
The K+-Dependent and -Independent Pyruvate Kinases Acquire the Active Conformation by Different Mechanisms

 ,
,  ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Effect of Li+, Na+, K+, NH4+, Rb+, Cs+, (CH3)4N+ and (CH3)3N+-CH2-CH2-OH (Choline) on the Activities of WT-RMPK and RMPK Mutants
2.2. Effect of Activating Monovalent Cations to WT-RMPK and RMPK Mutants in the Presence of 250 mM Ionic Strength Maintained with (CH3)4N+
2.3. Kinetics of WT-RMPK and RMPK Mutants
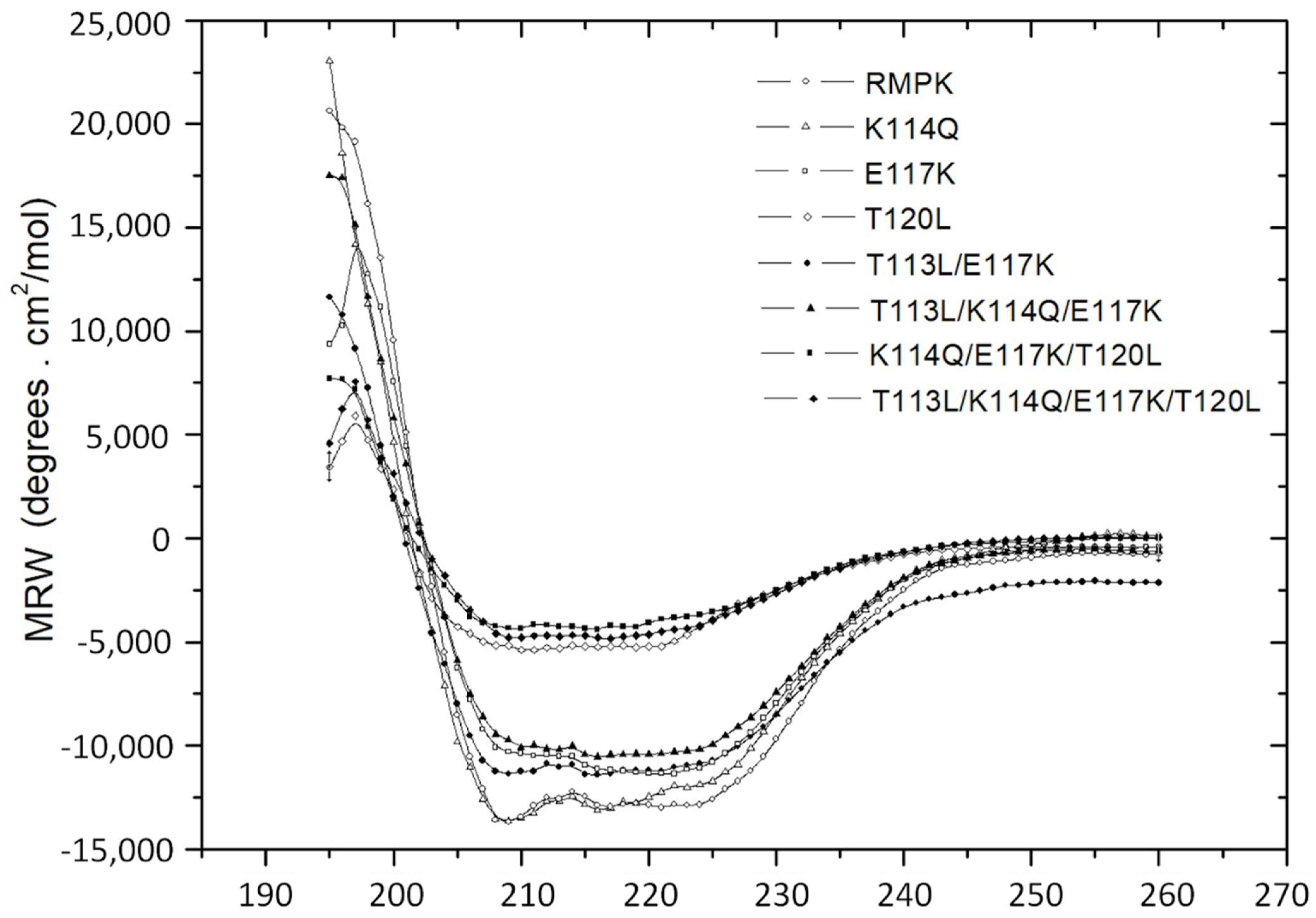
2.4. Circular Dichroism Spectra of WT-RMPK and RMPK Mutants
2.5. Structural Features of the Open and Closed Conformations of WT-RMPK and of MtbPK
2.6. Molecular Dynamics Simulations of WT-RMPK, T120L Mutant and MtbPK in the Absence or Presence of Ligands
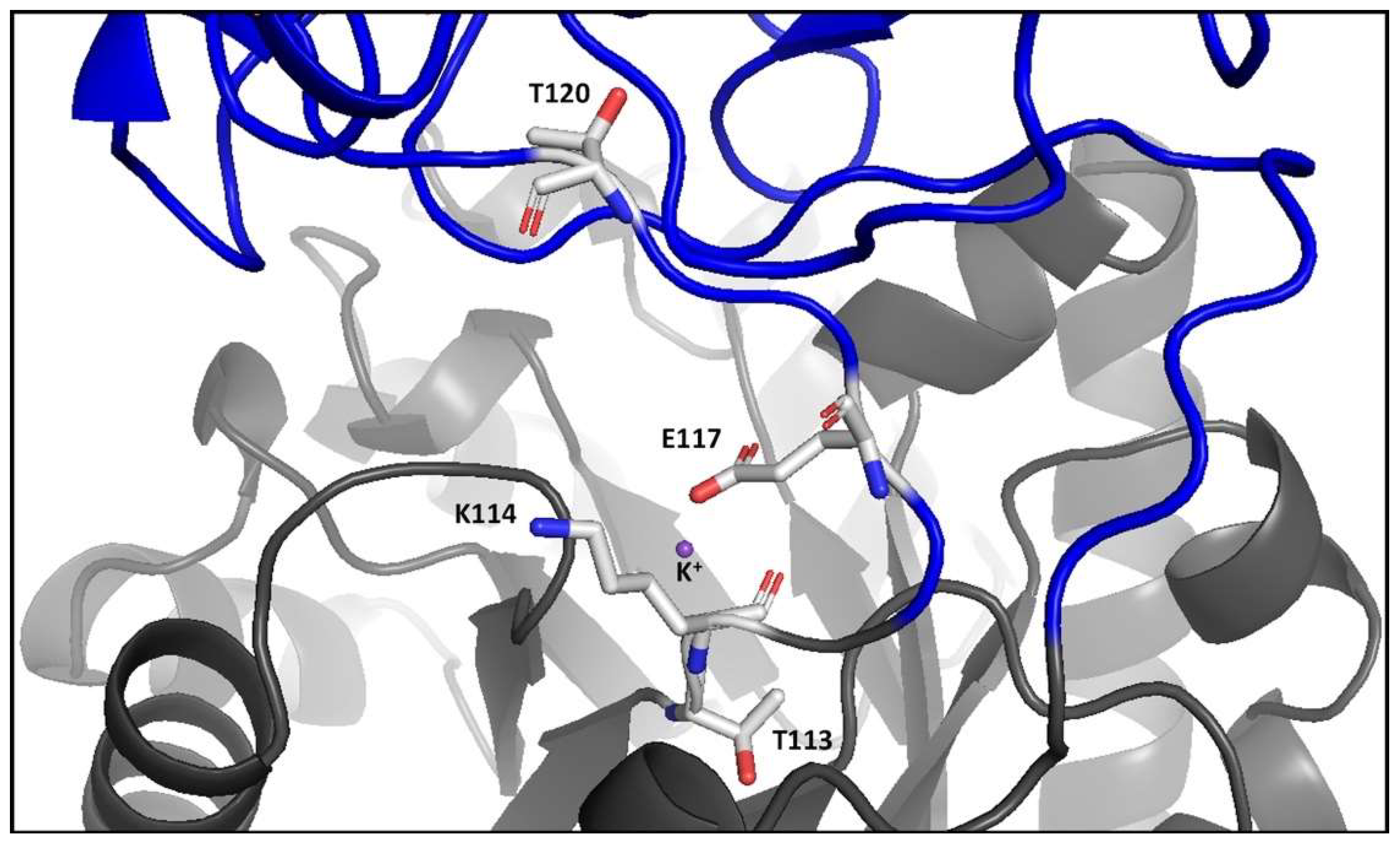
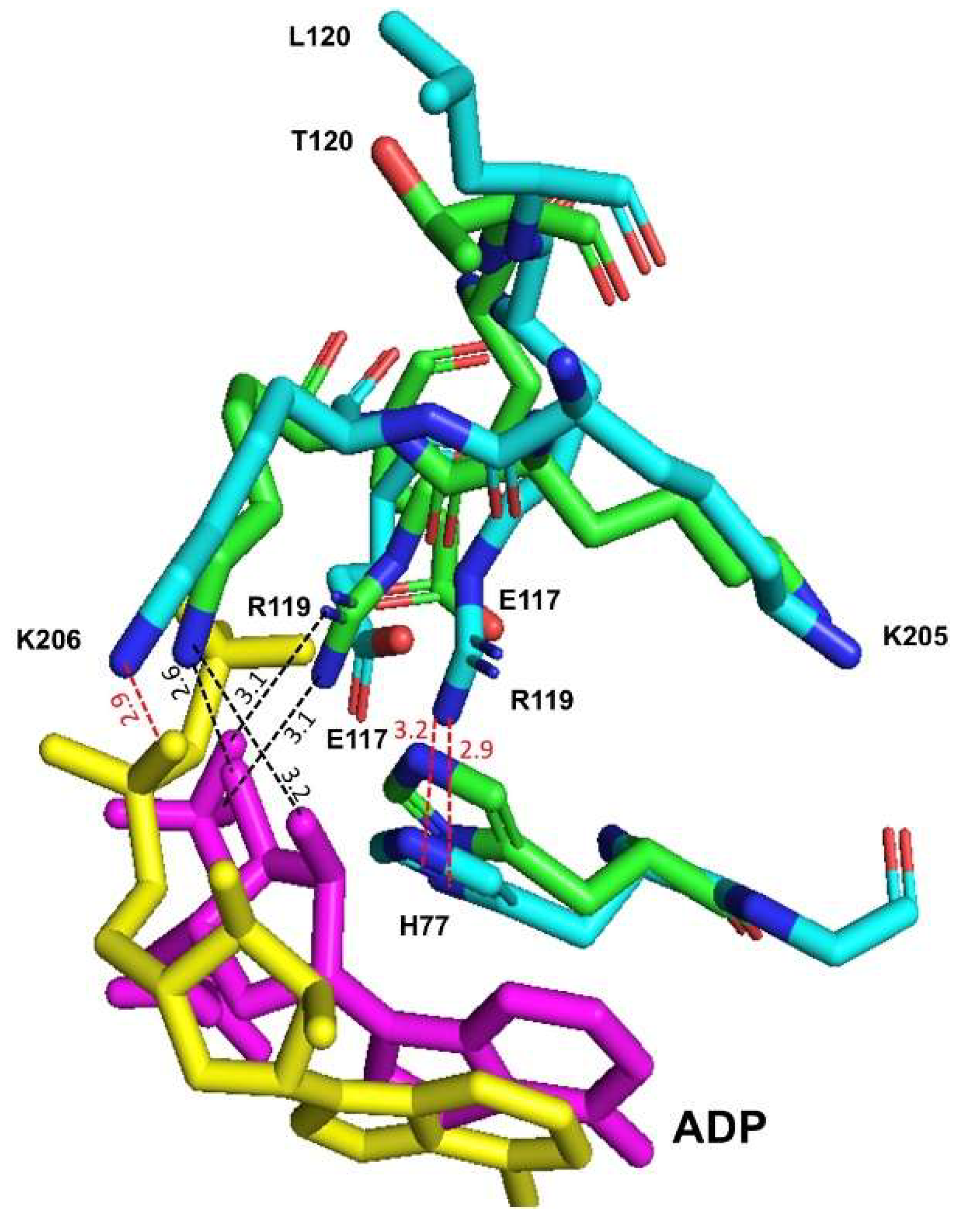
2.7. Structural Changes Induced by the Substitution of Thr for Leu 120 to the RMPK
2.8. Different Main Chain Orientations of Residue Lys117 in the E117K Mutant and the Corresponding Lys72 in the Apo and Holoenzyme of MtbPK
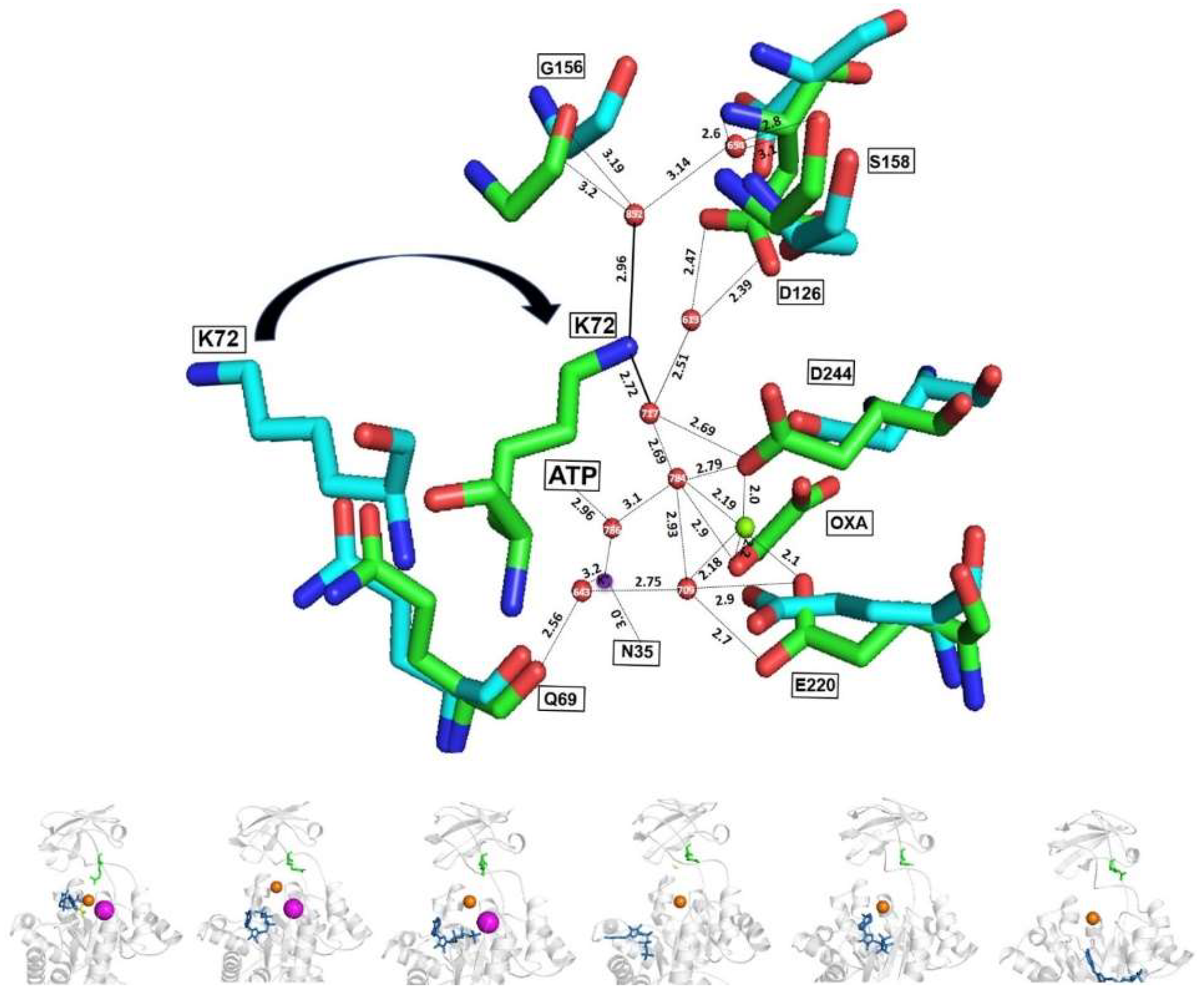
2.9. Different Interaction of MgATP to Oxalate, Mg2+ and K+ in the Active Sites of WT-RMPK and MtbPK
2.10. Active Sites of the Holo and Apo MtbPK
3. Conclusions
4. Experimental Procedures
4.1. Cloning and Expression of RMPK Mutants
4.2. Cell Growth and Purification of RMPK Mutants
4.3. Assays of Pyruvate Kinase Activity
4.4. Circular Dichroism Experiments
4.5. Molecular Dynamics Simulation
4.5.1. Preparation of Initial Coordinate Files
4.5.2. Molecular Dynamics Simulation
4.5.3. Trajectory Analysis
4.6. Crystallization of the Mutant E117K
4.7. Data Collection and Reduction
4.8. Molecular Replacement and Model Refinement
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Pyruvate kinase | PK |
| Wild type rabbit muscle pyruvate kinase | WT-RMPK |
