
Endocrine-Disrupting Chemicals and Their Adverse Effects on the Endoplasmic Reticulum




Abstract
:1. Introduction
1.1. Brief Status of EDC
1.2. EDCs and Related Disease
2. ER Stress and EDCs
2.1. The Link between EDCs and ER Stress
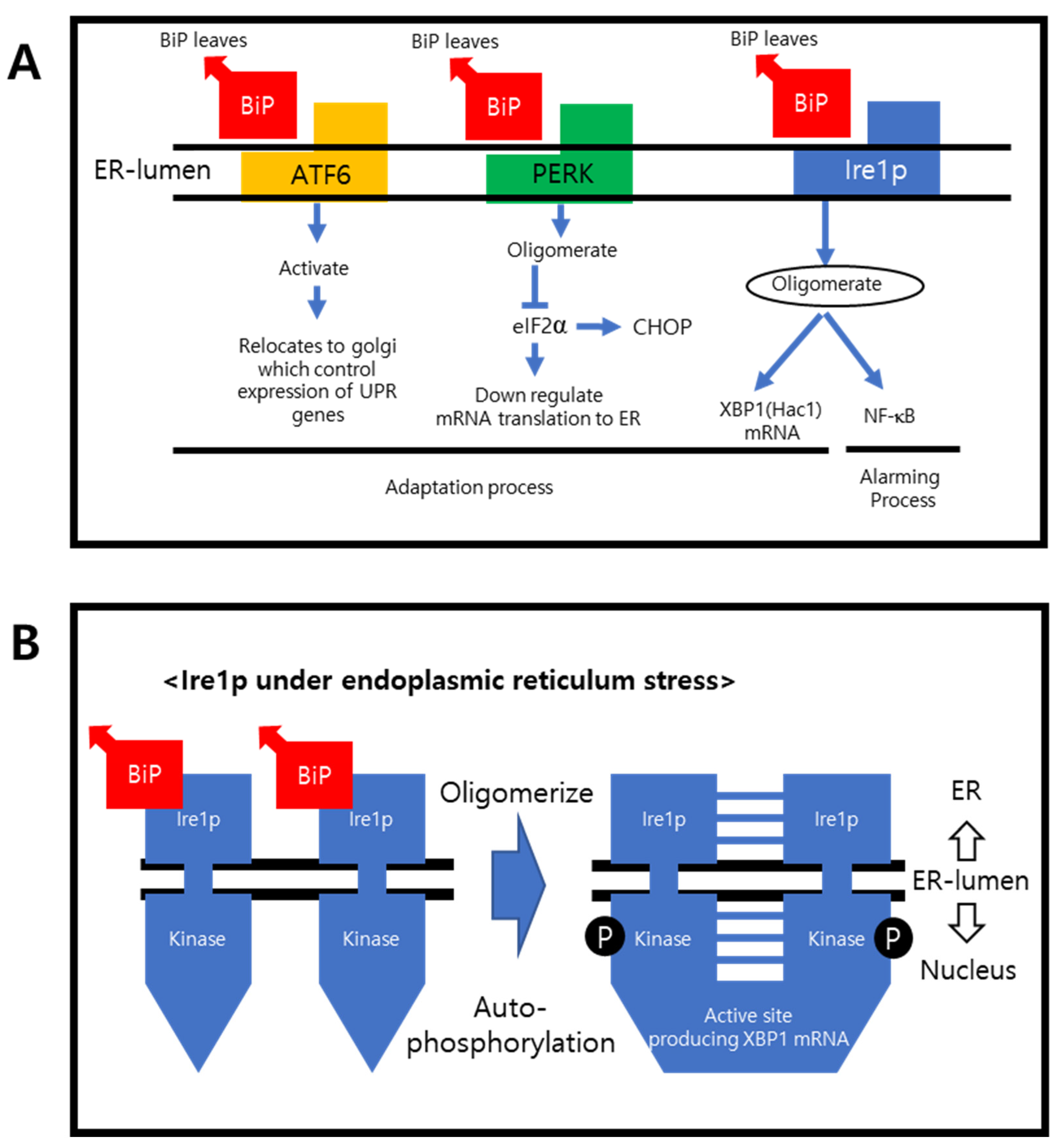
2.2. ER Stress
3. Specific Examples of EDCs Affecting the ER
3.1. Atrazine
3.2. Bisphenol A and Their Metabolites
3.3. Butylparaben
3.4. Chlorpyrifos
3.5. Dibutyl Phthalate
3.6. 2,4-Dichlorophenol
3.7. Dichloro Diphenyl Trichloro Ethane
3.8. Di (2-Ethylhexyl) Phthalate
3.9. Glyphosate
3.10. Lead
3.11. 4-Nonyl Phenol
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
References
- Colborn, T.; Clement, C. Wingspread consensus statement. In Chemically Induced Alterations in Sexual, Functional Development: The Wildlife/Human Connection; Princeton Scientific Publishing Princeton: Princeton, NJ, USA, 1992. [Google Scholar]
- Diamanti-Kandarakis, E.; Bourguignon, J.-P.; Giudice, L.C.; Hauser, R.; Prins, G.S.; Soto, A.M.; Zoeller, R.T.; Gore, A.C. Endocrine-Disrupting Chemicals: An Endocrine Society Scientific Statement. Endocr. Rev. 2009, 30, 293–342. [Google Scholar] [CrossRef] [PubMed]
- Lauretta, R.; Sansone, A.; Sansone, M.; Romanelli, F.; Appetecchia, M. Endocrine Disrupting Chemicals: Effects on Endocrine Glands. Front. Endocrinol. 2019, 10, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balaguer, P.; Delfosse, V.; Grimaldi, M.; Bourguet, W. Structural and functional evidences for the interactions between nuclear hormone receptors and endocrine dis-ruptors at low doses. Comptes Rendus Biol. 2017, 340, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Barouki, R. Endocrine disruptors: Revisiting concepts and dogma in toxicology. Comptes Rendus. Biol. 2017, 340, 410–413. [Google Scholar] [CrossRef] [PubMed]
- Taxvig, C.; Vinggaard, A.M.; Hass, U.; Axelstad, M.; Boberg, J.; Hansen, P.R.; Frederiksen, H.; Nellemann, C. Do parabens have the ability to interfere with steroidogenesis? Toxicol. Sci. 2008, 106, 206–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemini, C.; Hernández, A.; Jaimez, R.; Franco, Y.; Avila, M.E.; Castell, A. Morphometric analysis of mice uteri treated with the preservatives methyl, ethyl, propyl, and butylparaben. Toxicol. Ind. Health 2004, 20, 123–132. [Google Scholar] [CrossRef]
- Hu, Y.; Li, D.; Ma, X.; Liu, R.; Qi, Y.; Yuan, C.; Huang, D. Effects of 2,4-dichlorophenol exposure on zebrafish: Implications for the sex hormone synthesis. Aquat. Toxicol. 2021, 236, 105868. [Google Scholar] [CrossRef]
