
Therapeutic Treatments for Osteoporosis—Which Combination of Pills Is the Best among the Bad?

 ,
,  ,
,  and
and
Abstract
:1. Bone Remodeling Process in Healthy Individuals
1.1. Bone Development: Differentiation and Activation of Osteoblasts
1.2. Bone Autophagy: Differentiation and Activation of Osteoclasts
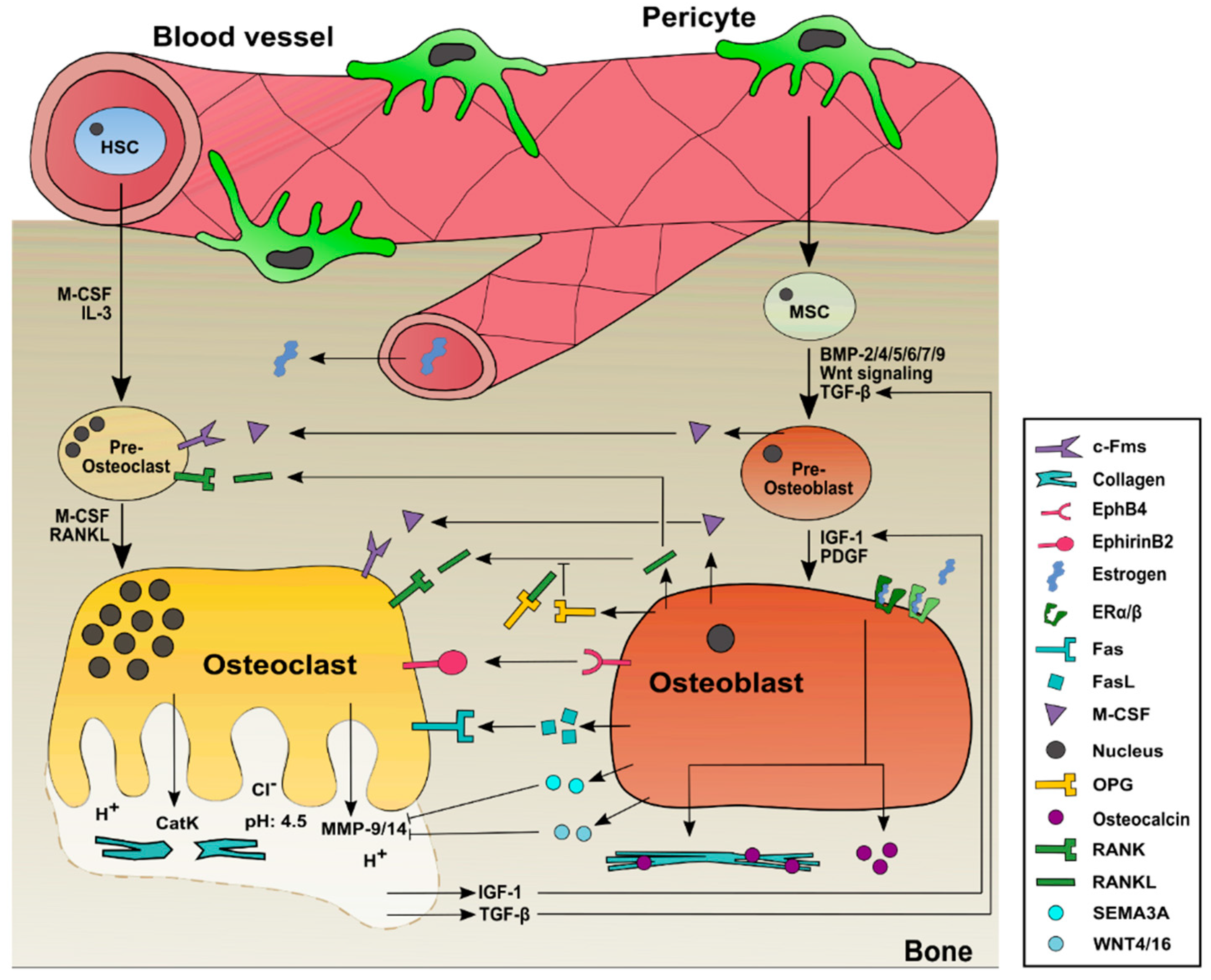
1.3. Interplay of the Major Cell Types Involved in Bone Remodeling: Osteoblasts and Osteoclasts
2. Changes in the Bone-Remodeling Process in Osteoporosis
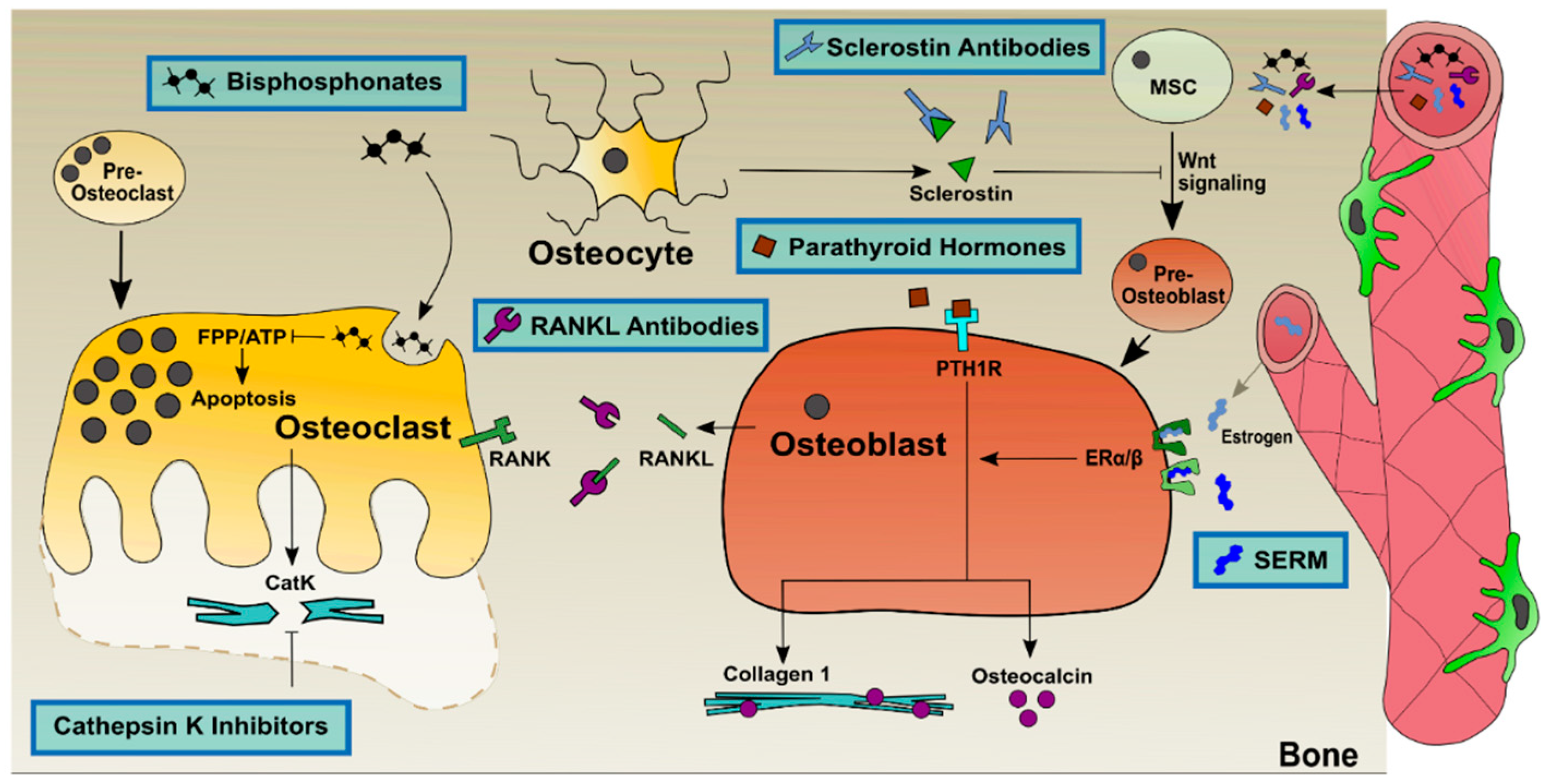
3. State of the Art Treatments against Osteoporosis
3.1. Anabolic Treatments of Osteoblasts to Improve Bone Growth
3.1.1. Parathyroid Hormones and Hormone-Related Bone Growth Agents Increase Osteoblast Activity
3.1.2. Monoclonal Sclerostin Antibodies Promote Wnt Signaling in Osteoblasts
3.2. Catabolic Treatments of Osteoclasts to Prevent Excessive Bone Resorption
3.2.1. Bisphosphonates Advance Osteoclasts toward Apoptosis
3.2.2. Selective Estrogen Receptor Modulators Inhibit the Activation of Osteoclasts
3.2.3. Monoclonal RANKL Antibody Prevents Receptor Activation on Osteoclasts
3.2.4. Cathepsin K Inhibitors Prevent Type I Collagen Degradation
3.3. Combinational Therapies to Improve Osteoporosis Treatment
3.4. Treatment of Vertebral and Critical Size Fractures of Osteoporosis Patients Using Scaffold Material and Controlled Drug Release
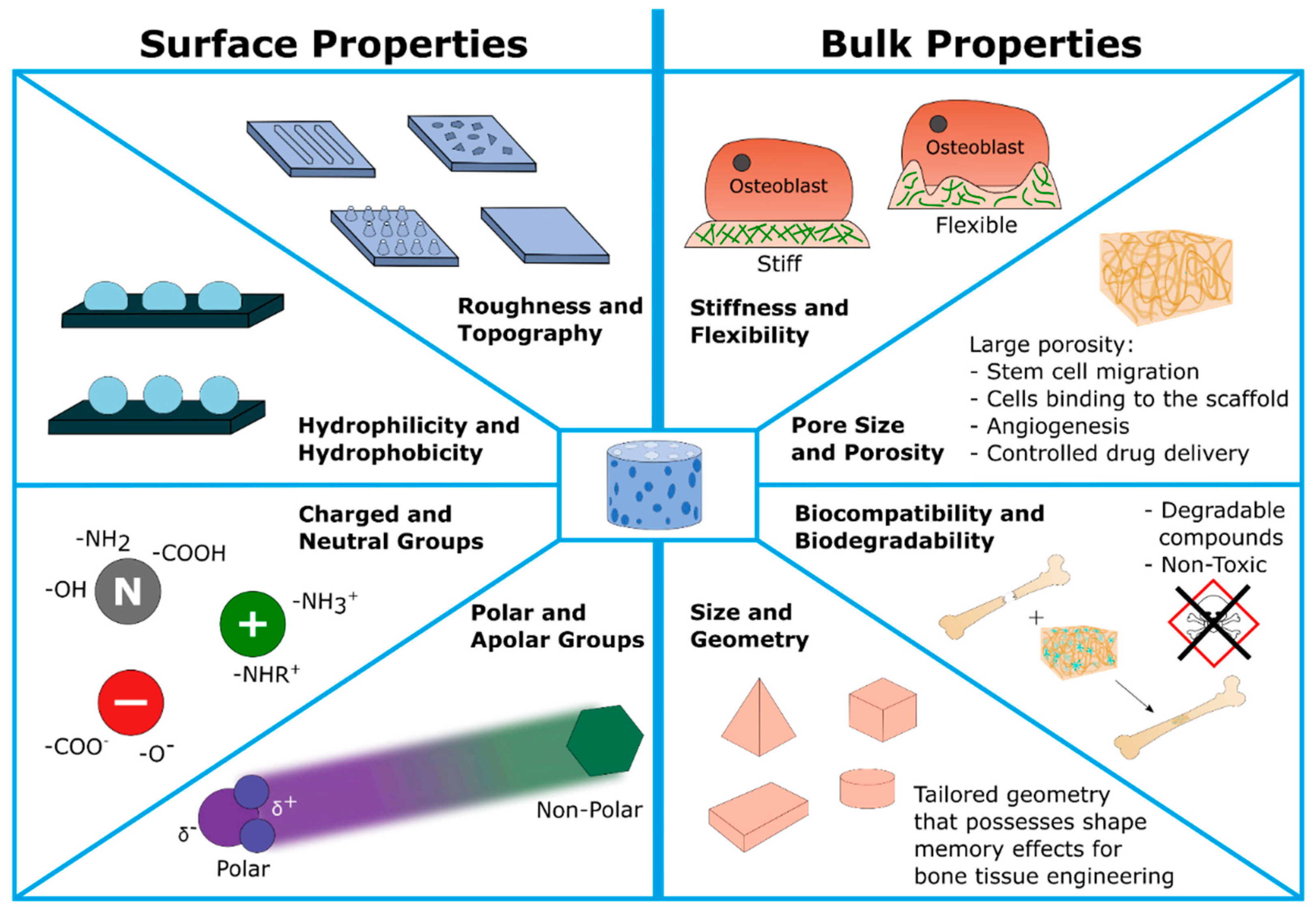
3.4.1. Scaffolds Used in Osteoporosis Treatment
- i.
- Bulk properties: including porosity/pore sizes, mechanical strength (stiffness and flexibility), biocompatibility, and last but not least, the (bio)degradability of scaffold components and metabolic products. In summary, these parameters are confirmed to guide the 3D cell migration, proliferation as well as final osteogenesis. Moreover, scaffolds’ development also should support angiogenesis, e.g., providing a tailored porosity for the ingrowth of cardiovascular tissue. Finally, the scaffold should be biodegradable in order to allow natural bone growth. Therefore, the products of this degradation are required to be non-toxic and non-mutagenic.
- ii.
- Surface properties: including hydrophilicity versus hydrophobicity due to corresponding functional groups (polar or apolar, charged or neutral chemical groups) attached to the surface and resulting in a certain wettability behavior; surface architecture including roughness and topography. For example, scaffold surfaces can be modified to allow a certain cell alignment. Function of the tailored surface include mimicking the surface of the ECM, guiding the cell adhesion, and final cell attachment.