| Choline | (CH3)3N+-CH2-CH2-OH |
| Circular dichroism | CD |
| Molecular dynamics | MD |
| Mycobacterium tuberculosis pyruvate kinase | MtbPK |
| N-(2-Hydroxyethyl)piperazine-N′-(2-ethanesulfonic acid) | HEPES |
| phosphoenolpyruvate | PEP |
| Luria Bertani | LB |
| hog muscle lactate dehydrogenase | LDH |
References
- Kachmar, J.F.; Boyer, P.D. Kinetic analysis of enzyme reactions: II. The potassium activation and calcium inhibition of pyruvic phosphoferase. J. Biol. Chem. 1953, 200, 669–682. [Google Scholar] [CrossRef]
- Ramírez-Silva, L.; Ferreira, S.T.; Nowak, T.; Tuena de Gómez-Puyou, M.; Gómez-Puyou, A. Dimethyl sulfoxide promotes the K+-independent activity of pyruvate kinase and the acquisition of the active catalytic conformation. Eur. J. Biochem. 2001, 268, 3267–3274. [Google Scholar] [CrossRef] [PubMed]
- Kayne, F.J. Thallium (I) activation of pyruvate kinase. Arch. Biochem. Biophys. 1971, 143, 232–239. [Google Scholar] [CrossRef]
- Nowak, T. Conformational changes required for pyruvate kinase activity as modulated by monovalent cations. J. Biol. Chem. 1976, 251, 73–78. [Google Scholar] [CrossRef]
- Benziman, M. Factors affecting the activity of pyruvate kinase of Acetobacter xylinum. Biochem. J. 1969, 112, 631–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, C.L.; Atkinson, D.E. Regulation at the phosphoenolpyruvate branchpoint in Azotobacter vinelandii: Pyruvate kinase. J. Bacteriol. 1971, 106, 37–44. [Google Scholar] [CrossRef] [Green Version]
- Smart, J.B.; Pritchard, G.G. Regulation of pyruvate kinase from Propionibacterium shermanii. Arch. Microbiol. 1979, 122, 281–288. [Google Scholar] [CrossRef]
- Kapoor, R.; Venkitasubramanian, T.A. Glucose 6-phosphate activation of pyruvate kinase from Mycobacterium smegmatis. Biochem. J. 1981, 193, 435–440. [Google Scholar] [CrossRef] [Green Version]
