
Non-Invasive Assessment of Locally Overexpressed Human Adenosine 2A Receptors in the Heart of Transgenic Mice

 ,
,  ,
,  , ,
, ,  ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Results
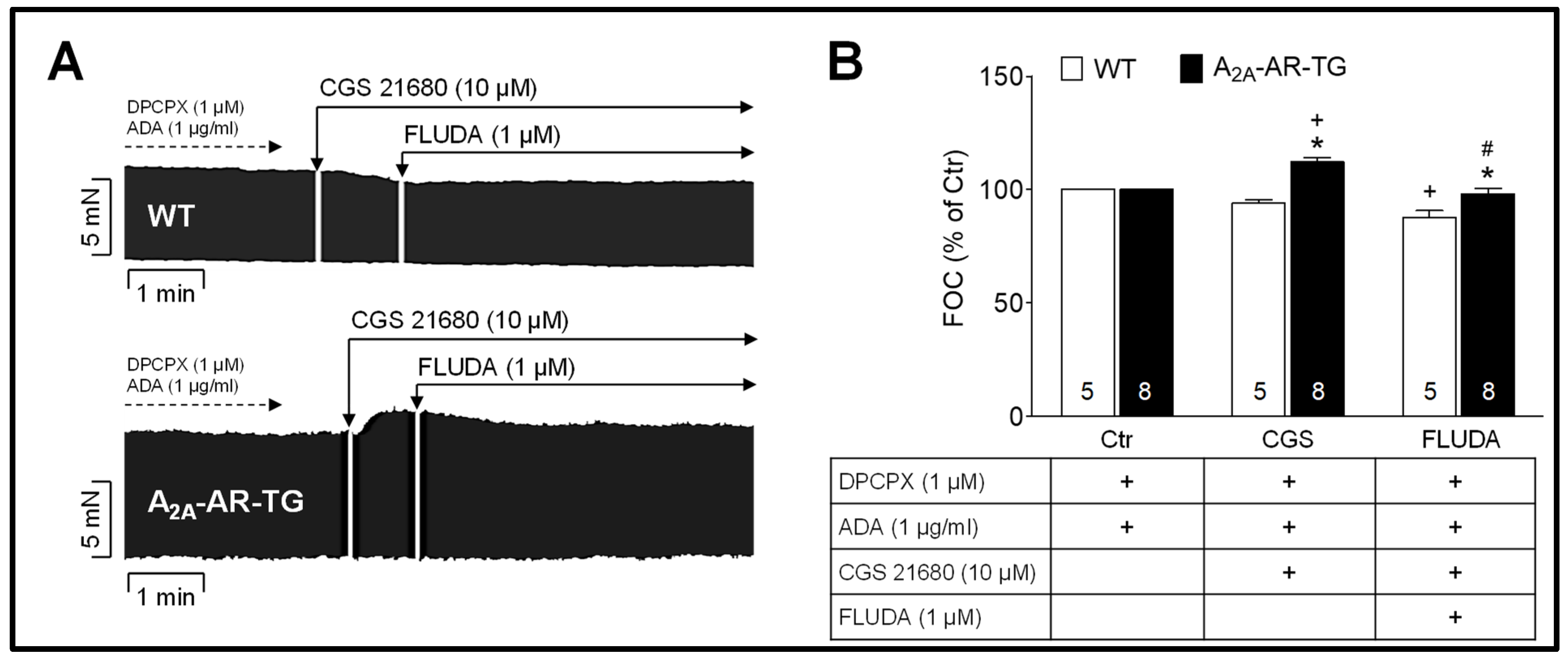
2.1. Impact of FLUDA on the Atrial Force of Contraction (FOC) in Electrically Stimulated Atrial Preparations
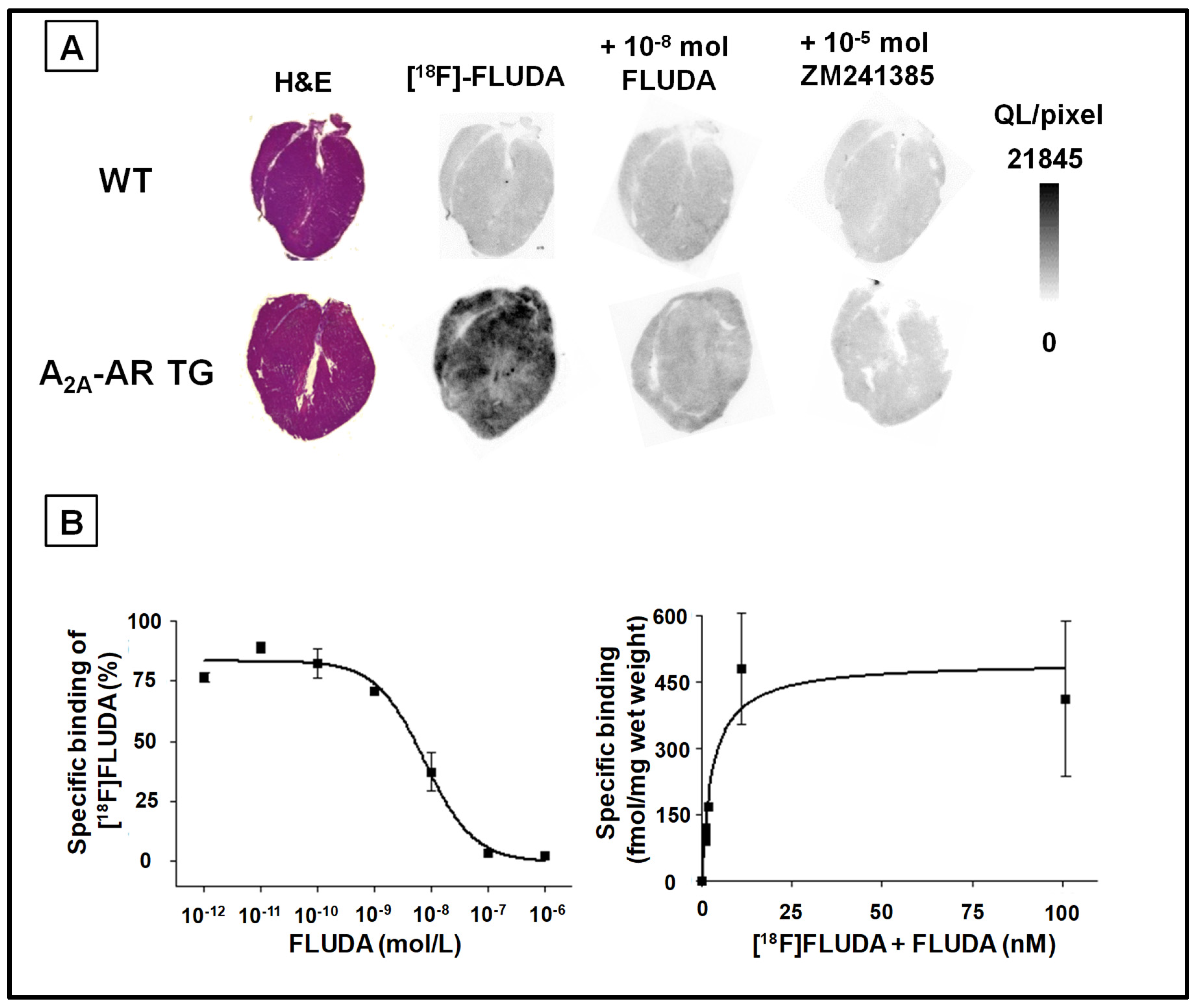
2.2. In Vitro Binding of [18F]FLUDA to the A2A-AR in Heart Samples
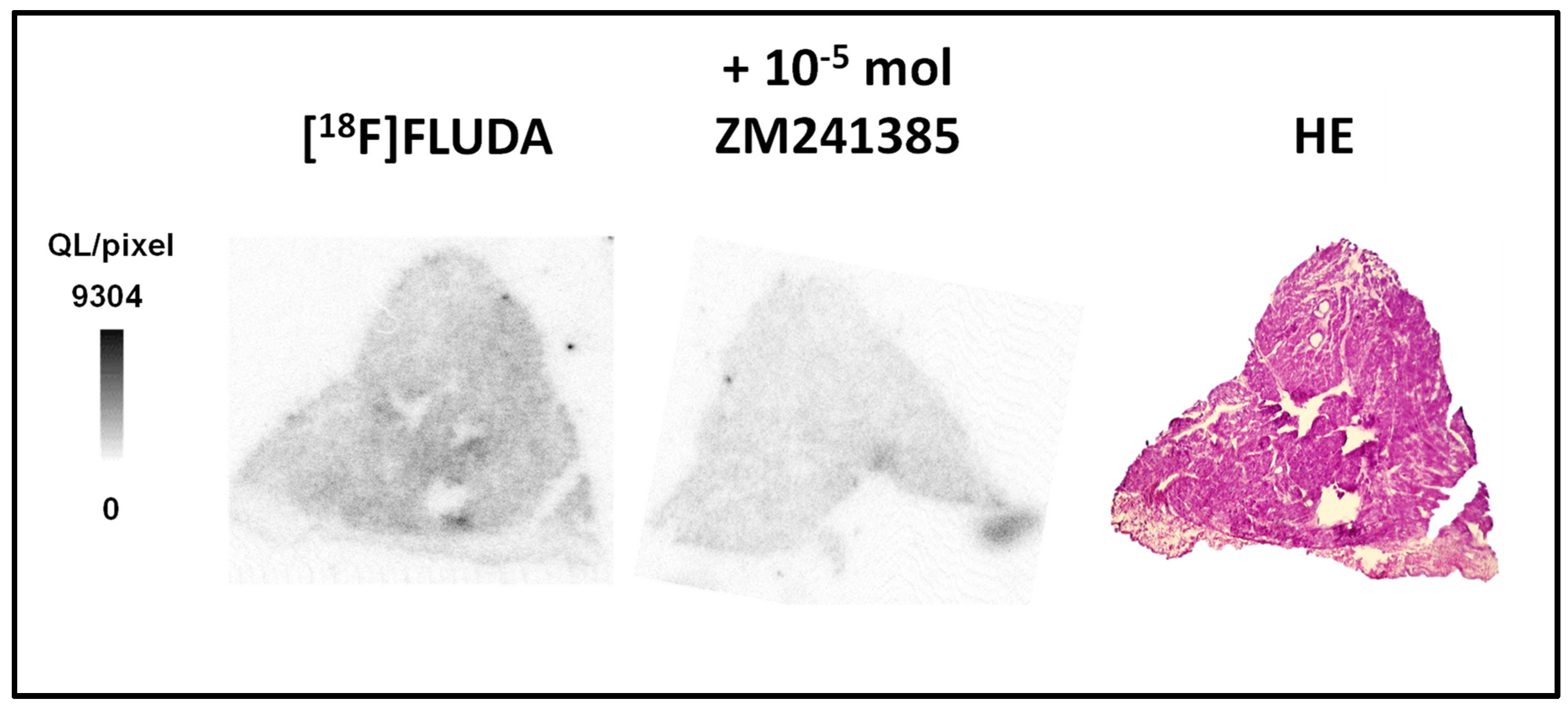
2.3. In Vitro Binding Studies of [18F]FLUDA in Human Atrial Samples
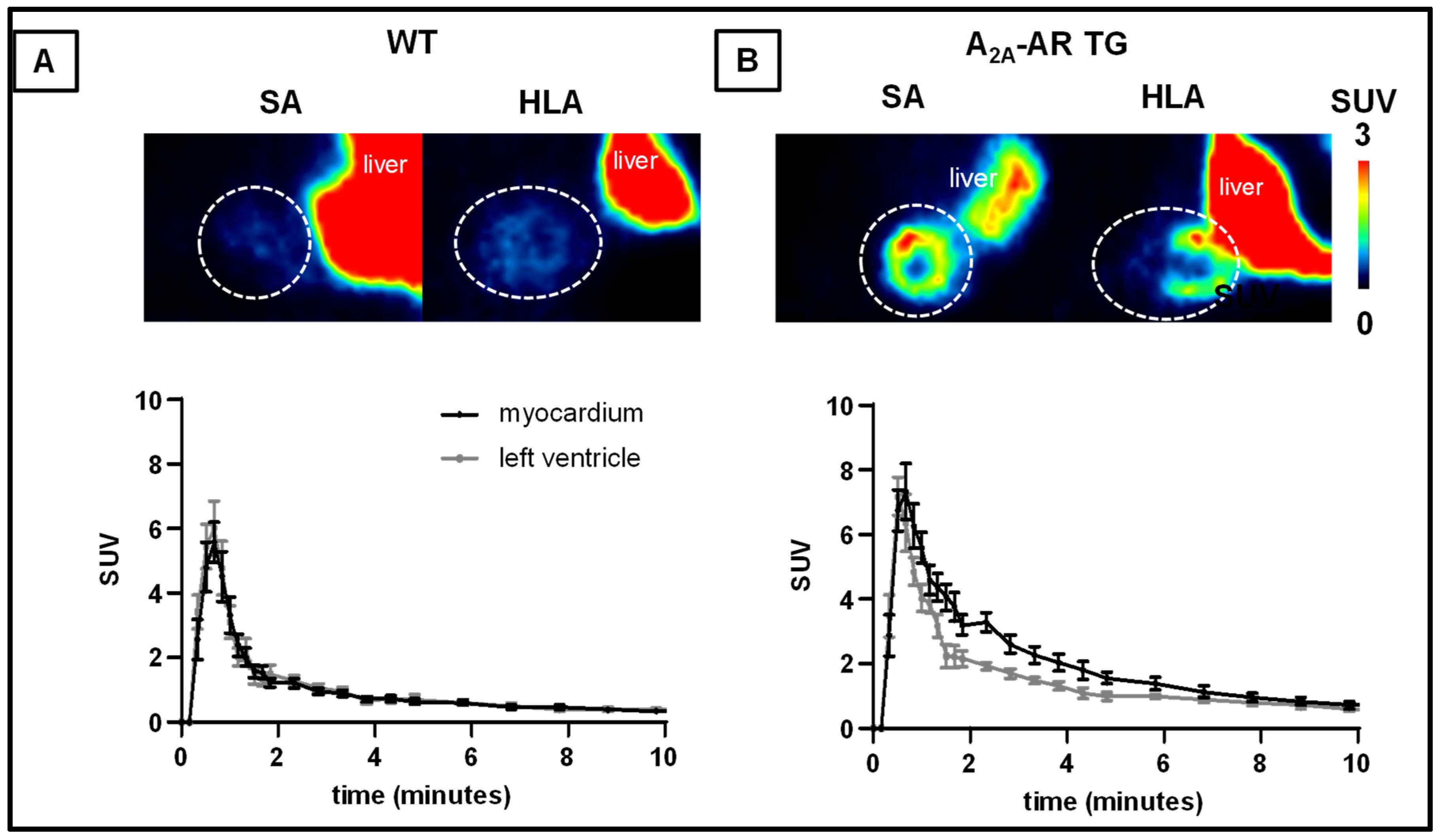
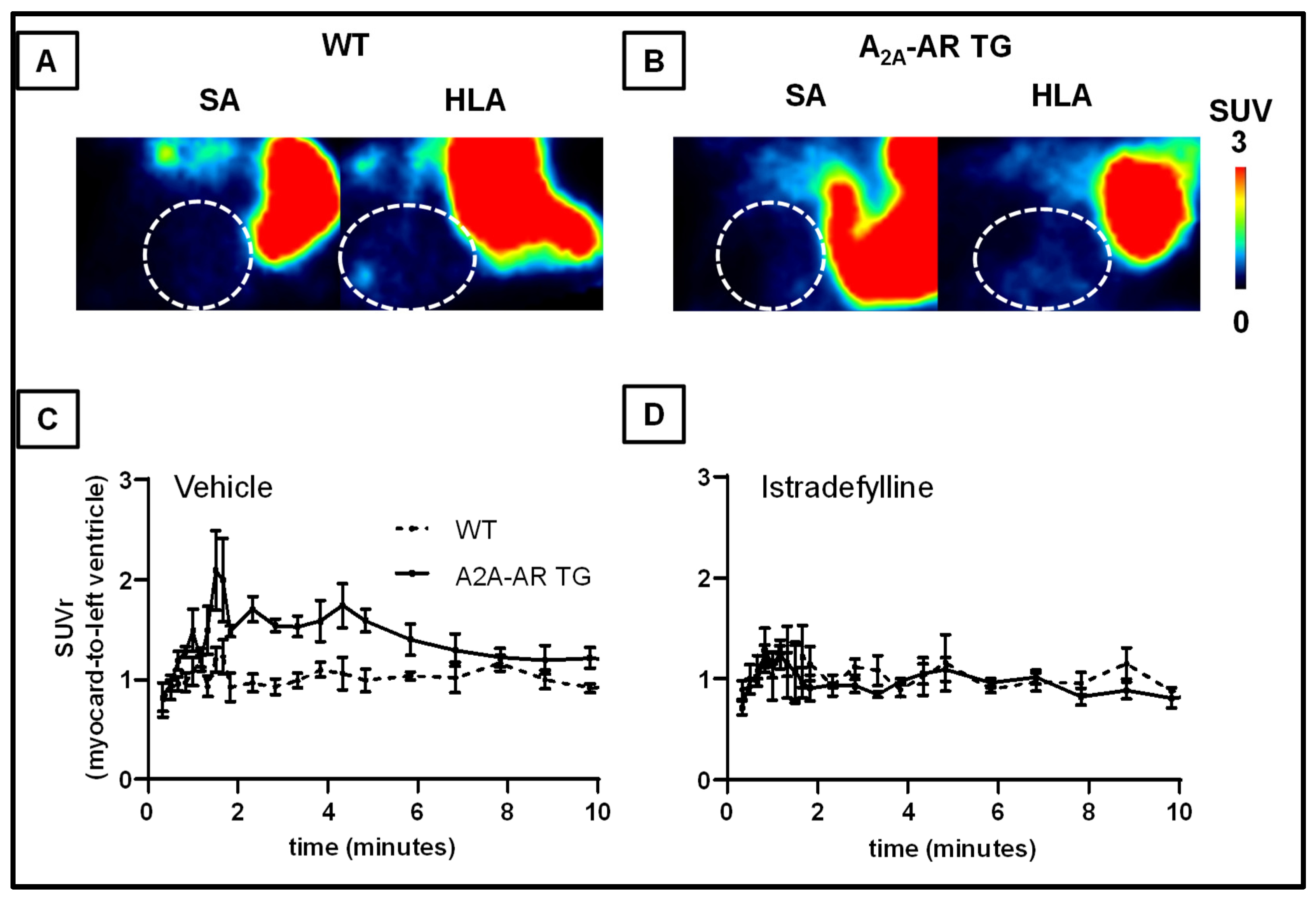
2.4. In Vivo Uptake into the Heart and Other Tissues of [18F]FLUDA in WT and A2A-AR TG under Baseline Conditions and after Blocking with Tozadenant and Istradefylline
3. Discussion
4. Materials and Methods
4.1. General Information
4.2. Animals
4.3. Human Atrial Preparations
4.4. Contractile Function
4.5. In Vitro Autoradiography
4.6. Small-Animal PET/MR Experiments
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reiss, A.B.; Grossfeld, D.; Kasselman, L.J.; Renna, H.A.; Vernice, N.A.; Drewes, W.; Konig, J.; Carsons, S.E.; DeLeon, J. Adenosine and the Cardiovascular System. Am. J. Cardiovasc. Drugs 2019, 19, 449–464. [Google Scholar] [CrossRef] [PubMed]
- Peart, J.N.; Headrick, J.P. Adenosinergic Cardioprotection: Multiple Receptors, Multiple Pathways. Pharmacol. Ther. 2007, 114, 208–221. [Google Scholar] [CrossRef] [PubMed]
- Olsson, R.A.; Pearson, J.D. Cardiovascular Purinoreceptors. Physiol. Rev. 1990, 70, 761–845. [Google Scholar] [CrossRef] [PubMed]
- Thiel, M.; Caldwell, C.C.; Sitkovsky, M.V. The Critical Role of Adenosine A2A Receptors in Downregulation of Inflammation and Immunity in the Pathogenesis of Infectious Diseases. Microbes Infect. 2003, 5, 515–526. [Google Scholar] [CrossRef]