- –
- Chemical and physical vapor deposition (CVD and PVD): used for surface functionalization and modification;
- –
- Self-assembly methods, applied to cover scaffold surfaces with polymer mono- and/or multilayers to modify the surface chemistry and/or topography in a very controlled manner, e.g., layer-by-layer (LbL) method and the Langmuir–Blodgett technique;
- –
- Electrospinning of fiber-based scaffolds that mimic the extracellular matrix. Nano-scaled fibrillary structures with diameters of 50–500 nm are accessible—enhancing the cell–matrix interactions and calcification. Increasing the length of electrospun fibers (via polymer concentration) results in enhanced fiber entanglements, thereby improving the osteoconductive activity;
- –
- Solid freeform rapid prototyping including selective laser sintering (SLS), selective laser ablation (SLA), and fused deposition methods (FDMs) are used for scaffold surface patterning in different micro- and nano-scales. FDMs allow easy and flexible material implementation for scaffold fabrication reported for bone tissue engineering [228,231]; 3D printing and 3D plotting combines rapid prototyping technologies to produce tailored 3D scaffolds. Thus, scaffold size, shape, and porosity can be adjusted. Interconnected pores are accessible supporting cell ingrowth, metabolic activity, and nutrient exchange [232,233,234,235];
- –
- Lithographic methods can be used, and finally;
- –
3.4.2. Controlled Drug Release Used in Osteoporosis Treatment
- Teriparatide therapy should generally be limited to 24 months;
- Raloxifene therapy has been shown to be beneficial for up to eight years;
- Bisphosphonate therapy has proven benefits for three to five years;
- Denosumab has a proven benefit for up to three years.
4. Future Perspectives of Anabolic and Catabolic Treatments for Osteoporosis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ABL | Abaloparatide |
| ALN | Alendronate |
| ALP | Alkaline phosphatase |
| ATF4 | Activating transcription factor 4 |
| ATP | Adenosine triphosphate |
| BM-MSC | Bone marrow mesenchymal stem cell |
| BMD | Bone mineral density |
| BMP | Bone morphogenetic protein |
| BMU | Basic multicellular unit |
| BP | Bisphosphonate |
| BSP | Bone sialoprotein |
| COL1A1 | Alpha-1 type I collagen |
| COVID-19 | Coronavirus disease 2019 |
| CTX-1 | C-terminal cross-linked telopeptide of type I collagen |
| DXA/DEXA | Dual-energy X-ray absorptiometry |
| ECM | Extracellular matrix |
| EMA | European Medicines Agency |
| EphB4 | EPH receptor B4 |
| ERα/β | Estrogen receptor-α/β |
| FASL | FAS ligand |
| FDA | Food and Drug Administration |
| FDM | Fused deposition method |
| FPP | Farnesyl pyrophosphatase |
| GM-CSF | Granulocyte-macrophage colony-stimulating factor |
| HA | Hydroxyapatite |
| HDAC | Histone deacetylase |
| HSC | Hematopoietic stem cell |
| IGF | Insulin-like growth factor |
| IGFBP | IGF binding protein |
| IL | Interleukin |
| LbL | Layer-by-layer |
| LRP | LDL-receptor-related protein |
| M-CSF | Macrophage colony-stimulating factor |
| MMP | Matrix metalloproteinase |
| MSC | Mesenchymal stem cell |
| NTX-1 | N-terminal cross-linked telopeptide of type I collagen |
| OB | Osteoblast |
| OC | Osteoclast |
| OCN | Osteocalcin |
| ON | Osteonectin |
| OP | Osteoporosis |
| OPG | Osteoprotegerin |
| OPN | Osteopontin |
| OSX | Osterix |
| OVX | Ovariectomy |
| PCL | Polycaprolactone |
| PGA | Polygluycolicacid |
| PLA | Polylacticacid |
| PLR | Perilacunar remodeling |
| PTH | Parathyroid hormone |
| PTH1/2R | Parathyroid hormone 1/2 receptor |
| PTHrP | Parathyroid hormone-related protein |
| RA | Retinoic acid |
| RANK | Receptor activator of nuclear factor kappa-Β |
| RANKL | Receptor activator of nuclear factor kappa-Β ligand |
| RUNX | Runt-related transcription factor |
| SERM | Selective estrogen receptor modulator |
| SIK | Salt-inducible kinase |
| SIM | Simvastatin |
| SLA | Selective laser ablation |
| SLS | Selective laser sintering |
| TCP | Tricalcium phosphates |
| TGF-β | Transforming growth factor-β |
| TPTD | Teriparatide |
| TRAP | Tartrate-resistant acid phosphatase |
| ZOL | Zoledronic acid/zoledronate |
References
- Clarke, B. Normal bone anatomy and physiology. Clin. J. Am. Soc. Nephrol. 2008, 3 (Suppl. 3), S131–S139. [Google Scholar] [CrossRef] [Green Version]
- Burr, D.B. Bone morphology and organization. In Basic and Applied Bone Biology; Elsevier: Cambridge, MA, USA, 2019; pp. 3–26. [Google Scholar]
- Le, B.Q.; Nurcombe, V.; Cool, S.M.; van Blitterswijk, C.A.; de Boer, J.; LaPointe, V.L.S. The Components of Bone and What They Can Teach Us about Regeneration. Materials 2017, 11, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, M.R.; Hock, J.M.; Burr, D.B. Periosteum: Biology, regulation, and response to osteoporosis therapies. Bone 2004, 35, 1003–1012. [Google Scholar] [CrossRef] [Green Version]
- Florencio-Silva, R.; Sasso, G.R.; Sasso-Cerri, E.; Simoes, M.J.; Cerri, P.S. Biology of Bone Tissue: Structure, Function, and Factors That Influence Bone Cells. BioMed Res. Int. 2015, 2015, 421746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nahian, A.; Chauhan, P.R. Histology, Periosteum and Endosteum. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Schepers, K.; Pietras, E.M.; Reynaud, D.; Flach, J.; Binnewies, M.; Garg, T.; Wagers, A.J.; Hsiao, E.C.; Passegue, E. Myeloproliferative neoplasia remodels the endosteal bone marrow niche into a self-reinforcing leukemic niche. Cell Stem Cell 2013, 13, 285–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charbord, P. Bone marrow mesenchymal stem cells: Historical overview and concepts. Hum. Gene Ther. 2010, 21, 1045–1056. [Google Scholar] [CrossRef] [Green Version]
- Morrison, S.J.; Scadden, D.T. The bone marrow niche for haematopoietic stem cells. Nature 2014, 505, 327–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marks, S.C., Jr.; Gartland, A.; Odgren, P.R. Skeletal Development; Elsevier: San Diego, CA, USA, 2004. [Google Scholar]
- Mackie, E.J.; Ahmed, Y.A.; Tatarczuch, L.; Chen, K.S.; Mirams, M. Endochondral ossification: How cartilage is converted into bone in the developing skeleton. Int. J. Biochem. Cell Biol. 2008, 40, 46–62. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.R.; Burr, D.B. Bone Growth, Modeling, and Remodeling. In Basic and Applied Bone Biology; Elsevier: Cambridge, MA, USA, 2019; pp. 85–100. [Google Scholar]
- Tobiasch, E. Adult human mesenchymal stem cells as source for future tissue engineering. In Forschungsspitzen und Spitzenforschung; Springer: Heidelberg, Germany, 2009; pp. 329–338. [Google Scholar]
- Tobiasch, E. Differentiation potential of adult human mesenchymal stem cells. In Stem Cell Engineering; Springer: Heidelberg, Germany, 2011; pp. 61–77. [Google Scholar]
- Seifert, A.; Werheid, D.F.; Knapp, S.M.; Tobiasch, E. Role of Hox genes in stem cell differentiation. World J. Stem Cells 2015, 7, 583–595. [Google Scholar] [CrossRef] [PubMed]
- Bellido, T.; Plotkin, L.I.; Bruzzaniti, A. Bone cells. In Basic and Applied Bone Biology; Elsevier: Cambridge, MA, USA, 2019; pp. 37–55. [Google Scholar]
- Tonk, C.H.; Witzler, M.; Schulze, M.; Tobiasch, E. Mesenchymal Stem Cells. In Essential Current Concepts in Stem Cell Biology; Springer: Basel, Switzerland, 2020; pp. 21–39. [Google Scholar]
- Siclari, V.A.; Zhu, J.; Akiyama, K.; Liu, F.; Zhang, X.; Chandra, A.; Nah, H.D.; Shi, S.; Qin, L. Mesenchymal progenitors residing close to the bone surface are functionally distinct from those in the central bone marrow. Bone 2013, 53, 575–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debnath, S.; Yallowitz, A.R.; McCormick, J.; Lalani, S.; Zhang, T.; Xu, R.; Li, N.; Liu, Y.; Yang, Y.S.; Eiseman, M.; et al. Discovery of a periosteal stem cell mediating intramembranous bone formation. Nature 2018, 562, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Su, P.; Tian, Y.; Yang, C.; Ma, X.; Wang, X.; Pei, J.; Qian, A. Mesenchymal Stem Cell Migration during Bone Formation and Bone Diseases Therapy. Int. J. Mol. Sci. 2018, 19, 2343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, W.; Gao, B.; Fan, J.; Cheng, P.; Hu, Y.; Jie, Q.; Luo, Z.; Yang, L. Mesenchymal Progenitors Derived from Different Locations in Long Bones Display Diverse Characteristics. Stem Cells Int. 2019, 2019, 5037578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yianni, V.; Sharpe, P.T. Perivascular-Derived Mesenchymal Stem Cells. J. Dent. Res. 2019, 98, 1066–1072. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Khan, D.; Delling, J.; Tobiasch, E. Mechanisms underlying the osteo- and adipo-differentiation of human mesenchymal stem cells. Sci. World J. 2012, 2012, 793823. [Google Scholar] [CrossRef] [Green Version]
- Miron, R.J.; Zhang, Y.F. Osteoinduction: A review of old concepts with new standards. J. Dent. Res. 2012, 91, 736–744. [Google Scholar] [CrossRef]
- Amarasekara, D.S.; Kim, S.; Rho, J. Regulation of Osteoblast Differentiation by Cytokine Networks. Int. J. Mol. Sci. 2021, 22, 2851. [Google Scholar] [CrossRef]
- Pansky, A.; Roitzheim, B.; Tobiasch, E. Differentiation potential of adult human mesenchymal stem cells. Clin. Lab. 2007, 53, 81–84. [Google Scholar]
- Zippel, N.; Limbach, C.A.; Ratajski, N.; Urban, C.; Luparello, C.; Pansky, A.; Kassack, M.U.; Tobiasch, E. Purinergic receptors influence the differentiation of human mesenchymal stem cells. Stem Cells Dev. 2012, 21, 884–900. [Google Scholar] [CrossRef]
- Chen, Q.; Shou, P.; Zheng, C.; Jiang, M.; Cao, G.; Yang, Q.; Cao, J.; Xie, N.; Velletri, T.; Zhang, X.; et al. Fate decision of mesenchymal stem cells: Adipocytes or osteoblasts? Cell Death Differ. 2016, 23, 1128–1139. [Google Scholar] [CrossRef] [Green Version]
- Ducy, P.; Zhang, R.; Geoffroy, V.; Ridall, A.L.; Karsenty, G. Osf2/Cbfa1: A transcriptional activator of osteoblast differentiation. Cell 1997, 89, 747–754. [Google Scholar] [CrossRef] [Green Version]
- Komori, T. Regulation of Proliferation, Differentiation and Functions of Osteoblasts by Runx2. Int. J. Mol. Sci. 2019, 20, 1694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maruyama, Z.; Yoshida, C.A.; Furuichi, T.; Amizuka, N.; Ito, M.; Fukuyama, R.; Miyazaki, T.; Kitaura, H.; Nakamura, K.; Fujita, T.; et al. Runx2 determines bone maturity and turnover rate in postnatal bone development and is involved in bone loss in estrogen deficiency. Dev. Dyn. 2007, 236, 1876–1890. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Jiang, Q.; Miyazaki, T.; Komori, T. Runx2 regulates cranial suture closure by inducing hedgehog, Fgf, Wnt and Pthlh signaling pathway gene expressions in suture mesenchymal cells. Hum. Mol. Genet. 2019, 28, 896–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakashima, K.; Zhou, X.; Kunkel, G.; Zhang, Z.; Deng, J.M.; Behringer, R.R.; de Crombrugghe, B. The novel zinc finger-containing transcription factor osterix is required for osteoblast differentiation and bone formation. Cell 2002, 108, 17–29. [Google Scholar] [CrossRef] [Green Version]
- Komori, T. Regulation of osteoblast differentiation by transcription factors. J. Cell. Biochem. 2006, 99, 1233–1239. [Google Scholar] [CrossRef]
- Luo, Y.; Zhang, Y.; Miao, G.; Zhang, Y.; Liu, Y.; Huang, Y. Runx1 regulates osteogenic differentiation of BMSCs by inhibiting adipogenesis through Wnt/beta-catenin pathway. Arch. Oral Biol. 2019, 97, 176–184. [Google Scholar] [CrossRef]
- Tang, C.Y.; Chen, W.; Luo, Y.; Wu, J.; Zhang, Y.; McVicar, A.; McConnell, M.; Liu, Y.; Zhou, H.D.; Li, Y.P. Runx1 up-regulates chondrocyte to osteoblast lineage commitment and promotes bone formation by enhancing both chondrogenesis and osteogenesis. Biochem. J. 2020, 477, 2421–2438. [Google Scholar] [CrossRef]
- Tang, J.; Xie, J.; Chen, W.; Tang, C.; Wu, J.; Wang, Y.; Zhou, X.D.; Zhou, H.D.; Li, Y.P. Runt-related transcription factor 1 is required for murine osteoblast differentiation and bone formation. J. Biol. Chem. 2020, 295, 11669–11681. [Google Scholar] [CrossRef]
- Tang, C.Y.; Wu, M.; Zhao, D.; Edwards, D.; McVicar, A.; Luo, Y.; Zhu, G.; Wang, Y.; Zhou, H.D.; Chen, W.; et al. Runx1 is a central regulator of osteogenesis for bone homeostasis by orchestrating BMP and WNT signaling pathways. PLoS Genet. 2021, 17, e1009233. [Google Scholar] [CrossRef]