- Waygood, E.B.; Rayman, M.K.; Sanwal, B.D. The control of pyruvate kinase of Escherichia coli. II. Effectors and regulatory properties of the enzyme activated by ribose 5-phosphate. Can. J. Biochem. 1975, 53, 444–454. [Google Scholar] [CrossRef]
- Busto, F.; Del Valle, P.; Soler, J. Some kinetic properties of pyruvate kinase from Phycomyces blakesleeanus. Biochem. Cell Biol. 1988, 66, 148–157. [Google Scholar] [CrossRef]
- Jetten, M.S.M.; Gubler, M.E.; Lee, S.H.; Sinskey, A.J. Structural and functional analysis of pyruvate kinase from Corynebacterium glutamicum. Appl. Environ. Microbiol. 1994, 60, 2501–2507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steiner, P.; Fussenegger, M.; Bailey, J.E.; Sauer, U. Cloning and expression of the Zymomonas mobilis pyruvate kinase gene in Escherichia coli. Gene 1988, 220, 31–38. [Google Scholar] [CrossRef]
- Schramm, A.; Siebers, B.; Tjaden, B.; Brinkmann, H.; Hensel, R. Pyruvate kinase of the hyperthermophilic Crenarchaeote Thermoproteus tenax: Physiological role and phylogenetic aspects. J. Bacteriol. 2000, 182, 2001–2009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnsen, U.; Hansen, T.; Schönheit, P. Comparative analysis of pyruvate kinases from the hyperthermophilic archaea Archaeoglobus fulgidus, Aeropyrum pernix, and Pyrobaculum aerophilum. J. Biol. Chem. 2003, 278, 25417–25427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laughlin, L.T.; Reed, G. The monovalent cation requirement of rabbit muscle pyruvate kinase is eliminated by substitution of lysine for glutamate 117. Arch. Biochem. Biophys. 1997, 348, 262–267. [Google Scholar] [CrossRef]
- Oria-Hernández, J.; Riveros-Rosas, H.; Ramírez-Silva, L. Dichotomic phylogenetic tree of the pyruvate kinase family. K+-dependent and –independent enzymes. J. Biol. Chem. 2006, 281, 30717–30724. [Google Scholar] [CrossRef] [Green Version]