- Dobson, J.G. Reduction by Adenosine of the Isoproterenol-Induced Increase in Cyclic Adenosine 3’,5’-Monophosphate Formation and Glycogen Phosphorylase Activity in Rat Heart Muscle. Circ. Res. 1978, 43, 785–792. [Google Scholar] [CrossRef] [Green Version]
- Dobson, J.G. Mechanism of Adenosine Inhibition of Catecholamine-Induced Responses in Heart. Circ. Res. 1983, 52, 151–160. [Google Scholar] [CrossRef] [Green Version]
- Makujina, S.R.; Sabouni, M.H.; Bhatia, S.; Douglas, F.L.; Mustafa, S.J. Vasodilatory Effects of Adenosine A2 Receptor Agonists CGS 21680 and CGS 22492 in Human Vasculature. Eur. J. Pharmacol. 1992, 221, 243–247. [Google Scholar] [CrossRef]
- Tikh, E.I.; Fenton, R.A.; Dobson, J.G. Contractile Effects of Adenosine A1 and A2A Receptors in Isolated Murine Hearts. Am. J. Physiol. Heart Circ. Physiol. 2006, 290, 348–356. [Google Scholar] [CrossRef] [Green Version]
- Lasley, R.D.; Kristo, G.; Keith, B.J.; Mentzer, R.M. The A2a/A2b Receptor Antagonist ZM-241385 Blocks the Cardioprotective Effect of Adenosine Agonist Pretreatment in in Vivo Rat Myocardium. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, 426–431. [Google Scholar] [CrossRef] [Green Version]
- Llach, A.; Molina, C.E.; Prat-Vidal, C.; Fernandes, J.; Casad, V.; Ciruela, F.; Llus, C.; Franco, R.; Cinca, J.; Hove-Madsen, L. Abnormal Calcium Handling in Atrial Fibrillation Is Linked to Up-Regulation of Adenosine A 2A Receptors. Eur. Heart J. 2011, 32, 721–729. [Google Scholar] [CrossRef] [Green Version]
- Molina, C.E.; Llach, A.; Herraiz-Martínez, A.; Tarifa, C.; Barriga, M.; Wiegerinck, R.F.; Fernandes, J.; Cabello, N.; Vallmitjana, A.; Benitéz, R.; et al. Prevention of Adenosine A2A Receptor Activation Diminishes Beat-to-Beat Alternation in Human Atrial Myocytes. Basic Res. Cardiol. 