- Xiao, G.; Jiang, D.; Ge, C.; Zhao, Z.; Lai, Y.; Boules, H.; Phimphilai, M.; Yang, X.; Karsenty, G.; Franceschi, R.T. Cooperative interactions between activating transcription factor 4 and Runx2/Cbfa1 stimulate osteoblast-specific osteocalcin gene expression. J. Biol. Chem. 2005, 280, 30689–30696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heo, J.S.; Lee, S.G.; Kim, H.O. Distal-less homeobox 5 is a master regulator of the osteogenesis of human mesenchymal stem cells. Int. J. Mol. Med. 2017, 40, 1486–1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hojo, H.; Ohba, S.; Chung, U.I. Signaling pathways regulating the specification and differentiation of the osteoblast lineage. Regen. Ther. 2015, 1, 57–62. [Google Scholar] [CrossRef] [Green Version]
- Houschyar, K.S.; Tapking, C.; Borrelli, M.R.; Popp, D.; Duscher, D.; Maan, Z.N.; Chelliah, M.P.; Li, J.; Harati, K.; Wallner, C.; et al. Wnt Pathway in Bone Repair and Regeneration—What Do We Know So Far. Front. Cell Dev. Biol. 2018, 6, 170. [Google Scholar] [CrossRef] [PubMed]
- Baron, R.; Kneissel, M. WNT signaling in bone homeostasis and disease: From human mutations to treatments. Nat. Med. 2013, 19, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Chen, G.; Li, Y.P. TGF-beta and BMP signaling in osteoblast, skeletal development, and bone formation, homeostasis and disease. Bone Res. 2016, 4, 16009. [Google Scholar] [CrossRef] [PubMed]
- Krishnakumar, G.S.; Roffi, A.; Reale, D.; Kon, E.; Filardo, G. Clinical application of bone morphogenetic proteins for bone healing: A systematic review. Int. Orthop. 2017, 41, 1073–1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaebisch, C.; Schipper, D.; Babczyk, P.; Tobiasch, E. The role of purinergic receptors in stem cell differentiation. Comput. Struct. Biotechnol. J. 2015, 13, 75–84. [Google Scholar] [CrossRef] [Green Version]
- Blair, H.C. How the osteoclast degrades bone. Bioessays 1998, 20, 837–846. [Google Scholar] [CrossRef]
- Bar-Shavit, Z. The osteoclast: A multinucleated, hematopoietic-origin, bone-resorbing osteoimmune cell. J. Cell. Biochem. 2007, 102, 1130–1139. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, N. Signaling Pathways in Osteoclast Differentiation. Chonnam Med. J. 2016, 52, 12–17. [Google Scholar] [CrossRef] [Green Version]
- Udagawa, N.; Koide, M.; Nakamura, M.; Nakamichi, Y.; Yamashita, T.; Uehara, S.; Kobayashi, Y.; Furuya, Y.; Yasuda, H.; Fukuda, C.; et al. Osteoclast differentiation by RANKL and OPG signaling pathways. J. Bone Miner. Metab. 2021, 39, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Longo, A.; Librizzi, M.; Naselli, F.; Caradonna, F.; Tobiasch, E.; Luparello, C. PTHrP in differentiating human mesenchymal stem cells: Transcript isoform expression, promoter methylation, and protein accumulation. Biochimie 2013, 95, 1888–1896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Udagawa, N.; Takahashi, N.; Yasuda, H.; Mizuno, A.; Itoh, K.; Ueno, Y.; Shinki, T.; Gillespie, M.T.; Martin, T.J.; Higashio, K.; et al. Osteoprotegerin produced by osteoblasts is an important regulator in osteoclast development and function. Endocrinology 2000, 141, 3478–3484. [Google Scholar] [CrossRef]
- Teitelbaum, S.L. Bone resorption by osteoclasts. Science 2000, 289, 1504–1508. [Google Scholar] [CrossRef] [PubMed]
- Vaananen, K. Mechanism of osteoclast mediated bone resorption—Rationale for the design of new therapeutics. Adv. Drug Deliv. Rev. 2005, 57, 959–971. [Google Scholar] [CrossRef] [PubMed]
- Gavali, S.; Gupta, M.K.; Daswani, B.; Wani, M.R.; Sirdeshmukh, R.; Khatkhatay, M.I. Estrogen enhances human osteoblast survival and function via promotion of autophagy. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 1498–1507. [Google Scholar] [CrossRef]
- Khosla, S.; Oursler, M.J.; Monroe, D.G. Estrogen and the skeleton. Trends Endocrinol. Metab. 2012, 23, 576–581. [Google Scholar] [CrossRef] [Green Version]
- Kular, J.; Tickner, J.; Chim, S.M.; Xu, J. An overview of the regulation of bone remodelling at the cellular level. Clin. Biochem. 2012, 45, 863–873. [Google Scholar] [CrossRef]
- Kim, J.M.; Lin, C.; Stavre, Z.; Greenblatt, M.B.; Shim, J.H. Osteoblast-Osteoclast Communication and Bone Homeostasis. Cells 2020, 9, 2073. [Google Scholar] [CrossRef]
- Krum, S.A.; Miranda-Carboni, G.A.; Hauschka, P.V.; Carroll, J.S.; Lane, T.F.; Freedman, L.P.; Brown, M. Estrogen protects bone by inducing Fas ligand in osteoblasts to regulate osteoclast survival. EMBO J. 2008, 27, 535–545. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Liu, S.; Zhao, Y.; Liu, D.; Liu, Y.; Chen, C.; Karray, S.; Shi, S.; Jin, Y. Osteoblast-induced osteoclast apoptosis by fas ligand/FAS pathway is required for maintenance of bone mass. Cell Death Differ. 2015, 22, 1654–1664. [Google Scholar] [CrossRef] [PubMed]
- Garcia, A.J.; Tom, C.; Guemes, M.; Polanco, G.; Mayorga, M.E.; Wend, K.; Miranda-Carboni, G.A.; Krum, S.A. ERalpha signaling regulates MMP3 expression to induce FasL cleavage and osteoclast apoptosis. J. Bone Miner. Res. 2013, 28, 283–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Wang, Z.; Duan, N.; Zhu, G.; Schwarz, E.M.; Xie, C. Osteoblast-osteoclast interactions. Connect. Tissue Res. 2018, 59, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Sambandam, Y.; Blanchard, J.J.; Daughtridge, G.; Kolb, R.J.; Shanmugarajan, S.; Pandruvada, S.N.; Bateman, T.A.; Reddy, S.V. Microarray profile of gene expression during osteoclast differentiation in modelled microgravity. J. Cell. Biochem. 2010, 111, 1179–1187. [Google Scholar] [CrossRef]
- Boyce, B.F.; Xing, L. Functions of RANKL/RANK/OPG in bone modeling and remodeling. Arch. Biochem. Biophys. 2008, 473, 139–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuoka, K.; Park, K.A.; Ito, M.; Ikeda, K.; Takeshita, S. Osteoclast-derived complement component 3a stimulates osteoblast differentiation. J. Bone Miner. Res. 2014, 29, 1522–1530. [Google Scholar] [CrossRef]
- Prideaux, M.; Findlay, D.M.; Atkins, G.J. Osteocytes: The master cells in bone remodelling. Curr. Opin. Pharmacol. 2016, 28, 24–30. [Google Scholar] [CrossRef]
- Frost, H.M. The mechanostat: A proposed pathogenic mechanism of osteoporoses and the bone mass effects of mechanical and nonmechanical agents. Bone Miner. 1987, 2, 73–85. [Google Scholar]
- Nakashima, T.; Hayashi, M.; Fukunaga, T.; Kurata, K.; Oh-Hora, M.; Feng, J.Q.; Bonewald, L.F.; Kodama, T.; Wutz, A.; Wagner, E.F.; et al. Evidence for osteocyte regulation of bone homeostasis through RANKL expression. Nat. Med. 2011, 17, 1231–1234. [Google Scholar] [CrossRef]
- Xiong, J.; Onal, M.; Jilka, R.L.; Weinstein, R.S.; Manolagas, S.C.; O’Brien, C.A. Matrix-embedded cells control osteoclast formation. Nat. Med. 2011, 17, 1235–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dallas, S.L.; Prideaux, M.; Bonewald, L.F. The osteocyte: An endocrine cell... and more. Endocr. Rev. 2013, 34, 658–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atkins, G.J.; Rowe, P.S.; Lim, H.P.; Welldon, K.J.; Ormsby, R.; Wijenayaka, A.R.; Zelenchuk, L.; Evdokiou, A.; Findlay, D.M. Sclerostin is a locally acting regulator of late-osteoblast/preosteocyte differentiation and regulates mineralization through a MEPE-ASARM-dependent mechanism. J. Bone Miner. Res. 2011, 26, 1425–1436. [Google Scholar] [CrossRef] [Green Version]
- Kogawa, M.; Wijenayaka, A.R.; Ormsby, R.T.; Thomas, G.P.; Anderson, P.H.; Bonewald, L.F.; Findlay, D.M.; Atkins, G.J. Sclerostin regulates release of bone mineral by osteocytes by induction of carbonic anhydrase 2. J. Bone Miner. Res. 2013, 28, 2436–2448. [Google Scholar] [CrossRef] [PubMed]
- McClung, M.R.; Grauer, A.; Boonen, S.; Bolognese, M.A.; Brown, J.P.; Diez-Perez, A.; Langdahl, B.L.; Reginster, J.Y.; Zanchetta, J.R.; Wasserman, S.M.; et al. Romosozumab in postmenopausal women with low bone mineral density. N. Engl. J. Med. 2014, 370, 412–420. [Google Scholar] [CrossRef] [Green Version]
- Belanger, L.F.; Belanger, C.; Semba, T. Technical approaches leading to the concept of osteocytic osteolysis. Clin. Orthop. Relat. Res. 1967, 54, 187–196. [Google Scholar]
- Qing, H.; Ardeshirpour, L.; Pajevic, P.D.; Dusevich, V.; Jahn, K.; Kato, S.; Wysolmerski, J.; Bonewald, L.F. Demonstration of osteocytic perilacunar/canalicular remodeling in mice during lactation. J. Bone Miner. Res. 2012, 27, 1018–1029. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.S.; Wang, L.; de Bakker, C.M.J.; Lai, X. Mechanical Regulation of the Maternal Skeleton during Reproduction and Lactation. Curr. Osteoporos. Rep. 2019, 17, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Paul Tuck, S.; Layfield, R.; Walker, J.; Mekkayil, B.; Francis, R. Adult Paget’s disease of bone: A review. Rheumatology 2017, 56, 2050–2059. [Google Scholar] [CrossRef] [Green Version]
- Drake, M.T.; Clarke, B.L.; Lewiecki, E.M. The Pathophysiology and Treatment of Osteoporosis. Clin. Ther. 2015, 37, 1837–1850. [Google Scholar] [CrossRef]
- Kanis, J.A.; Cooper, C.; Rizzoli, R.; Reginster, J.Y.; Scientific Advisory Board of the European Society for Clinical; Economic Aspects of Osteoporosis; the Committees of Scientific Advisors; National Societies of the International Osteoporosis Foundation. European guidance for the diagnosis and management of osteoporosis in postmenopausal women. Osteoporos. Int. 2019, 30, 3–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alswat, K.A. Gender Disparities in Osteoporosis. J. Clin. Med. Res. 2017, 9, 382–387. [Google Scholar] [CrossRef] [Green Version]
- Manolagas, S.C. From estrogen-centric to aging and oxidative stress: A revised perspective of the pathogenesis of osteoporosis. Endocr. Rev. 2010, 31, 266–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albright, F.; Sulkowitch, H.; Bloomberg, E.; Smith, P.H. Post-Menopausal Osteoporosis. Trans. A Am. Physicians 1940, 55, 298–305. [Google Scholar]
- Raisz, L.G. Pathogenesis of osteoporosis: Concepts, conflicts, and prospects. J. Clin. Investig. 2005, 115, 3318–3325. [Google Scholar] [CrossRef] [Green Version]
- Khosla, S.; Atkinson, E.J.; Melton, L.J., 3rd; Riggs, B.L. Effects of age and estrogen status on serum parathyroid hormone levels and biochemical markers of bone turnover in women: A population-based study. J. Clin. Endocrinol. Metab. 1997, 82, 1522–1527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sims, N.A.; Clement-Lacroix, P.; Minet, D.; Fraslon-Vanhulle, C.; Gaillard-Kelly, M.; Resche-Rigon, M.; Baron, R. A functional androgen receptor is not sufficient to allow estradiol to protect bone after gonadectomy in estradiol receptor-deficient mice. J. Clin. Investig. 2003, 111, 1319–1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maatta, J.A.; Buki, K.G.; Gu, G.; Alanne, M.H.; Vaaraniemi, J.; Liljenback, H.; Poutanen, M.; Harkonen, P.; Vaananen, K. Inactivation of estrogen receptor alpha in bone-forming cells induces bone loss in female mice. FASEB J. 2013, 27, 478–488. [Google Scholar] [CrossRef]
- Melville, K.M.; Kelly, N.H.; Khan, S.A.; Schimenti, J.C.; Ross, F.P.; Main, R.P.; van der Meulen, M.C. Female mice lacking estrogen receptor-alpha in osteoblasts have compromised bone mass and strength. J. Bone Miner. Res. 2014, 29, 370–379. [Google Scholar] [CrossRef]
- Kondoh, S.; Inoue, K.; Igarashi, K.; Sugizaki, H.; Shirode-Fukuda, Y.; Inoue, E.; Yu, T.; Takeuchi, J.K.; Kanno, J.; Bonewald, L.F.; et al. Estrogen receptor alpha in osteocytes regulates trabecular bone formation in female mice. Bone 2014, 60, 68–77. [Google Scholar] [CrossRef] [Green Version]
- Hofbauer, L.C.; Khosla, S.; Dunstan, C.R.; Lacey, D.L.; Spelsberg, T.C.; Riggs, B.L. Estrogen stimulates gene expression and protein production of osteoprotegerin in human osteoblastic cells. Endocrinology 1999, 140, 4367–4370. [Google Scholar] [CrossRef]
- Streicher, C.; Heyny, A.; Andrukhova, O.; Haigl, B.; Slavic, S.; Schuler, C.; Kollmann, K.; Kantner, I.; Sexl, V.; Kleiter, M.; et al. Estrogen Regulates Bone Turnover by Targeting RANKL Expression in Bone Lining Cells. Sci. Rep. 2017, 7, 6460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pepene, C.E.; Seck, T.; Diel, I.; Minne, H.W.; Ziegler, R.; Pfeilschifter, J. Influence of fluor salts, hormone replacement therapy and calcitonin on the concentration of insulin-like growth factor (IGF)-I, IGF-II and transforming growth factor-beta 1 in human iliac crest bone matrix from patients with primary osteoporosis. Eur. J. Endocrinol. 2004, 150, 81–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehnert, S.; Baur, J.; Schmitt, A.; Neumaier, M.; Lucke, M.; Dooley, S.; Vester, H.; Wildemann, B.; Stockle, U.; Nussler, A.K. TGF-beta1 as possible link between loss of bone mineral density and chronic inflammation. PLoS ONE 2010, 5, e14073. [Google Scholar] [CrossRef] [PubMed]