- Larsen, T.M.; Benning, M.M.; Rayment, I.; Reed, G.H. Structure of the bis(Mg2+)-ATP-oxalate complex of the rabbit muscle pyruvate kinase at 2.1 Å resolution: ATP binding over a barrel. Biochemistry 1998, 37, 6247–6255. [Google Scholar] [CrossRef]
- Zhong, W.; Cui, L.; Goh, B.C.; Cai, O.; Ho, P.; Chionh, Y.H.; Yuan, M.; El Sahili, A.; Fothergill-Gilmore, L.; Walkinshaw, M.D.; et al. Allosteric pyruvate kinase-based “logic gate” synergistically senses energy and sugar levels in Mycobacterium tuberculosis. Nat. Commun. 2017, 8, 1986. [Google Scholar] [CrossRef]
- Suelter, C.H. Monovalent cations in enzyme-catalyzed reactions. In Metal Ions in Biological Systems; Sigel, H., Ed.; Dekker: New York, NY, USA, 1974; Volume 3, pp. 201–251. [Google Scholar]
- Oria-Hernández, J.; Cabrera, N.; Pérez-Montfort, R.; Ramírez-Silva, L. Pyruvate kinase revisited. The activating efffect of K+. J. Biol. Chem. 2005, 280, 37924–37929. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Silva, L.; Guerrero-Mendiola, C.; Cabrera, N. The importance of polarity in the evolution of the K+-binding site of pyruvate kinase. Int. J. Mol. Sci. 2014, 15, 22214–22226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Cera, E. A structural perspective on enzymes activated by monovalent cations. J. Biol. Chem. 2006, 281, 1305–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, M.J.; Di Cera, E. Role of Na+ and K+ in enzyme function. Physiol. Rev. 2006, 86, 1049–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerrero-Mendiola, C.; García-Trejo, J.J.; Encalada, R.; Saavedra, E.; Ramírez-Silva, L. The contribution of two isozymes to the pyruvate kinase activity of Vibrio cholera: One K+-dependent constitutively active and another K+-independent with essential allosteric activation. PLoS ONE 2017, 12, e0178673. [Google Scholar] [CrossRef] [PubMed]