- Brodkin, M.A.; Madhoun, H.; Rameswaran, M.; Vatnick, I. Atrazine is an immune disruptor in adult northern leopard frogs (Rana pipiens). Environ. Toxicol. Chem. 2007, 26, 80–84. [Google Scholar] [CrossRef]
- Lind, M.P.; Lind, L. Circulating levels of bisphenol A and phthalates are related to carotid atherosclerosis in the elderly. Atherosclerosis 2011, 218, 207–213. [Google Scholar] [CrossRef]
- Kubo, K.; Arai, O.; Omura, M.; Watanabe, R.; Ogata, R.; Aou, S. Low dose effects of bisphenol A on sexual differentiation of the brain and behavior in rats. Neurosci. Res. 2003, 45, 345–356. [Google Scholar] [CrossRef]
- Tarazona, J.V.; Court-Marques, D.; Tiramani, M.; Reich, H.; Pfeil, R.; Istace, F.; Crivellente, F. Glyphosate toxicity and carcinogenicity: A review of the scientific basis of the European Union assessment and its differences with IARC. Arch. Toxicol. 2017, 91, 2723–2743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, M.; Bowser, D.; Costa, M. Carcinogenicity and genotoxicity of lead, beryllium, and other metals. Toxicol. Met. 1996, 253–284. [Google Scholar]
- Masuo, Y.; Ishido, M. Neurotoxicity of Endocrine Disruptors: Possible Involvement in Brain Development and Neurodegeneration. J. Toxicol. Environ. Heal. Part B 2011, 14, 346–369. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Park, S.M.; Jeong, S.; Kim, K.; Jeung, E.B. Combined Exposure to Diazinon and Nicotine Exerts a Synergistic Adverse Effect In Vitro and Disrupts Brain Development and Behaviors In Vivo. Int. J. Mol. Sci. 2021, 22, 7742. [Google Scholar] [CrossRef]
- Tran, D.N.; Park, S.-M.; Jung, E.-M.; Jeung, E.-B. Prenatal Octamethylcyclotetrasiloxane Exposure Impaired Proliferation of Neuronal Progenitor, Leading to Motor, Cognition, Social and Behavioral Functions. Int. J. Mol. Sci. 2021, 22, 12949. [Google Scholar] [CrossRef]
- Lang, I.A.; Galloway, T.S.; Scarlett, A.; Henley, W.E.; Depledge, M.; Wallace, R.B.; Melzer, D. Association of Urinary Bisphenol A Concentration with Medical Disorders and Laboratory Abnormalities in Adults. JAMA 2008, 300, 1303–1310. [Google Scholar] [CrossRef]
- Liu, Z.-W.; Zhu, H.-T.; Chen, K.-L.; Dong, X.; Wei, J.; Qiu, C.; Xue, J.-H. Protein kinase RNA- like endoplasmic reticulum kinase (PERK) signaling pathway plays a major role in reactive oxygen species (ROS)-mediated endoplasmic reticulum stress-induced apoptosis in diabetic cardiomyopathy. Cardiovasc. Diabetol. 2013, 12, 158. [Google Scholar] [CrossRef] [Green Version]
- Moncan, M.; Mnich, K.; Blomme, A.; Almanza, A.; Samali, A.; Gorman, A.M. Regulation of lipid metabolism by the unfolded protein response. J. Cell. Mol. Med. 2021, 25, 1359–1370. [Google Scholar] [CrossRef]