- Modder, U.I.; Clowes, J.A.; Hoey, K.; Peterson, J.M.; McCready, L.; Oursler, M.J.; Riggs, B.L.; Khosla, S. Regulation of circulating sclerostin levels by sex steroids in women and in men. J. Bone Miner. Res. 2011, 26, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Clarke, B.L.; Drake, M.T. Clinical utility of serum sclerostin measurements. Bonekey Rep. 2013, 2, 361. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, A.; Rotwein, P. Insulin-like growth factor-binding protein-5 inhibits osteoblast differentiation and skeletal growth by blocking insulin-like growth factor actions. Mol. Endocrinol. 2008, 22, 1238–1250. [Google Scholar] [CrossRef] [Green Version]
- Ueland, T.; Brixen, K.; Mosekilde, L.; Mosekilde, L.; Flyvbjerg, A.; Bollerslev, J. Age-related changes in cortical bone content of insulin-like growth factor binding protein (IGFBP)-3, IGFBP-5, osteoprotegerin, and calcium in postmenopausal osteoporosis: A cross-sectional study. J. Clin. Endocrinol. Metab. 2003, 88, 1014–1018. [Google Scholar] [CrossRef] [Green Version]
- Ye, C.; Hou, W.; Chen, M.; Lu, J.; Chen, E.; Tang, L.; Hang, K.; Ding, Q.; Li, Y.; Zhang, W.; et al. IGFBP7 acts as a negative regulator of RANKL-induced osteoclastogenesis and oestrogen deficiency-induced bone loss. Cell Prolif. 2020, 53, e12752. [Google Scholar] [CrossRef] [Green Version]
- Taxel, P.; Choksi, P.; Van Poznak, C. The management of osteoporosis in breast cancer survivors. Maturitas 2012, 73, 275–279. [Google Scholar] [CrossRef] [Green Version]
- Muhammad, A.; Mada, S.B.; Malami, I.; Forcados, G.E.; Erukainure, O.L.; Sani, H.; Abubakar, I.B. Postmenopausal osteoporosis and breast cancer: The biochemical links and beneficial effects of functional foods. Biomed. Pharmacother. 2018, 107, 571–582. [Google Scholar] [CrossRef] [PubMed]
- Llorente, I.; Garcia-Castaneda, N.; Valero, C.; Gonzalez-Alvaro, I.; Castaneda, S. Osteoporosis in Rheumatoid Arthritis: Dangerous Liaisons. Front. Med. 2020, 7, 601618. [Google Scholar] [CrossRef] [PubMed]
- Wysham, K.D.; Baker, J.F.; Shoback, D.M. Osteoporosis and fractures in rheumatoid arthritis. Curr. Opin. Rheumatol. 2021, 33, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Zhang, J.; Wang, J.; Lv, X.; Pu, D.; Wang, Y.; Men, Q.; He, L. Uncoupled bone remodeling is characteristic of bone damage in premenopausal women with new-onset systemic lupus erythematosus. Lupus 2021, 30, 1116–1123. [Google Scholar] [CrossRef]
- Nitta, K.; Yajima, A.; Tsuchiya, K. Management of Osteoporosis in Chronic Kidney Disease. Intern. Med. 2017, 56, 3271–3276. [Google Scholar] [CrossRef] [Green Version]
- Guanabens, N.; Pares, A. Osteoporosis in chronic liver disease. Liver Int. 2018, 38, 776–785. [Google Scholar] [CrossRef] [Green Version]
- Kurra, S.; Fink, D.A.; Siris, E.S. Osteoporosis-associated fracture and diabetes. Endocrinol. Metab. Clin. N. Am. 2014, 43, 233–243. [Google Scholar] [CrossRef]
- Farr, J.N.; Drake, M.T.; Amin, S.; Melton, L.J., 3rd; McCready, L.K.; Khosla, S. In vivo assessment of bone quality in postmenopausal women with type 2 diabetes. J. Bone Miner. Res. 2014, 29, 787–795. [Google Scholar] [CrossRef]
- Invernizzi, M.; Carda, S.; Viscontini, G.S.; Cisari, C. Osteoporosis in Parkinson’s disease. Parkinsonism Relat. Disord. 2009, 15, 339–346. [Google Scholar] [CrossRef]
- Mumford, E.R.; Raffles, S.; Reynolds, P. Coexistent osteoporosis and multiple myeloma: When to investigate further in osteoporosis. BMJ Case Rep. 2015, 2015, bcr2015210896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinman, J.; Shibli-Rahhal, A. Anorexia Nervosa and Osteoporosis: Pathophysiology and Treatment. J. Bone Metab. 2019, 26, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Kelsey, J.L. Risk factors for osteoporosis and associated fractures. Public Health Rep. 1989, 104, 14–20. [Google Scholar]
- Nordin, B.E. Calcium and osteoporosis. Nutrition 1997, 13, 664–686. [Google Scholar] [CrossRef]
- McGraw, R.L.; Riggs, J.E. Osteoporosis, sedentary lifestyle, and increasing hip fractures: Pathogenic relationship or differential survival bias. Calcif. Tissue Int. 1994, 55, 87–89. [Google Scholar] [CrossRef] [PubMed]
- de Vernejoul, M.C.; Bielakoff, J.; Herve, M.; Gueris, J.; Hott, M.; Modrowski, D.; Kuntz, D.; Miravet, L.; Ryckewaert, A. Evidence for defective osteoblastic function. A role for alcohol and tobacco consumption in osteoporosis in middle-aged men. Clin. Orthop. Relat. Res. 1983, 179, 107–115. [Google Scholar] [CrossRef]
- Cheraghi, Z.; Doosti-Irani, A.; Almasi-Hashiani, A.; Baigi, V.; Mansournia, N.; Etminan, M.; Mansournia, M.A. The effect of alcohol on osteoporosis: A systematic review and meta-analysis. Drug Alcohol. Depend. 2019, 197, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Daniell, H.W. Osteoporosis and smoking. JAMA 1972, 221, 509. [Google Scholar] [CrossRef]
- Walker, L.M.; Preston, M.R.; Magnay, J.L.; Thomas, P.B.; El Haj, A.J. Nicotinic regulation of c-fos and osteopontin expression in human-derived osteoblast-like cells and human trabecular bone organ culture. Bone 2001, 28, 603–608. [Google Scholar] [CrossRef]
- Nicholls, A.R.; Holt, R.I. Growth Hormone and Insulin-Like Growth Factor-1. Front. Horm. Res. 2016, 47, 101–114. [Google Scholar] [CrossRef]
- Christianson, M.S.; Shen, W. Osteoporosis prevention and management: Nonpharmacologic and lifestyle options. Clin. Obstet. Gynecol. 2013, 56, 703–710. [Google Scholar] [CrossRef]
- Malorgio, A.; Malorgio, M.; Benedetti, M.; Casarosa, S.; Cannataro, R. High intensity resistance training as intervention method to knee osteoarthritis. Sports Med. Health Sci. 2021, 3, 46–48. [Google Scholar] [CrossRef]
- Moreira, L.D.F.; Oliveira, M.L.d.; Lirani-Galvão, A.P.; Marin-Mio, R.V.; Santos, R.N.d.; Lazaretti-Castro, M. Physical exercise and osteoporosis: Effects of different types of exercises on bone and physical function of postmenopausal women. Arq. Bras. Endocrinol. Metabol. 2014, 58, 514–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, P.C.; Siu, W.S. Herbal treatment for osteoporosis: A current review. J. Tradit. Complement. Med. 2013, 3, 82–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russow, G.; Jahn, D.; Appelt, J.; Mardian, S.; Tsitsilonis, S.; Keller, J. Anabolic Therapies in Osteoporosis and Bone Regeneration. Int. J. Mol. Sci. 2018, 20, 83. [Google Scholar] [CrossRef] [Green Version]
- Bi, H.; Chen, X.; Gao, S.; Yu, X.; Xiao, J.; Zhang, B.; Liu, X.; Dai, M. Key Triggers of Osteoclast-Related Diseases and Available Strategies for Targeted Therapies: A Review. Front. Med. 2017, 4, 234. [Google Scholar] [CrossRef]
- Brewer, H.B., Jr.; Fairwell, T.; Ronan, R.; Sizemore, G.W.; Arnaud, C.D. Human parathyroid hormone: Amino-acid sequence of the amino-terminal residues 1-34. Proc. Natl. Acad. Sci. USA 1972, 69, 3585–3588. [Google Scholar] [CrossRef] [Green Version]
- Librizzi, M.; Tobiasch, E.; Luparello, C. The conditioned medium from osteo-differentiating human mesenchymal stem cells affects the viability of triple negative MDA-MB231 breast cancer cells. Cell Biochem. Funct. 2016, 34, 7–15. [Google Scholar] [CrossRef]
- Mannstadt, M.; Juppner, H.; Gardella, T.J. Receptors for PTH and PTHrP: Their biological importance and functional properties. Am. J. Physiol. 1999, 277, F665–F675. [Google Scholar] [CrossRef]
- Altarejos, J.Y.; Montminy, M. CREB and the CRTC co-activators: Sensors for hormonal and metabolic signals. Nat. Rev. Mol. Cell Biol. 2011, 12, 141–151. [Google Scholar] [CrossRef] [Green Version]
- Wein, M.N.; Liang, Y.; Goransson, O.; Sundberg, T.B.; Wang, J.; Williams, E.A.; O’Meara, M.J.; Govea, N.; Beqo, B.; Nishimori, S.; et al. SIKs control osteocyte responses to parathyroid hormone. Nat. Commun. 2016, 7, 13176. [Google Scholar] [CrossRef]
- Podbesek, R.; Edouard, C.; Meunier, P.J.; Parsons, J.A.; Reeve, J.; Stevenson, R.W.; Zanelli, J.M. Effects of two treatment regimes with synthetic human parathyroid hormone fragment on bone formation and the tissue balance of trabecular bone in greyhounds. Endocrinology 1983, 112, 1000–1006. [Google Scholar] [CrossRef] [PubMed]
- Neer, R.M.; Arnaud, C.D.; Zanchetta, J.R.; Prince, R.; Gaich, G.A.; Reginster, J.Y.; Hodsman, A.B.; Eriksen, E.F.; Ish-Shalom, S.; Genant, H.K.; et al. Effect of parathyroid hormone (1-34) on fractures and bone mineral density in postmenopausal women with osteoporosis. N. Engl. J. Med. 2001, 344, 1434–1441. [Google Scholar] [CrossRef] [PubMed]
- Vahle, J.L.; Sato, M.; Long, G.G.; Young, J.K.; Francis, P.C.; Engelhardt, J.A.; Westmore, M.S.; Linda, Y.; Nold, J.B. Skeletal changes in rats given daily subcutaneous injections of recombinant human parathyroid hormone (1-34) for 2 years and relevance to human safety. Toxicol. Pathol. 2002, 30, 312–321. [Google Scholar] [CrossRef]
- Ehrenmann, J.; Schoppe, J.; Klenk, C.; Rappas, M.; Kummer, L.; Dore, A.S.; Pluckthun, A. High-resolution crystal structure of parathyroid hormone 1 receptor in complex with a peptide agonist. Nat. Struct. Mol. Biol. 2018, 25, 1086–1092. [Google Scholar] [CrossRef] [PubMed]
- Migliaccio, S.; Brama, M.; Malavolta, N. Management of glucocorticoids-induced osteoporosis: Role of teriparatide. Ther. Clin. Risk Manag. 2009, 5, 305–310. [Google Scholar] [CrossRef]
- Karvande, A.; Kushwaha, P.; Ahmad, N.; Adhikary, S.; Kothari, P.; Tripathi, A.K.; Khedgikar, V.; Trivedi, R. Glucose dependent miR-451a expression contributes to parathyroid hormone mediated osteoblast differentiation. Bone 2018, 117, 98–115. [Google Scholar] [CrossRef]
- Saag, K.G.; Shane, E.; Boonen, S.; Marin, F.; Donley, D.W.; Taylor, K.A.; Dalsky, G.P.; Marcus, R. Teriparatide or alendronate in glucocorticoid-induced osteoporosis. N. Engl. J. Med. 2007, 357, 2028–2039. [Google Scholar] [CrossRef] [Green Version]
- Miller, P.D.; Hattersley, G.; Riis, B.J.; Williams, G.C.; Lau, E.; Russo, L.A.; Alexandersen, P.; Zerbini, C.A.; Hu, M.Y.; Harris, A.G.; et al. Effect of Abaloparatide vs Placebo on New Vertebral Fractures in Postmenopausal Women with Osteoporosis: A Randomized Clinical Trial. JAMA 2016, 316, 722–733. [Google Scholar] [CrossRef]
- Cupp, M.E.; Nayak, S.K.; Adem, A.S.; Thomsen, W.J. Parathyroid hormone (PTH) and PTH-related peptide domains contributing to activation of different PTH receptor-mediated signaling pathways. J. Pharmacol. Exp. Ther. 2013, 345, 404–418. [Google Scholar] [CrossRef] [Green Version]
- Leali, P.T.; Balsano, M.; Maestretti, G.; Brusoni, M.; Amorese, V.; Ciurlia, E.; Andreozzi, M.; Caggiari, G.; Doria, C. Efficacy of teriparatide vs neridronate in adults with osteogenesis imperfecta type I: A prospective randomized international clinical study. Clin. Cases Miner. Bone Metab. 2017, 14, 153–156. [Google Scholar] [CrossRef]
- Koski, A.M.; Sikio, A.; Forslund, T. Teriparatide treatment complicated by malignant myeloma. BMJ Case Rep. 2010, 2010, bcr0120102681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makino, A.; Takagi, H.; Takahashi, Y.; Hase, N.; Sugiyama, H.; Yamana, K.; Kobayashi, T. Abaloparatide Exerts Bone Anabolic Effects with Less Stimulation of Bone Resorption-Related Factors: A Comparison with Teriparatide. Calcif. Tissue Int. 2018, 103, 289–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leder, B.Z.; O’Dea, L.S.; Zanchetta, J.R.; Kumar, P.; Banks, K.; McKay, K.; Lyttle, C.R.; Hattersley, G. Effects of abaloparatide, a human parathyroid hormone-related peptide analog, on bone mineral density in postmenopausal women with osteoporosis. J. Clin. Endocrinol. Metab. 2015, 100, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Ricarte, F.R.; Le Henaff, C.; Kolupaeva, V.G.; Gardella, T.J.; Partridge, N.C. Parathyroid hormone(1-34) and its analogs differentially modulate osteoblastic Rankl expression via PKA/SIK2/SIK3 and PP1/PP2A-CRTC3 signaling. J. Biol. Chem. 2018, 293, 20200–20213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, P.D.; Bilezikian, J.P.; Fitzpatrick, L.A.; Mitlak, B.; McCloskey, E.V.; Cosman, F.; Bone, H.G. Abaloparatide: An anabolic treatment to reduce fracture risk in postmenopausal women with osteoporosis. Curr. Med. Res. Opin. 2020, 36, 1861–1872. [Google Scholar] [CrossRef]