- Geenfield, N. Using circular dichroism spectra to estimate protein secondary structure. Nat. Protoc. 2006, 1, 2876–2890. [Google Scholar] [CrossRef] [PubMed]
- Allen, S.C.; Muirhead, H. Refined three-dimensional structure of cat muscle (M1) pyruvate kinase at a resolution of 2.6 Å. Acta Crystallogr. Sect. D Biol. Crystallogr. 1996, 52, 499–504. [Google Scholar] [CrossRef]
- Larsen, T.M.; Laughlin, L.T.; Holden, H.M.; Rayment, I.; Reed, G.H. Structure of Rabbit Muscle Pyruvate Kinase Complexed with Mn2+, K+, and Pyruvate. Biochemistry 1994, 33, 6301–6309. [Google Scholar] [CrossRef]
- Larsen, T.M.; Benning, M.M.; Wesenberg, G.E.; Rayment, I.; Reed, G.H. Ligand-Induced Domain Movement in Pyruvate Kinase: Structure of the Enzyme from Rabbit Muscle with Mg2+, K+, and L-Phospholactate at 2.7 Å Resolution. Arch. Biochem. Biophys. 1997, 354, 199–206. [Google Scholar] [CrossRef]
- Schormann, N.; Hayden, K.L.; Lee, P.; Banerjee, S.; Chattopadhyay, D. An overview of structure, function, and regulation of pyruvate kinases. Protein Sci. 2019, 28, 1771–1784. [Google Scholar] [CrossRef]
- Solomons, T.G.; Johnsen, U.; Schönheit, P.; Davies, C. 3-Phosphoglycerate is an Allosteric Activator of Pyruvate Kinase from the Hyperthermophilic Archaeon Pyrobaculum aerophilum. Biochemistry 2013, 52, 5865–5875. [Google Scholar] [CrossRef] [PubMed]
- Abdelhamid, Y.; Brear, P.; Greenhalgh, J.; Chee, X.; Rahman, T.; Welch, M. Evolutionary plasticity in the allosteric regulator binding site of pyruvate kinase isoform PykA from Pseudomonas aeruginosa. J. Biol. Chem. 2019, 294, 15505–15528. [Google Scholar] [CrossRef]
- Roe, D.R.; Cheatham, T.E., III. PTRAJ and CPPTRAJ: Software for Processing and Analysis of Molecular Dynamics Trajectory Data. J. Chem. Theory Comput. 2013, 9, 3084–3095. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Silva, L.; Tuena de Gómez-Puyou, M.; Gómez-Puyou, A. Water-Induced transitions in the K+ requirements for the activity of Pyruvate Kinase entrapped in Reverse Micelles. Biochemistry 1993, 32, 5332–5338. [Google Scholar] [CrossRef] [PubMed]