2016, 111, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asakura, M.; Asanuma, H.; Kim, J.; Liao, Y.; Nakamaru, K.; Fujita, M.; Komamura, K.; Isomura, T.; Fukurawa, H.; Tomoike, H.; et al. Impact of Adenosine Receptor Signaling and Metabolism on Pathophysiology in Patients with Chronic Heart Failure. Hypertens. Res. 2007, 30, 781–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Silva, J.S.; Gabriel-Costa, D.; Sudo, R.T.; Wang, H.; Groban, L.; Ferraz, E.B.; Nascimento, J.H.M.; Fraga, C.A.M.; Barreiro, E.J.; Zapata-Sudo, G. Adenosine A2A Receptor Agonist Prevents Cardiac Remodeling and Dysfunction in Spontaneously Hypertensive Male Rats after Myocardial Infarction. Drug Des. Dev. Ther. 2017, 11, 553–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glover, D.K.; Ruiz, M.; Takehana, K.; Petruzella, F.D.; Rieger, J.M.; Macdonald, T.L.; Watson, D.D.; Linden, J.; Beller, G.A. Cardioprotection by Adenosine A2A Agonists in a Canine Model of Myocardial Stunning Produced by Multiple Episodes of Transient Ischemia. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H3164–H3171. [Google Scholar] [CrossRef] [Green Version]
- Laubach, V.E.; French, B.A.; Okusa, M.D. Targeting of Adenosine Receptors in Ischemia-Reperfusion Injury. Expert Opin. Targets 2011, 15, 103–118. [Google Scholar] [CrossRef]
- Sakata, M.; Ishibashi, K.; Imai, M.; Wagatsuma, K.; Ishii, K.; Zhou, X.; de Vries, E.F.J.; Elsinga, P.H.; Ishiwata, K.; Toyohara, J. Initial Evaluation of an Adenosine A 2A Receptor Ligand, 11 C-Preladenant, in Healthy Human Subjects. J. Nucl. Med. 2017, 58, 1464–1470. [Google Scholar] [CrossRef] [Green Version]
- Barret, O.; Hannestad, J.; Vala, C.; Alagille, D.; Tavares, A.; Laruelle, M.; Jennings, D.; Marek, K.; Russell, D.; Seibyl, J.; et al. Characterization in Humans of 18F-MNI-444, a PET Radiotracer for Brain Adenosine 2A Receptors. J. Nucl. Med. 2015, 56, 586–591. [Google Scholar] [CrossRef] [Green Version]