- Cosman, F.; Peterson, L.R.; Towler, D.A.; Mitlak, B.; Wang, Y.; Cummings, S.R. Cardiovascular Safety of Abaloparatide in Postmenopausal Women with Osteoporosis: Analysis from the ACTIVE Phase 3 Trial. J. Clin. Endocrinol. Metab. 2020, 105. [Google Scholar] [CrossRef]
- Yang, Y.; Lei, H.; Wang, B. Effect of the PTHrP(1-34) analog abaloparatide on inducing chondrogenesis involves inhibition of intracellular reactive oxygen species production. Biochem. Biophys. Res. Commun. 2019, 509, 960–965. [Google Scholar] [CrossRef]
- Minisola, S. Romosozumab: From basic to clinical aspects. Expert Opin. Biol. Ther. 2014, 14, 1225–1228. [Google Scholar] [CrossRef] [Green Version]
- Larsson, S. Anti-sclerostin—Is there an indication? Injury 2016, 47 (Suppl. 1), S31–S35. [Google Scholar] [CrossRef]
- van Dinther, M.; Zhang, J.; Weidauer, S.E.; Boschert, V.; Muth, E.M.; Knappik, A.; de Gorter, D.J.; van Kasteren, P.B.; Frisch, C.; Mueller, T.D.; et al. Anti-Sclerostin antibody inhibits internalization of Sclerostin and Sclerostin-mediated antagonism of Wnt/LRP6 signaling. PLoS ONE 2013, 8, e62295. [Google Scholar] [CrossRef] [Green Version]
- Glorieux, F.H.; Devogelaer, J.P.; Durigova, M.; Goemaere, S.; Hemsley, S.; Jakob, F.; Junker, U.; Ruckle, J.; Seefried, L.; Winkle, P.J. BPS804 Anti-Sclerostin Antibody in Adults With Moderate Osteogenesis Imperfecta: Results of a Randomized Phase 2a Trial. J. Bone Miner. Res. 2017, 32, 1496–1504. [Google Scholar] [CrossRef] [PubMed]
- Cosman, F.; Crittenden, D.B.; Adachi, J.D.; Binkley, N.; Czerwinski, E.; Ferrari, S.; Hofbauer, L.C.; Lau, E.; Lewiecki, E.M.; Miyauchi, A.; et al. Romosozumab Treatment in Postmenopausal Women with Osteoporosis. N. Engl. J. Med. 2016, 375, 1532–1543. [Google Scholar] [CrossRef] [PubMed]
- Saag, K.G.; Petersen, J.; Brandi, M.L.; Karaplis, A.C.; Lorentzon, M.; Thomas, T.; Maddox, J.; Fan, M.; Meisner, P.D.; Grauer, A. Romosozumab or Alendronate for Fracture Prevention in Women with Osteoporosis. N. Engl. J. Med. 2017, 377, 1417–1427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, M.; Sansalone, V.; Cooper, D.M.L.; Forwood, M.R.; Pivonka, P. Assessment of romosozumab efficacy in the treatment of postmenopausal osteoporosis: Results from a mechanistic PK-PD mechanostat model of bone remodeling. Bone 2020, 133, 115223. [Google Scholar] [CrossRef] [PubMed]
- Heaney, R.P.; Saville, P.D. Etidronate disodium in postmenopausal osteoporosis. Clin. Pharmacol. Ther. 1976, 20, 593–604. [Google Scholar] [CrossRef]
- Lepore, L.; Pennesi, M.; Barbi, E.; Pozzi, R. Treatment and prevention of osteoporosis in juvenile chronic arthritis with disodium clodronate. Clin. Exp. Rheumatol. 1991, 9 (Suppl. 6), 33–35. [Google Scholar]
- Chesnut, C.H., 3rd. Tiludronate: Development as an osteoporosis therapy. Bone 1995, 17, 517S–519S. [Google Scholar] [CrossRef]
- Adami, S.; Baroni, M.C.; Broggini, M.; Carratelli, L.; Caruso, I.; Gnessi, L.; Laurenzi, M.; Lombardi, A.; Norbiato, G.; Ortolani, S.; et al. Treatment of postmenopausal osteoporosis with continuous daily oral alendronate in comparison with either placebo or intranasal salmon calcitonin. Osteoporos. Int. 1993, 3 (Suppl. 3), S21–S27. [Google Scholar] [CrossRef]
- Rossini, M.; Gatti, D.; Zamberlan, N.; Braga, V.; Dorizzi, R.; Adami, S. Long-term effects of a treatment course with oral alendronate of postmenopausal osteoporosis. J. Bone Miner. Res. 1994, 9, 1833–1837. [Google Scholar] [CrossRef]
- Black, D.M.; Cummings, S.R.; Karpf, D.B.; Cauley, J.A.; Thompson, D.E.; Nevitt, M.C.; Bauer, D.C.; Genant, H.K.; Haskell, W.L.; Marcus, R.; et al. Randomised trial of effect of alendronate on risk of fracture in women with existing vertebral fractures. Fracture Intervention Trial Research Group. Lancet 1996, 348, 1535–1541. [Google Scholar] [CrossRef]
- Clemmesen, B.; Ravn, P.; Zegels, B.; Taquet, A.N.; Christiansen, C.; Reginster, J.Y. A 2-year phase II study with 1-year of follow-up of risedronate (NE-58095) in postmenopausal osteoporosis. Osteoporos. Int. 1997, 7, 488–495. [Google Scholar] [CrossRef] [PubMed]
- Harris, S.T.; Watts, N.B.; Genant, H.K.; McKeever, C.D.; Hangartner, T.; Keller, M.; Chesnut, C.H., 3rd; Brown, J.; Eriksen, E.F.; Hoseyni, M.S.; et al. Effects of risedronate treatment on vertebral and nonvertebral fractures in women with postmenopausal osteoporosis: A randomized controlled trial. Vertebral Efficacy with Risedronate Therapy (VERT) Study Group. JAMA 1999, 282, 1344–1352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braga, V.; Gatti, D.; Colapietro, F.; Battaglia, E.; Righetti, D.; Prizzi, R.; Rossini, M.; Adami, S. Intravenous intermittent neridronate in the treatment of postmenopausal osteoporosis. Bone 2003, 33, 342–345. [Google Scholar] [CrossRef]
- Cascella, T.; Musella, T.; Orio, F., Jr.; Palomba, S.; Bifulco, G.; Nappi, C.; Lombardi, G.; Colao, A.; Tauchmanova, L. Effects of neridronate treatment in elderly women with osteoporosis. J. Endocrinol. Investig. 2005, 28, 202–208. [Google Scholar] [CrossRef]
- Ravn, P.; Clemmesen, B.; Riis, B.J.; Christiansen, C. The effect on bone mass and bone markers of different doses of ibandronate: A new bisphosphonate for prevention and treatment of postmenopausal osteoporosis: A 1-year, randomized, double-blind, placebo-controlled dose-finding study. Bone 1996, 19, 527–533. [Google Scholar] [CrossRef]
- Chesnut, C.H., 3rd; Skag, A.; Christiansen, C.; Recker, R.; Stakkestad, J.A.; Hoiseth, A.; Felsenberg, D.; Huss, H.; Gilbride, J.; Schimmer, R.C.; et al. Effects of oral ibandronate administered daily or intermittently on fracture risk in postmenopausal osteoporosis. J. Bone Miner. Res. 2004, 19, 1241–1249. [Google Scholar] [CrossRef] [Green Version]
- Roldan, E.J.; Perez-Llore, A.; Ferretti, J.L. Olpadronate: A new amino-bisphosphonate for the treatment of medical osteopathies. Expert Opin. Investig. Drugs 1998, 7, 1521–1538. [Google Scholar] [CrossRef]
- Mallmin, H.; Ljunghall, S.; Larsson, K.; Lindh, E. Short-term effects of pamidronate on biochemical markers of bone metabolism in osteoporosis—A placebo-controlled dose-finding study. Upsala J. Med. Sci. 1991, 96, 205–212. [Google Scholar] [CrossRef]
- Perifanis, V.; Vyzantiadis, T.; Vakalopoulou, S.; Tziomalos, K.; Garypidou, V.; Athanassiou-Metaxa, M.; Harsoulis, F. Treatment of beta-thalassaemia-associated osteoporosis with zoledronic acid. Br. J. Haematol. 2004, 125, 91–92. [Google Scholar] [CrossRef]
- Baron, R.; Ferrari, S.; Russell, R.G. Denosumab and bisphosphonates: Different mechanisms of action and effects. Bone 2011, 48, 677–692. [Google Scholar] [CrossRef]
- Russell, R.G.; Watts, N.B.; Ebetino, F.H.; Rogers, M.J. Mechanisms of action of bisphosphonates: Similarities and differences and their potential influence on clinical efficacy. Osteoporos. Int. 2008, 19, 733–759. [Google Scholar] [CrossRef] [PubMed]
- Frith, J.C.; Monkkonen, J.; Auriola, S.; Monkkonen, H.; Rogers, M.J. The molecular mechanism of action of the antiresorptive and antiinflammatory drug clodronate: Evidence for the formation in vivo of a metabolite that inhibits bone resorption and causes osteoclast and macrophage apoptosis. Arthritis Rheum. 2001, 44, 2201–2210. [Google Scholar] [CrossRef]
- Nancollas, G.H.; Tang, R.; Phipps, R.J.; Henneman, Z.; Gulde, S.; Wu, W.; Mangood, A.; Russell, R.G.; Ebetino, F.H. Novel insights into actions of bisphosphonates on bone: Differences in interactions with hydroxyapatite. Bone 2006, 38, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Tsai, J.N.; Lee, H.; David, N.L.; Eastell, R.; Leder, B.Z. Combination denosumab and high dose teriparatide for postmenopausal osteoporosis (DATA-HD): A randomised, controlled phase 4 trial. Lancet Diabetes Endocrinol. 2019, 7, 767–775. [Google Scholar] [CrossRef] [Green Version]
- Naylor, K.E.; McCloskey, E.V.; Jacques, R.M.; Peel, N.F.A.; Paggiosi, M.A.; Gossiel, F.; Walsh, J.S.; Eastell, R. Clinical utility of bone turnover markers in monitoring the withdrawal of treatment with oral bisphosphonates in postmenopausal osteoporosis. Osteoporos. Int. 2019, 30, 917–922. [Google Scholar] [CrossRef] [Green Version]
- Aki, T.; Hashimoto, K.; Uozumi, H.; Saito, M.; Sugawara, K.; Suzuki, M.; Hamada, S.; Ito, A.; Itoi, E. Morphological and Morphometrical Analyses of Fracture-Healing Sites of an Atypical Femoral Fracture in Patients with and without Long-Term Bisphosphonate Treatment for Osteoporosis: A Report of Two Cases. Tohoku J. Exp. Med. 2021, 253, 261–267. [Google Scholar] [CrossRef]
- Rosen, C.J.; Brown, S. Severe hypocalcemia after intravenous bisphosphonate therapy in occult vitamin D deficiency. N. Engl. J. Med. 2003, 348, 1503–1504. [Google Scholar] [CrossRef]
- Schussheim, D.H.; Jacobs, T.P.; Silverberg, S.J. Hypocalcemia associated with alendronate. Ann. Intern. Med. 1999, 130, 329. [Google Scholar] [CrossRef]
- Aparecida Cariolatto, F.; Carelli, J.; de Campos Moreira, T.; Pietrobon, R.; Rodrigues, C.; Bonilauri Ferreira, A.P. Recommendations for the Prevention of Bisphosphonate-Related Osteonecrosis of the Jaw: A Systematic Review. J. Evid. Based. Dent. Pract. 2018, 18, 142–152. [Google Scholar] [CrossRef]
- Ganesan, K.; Bansal, P.; Goyal, A.; Roane, D. Bisphosphonate. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Cosman, F.; Lindsay, R. Selective estrogen receptor modulators: Clinical spectrum. Endocr. Rev. 1999, 20, 418–434. [Google Scholar] [CrossRef]
- Obrecht, J.P. Tamoxifen for osteoporosis prevention? Dtsch. Med. Wochenschr. 1989, 114, 1465–1466. [Google Scholar] [PubMed]
- Frolik, C.A.; Bryant, H.U.; Black, E.C.; Magee, D.E.; Chandrasekhar, S. Time-dependent changes in biochemical bone markers and serum cholesterol in ovariectomized rats: Effects of raloxifene HCl, tamoxifen, estrogen, and alendronate. Bone 1996, 18, 621–627. [Google Scholar] [CrossRef]
- Rey, J.R.; Cervino, E.V.; Rentero, M.L.; Crespo, E.C.; Alvaro, A.O.; Casillas, M. Raloxifene: Mechanism of action, effects on bone tissue, and applicability in clinical traumatology practice. Open Orthop. J. 2009, 3, 14–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silverman, S.L.; Christiansen, C.; Genant, H.K.; Vukicevic, S.; Zanchetta, J.R.; de Villiers, T.J.; Constantine, G.D.; Chines, A.A. Efficacy of bazedoxifene in reducing new vertebral fracture risk in postmenopausal women with osteoporosis: Results from a 3-year, randomized, placebo-, and active-controlled clinical trial. J. Bone Miner. Res. 2008, 23, 1923–1934. [Google Scholar] [CrossRef]
- Ke, H.Z.; Brown, T.A.; Thompson, D.D. Lasofoxifene (CP-336,156), a novel selective estrogen receptor modulator, in preclinical studies. J. Am. Aging Assoc. 2002, 25, 87–99. [Google Scholar] [CrossRef] [Green Version]
- Jordan, V.C. Long-term tamoxifen therapy to control or to prevent breast cancer: Laboratory concept to clinical trials. Prog. Clin. Biol. Res. 1988, 262, 105–123. [Google Scholar]
- Gianni, W.; Ricci, A.; Gazzaniga, P.; Brama, M.; Pietropaolo, M.; Votano, S.; Patane, F.; Agliano, A.M.; Spera, G.; Marigliano, V.; et al. Raloxifene modulates interleukin-6 and tumor necrosis factor-alpha synthesis in vivo: Results from a pilot clinical study. J. Clin. Endocrinol. Metab. 2004, 89, 6097–6099. [Google Scholar] [CrossRef] [Green Version]
- Jilka, R.L.; Hangoc, G.; Girasole, G.; Passeri, G.; Williams, D.C.; Abrams, J.S.; Boyce, B.; Broxmeyer, H.; Manolagas, S.C. Increased osteoclast development after estrogen loss: Mediation by interleukin-6. Science 1992, 257, 88–91. [Google Scholar] [CrossRef]
- Kumar, V.; Green, S.; Stack, G.; Berry, M.; Jin, J.R.; Chambon, P. Functional domains of the human estrogen receptor. Cell 1987, 51, 941–951. [Google Scholar] [CrossRef]
- Johnston, C.C., Jr.; Bjarnason, N.H.; Cohen, F.J.; Shah, A.; Lindsay, R.; Mitlak, B.H.; Huster, W.; Draper, M.W.; Harper, K.D.; Heath, H., 3rd; et al. Long-term effects of raloxifene on bone mineral density, bone turnover, and serum lipid levels in early postmenopausal women: Three-year data from 2 double-blind, randomized, placebo-controlled trials. Arch. Intern. Med. 2000, 160, 3444–3450. [Google Scholar] [CrossRef] [PubMed]