- Büchner, T.; Pleiderer, G. Pyruvate kinase from muscle. In Methods in Enzymology; Colowick, S., Kaplan, N., Eds.; Academic Press: New York, NY, USA, 1955; Volume 1, pp. 435–440. [Google Scholar]
- Shoemakers, T.J.M.; Visser, G.J.; Flik, G.; Theuvent, P.R. CHELATOR: An improved method for computing metal ion concentrations in physiological solutions. Biotechniques 1992, 12, 870–879. [Google Scholar]
- Doughtery, T.M.; Cleland, W.W. pH studies of the chemical mechanism of rabbit muscle pyruvate kinase. 2. Physiological substrates and phosphoenol-αbutyrate. Biochemistry 1985, 24, 5875–5880. [Google Scholar]
- Cottam, G.L.; Hollenberg, P.F.; Coon, M.J. Subunit structure of rabbit muscle pyruvate kinase. J. Biol. Chem. 1969, 244, 1481–1486. [Google Scholar] [CrossRef]
- DeLano, W.L. Use of PYMOL as a communications tool for molecular science. Am. Chem. Soc. 2004, 228, U313–U314. [Google Scholar]
- Ferrari, A.J.R.; Gozzo, F.C.; Martinez, L. Statistical force-field for structural modeling using chemical cross-linking/mass spectrometry distance constraints. Bioinformatics 2019, 35, 3005–3012. [Google Scholar] [CrossRef]
- David, A.C.; Josh, B.; Betz, R.M.; Cerutti, D.S.; Cheatham, T.E., III; Darden, T.A.; Duke, R.E.; Giese, T.J.; Gohlke, H.; Goetz, A.W.; et al. AMBER 2015. 2015. Available online: https://ambermd.org/doc12/Amber15.pdf (accessed on 13 January 2022).
- Case, D.A.; Darden, T.A.; Cheatham, T.E., III; Simmerling, C.L.; Wang, J.; Duke, R.E.; Luo, R.; Walker, R.C.; Zhang, W.; Merz, K.M.; et al. AMBER 12. 2012. Available online: https://ambermd.org/doc12/Amber12.pdf (accessed on 13 January 2022).