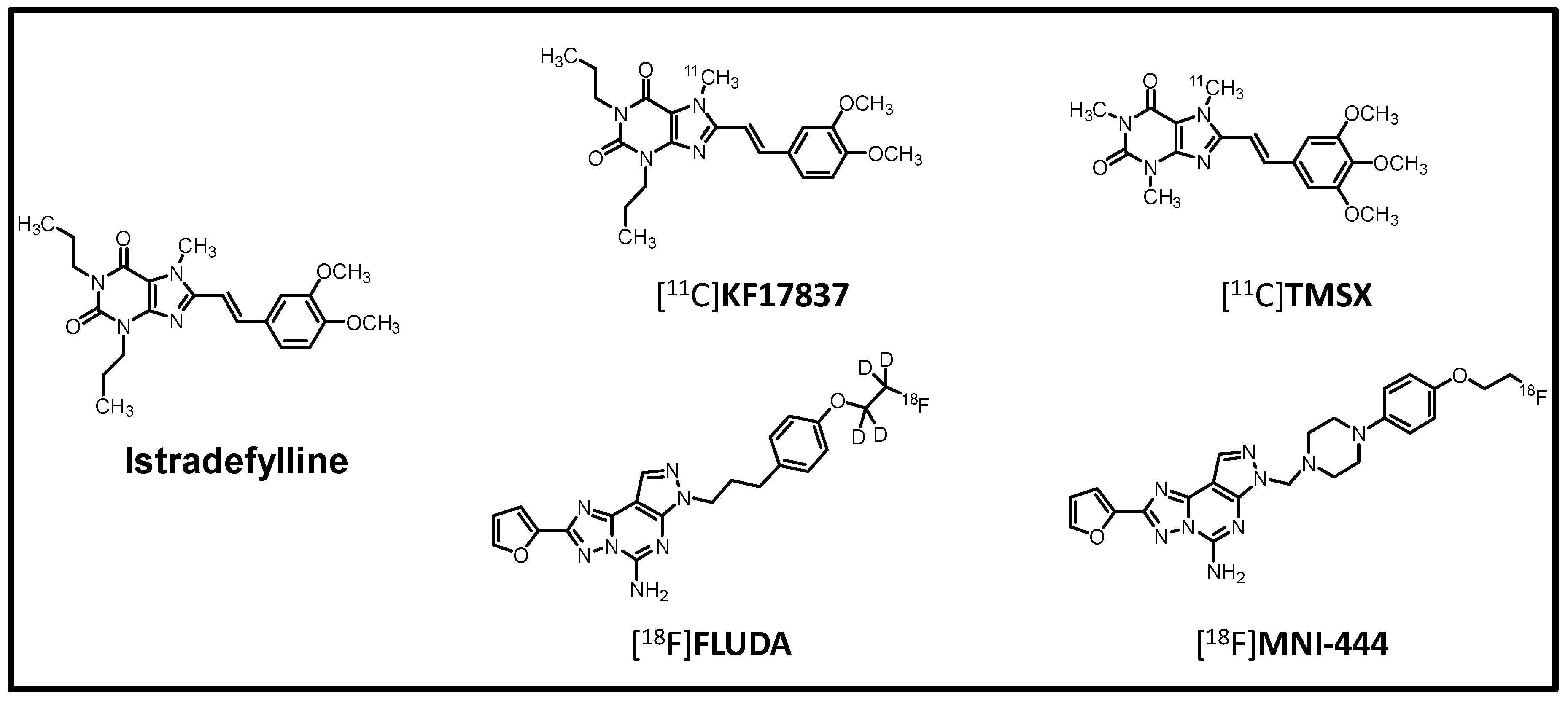
- Lai, T.H.; Toussaint, M.; Teodoro, R.; Dukić-Stefanović, S.; Gündel, D.; Ludwig, F.; Wenzel, B.; Schröder, S.; Sattler, B.; Moldovan, R.-P.; et al. Improved in vivo PET Imaging of the Adenosine A2A Receptor in the Brain Using [18F]FLUDA, a Deuterated Radiotracer with High Metabolic Stability. Eur. J. Nucl. Med. Mol. Imaging 2021, 48, 2727–2736. [Google Scholar] [CrossRef]
- Ishiwata, K.; Sakiyama, Y.; Sakiyama, T.; Shimada, J.; Toyama, H.; Oda, K.; Suzuki, F.; Senda, M. Myocardial Adenosine A2a Receptor Imaging of Rabbit by PET with [11C]KF17837. Ann. Nucl. Med. 1997, 11, 219–225. [Google Scholar] [CrossRef]
- Ishiwata, K.; Kawamura, K.; Kimura, Y.; Oda, K.; Ishii, K. Potential of an Adenosine A2A Receptor Antagonist [ 11C]TMSX for Myocardial Imaging by Positron Emission Tomography: A First Human Study. Ann. Nucl. Med. 2003, 17, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Boknik, P.; Drzewiecki, K.; Eskandar, J.; Gergs, U.; Grote-Wessels, S.; Fabritz, L.; Kirchhof, P.; Müller, F.U.; Stümpel, F.; Schmitz, W.; et al. Phenotyping of Mice with Heart Specific Overexpression of A2A-Adenosine Receptors: Evidence for Cardioprotective Effects of A2A-Adenosine Receptors. Front. Pharmacol. 2018, 9, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinonen, I.; Nesterov, S.V.; Liukko, K.; Kemppainen, J.; Någren, K.; Luotolahti, M.; Virsu, P.; Oikonen, V.; Nuutila, P.; Kujala, U.M.; et al. Myocardial Blood Flow and Adenosine A2A Receptor Density in Endurance Athletes and Untrained Men. J. Physiol. 2008, 586, 5193–5202. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, M.; Kimura, Y.; Tokizawa, K.; Ishii, K.; Oda, K.; Sasaki, T.; Nakamura, Y.; Muraoka, I.; Ishiwata, K. Greater Adenosine A2A Receptor Densities in Cardiac and Skeletal Muscle in Endurance-Trained Men: A [11C]TMSX PET Study. Nucl. Med. Biol. 2005, 32, 831–836. [Google Scholar] [CrossRef] [PubMed]