- Reddy, G.K.; Nadler, E.; Jain, V.K. Denosumab (AMG 162), a Fully Human Monoclonal Antibody against RANK Ligand Activity. Support. Cancer Ther. 2005, 3, 14–15. [Google Scholar] [CrossRef]
- Bord, S.; Ireland, D.C.; Beavan, S.R.; Compston, J.E. The effects of estrogen on osteoprotegerin, RANKL, and estrogen receptor expression in human osteoblasts. Bone 2003, 32, 136–141. [Google Scholar] [CrossRef]
- McClung, M.R. Inhibition of RANKL as a treatment for osteoporosis: Preclinical and early clinical studies. Curr. Osteoporos. Rep. 2006, 4, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Cummings, S.R.; San Martin, J.; McClung, M.R.; Siris, E.S.; Eastell, R.; Reid, I.R.; Delmas, P.; Zoog, H.B.; Austin, M.; Wang, A.; et al. Denosumab for prevention of fractures in postmenopausal women with osteoporosis. N. Engl. J. Med. 2009, 361, 756–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, N.; Zhang, Z.K.; Yu, Y.; Zhuo, Z.; Zhang, G.; Zhang, B.T. Pros and Cons of Denosumab Treatment for Osteoporosis and Implication for RANKL Aptamer Therapy. Front. Cell Dev. Biol. 2020, 8, 325. [Google Scholar] [CrossRef] [PubMed]
- Tsourdi, E.; Zillikens, M.C.; Meier, C.; Body, J.J.; Gonzalez Rodriguez, E.; Anastasilakis, A.D.; Abrahamsen, B.; McCloskey, E.; Hofbauer, L.C.; Guanabens, N.; et al. Fracture risk and management of discontinuation of denosumab therapy: A systematic review and position statement by ECTS. J. Clin. Endocrinol. Metab. 2021, 106, 264–281. [Google Scholar] [CrossRef] [PubMed]
- Nicolatou-Galitis, O.; Schiodt, M.; Mendes, R.A.; Ripamonti, C.; Hope, S.; Drudge-Coates, L.; Niepel, D.; Van den Wyngaert, T. Medication-related osteonecrosis of the jaw: Definition and best practice for prevention, diagnosis, and treatment. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2019, 127, 117–135. [Google Scholar] [CrossRef] [Green Version]
- Boquete-Castro, A.; Gomez-Moreno, G.; Calvo-Guirado, J.L.; Aguilar-Salvatierra, A.; Delgado-Ruiz, R.A. Denosumab and osteonecrosis of the jaw. A systematic analysis of events reported in clinical trials. Clin. Oral Implants Res. 2016, 27, 367–375. [Google Scholar] [CrossRef]
- Bossard, M.J.; Tomaszek, T.A.; Thompson, S.K.; Amegadzie, B.Y.; Hanning, C.R.; Jones, C.; Kurdyla, J.T.; McNulty, D.E.; Drake, F.H.; Gowen, M.; et al. Proteolytic activity of human osteoclast cathepsin K. Expression, purification, activation, and substrate identification. J. Biol. Chem. 1996, 271, 12517–12524. [Google Scholar] [CrossRef] [Green Version]
- Drake, F.H.; Dodds, R.A.; James, I.E.; Connor, J.R.; Debouck, C.; Richardson, S.; Lee-Rykaczewski, E.; Coleman, L.; Rieman, D.; Barthlow, R.; et al. Cathepsin K, but not cathepsins B, L, or S, is abundantly expressed in human osteoclasts. J. Biol. Chem. 1996, 271, 12511–12516. [Google Scholar] [CrossRef] [Green Version]
- Costa, A.G.; Cusano, N.E.; Silva, B.C.; Cremers, S.; Bilezikian, J.P. Cathepsin K: Its skeletal actions and role as a therapeutic target in osteoporosis. Nat. Rev. Rheumatol. 2011, 7, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Garnero, P.; Borel, O.; Byrjalsen, I.; Ferreras, M.; Drake, F.H.; McQueney, M.S.; Foged, N.T.; Delmas, P.D.; Delaisse, J.M. The collagenolytic activity of cathepsin K is unique among mammalian proteinases. J. Biol. Chem. 1998, 273, 32347–32352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kafienah, W.; Bromme, D.; Buttle, D.J.; Croucher, L.J.; Hollander, A.P. Human cathepsin K cleaves native type I and II collagens at the N-terminal end of the triple helix. Biochem. J. 1998, 331 Pt 3, 727–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bromme, D.; Lecaille, F. Cathepsin K inhibitors for osteoporosis and potential off-target effects. Expert Opin. Investig. Drugs 2009, 18, 585–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peroni, A.; Zini, A.; Braga, V.; Colato, C.; Adami, S.; Girolomoni, G. Drug-induced morphea: Report of a case induced by balicatib and review of the literature. J. Am. Acad. Dermatol. 2008, 59, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Runger, T.M.; Adami, S.; Benhamou, C.L.; Czerwinski, E.; Farrerons, J.; Kendler, D.L.; Mindeholm, L.; Realdi, G.; Roux, C.; Smith, V. Morphea-like skin reactions in patients treated with the cathepsin K inhibitor balicatib. J. Am. Acad. Dermatol. 2012, 66, e89–e96. [Google Scholar] [CrossRef]
- Tanaka, M.; Hashimoto, Y.; Hasegawa, C.; Deacon, S.; Eastell, R. Antiresorptive effect of a cathepsin K inhibitor ONO-5334 and its relationship to BMD increase in a phase II trial for postmenopausal osteoporosis. BMC Musculoskelet. Disord. 2017, 18, 267. [Google Scholar] [CrossRef] [Green Version]
- Yamada, H.; Mori, H.; Nakanishi, Y.; Nishikawa, S.; Hashimoto, Y.; Ochi, Y.; Tanaka, M.; Kawabata, K. Effects of the Cathepsin K Inhibitor ONO-5334 and Concomitant Use of ONO-5334 with Methotrexate on Collagen-Induced Arthritis in Cynomolgus Monkeys. Int. J. Rheumatol. 2019, 2019, 5710340. [Google Scholar] [CrossRef]
- McClung, M.R.; O’Donoghue, M.L.; Papapoulos, S.E.; Bone, H.; Langdahl, B.; Saag, K.G.; Reid, I.R.; Kiel, D.P.; Cavallari, I.; Bonaca, M.P.; et al. Odanacatib for the treatment of postmenopausal osteoporosis: Results of the LOFT multicentre, randomised, double-blind, placebo-controlled trial and LOFT Extension study. Lancet Diabetes Endocrinol. 2019, 7, 899–911. [Google Scholar] [CrossRef]
- Solomon, D.H.; Rekedal, L.; Cadarette, S.M. Osteoporosis treatments and adverse events. Curr. Opin. Rheumatol. 2009, 21, 363–368. [Google Scholar] [CrossRef]
- Wei, K.; Qu, Y.; Gao, Y.; Ma, Y. Comparison of Efficacy of Teriparatide (Parathyroid Hormone 1-34) Alone and in Combination with Zoledronic Acid for Osteoporosis in Postmenopausal Women. J. Coll. Physicians Surg. Pak. 2021, 31, 240–242. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Arita, K.; Murota, E.; Hiratsuka, S.; Fujita, R.; Ishizu, H.; Asano, T.; Takahashi, D.; Takahata, M.; Iwasaki, N. Effects after starting or switching from bisphosphonate to romosozumab or denosumab in Japanese postmenopausal patients. J. Bone Miner. Metab. 2021, 39, 868–875. [Google Scholar] [CrossRef] [PubMed]
- Santa Maria, C.; Cheng, Z.; Li, A.; Wang, J.; Shoback, D.; Tu, C.L.; Chang, W. Interplay between CaSR and PTH1R signaling in skeletal development and osteoanabolism. Semin. Cell Dev. Biol. 2016, 49, 11–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etani, Y.; Ebina, K.; Hirao, M.; Kitaguchi, K.; Kashii, M.; Ishimoto, T.; Nakano, T.; Okamura, G.; Miyama, A.; Takami, K.; et al. Combined effect of teriparatide and an anti-RANKL monoclonal antibody on bone defect regeneration in mice with glucocorticoid-induced osteoporosis. Bone 2020, 139, 115525. [Google Scholar] [CrossRef] [PubMed]
- Khajuria, D.K.; Razdan, R.; Mahapatra, D.R. The combination therapy with zoledronic Acid and propranolol improves the trabecular microarchitecture and mechanical property in an rat model of postmenopausal osteoporosis. J. Osteoporos. 2014, 2014, 586431. [Google Scholar] [CrossRef] [Green Version]
- Von Schacht, E.; Dambacher, M.A.; Ringe, J.D.; Dukas, L. Combination of alendronate plus alfacalcidol in the treatment of osteoporosis. Rationale, preclinical data and clinical evidence. MMW Fortschr. Med. 2012, 154 (Suppl. 1), 10–21. [Google Scholar]
- Lou, S.; Lv, H.; Li, Z.; Zhang, L.; Tang, P. Combination therapy of anabolic agents and bisphosphonates on bone mineral density in patients with osteoporosis: A meta-analysis of randomised controlled trials. BMJ Open 2018, 8, e015187. [Google Scholar] [CrossRef] [Green Version]
- Cosman, F.; Dempster, D.W. Anabolic Agents for Postmenopausal Osteoporosis: How Do You Choose? Curr. Osteoporos. Rep. 2021, 19, 189–205. [Google Scholar] [CrossRef]
- Cosman, F.; Lewiecki, E.M.; Ebeling, P.R.; Hesse, E.; Napoli, N.; Matsumoto, T.; Crittenden, D.B.; Rojeski, M.; Yang, W.; Libanati, C. T-score as an indicator of fracture risk during treatment with romosozumab or alendronate in the ARCH trial. J. Bone Miner. Res. 2020, 35, 1333–1342. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, Z.; Jiang, Y.; Liu, H.; Feng, Y.; Wang, Z.; Liu, H.; Wang, J.; Yang, B.; Lin, Q. Bioinspired mineral hydrogels as nanocomposite scaffolds for the promotion of osteogenic marker expression and the induction of bone regeneration in osteoporosis. Acta Biomater. 2020, 113, 614–626. [Google Scholar] [CrossRef]
- Zhu, Y.; Li, Z.; Zhang, Y.; Lan, F.; He, J.; Wu, Y. The essential role of osteoclast-derived exosomes in magnetic nanoparticle-infiltrated hydroxyapatite scaffold modulated osteoblast proliferation in an osteoporosis model. Nanoscale 2020, 12, 8720–8726. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, N.; Sukegawa, S.; Kitamura, A.; Goto, R.; Noda, T.; Nakano, K.; Takabatake, K.; Kawai, H.; Nagatsuka, H.; Kawasaki, K.; et al. Deep Learning for Osteoporosis Classification Using Hip Radiographs and Patient Clinical Covariates. Biomolecules 2020, 10, 1534. [Google Scholar] [CrossRef] [PubMed]
- Codrea, C.I.; Croitoru, A.-M.; Baciu, C.C.; Melinescu, A.; Ficai, D.; Fruth, V.; Ficai, A. Advances in Osteoporotic Bone Tissue Engineering. J. Clin. Med. 2021, 10, 253. [Google Scholar] [CrossRef]
- Hench, L.L.; Polak, J.M. Third-generation biomedical materials. Science 2002, 295, 1014–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, X.; Zhang, X.; Song, J.; Xu, X.; Xu, A.; Wang, M.; Xie, B.; Huang, E.; Deng, F.; Wei, S. Osteoinductive peptide-functionalized nanofibers with highly ordered structure as biomimetic scaffolds for bone tissue engineering. Int. J. Nanomed. 2015, 10, 7109–7128. [Google Scholar]
- Delgado-Ruiz, R.; Swanson, P.; Romanos, G. Systematic review of the long-term effects of transgender hormone therapy on bone markers and bone mineral density and their potential effects in implant therapy. J. Clin. Med. 2019, 8, 784. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Guo, J.L.; Wang, J.; Mikos, A.G.; Zhang, S. Hierarchically designed bone scaffolds: From internal cues to external stimuli. Biomaterials 2019, 218, 119334. [Google Scholar] [CrossRef]
- Yuan, B.; Wang, L.; Zhao, R.; Yang, X.; Yang, X.; Zhu, X.; Liu, L.; Zhang, K.; Song, Y.; Zhang, X. A biomimetically hierarchical polyetherketoneketone scaffold for osteoporotic bone repair. Sci. Adv. 2020, 6, eabc4704. [Google Scholar] [CrossRef]
- Amani, N.; Javar, H.A.; Dorkoosh, F.A.; Rouini, M.R.; Amini, M.; Sharifzadeh, M.; Boumi, S. Preparation and pulsatile release evaluation of teriparatide-loaded multilayer implant composed of polyanhydride-hydrogel layers using spin coating for the treatment of osteoporosis. J. Pharm. Innov. 2020, 16, 337–358. [Google Scholar] [CrossRef]
- Salamanna, F.; Gambardella, A.; Contartese, D.; Visani, A.; Fini, M. Nano-Based Biomaterials as Drug Delivery Systems against Osteoporosis: A Systematic Review of Preclinical and Clinical Evidence. Nanomaterials 2021, 11, 530. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, T.; Geng, T.; Wang, X.; Lin, K.; Wang, P. Dental Implants Loaded With Bioactive Agents Promote Osseointegration in Osteoporosis: A Review. Front. Bioeng. Biotechnol. 2021, 9, 591796. [Google Scholar] [CrossRef] [PubMed]
- Mohseni, M.; Hutmacher, D.W.; Castro, N.J. Independent Evaluation of Medical-Grade Bioresorbable Filaments for Fused Deposition Modelling/Fused Filament Fabrication of Tissue Engineered Constructs. Polymers 2018, 10, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngo, T.; Kashani, A.; Imbalzano, G.; Nguyen, K.; Hui, D. Additive manufacturing (3D printing): A review of materials, methods, applications and challenges. Compos. Part B-Eng. 2018, 143, 172–196. [Google Scholar] [CrossRef]
- Lee, H.; Yang, G.H.; Kim, M.; Lee, J.; Huh, J.; Kim, G. Fabrication of micro/nanoporous collagen/dECM/silk-fibroin biocomposite scaffolds using a low temperature 3D printing process for bone tissue regeneration. Mater. Sci. Eng. C 2018, 84, 140–147. [Google Scholar] [CrossRef]
- Kim, H.; Yang, G.H.; Choi, C.H.; Cho, Y.S.; Kim, G. Gelatin/PVA scaffolds fabricated using a 3D-printing process employed with a low-temperature plate for hard tissue regeneration: Fabrication and characterizations. Int. J. Biol. Macromol. 2018, 120, 119–127. [Google Scholar] [CrossRef]