- Case, D.A.; Cheatham, T.E., III; Darden, T.; Gohlke, H.; Luo, R.; Merz, K.M.; Onufriev, A.; Simmerling, C.; Wang, B.; Woods, R.J. The Amber biomolecular simulation programs. J. Comput. Chem. 2005, 26, 1668–1688. [Google Scholar] [CrossRef] [Green Version]
- Walker, R.C.; Crowley, M.F.; Case, D.A. The implementation of a fast and accurate QM/MM potential method in Amber. J. Comput. Chem. 2008, 29, 1019–1031. [Google Scholar] [CrossRef] [Green Version]
- Tian, C.; Kasavajhala, K.; Belfon, K.A.A.; Raguette, L.; Huang, H.; Migues, A.N.; Bickel, J.; Wang, Y.; Pincay, J.; Wu, Q.; et al. ff19SB: Amino-Acid-Specific Protein Backbone Parameters Trained against Quantum Mechanics Energy Surfaces in Solution. J. Chem. Theory Comput. 2020, 16, 528–552. [Google Scholar] [CrossRef]
- Salomon-Ferrer, R.; Gotz, A.W.; Poole, D.; Le Grand, S.; Walker, R.C. Routine Microsecond Molecular Dynamics Simulations with AMBER on GPUs. 2. Explicit Solvent Particle Mesh Ewald. J. Chem. Theory Comput. 2013, 9, 3878–3888. [Google Scholar] [CrossRef] [PubMed]
- Kabsch, W. Xds. Acta. Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 125–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, P.R. Scaling and assessment of data quality. Acta. Crystallogr. Sect. D Biol. Crystallogr. 2006, 62, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Adams, P.D.; Afonine, P.V.; Bunkóczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.-W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. A comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 213–221. [Google Scholar] [CrossRef] [Green Version]
- McCoy, A.J.; Grosse-Kunstleve, R.W.; Adams, P.D.; Winn, M.D.; Storoni, L.C.; Read, R.J. Phaser crystallographic software. J. Appl. Crystallogr. 2007, 40, 658–674. [Google Scholar] [CrossRef] [Green Version]
- Afonine, P.V.; Grosse-Kunstleve, R.W.; Echols, N.; Headd, J.J.; Moriarty, N.W.; Mustyakimov, M.; Terwilliger, T.C.; Urzhumtsev, A.; Zwart, P.H.; Adams, P.D. Towards automated crystallographic structure refinement with phenix refine. Acta. Crystallogr. D. Biol. Crystallogr. 2012, 68, 352–367. [Google Scholar] [CrossRef] [Green Version]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. Sect. D Biol. Crystallogr. 2004, 60, 2126–2132. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Cooper, D.R.; Porebski, P.J.; Shabalin, I.G.; Handing, K.B.; Minor, W. CheckMyMetal: A macromolecular metal-binding validation tool. Acta Crystallogr. Sect. D Biol. Crystallogr. 2017, 73, 223–233. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| M+.Ionic radius (pm) | WT-RMPK | K114Q | ||||||
|---|---|---|---|---|---|---|---|---|
| kcat (s−1) | K (mM) | n | log E (s−1 M−1) | kcat (s−1) | K (mM) | n | log E (s−1 M−1) | |
| Li+69 | 16 ± 3 | 53 ± 25 | ___ | 0.47 | N.D. | N.D. | N.D. | N.D. |
| Na+102 | 138 ± 8 | 48 ± 4 | 1.7 ± 0.1 | 3.46 | N.D. | N.D. | N.D. | N.D. |
| K+138 | 857 ± 16 | 5.8 ± 0.2 | 1.5 ± 0.1 | 5.17 | 970 ± 24 | 11 ± 0.6 | 1.4 ± 0.1 | 4.95 |
| NH4+147 | 596 ± 24 | 2.5 ± 0.2 | 1.8 ± 0.2 | 5.38 | 742 ± 25 | 3.5 ± 0.2 | 1.6 ± 0.1 | 5.33 |
| Rb+148 | 462 ± 20 | 5.4 ± 0.4 | 1.4 ± 0.2 | 4.93 | 691 ± 24 | 7.0 ± 0.5 | 1.5 ± 0.1 | 4.99 |
| Cs+170 | 198 ± 12 | 20 ± 3 | 1.5 ± 0.1 | 3.99 | 223 ± 13 | 15 ± 1.8 | 1.4 ± 0.2 | 4.17 |
| RMPK & RMPK Mutants | With E117 | RMPK Mutants | With K117 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| kcat (s−1) | K (mM) | n | Log kcat/K (s−1 M−1) | kcat (s−1) | K (mM) | n | Log kcat/K (s−1 M−1) | ||
| PEP | PEP | ||||||||
| WT | 703 ± 28 | 0.063 ± 0.008 | __ | 7.05 | E117K | 249 ± 16 | 0.063 ± 0.00 | __ | 6.60 |
| aT113L | 725 ± 49 | 0.108 ± 0.018 | __ | 6.83 | T113L/E117K | 280 ± 8 | 0.048 ± 0.002 | __ | 6.77 |
| K114Q | 805 ± 26 | 0.15 ± 0.014 | __ | 6.74 | T113L/K114Q/E117K | 363 ± 4 | 0.067 ± 0.00 | 1.82 ± 0.17 | 6.73 |
| MgADP | MgADP | ||||||||