- Khanapur, S.; Paul, S.; Shah, A.; Vatakuti, S.; Koole, M.J.B.; Zijlma, R.; Dierckx, R.A.J.O.; Luurtsema, G.; Garg, P.; van Waarde, A.; et al. Development of [18F]-Labeled Pyrazolo[4,3-e ]-1,2,4-Triazolo[1, 5-c ]Pyrimidine (SCH442416) Analogs for the Imaging of Cerebral Adenosine A 2A Receptors with Positron Emission Tomography. J. Med. Chem. 2014, 57, 6765–6780. [Google Scholar] [CrossRef]
- Khanapur, S.; van Waarde, A.; Dierckx, R.A.J.O.; Elsinga, P.H.; Koole, M.J.B. Preclinical Evaluation and Quantification of 18F-Fluoroethyl and 18F-Fluoropropyl Analogs of SCH442416 as Radioligands for PET Imaging of the Adenosine A2A Receptor in Rat Brain. J. Nucl. Med. 2017, 58, 466–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hove-Madsen, L.; Prat-Vidal, C.; Llach, A.; Ciruela, F.; Casadó, V.; Lluis, C.; Bayes-Genis, A.; Cinca, J.; Franco, R. Adenosine A2A Receptors Are Expressed in Human Atrial Myocytes and Modulate Spontaneous Sarcoplasmic Reticulum Calcium Release. Cardiovasc. Res. 2006, 72, 292–302. [Google Scholar] [CrossRef] [Green Version]
- Varani, K.; Laghi-Pasini, F.; Camurri, A.; Capecchi, P.L.; Maccherini, M.; Diciolla, F.; Ceccatelli, L.; Enea Lazzerini, P.; Ulouglu, C.; Cattabeni, F.; et al. Changes of Peripheral A 2A Adenosine Receptors in Chronic Heart Failure and Cardiac Transplantation. FASEB J. 2003, 17, 280–282. [Google Scholar] [CrossRef] [Green Version]
- Elkholy, K.O.; Hegazy, O.; Okunade, A.; Aktas, S.; Ajibawo, T. Regadenoson Stress Testing: A Comprehensive Review With a Focused Update. Cureus 2021, 13, e15940. [Google Scholar] [CrossRef]