- Song, X.; Tetik, H.; Jirakittsonthon, T.; Parandoush, P.; Yang, G.; Lee, D.; Ryu, S.; Lei, S.; Weiss, M.L.; Lin, D. Biomimetic 3D printing of hierarchical and interconnected porous hydroxyapatite structures with high mechanical strength for bone cell culture. Adv. Eng. Mater. 2019, 21, 1800678. [Google Scholar] [CrossRef] [Green Version]
- Lipp, L.; Sharma, D.; Banerjee, A.; Singh, J. Controlled delivery of Salmon calcitonin using thermosensitive triblock copolymer depot for treatment of osteoporosis. ACS Omega 2019, 4, 1157–1166. [Google Scholar] [CrossRef] [Green Version]
- Mora-Raimundo, P.; Lozano, D.; Manzano, M.; Vallet-Regi, M. Nanoparticles to Knockdown Osteoporosis-Related Gene and Promote Osteogenic Marker Expression for Osteoporosis Treatment. ACS Nano 2019, 13, 5451–5464. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Chen, G.; Liu, M.; Xu, Z.; Chen, H.; Yang, L.; Lv, Y. Scaffold strategies for modulating immune microenvironment during bone regeneration. Mater. Sci. Eng. C 2020, 108, 110411. [Google Scholar] [CrossRef]
- Wu, T.; Sun, J.; Tan, L.; Yan, Q.; Li, L.; Chen, L.; Liu, X.; Bin, S. Enhanced osteogenesis and therapy of osteoporosis using simvastatin loaded hybrid system. Bioact. Mater. 2020, 5, 348–357. [Google Scholar] [CrossRef]
- Rahman, S.U.; Nagrath, M.; Ponnusamy, S.; Arany, P.R. Nanoscale and Macroscale Scaffolds with Controlled-Release Polymeric Systems for Dental Craniomaxillofacial Tissue Engineering. Materials 2018, 11, 1478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Bachhuka, A.; Han, S.; Wei, F.; Lu, S.; Visalakshan, R.M.; Vasilev, K.; Xiao, Y. Tuning chemistry and topography of nanoengineered surfaces to manipulate immune response for bone regeneration applications. ACS Nano 2017, 11, 4494–4506. [Google Scholar] [CrossRef] [PubMed]
- Seidenstuecker, M.; Kerr, L.; Bernstein, A.; Mayr, H.O.; Suedkamp, N.P.; Gadow, R.; Krieg, P.; Hernandez Latorre, S.; Thomann, R.; Syrowatka, F.; et al. 3D Powder Printed Bioglass and beta-Tricalcium Phosphate Bone Scaffolds. Materials 2017, 11, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Deng, C.; Li, J.; Yao, Q.; Chang, J.; Wang, L.; Wu, C. 3D printing of a lithium-calcium-silicate crystal bioscaffold with dual bioactivities for osteochondral interface reconstruction. Biomaterials 2019, 196, 138–150. [Google Scholar] [CrossRef] [PubMed]
- Casarrubios, L.; Gómez-Cerezo, N.; Sánchez-Salcedo, S.; Feito, M.; Serrano, M.; Saiz-Pardo, M.; Ortega, L.; De Pablo, D.; Díaz-Güemes, I.; Fernández-Tomé, B. Silicon substituted hydroxyapatite/VEGF scaffolds stimulate bone regeneration in osteoporotic sheep. Acta Biomater. 2020, 101, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Sithole, M.N.; Kumar, P.; du Toit, L.C.; Marimuthu, T.; Choonara, Y.E.; Pillay, V. A 3D bioprinted in situ conjugated-co-fabricated scaffold for potential bone tissue engineering applications. J. Biomed. Mater. Res. A 2018, 106, 1311–1321. [Google Scholar] [CrossRef]
- Kosik-Kozioł, A.; Graham, E.; Jaroszewicz, J.; Chlanda, A.; Kumar, P.S.; Ivanovski, S.; Swieszkowski, W.; Vaquette, C. Surface modification of 3D printed polycaprolactone constructs via a solvent treatment: Impact on physical and osteogenic properties. ACS Biomater. Sci. Eng. 2018, 5, 318–328. [Google Scholar] [CrossRef]
- Grémare, A.; Guduric, V.; Bareille, R.; Heroguez, V.; Latour, S.; L’heureux, N.; Fricain, J.C.; Catros, S.; Le Nihouannen, D. Characterization of printed PLA scaffolds for bone tissue engineering. J. Biomed. Mater. Res. Part A 2018, 106, 887–894. [Google Scholar] [CrossRef]
- Liu, Y.; Yu, P.; Peng, X.; Huang, Q.; Ding, M.; Chen, Y.; Jin, R.; Xie, J.; Zhao, C.; Li, J. Hexapeptide-conjugated calcitonin for targeted therapy of osteoporosis. J. Control. Release 2019, 304, 39–50. [Google Scholar] [CrossRef]
- Gao, F.; Xu, Z.; Liang, Q.; Liu, B.; Li, H.; Wu, Y.; Zhang, Y.; Lin, Z.; Wu, M.; Ruan, C. Direct 3D printing of high strength biohybrid gradient hydrogel scaffolds for efficient repair of osteochondral defect. Adv. Funct. Mater. 2018, 28, 1706644. [Google Scholar] [CrossRef]
- Gao, F.; Jiao, C.; Yu, B.; Cong, H.; Shen, Y. Preparation and Biomedical Application of Injectable Hydrogels. Mater. Chem. Front. 2021, 5, 4912–4936. [Google Scholar] [CrossRef]
- Zeng, Y.; Zhou, M.; Mou, S.; Yang, J.; Yuan, Q.; Guo, L.; Zhong, A.; Wang, J.; Sun, J.; Wang, Z. Sustained delivery of alendronate by engineered collagen scaffold for the repair of osteoporotic bone defects and resistance to bone loss. J. Biomed. Mater. Res. A 2020, 108, 2460–2472. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Wufuer, M.; Kim, I.; Choi, T.H.; Kim, B.J.; Jung, H.G.; Jeon, B.; Lee, G.; Jeon, O.H.; Chang, H. Sequential dual-drug delivery of BMP-2 and alendronate from hydroxyapatite-collagen scaffolds for enhanced bone regeneration. Sci. Rep. 2021, 11, 746. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, R.; Pandey, R.; Kumar, S.; Mehrotra, D. Poly hydroxyalkanoates (PHA): Role in bone scaffolds. J. Oral Biol. Craniofacial Res. 2020, 10, 389–392. [Google Scholar] [CrossRef]
- Wei, H.; Xu, Y.; Wang, Y.; Xu, L.; Mo, C.; Li, L.; Shen, B.; Sun, Y.; Cheng, P.; Yang, L.; et al. Identification of Fibroblast Activation Protein as an Osteogenic Suppressor and Anti-osteoporosis Drug Target. Cell Rep. 2020, 33, 108252. [Google Scholar] [CrossRef]
- Starling, S. New anti-osteoporosis drug target identified. Nat. Rev. Endocrinol. 2021, 17, 4–5. [Google Scholar] [CrossRef]
- Kadaru, T.; Shibli-Rahhal, A. Zoledronic Acid after Treatment with Denosumab is Associated with Bone Loss within 1 Year. J. Bone Metab. 2021, 28, 51–58. [Google Scholar] [CrossRef]
- Rozenberg, S.; Al-Daghri, N.; Aubertin-Leheudre, M.; Brandi, M.L.; Cano, A.; Collins, P.; Cooper, C.; Genazzani, A.R.; Hillard, T.; Kanis, J.A.; et al. Is there a role for menopausal hormone therapy in the management of postmenopausal osteoporosis? Osteoporos. Int. 2020, 31, 2271–2286. [Google Scholar] [CrossRef]
- Pazianas, M.; Miller, P.D. Osteoporosis and Chronic Kidney Disease-Mineral and Bone Disorder (CKD-MBD): Back to Basics. Am. J. Kidney Dis. 2021, 78, 582–589. [Google Scholar] [CrossRef]
- Ottensmeyer, P.F.; Witzler, M.; Schulze, M.; Tobiasch, E. Small Molecules Enhance Scaffold-Based Bone Grafts via Purinergic Receptor Signaling in Stem Cells. Int. J. Mol. Sci. 2018, 19, 3601. [Google Scholar] [CrossRef] [Green Version]
- Witzler, M.; Ottensmeyer, P.F.; Gericke, M.; Heinze, T.; Tobiasch, E.; Schulze, M. Non-Cytotoxic Agarose/Hydroxyapatite Composite Scaffolds for Drug Release. Int. J. Mol. Sci. 2019, 20, 3565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witzler, M.; Buchner, D.; Shoushrah, S.H.; Babczyk, P.; Baranova, J.; Witzleben, S.; Tobiasch, E.; Schulze, M. Polysaccharide-Based Systems for Targeted Stem Cell Differentiation and Bone Regeneration. Biomolecules 2019, 9, 840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witzler, M.; Vermeeren, S.; Kolevatov, R.O.; Haddad, R.; Gericke, M.; Heinze, T.; Schulze, M. Evaluating Release Kinetics from Alginate Beads Coated with Polyelectrolyte Layers for Sustained Drug Delivery. ACS Appl. Bio Mater. 2021, 4, 6719–6731. [Google Scholar] [CrossRef] [PubMed]
- Ning, Z.; Tan, B.; Chen, B.; Lau, D.S.A.; Wong, T.M.; Sun, T.; Peng, S.; Li, Z.; Lu, W.W. Precisely Controlled Delivery of Abaloparatide through Injectable Hydrogel to Promote Bone Regeneration. Macromol. Biosci. 2019, 19, e1900020. [Google Scholar] [CrossRef] [PubMed]
- Pei, B.; Wang, W.; Dunne, N.; Li, X. Applications of Carbon Nanotubes in Bone Tissue Regeneration and Engineering: Superiority, Concerns, Current Advancements, and Prospects. Nanomaterials 2019, 9, 1501. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Chen, X.; Geng, S.; Wei, L.; Miron, R.J.; Zhao, Y.; Zhang, Y. Nanogel-based scaffolds fabricated for bone regeneration with mesoporous bioactive glass and strontium: In vitro and in vivo characterization. J. Biomed. Mater. Res. A 2017, 105, 1175–1183. [Google Scholar] [CrossRef]
- Wang, X.; Zeng, D.; Weng, W.; Huang, Q.; Zhang, X.; Wen, J.; Wu, J.; Jiang, X. Alendronate delivery on amino modified mesoporous bioactive glass scaffolds to enhance bone regeneration in osteoporosis rats. Artif. Cells Nanomed. Biotechnol. 2018, 46, 171–181. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.J.; Wang, F.Q.; Lu, C.B.; Zou, J.W.; Hu, J.B.; Yang, Z.; Sang, H.X.; Zhang, Y. Modulation of bone formation and resorption using a novel zoledronic acid loaded gelatin nanoparticles integrated porous titanium scaffold: An in vitro and in vivo study. Biomed. Mater. 2020, 15, 055013. [Google Scholar] [CrossRef] [Green Version]
- Qayoom, I.; Teotia, A.K.; Kumar, A. Nanohydroxyapatite Based Ceramic Carrier Promotes Bone Formation in a Femoral Neck Canal Defect in Osteoporotic Rats. Biomacromolecules 2020, 21, 328–337. [Google Scholar] [CrossRef]
- Fouad-Elhady, E.A.; Aglan, H.A.; Hassan, R.E.; Ahmed, H.H.; Sabry, G.M. Modulation of bone turnover aberration: A target for management of primary osteoporosis in experimental rat model. Heliyon 2020, 6, e03341. [Google Scholar] [CrossRef]
- Zhang, B.; Wang, L.; Song, P.; Pei, X.; Sun, H.; Wu, L.; Zhou, C.; Wang, K.; Fan, Y.; Zhang, X. 3D printed bone tissue regenerative PLA/HA scaffolds with comprehensive performance optimizations. Mater. Des. 2021, 201, 109490. [Google Scholar] [CrossRef]
- Kettenberger, U.; Luginbuehl, V.; Procter, P.; Pioletti, D.P. In vitro and in vivo investigation of bisphosphonate-loaded hydroxyapatite particles for peri-implant bone augmentation. J. Tissue Eng. Regen. Med. 2017, 11, 1974–1985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohl, F.; Schmitz, J.; Furtmann, N.; Schulz-Fincke, A.C.; Mertens, M.D.; Kuppers, J.; Benkhoff, M.; Tobiasch, E.; Bartz, U.; Bajorath, J.; et al. Design, characterization and cellular uptake studies of fluorescence-labeled prototypic cathepsin inhibitors. Org. Biomol. Chem. 2015, 13, 10310–10323. [Google Scholar] [CrossRef] [PubMed]
- Lindstrom, E.; Rizoska, B.; Henderson, I.; Terelius, Y.; Jerling, M.; Edenius, C.; Grabowska, U. Nonclinical and clinical pharmacological characterization of the potent and selective cathepsin K inhibitor MIV-711. J. Transl. Med. 2018, 16, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visetvichaporn, V.; Kim, K.H.; Jung, K.; Cho, Y.S.; Kim, D.D. Formulation of self-microemulsifying drug delivery system (SMEDDS) by D-optimal mixture design to enhance the oral bioavailability of a new cathepsin K inhibitor (HL235). Int. J. Pharm. 2020, 573, 118772. [Google Scholar] [CrossRef] [PubMed]
- Pathak, J.L.; Bravenboer, N.; Klein-Nulend, J. The Osteocyte as the New Discovery of Therapeutic Options in Rare Bone Diseases. Front. Endocrinol. 2020, 11, 405. [Google Scholar] [CrossRef]
- Rochefort, G.Y. The osteocyte as a therapeutic target in the treatment of osteoporosis. Ther. Adv. Musculoskelet. Dis. 2014, 6, 79–91. [Google Scholar] [CrossRef] [Green Version]
- Ghorbaninejad, M.; Khademi-Shirvan, M.; Hosseini, S.; Baghaban Eslaminejad, M. Epidrugs: Novel epigenetic regulators that open a new window for targeting osteoblast differentiation. Stem Cell Res. Ther. 2020, 11, 456. [Google Scholar] [CrossRef]
- Wang, P.; Perche, F.; Logeart-Avramoglou, D.; Pichon, C. RNA-based therapy for osteogenesis. Int. J. Pharm. 2019, 569, 118594. [Google Scholar] [CrossRef]
- Ultimo, S.; Zauli, G.; Martelli, A.M.; Vitale, M.; McCubrey, J.A.; Capitani, S.; Neri, L.M. Influence of physical exercise on microRNAs in skeletal muscle regeneration, aging and diseases. Oncotarget 2018, 9, 17220. [Google Scholar] [CrossRef] [Green Version]
- Cannataro, R.; Carbone, L.; Petro, J.L.; Cione, E.; Vargas, S.; Angulo, H.; Forero, D.A.; Odriozola-Martínez, A.; Kreider, R.B.; Bonilla, D.A. Sarcopenia: Etiology, Nutritional Approaches, and miRNAs. Int. J. Mol. Sci. 2021, 22, 9724. [Google Scholar] [CrossRef] [PubMed]
