Biol. 2008, 110, 116–124. [Google Scholar] [CrossRef]
- Konakchieva, R.; Mitev, Y.; Almeida, O.F.; Patchev, V.K. Chronic melatonin treatment counteracts glucocorticoid-induced dysregulation of the hypothalamic-pituitary-adrenal axis in the rat. Neuroendocrinology 1998, 67, 171–180. [Google Scholar] [CrossRef]
- Cardinali, D.P.; Fontao, R.; Falco, N.; Ruiz, J.I.; Caccuri, R.; Furio, A.M. Neuroprotective effect of melatonin on glucocorticoid toxicity in the rat hippocampus. Open Physiol. J. 2008, 1, 23–27. [Google Scholar] [CrossRef]
- Gomaa, A.M.; Galal, H.M.; Abou-Elgait, A.T. Neuroprotective effects of melatonin administration against chronic immobilization stress in rats. Int. J. Physiol. Pathophysiol. Pharmacol. 2017, 9, 16–27. [Google Scholar] [PubMed]
- Wang, X.; Wang, Z.; Cao, J.; Dong, Y.; Chen, Y. Melatonin ameliorates anxiety-like behaviors induced by sleep deprivation in mice: Role of oxidative stress, neuroinflammation, autophagy and apoptosis. Brain Res. Bull 2021, 174, 161–172. [Google Scholar] [CrossRef]
- Castaño, M.Y.; Garrido, M.; Rodríguez, A.B.; Gómez, M.Á. Melatonin improves mood status and quality of life and decreases cortisol levels in fibromyalgia. Biol. Res. Nurs. 2019, 21, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.-X.; Manchester, L.C.; Terron, M.P.; Flores, L.J.; Reiter, R.J. One molecule, many derivatives: A never-ending interaction of melatonin with reactive oxygen and nitrogen species? J. Pineal. Res. 2007, 42, 28–42. [Google Scholar] [CrossRef]
- Bettahi, I.; Pozo, D.; Osuna, C.; Reiter, R.J.; Acuña-Castroviejo, D.; Guerrero, J.M. Melatonin reduces nitric oxide synthase activity in rat hypothalamus. J. Pineal. Res. 1996, 20, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Zarezadeh, M.; Barzegari, M.; Aghapour, B.; Adeli, S.; Khademi, F.; Musazadeh, V.; Jamilian, P.; Jamilian, P.; Fakhr, L.; Chehregosha, F.; et al. Melatonin effectiveness in amelioration of oxidative stress and strengthening of antioxidant defense system: Findings from a systematic review and dose-response meta-analysis of controlled clinical trials. Clin. Nutr. ESPEN 2022, 48, 109–120. [Google Scholar] [CrossRef]
- Morvaridzadeh, M.; Sadeghi, E.; Agah, S.; Nachvak, S.M.; Fazelian, S.; Moradi, F.; Persad, E.; Heshmati, J. Effect of melatonin supplementation on oxidative stress parameters: A systematic review and meta-analysis. Pharmacol. Res. 2020, 161, 105210. [Google Scholar] [CrossRef]
- Zhang, L.; Guo, H.-L.; Zhang, H.-Q.; Xu, T.-Q.; He, B.; Wang, Z.-H.; Yang, Y.-P.; Tang, X.-D.; Zhang, P.; Liu, F.-E. Melatonin prevents sleep deprivation-associated anxiety-like behavior in rats: Role of oxidative stress and balance between GABAergic and glutamatergic transmission. Am. J. Transl. Res. 2017, 9, 2231–2242. [Google Scholar]
- Ergenc, M.; Ozacmak, H.S.; Turan, I.; Ozacmak, V.H. Melatonin reverses depressive and anxiety like-behaviours induced by diabetes: Involvement of oxidative stress, age, rage and S100B levels in the hippocampus and prefrontal cortex of rats. Arch. Physiol. Biochem. 2022, 128, 402–410. [Google Scholar] [CrossRef]
- Lacoste, B.; Angeloni, D.; Dominguez-Lopez, S.; Calderoni, S.; Mauro, A.; Fraschini, F.; Descarries, L.; Gobbi, G. Anatomical and cellular localization of melatonin MT1 and MT2 receptors in the adult rat brain. J. Pineal. Res. 2015, 58, 397–417. [Google Scholar] [CrossRef] [PubMed]