- Ishiwata, K.; Noguchi, J.; Wakabayashi, S.; Shimada, J.; Ogi, N.; Nariai, T.; Tanaka, A.; Endo, K.; Suzuki, F.; Senda, M. 11C-Labeled KF18446: A Potential Central Nervous System Adenosine A2a Receptor Ligand. J. Nucl. Med. Off. Publ. Soc. Nucl. Med. 2000, 41, 345–354. [Google Scholar]
- Naganawa, M.; Kimura, Y.; Mishina, M.; Manabe, Y.; Chihara, K.; Oda, K.; Ishii, K.; Ishiwata, K. Quantification of Adenosine A2A Receptors in the Human Brain Using [11C]TMSX and Positron Emission Tomography. Eur. J. Nucl. Med. Mol. Imaging 2007, 34, 679–687. [Google Scholar] [CrossRef]
- Gergs, U.; Büxel, M.L.; Bresinsky, M.; Kirchhefer, U.; Fehse, C.; Höring, C.; Hofmann, B.; Marusakova, M.; Čináková, A.; Schwarz, R.; et al. Cardiac Effects of Novel Histamine H 2 Receptor Agonists. J. Pharmacol. Exp. Ther. 2021, 379, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Gergs, U.; Rothkirch, D.; Hofmann, B.; Treede, H.; Robaye, B.; Simm, A.; Müller, C.E.; Neumann, J. Mechanism Underlying the Contractile Activity of UTP in the Mammalian Heart. Eur. J. Pharmacol. 2018, 830, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Motulsky, H. The GraphPad Guide to Analyzing Radioligand Binding Data; GraphPad: San Diego, CA, USA, 1995. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tissue | Time to Peak (min) | TAC Peak Value (SUV) | AUC0–60 (SUV·min) | AUC1–10 (SUV·min) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WT | TG | p-Value | WT | TG | p-Value | WT | TG | p-Value | WT | TG | p-Value | |
| Myocardium | 0.6 ± 0.1 | 0.6 ± 0.0 | 0.105 | 6.3 ± 0.6 | 8.1 ± 0.7 | 0.0437 | 19.6 ± 2.0 | 35.0 ± 3.1 | 0.001 | 8.4 ± 0.9 | 19.3 ± 2.2 | < 0.001 |
| Blood | 0.7 ± 0.0 | 0.5 ± 0.0 | 0.011 | 6.9 ± 0.9 | 7.7 ± 0.7 | 0.242 | 20.4 ± 2.4 | 29.0 ± 1.8 | 0.008 | 8.6 ± 1.2 | 13.9 ± 1.1 | 0.004 |
| Muscle | 3.3 ± 1.0 | 4.0 ± 0.9 | 0.303 | 0.5 ± 0.1 | 0.5 ± 0.1 | 0.327 | 9.6 ± 1.4 | 12.1 ± 1.9 | 0.153 | 3.8 ± 0.5 | 4.7 ± 0.8 | 0.175 |