| WT | 691 ± 16 | 0.23 ± 0.010 | __ | 6.48 | E117K | 261 ± 12 | 0.360 ± 0.035 | 1.23 ± 0.09 | 5.86 |
| bT113L | 831 ± 120 | 1.07 ± 0.26 | __ | 5.89 | T113L/E117K | 363 ± 28 | 0.100 ± 0.030 | __ | 6.56 |
| K14Q | 690 ± 19 | 0.54 ± 0.034 | __ | 6.10 | T113L/K114Q/E117K | 419 ± 8 | 0.640 ± 0.030 | 1.82 ± 0.17 | 5.82 |
| Mg2+free | Mg2+free | ||||||||
| WT | 865 ± 12 | 0.17 ± 0.010 | 1.46 ± 0.1 | 6.71 | E117K | 284 ± 4 | 4.9 ± 0.3 | __ | 4.76 |
| K114Q | N.D. | N.D. | N.D. | N.D. | T113L/E117K | 458 ± 75 | 1.6 ± 0.5 | 1.39 ± 0.3 | 5.46 |
| T113L | N.D. | N.D. | N.D. | N.D. | T113L/K114Q/E117K | 371 ± 16 | 8.7 ± 0.7 | 1.44 ± 0.1 | 4.63 |
| Name | Template DNA | Primer Sequence 5′→3′ |
|---|---|---|
| K114Q | WT-RMPK | FW_CTGGACACTCAGGGACCCGAG Rv_CTCGGGTCCCTGAGTGTCCAG |
| T120L | WT-RMPK | Fw_ATCCGGCTAGGCCTCATCAAGGGC Rv_GCCCTTGATGAGGCCTAGCCGGAT |
| T113L/E117K | E117K mutant 1 | Fw_CTGGACCTTAAGGGACCCAAGATCCGGACG Rv_AGTCCGGATCTTGGGTCCCTTAAGGTCCAG |
| T113L/K114Q/E117K | E117K mutant 1 | Fw_CTCTGGACCTGCAGGGACCC Rv_GGGTCCCTGCAGGTCCAGAG |
| K114Q/E117K/T120L | K114Q mutant | Fw_GGACCCAAGATCCGGCTAGGCCTCATCAAGGGC Rv_GCCCTTGATGAGGCCTAGCCGGATCTTGGGTCC |
| T113L/K114K/E117K/ T120L | T113L/K114Q/E117K mutant | |
| External (pTRC99A) | Fw_TAATCATCCGGCTCGTATAATGTG Rv_GGCTGAAAATCTTCTCTCATCCGC | |
| Data Collection | |
| Wavelength [Å] | 0.98 |
| Space group | P212121 |
| Unit cell dimensions a, b, c [Å] | 108.34, 121.83, 161.37 |
| Resolution range [Å] a | 48.18–2.25 (2.37–2.25) |
| Rmerge (%) b | 0.093 (0.57) |
| I/σI | 17.5 (3.0) |
| Observations Total Unique | 733102 (89732) 101836 (14698) |
| Completeness | 99.9 (100.0) |
| Redundancy | 7.2 (6.1) |
| CC1/2 | 0.99 (0.81) |
| Refinement Statistics | |
| Rwork/Rfree (%) c | 0.20/0.25 |
| No. of atoms: | 15009 |
| Protein | 14307 |
| Ligands | 60 |
| Metal Ions | 4 |
| Solvent | 638 |
| B value (Å2) | 34.0 |
| RMSD bond lengths (Å) | 0.006 |
| RMSD bond angles (°) | 0.898 |
| Ramachandran favored (%) | 98.73 |
| Ramachandran allowed (%) | 1.11 |
| Ramachandran outliers (%) | 0.16 |
| PDB code | 7R6Y |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramírez-Silva, L.; Hernández-Alcántara, G.; Guerrero-Mendiola, C.; González-Andrade, M.; Rodríguez-Romero, A.; Rodríguez-Hernández, A.; Lugo-Munguía, A.; Gómez-Coronado, P.A.; Rodríguez-Méndez, C.; Vega-Segura, A. The K+-Dependent and -Independent Pyruvate Kinases Acquire the Active Conformation by Different Mechanisms. Int. J. Mol. Sci. 2022, 23, 1347. https://doi.org/10.3390/ijms23031347
Ramírez-Silva L, Hernández-Alcántara G, Guerrero-Mendiola C, González-Andrade M, Rodríguez-Romero A, Rodríguez-Hernández A, Lugo-Munguía A, Gómez-Coronado PA, Rodríguez-Méndez C, Vega-Segura A. The K+-Dependent and -Independent Pyruvate Kinases Acquire the Active Conformation by Different Mechanisms. International Journal of Molecular Sciences. 2022; 23(3):1347. https://doi.org/10.3390/ijms23031347
Chicago/Turabian StyleRamírez-Silva, Leticia, Gloria Hernández-Alcántara, Carlos Guerrero-Mendiola, Martin González-Andrade, Adela Rodríguez-Romero, Annia Rodríguez-Hernández, Alan Lugo-Munguía, Paul A. Gómez-Coronado, Cristina Rodríguez-Méndez, and Alicia Vega-Segura. 2022. "The K+-Dependent and -Independent Pyruvate Kinases Acquire the Active Conformation by Different Mechanisms" International Journal of Molecular Sciences 23, no. 3: 1347. https://doi.org/10.3390/ijms23031347
APA StyleRamírez-Silva, L., Hernández-Alcántara, G., Guerrero-Mendiola, C., González-Andrade, M., Rodríguez-Romero, A., Rodríguez-Hernández, A., Lugo-Munguía, A., Gómez-Coronado, P. A., Rodríguez-Méndez, C., & Vega-Segura, A. (2022). The K+-Dependent and -Independent Pyruvate Kinases Acquire the Active Conformation by Different Mechanisms. International Journal of Molecular Sciences, 23(3), 1347. https://doi.org/10.3390/ijms23031347