- Comai, S.; Lopez-Canul, M.; De Gregorio, D.; Posner, A.; Ettaoussi, M.; Guarnieri, F.C.; Gobbi, G. Melatonin MT1 receptor as a novel target in neuropsychopharmacology: MT1 ligands, pathophysiological and therapeutic implications, and perspectives. Pharmacol. Res. 2019, 144, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Gobbi, G.; Blier, P. Effect of neurokinin-1 receptor antagonists on serotoninergic, noradrenergic and hippocampal neurons: Comparison with antidepressant drugs. Peptides 2005, 26, 1383–1393. [Google Scholar] [CrossRef] [PubMed]
- Kholghi, G.; Eskandari, M.; Shokouhi Qare Saadlou, M.-S.; Zarrindast, M.-R.; Vaseghi, S. Night shift hormone: How does melatonin affect depression? Physiol. Behav. 2022, 252, 113835. [Google Scholar] [CrossRef] [PubMed]
- Naguib, M.; Gottumukkala, V.; Goldstein, P.A. Melatonin and anesthesia: A clinical perspective. J. Pineal. Res. 2007, 42, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Popova, E. Role of dopamine in retinal function. In Webvision: The Organization of the Retina and Visual System; Kolb, H., Fernandez, E., Nelson, R., Eds.; University of Utah Health Sciences Center: Salt Lake City, UT, USA, 1995. [Google Scholar]
- González, S.; Moreno-Delgado, D.; Moreno, E.; Pérez-Capote, K.; Franco, R.; Mallol, J.; Cortés, A.; Casadó, V.; Lluís, C.; Ortiz, J.; et al. Circadian-related heteromerization of adrenergic and dopamine d4 receptors modulates melatonin synthesis and release in the pineal gland. PLoS Biol. 2012, 10, e1001347. [Google Scholar] [CrossRef]
- Pandi-Perumal, S.R.; Trakht, I.; Srinivasan, V.; Spence, D.W.; Maestroni, G.J.M.; Zisapel, N.; Cardinali, D.P. Physiological effects of melatonin: Role of melatonin receptors and signal transduction pathways. Prog. Neurobiol. 2008, 85, 335–353. [Google Scholar] [CrossRef]
- Zisapel, N. Melatonin-dopamine interactions: From basic neurochemistry to a clinical setting. Cell Mol. Neurobiol. 2001, 21, 605–616. [Google Scholar] [CrossRef]
- Firsov, M.L.; Astakhova, L.A. Role of dopamine as a regulator of vertebrate photoreceptors. Ross Fiziol Zh Im I M Sechenova 2014, 100, 777–790. [Google Scholar]
- Reiter, R.J.; Sharma, R.; Rosales-Corral, S.; de Mange, J.; Phillips, W.T.; Tan, D.X.; Bitar, R.D. Melatonin in ventricular and subarachnoid cerebrospinal fluid: Its function in the neural glymphatic network and biological significance for neurocognitive health. Biochem. Biophys. Res. Commun. 2022, 605, 70–81. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Repova, K.; Baka, T.; Krajcirovicova, K.; Stanko, P.; Aziriova, S.; Reiter, R.J.; Simko, F. Melatonin as a Potential Approach to Anxiety Treatment. Int. J. Mol. Sci. 2022, 23, 16187. https://doi.org/10.3390/ijms232416187
Repova K, Baka T, Krajcirovicova K, Stanko P, Aziriova S, Reiter RJ, Simko F. Melatonin as a Potential Approach to Anxiety Treatment. International Journal of Molecular Sciences. 2022; 23(24):16187. https://doi.org/10.3390/ijms232416187
Chicago/Turabian StyleRepova, Kristina, Tomas Baka, Kristina Krajcirovicova, Peter Stanko, Silvia Aziriova, Russel J. Reiter, and Fedor Simko. 2022. "Melatonin as a Potential Approach to Anxiety Treatment" International Journal of Molecular Sciences 23, no. 24: 16187. https://doi.org/10.3390/ijms232416187
APA StyleRepova, K., Baka, T., Krajcirovicova, K., Stanko, P., Aziriova, S., Reiter, R. J., & Simko, F. (2022). Melatonin as a Potential Approach to Anxiety Treatment. International Journal of Molecular Sciences, 23(24), 16187. https://doi.org/10.3390/ijms232416187