| Lung | 0.5 ± 0.1 | 0.5 ± 0.0 | 0.500 | 4.2 ± 0.4 | 4.7 ± 0.4 | 0.243 | 5.4 ± 0.5 | 17.8 ± 3.7 | 0.135 | 7.5 ± 0.7 | 7.3 ± 1.5 | 0.128 |
| Liver | 3.5 ± 0.5 | 5.1 ± 1.3 | 0.123 | 6.7 ± 1.1 | 6.1 ± 0.5 | 0.313 | 126 ± 17 | 143 ± 9 | 0.200 | 74.4 ± 9.3 | 61.5 ± 4.4 | 0.119 |
| A2A-AR Group | Tissue | Time to Peak | TAC Peak Value | AUC0–60 | AUC1–10 | ||||
|---|---|---|---|---|---|---|---|---|---|
| Istradyfelline | p-Value | Istradyfelline | p-Value | Istradyfelline | p-Value | Istradyfelline | p-Value | ||
| WT | Myocardium | 87 ± 9 | 0.175 | 130 ± 18 | 0.082 | 116 ± 4 | 0.166 | 87 ± 9 | 0.175 |
| Blood | 75 ± 0 | 0.017 | 125 ± 11 | 0.125 | 117 ± 6 | 0.179 | 106 ± 6 | 0.381 | |
| Muscle | 44 ± 21 | 0.138 | 180 ± 24 | 0.027 | 119 ± 17 | 0.229 | 142 ± 19 | 0.061 | |
| Lung | 84 ± 11 | 0.223 | 109 ± 31 | 0.364 | 131 ± 32 | 0.130 | 133 ± 38 | 0.146 | |
| Liver | 100 ± 25 | 0.500 | 100 ± 11 | 0.499 | 83 ± 3 | 0.210 | 94 ± 5 | 0.410 | |
| TG | Myocardium | 120 ± 17 | 0.107 | 91 ± 15 | 0.302 | 78 ± 8 | 0.083 | 61 ± 9 | 0.032 |
| Blood | 105 ± 11 | 0.313 | 98 ± 9 | 0.442 | 100 ± 11 | 0.500 | 90 ± 9 | 0.222 | |
| Muscle | 58 ± 13 | 0.130 | 138 ± 50 | 0.211 | 84 ± 14 | 0.268 | 99 ± 21 | 0.483 | |
| Lung | 116 ± 11 | 0.090 | 95 ± 6 | 0.380 | 102 ± 2 | 0.470 | 92 ± 5 | 0.401 | |
| Liver | 54 ± 12 | 0.124 | 117 ± 3 | 0.090 | 95 ± 5 | 0.315 | 102 ± 0 | 0.431 | |
| Treatment | Tissue | AUC1–10 Ratio (Tissue-to-Left Ventricle) | AUC1–10 Ratio (Tissue-to-Muscle) | ||||
|---|---|---|---|---|---|---|---|
| WT | A2A-AR TG | p-Value | WT | A2A-AR TG | p-Value | ||
| Vehicle | Myocardium | 1.0 ± 0.0 | 1.4 ± 0.1 | 0.001 | 2.4 ± 0.5 | 4.8 ± 1.2 | 0.028 |
| Lung | 0.7 ± 0.1 | 0.5 ± 0.1 | 0.081 | 1.5 ± 0.2 | 1.7 ± 0.2 | 0.297 | |
| Liver | 7.8 ± 1.0 | 6.0 ± 1.6 | 0.190 | 20.6 ± 2.8 | 16.5 ± 4.4 | 0.223 | |