- Tsourdi, E.; Yu, E.W.; Jan de Beur, S.M.; Drake, M.T. Vaccination for COVID-19 and relationship to osteoporosis care: Current evidence and suggested approaches. J. Bone Miner. Res. 2021, 36, 1042–1047. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Zhang, Q.; Yin, Y.; Zhu, Y.; Wang, J.; Hou, Z.; Zhang, Y.; Chen, W. Epidemiologic characteristics of traumatic fractures during the outbreak of coronavirus disease 2019 (COVID-19) in China: A retrospective & comparative multi-center study. Injury 2020, 51, 1698–1704. [Google Scholar] [CrossRef] [PubMed]
- Polan, C.; Meyer, H.-L.; Burggraf, M.; Herten, M.; Beck, P.; Braitsch, H.; Becker, L.; Vogel, C.; Dudda, M.; Kauther, M.D. Geriatric Proximal Femur Fractures During the COVID-19 Pandemic-Fewer Cases, But More Comorbidities. Geriatr. Orthop. Surg. Rehabil. 2021, 12, 21514593211009657. [Google Scholar] [CrossRef] [PubMed]
- Fuggle, N.; Singer, A.; Gill, C.; Patel, A.; Medeiros, A.; Mlotek, A.S.; Pierroz, D.D.; Halbout, P.; Harvey, N.; Reginster, J.-Y. How has COVID-19 affected the treatment of osteoporosis? An IOF-NOF-ESCEO global survey. Osteoporos. Int. 2021, 32, 611–617. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Risk Factor Group | Risk Factor Subgroup | Effect/Influence | References |
|---|---|---|---|
| Hormone reduction | Estrogen/Estrone reduction | Reduction in OPG expression | [89] |
| Lack of ER-α-mediated suppression of RANKL expression | [90] | ||
| Lack of growth factor production such as IGFs or TGF-β | [91,92] | ||
| Lack of suppressive effect on Wnt-signaling antagonist sclerostin | [93] | ||
| IGF-1/2 reduction | Reduction in osteoblasts’ activation and differentiation | [117] | |
| Medical disorders and medication | Cancer/Breast cancer | Estrogen can influence breast cancer and treatment with anti-estrogen drugs can cause osteoporosis | [98,99] |
| Rheumatoid disorders | Glucocorticoid treatment in rheumatoid disorders increases the risk of osteoporosis | [100,101] | |
| Systemic inflammations leading to bone erosion due to a local effect of immune cells | [100] | ||
| Chronic kidney disease | Therapeutic drugs against osteoporosis can affect the renal function | [103] | |
| Chronic liver disease | Bilirubin and bile acids are retained factors of cholestasis and can decrease bone formation | [104] | |
| Diabetes mellitus | Hyperglycemia can be damaging to bone, since glucose can be toxic to osteoblasts | [105,106] | |
| Parkinson´s disease | Reduced mobility can cause reduced bone mass | [107] | |
| Multiple myeloma | Changes in the bone marrow microenvironment can lead to a dysregulation of bone turnover | [108] | |
| Poor nutrition/dietary factors | Low calcium intake | Calcium is an essential nutrient for bone growth, and a low intake reduces the bone density | [111] |
| Eating disorders/Anorexia nervosa | Low body weight can induce bone loss | [109] | |
| Gonadal function is decreased and can cause reduced bone mass | |||
| Metabolic disorders such as growth hormone resistance, low leptin concentrations and hypercortisolemia can induce bone loss | |||
| Lifestyle choices | Sedentary lifestyle | Reduced mobility can cause reduced bone mass | [112] |
| Excessive use of alcohol | Alcohol decreases the absorption of calcium and vitamin D | [113] | |
| Alcohol slows the bone turnover down | [114] | ||
| Use of tobacco | Indirect effect: the alteration of parathyroid hormone, adrenal hormones (leading to hypercortisolism), gonadal hormones, and increased oxidative stress | [113,115] | |
| Direct effect: binding of nicotine to its receptor on osteoblasts and inhibiting proliferation | [116] |
| Scaffold Type | Chemical Composition | Advantages | Disadvantages | References |
|---|---|---|---|---|
| Noble Metals | Titanium (Ti) and corresponding Ti alloys | Inertness, good biocompatibility, high mechanical strength; hydrophilic surface with reduced macrophage activation providing anti-inflammatory microenvironment improving osteogenesis | Strictly limited flexibility, no inherent bioactivity | [223,240] |
| Gold (Au) prepared as nanoparticles | Gold nanoparticles available in varying sizes (about 10–70 nm) with specific nano-topography that guides the cell attachment; gold with influence on expression of cytokines as well as different factors (e.g., osteogenic, fibrogenic, and angiogenic factors) | [238,241] | ||
| Minerals and Ceramics | Tricalcium phosphate (TCP); hydroxyapatite (HA) | Chemical composition with similarity to native bone tissue, high tensile strength | Low compressive strength and comprehensive modulus; limited option to change surface chemistry (number/nature of functional groups) | [235,242,243] |
| Sr–Ca–Si-doped, HA-based scaffolds | Doping enhances biomineralization capacity (compared to non-doped scaffolds) resulting in increased osteogenic activity | [219,239,244] | ||
| Hydrogels based on inorganic minerals and/or organic/polymeric materials | Poly(anhydride)s layers loaded with drugs (e.g., teriparatide) | Hydrogels possess a porous structure in micro- and nano-scale for tailored cell adhesion; option for surface modification/functionalization; high tensile strength, in situ formability and in situ drug delivery, injectability; high targeting and ability to allow uniform incorporation of therapeutic molecules and cells, resp., without need for further surgery; good biocompatibility and biodegradability resulting in fewer side effects, good cell colonization and proliferation | Low mechanical/compressive strength; need to combine hydrogels with other components in order to meet mechanical requirements; fast degradation (in case of poly(anhydrides) | [228] |
| Bioinspired mineral HA-based hydrogels as nanocomposite scaffolds for the promotion of osteogenic marker expression and the induction of bone regeneration in osteoporosis | [219,245] | |||
| Scaffolds based on synthetic polymers | Poly(caprolactone) (PCL) | Proven biocompatibility, option to tailor chemical composition and 3D structure of bilk polymers and surfaces to reach high porosity and tunable pore sizes | Low mechanical strength, premature degradation | [238,246] |
| Poly(vinyl alcohol) (PVA) modified with gelatin | [234] | |||
| Polyesters such as poly(lactic acid) (PLA), poly(glycolic acid) (PGA), and copolymers (PLA/PGA) | [231,247,248] | |||
| Poly(ethylene glycol diacrylate) (PEGDA) combined with laponite nanoclay (a mineral consisting of magnesium (Mg), lithium (Li), and silicon(Si)) | [249,250] | |||
| Polyetherketoneketone scaffold with a functionalized Sr-doped nano-HA coating | Biomimetically hierarchical structures; due to local release of Sr, enhancement of the intrinsic mechanical strength at microlevel resulting in improved bonding strength (of scaffold and host bone) | Advanced synthetic approach, hydrophobicity of the majority of synthetic polymers | [227] | |
| Scaffolds based on natural polymers | Polypeptides, such as collagen-based scaffolds (including different types I-V, focus on collagen type I); micro/nanoporous collagen modified with silk-fibroin | Collagens (combined with HA) enable the mimicking of chemical composition of natural bone | Weak mechanical strength requires a combination of collagen with minerals such as HA to mimic the chemical composition of bone | [251,252] |
| Polyesters such as poly(hydroxy alkanoates) (PHA), poly(hydroxy butyrates) (PHB), and poly(alginates) | Polyesters enable the mimicking of the natural tissue such as ECM; PHA show good biocompatibility and biodegradability, exhibit good tensile strength, thermoplasticity and elastomeric nature | Advanced preparation methods of PHA, PHB using enzymes/bacteria | [253] |
| Release Material | Chemical Composition | Encapsulated/Released Drug | Release Results | References |
|---|---|---|---|---|
| Inorganic/organic hybrid materials | Drug-functionalized HA combined with biodegradable collagen microspheres | Dual release of bisphosphonates (Alendronate, ALN) combined with BMP-2 | Initial release of BMP-2 for a few days, followed by sequential ALN release after two weeks; finally, increased osteogenic activity was observed due to synergistic effect of BMP-2/ALN and enhanced bone regeneration at eight weeks post-implantation (rat 8 mm critical-sized defect). | [252] |
| Thermo-sensitive triblock copolymer depot for treatment of osteoporosis | Salmon Calcitonin (sCT) | In vitro and in vivo studies using injectable depot materials doped with sCT to prevent osteoporosis side effects; moreover, the copolymeric release system maintained sCT in a conformationally stable form for the entire release process. | [236] | |
| Hydrogels based on pure polymers and/or hybrid materials | Methylacrylated gelatin | Abalo-paratide (analog of parathyroid hormone-related protein PTHrP 1-36) | Controlled release of Abaloparatide via injectable hydrogel—resulting in promoted pre-osteoblast differentiation and final bone regeneration. | [143,218,263] |
| PTH—hylaluronic acid hydrogel | Teriparatide (recombinant N-terminal fragment (rhPTH1-34) of the human parathyroid hormone) | Pulsatile drug delivery system, promoting angiogenesis. | [228] | |
| Collagen-based hydrogels | Sustained delivery of Alendronate | Improved repair of osteoporotic bone defects and resistance to bone loss. Kinetic studies confirmed a sustained ALN release, resulting in repair effects of collagen–ALN scaffolds for osteoporotic defects (5 mm cranial defects in ovary ectomized rats). | [251] | |
| Nanomaterials, e.g., nanoparticles, nanotubes, nanogels | HA-based nanoparticles | Bisphosphonates (ALN, ZOL) | Zn, Sr, and Ag incorporation for invigorating bone growth; partial Ca-substitution with cobalt favor osteogenesis process. | [229] |
| Titanium Sr/Ag loaded into Ti nanotubes | Strontium (Sr) and gold (Au) | Controlled drug release by varying nanotube diameter. | [229,264] | |
| Nanogel scaffolds consisting of bioactive glasses | Strontium (Sr) | Enhancement of osteoblast differentiation; enhanced bone regeneration in osteoporosis rats; targeted therapy of osteoporosis. | [229,265,266] | |
| Amino modified mesoporous bioactive glass (MBG) scaffolds | Alendronate (ALN) | |||
| Injectable nanogels consisting of triblock-copolymers | bone-seeking hexapeptide (Asp6)-conjugated sCT (sCT-Mal-Asp6) for the targeted treatment of osteoporosis | [248] | ||
| ZOL-loaded gelatin nanoparticles, integrated porous Ti scaffold implanted in a femoral defect | Zoledronate (ZOL) | [267] | ||
| Mesoporous hydroxyapatite (MHA) modified with poly (N-isopropylacrylamide) (PAA) brushes | Simvastatin (SIM) | Anti-osteoporotic effect of the SIM-loaded PAA/HA system studied in vivo on femur defect. | [239] | |
| Calcium sulfate/nano-HA-based nanocomposite carrier of BMP-2 and ZOL | BMP-2 and Zoledronate (ZOL) | BMSC-derived EXO, implanted in a femur defect (Sprague Dawley OVX rats) resulting in improved osteogenesis of the BMSCs. | [268] | |
| Nano-HA, nCh/HA, and nAg/HA delivered intravenously to female albino Wistar OVX rats | Alendronate (ALN) | [269] | ||
| Calcium citrate homogenized to nanoparticles (NPs), combined with PLA- and PLGA-based NPs | 17-beta-estradiol | Hormone replacement therapy; polyurethane nano-micelles deliver mRNA. | [229,270,271] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tonk, C.H.; Shoushrah, S.H.; Babczyk, P.; El Khaldi-Hansen, B.; Schulze, M.; Herten, M.; Tobiasch, E. Therapeutic Treatments for Osteoporosis—Which Combination of Pills Is the Best among the Bad? Int. J. Mol. Sci. 2022, 23, 1393. https://doi.org/10.3390/ijms23031393
Tonk CH, Shoushrah SH, Babczyk P, El Khaldi-Hansen B, Schulze M, Herten M, Tobiasch E. Therapeutic Treatments for Osteoporosis—Which Combination of Pills Is the Best among the Bad? International Journal of Molecular Sciences. 2022; 23(3):1393. https://doi.org/10.3390/ijms23031393
Chicago/Turabian StyleTonk, Christian Horst, Sarah Hani Shoushrah, Patrick Babczyk, Basma El Khaldi-Hansen, Margit Schulze, Monika Herten, and Edda Tobiasch. 2022. "Therapeutic Treatments for Osteoporosis—Which Combination of Pills Is the Best among the Bad?" International Journal of Molecular Sciences 23, no. 3: 1393. https://doi.org/10.3390/ijms23031393
APA StyleTonk, C. H., Shoushrah, S. H., Babczyk, P., El Khaldi-Hansen, B., Schulze, M., Herten, M., & Tobiasch, E. (2022). Therapeutic Treatments for Osteoporosis—Which Combination of Pills Is the Best among the Bad? International Journal of Molecular Sciences, 23(3), 1393. https://doi.org/10.3390/ijms23031393