| Myocardium | 1.0 ± 0.0 | 0.9 ± 0.1 | 0.570 | 1.8 ± 0.4 | 3.0 ± 1.1 | 0.334 | |
| Istradefylline | Lung | 0.8 ± 0.2 | 0.6 ± 0.1 | 0.400 | 1.3 ± 0.2 | 1.6 ± 0.3 | 0.483 |
| Liver | 6.5 ± 0.4 | 5.2 ± 0.8 | 0.193 | 11.5 ± 3.2 | 14.6 ± 2.7 | 0.390 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gündel, D.; Lai, T.H.; Dukic-Stefanovic, S.; Teodoro, R.; Deuther-Conrad, W.; Toussaint, M.; Kopka, K.; Moldovan, R.-P.; Boknik, P.; Hofmann, B.; et al. Non-Invasive Assessment of Locally Overexpressed Human Adenosine 2A Receptors in the Heart of Transgenic Mice. Int. J. Mol. Sci. 2022, 23, 1025. https://doi.org/10.3390/ijms23031025
Gündel D, Lai TH, Dukic-Stefanovic S, Teodoro R, Deuther-Conrad W, Toussaint M, Kopka K, Moldovan R-P, Boknik P, Hofmann B, et al. Non-Invasive Assessment of Locally Overexpressed Human Adenosine 2A Receptors in the Heart of Transgenic Mice. International Journal of Molecular Sciences. 2022; 23(3):1025. https://doi.org/10.3390/ijms23031025
Chicago/Turabian StyleGündel, Daniel, Thu Hang Lai, Sladjana Dukic-Stefanovic, Rodrigo Teodoro, Winnie Deuther-Conrad, Magali Toussaint, Klaus Kopka, Rareş-Petru Moldovan, Peter Boknik, Britt Hofmann, and et al. 2022. "Non-Invasive Assessment of Locally Overexpressed Human Adenosine 2A Receptors in the Heart of Transgenic Mice" International Journal of Molecular Sciences 23, no. 3: 1025. https://doi.org/10.3390/ijms23031025
APA StyleGündel, D., Lai, T. H., Dukic-Stefanovic, S., Teodoro, R., Deuther-Conrad, W., Toussaint, M., Kopka, K., Moldovan, R. -P., Boknik, P., Hofmann, B., Gergs, U., Neumann, J., & Brust, P. (2022). Non-Invasive Assessment of Locally Overexpressed Human Adenosine 2A Receptors in the Heart of Transgenic Mice. International Journal of Molecular Sciences, 23(3), 1025. https://doi.org/10.3390/ijms23031025