- Hotamisligil, G.S. Endoplasmic Reticulum Stress and the Inflammatory Basis of Metabolic Disease. Cell 2010, 140, 900–917. [Google Scholar] [CrossRef] [Green Version]
- Alberts, B.; Johnson, A.; Lewis, J.; Morgan, D.; Raff, M.; Roberts, K.; Walter, P. Molecular Biology of the Cell; WW Norton & Company: New York, NY, USA, 2017. [Google Scholar]
- Bonomini, F.; Rodella, L.F.; Rezzani, R. Metabolic Syndrome, Aging and Involvement of Oxidative Stress. Aging Dis. 2015, 6, 109–120. [Google Scholar] [CrossRef] [Green Version]
- Owen, C.R.; Kumar, R.; Zhang, P.; McGrath, B.C.; Cavener, D.R.; Krause, G.S. PERK is responsible for the increased phosphorylation of eIF2α and the severe inhibition of protein synthesis after transient global brain ischemia. J. Neurochem. 2005, 94, 1235–1242. [Google Scholar] [CrossRef] [PubMed]
- Meusser, B.; Hirsch, C.R.; Jarosch, E.; Sommer, T. ERAD: The long road to destruction. Nat. Cell Biol. 2005, 7, 766–772. [Google Scholar] [CrossRef] [PubMed]
- Hetz, C. The unfolded protein response: Controlling cell fate decisions under ER stress and beyond. Nat. Rev. Mol. Cell Biol. 2012, 13, 89–102. [Google Scholar] [CrossRef]
- Patil, C.; Walter, P. Intracellular signaling from the endoplasmic reticulum to the nucleus: The unfolded protein response in yeast and mammals. Curr. Opin. Cell Biol. 2001, 13, 349–355. [Google Scholar] [CrossRef]
- Calfon, M.; Zeng, H.; Urano, F.; Till, J.H.; Hubbard, S.R.; Harding, H.; Clark, S.G.; Ron, D. IRE1 couples endoplasmic reticulum load to secretory capacity by processing the XBP-1 mRNA. Nature 2002, 415, 92–96. [Google Scholar] [CrossRef]
- Kishino, A.; Hayashi, K.; Hidai, C.; Masuda, T.; Nomura, Y.; Oshima, T. XBP1-FoxO1 interaction regulates ER stress-induced autophagy in auditory cells. Sci. Rep. 2017, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Lemos, M.F.; Esteves, A.C.; Samyn, B.; Timperman, I.; van Beeumen, J.; Correia, A.; van Gestel, C.A.; Soares, A.M. Protein differential expression induced by endocrine disrupting compounds in a terrestrial isopod. Chemosphere 2010, 79, 570–576. [Google Scholar] [CrossRef]
- Du, Z.-H.; Qin, L.; Lin, J.; Sun, Y.-C.; Xia, J.; Zhang, C.; Li, X.-N.; Li, J.-L. Activating nuclear xenobiotic receptors and triggering ER stress and hepatic cytochromes P450 systems in quails (Coturnix C. coturnix) during atrazine exposure. Environ. Toxicol. 2017, 32, 1813–1822. [Google Scholar] [CrossRef]
- Sharma, S.; Sarkar, J.; Haldar, C.; Sinha, S. Melatonin reverses Fas, E2F-1 and endoplasmic reticulum stress mediated apoptosis and dysregulation of autophagy induced by the herbicide atrazine in murine splenocytes. PLoS ONE 2014, 9, e108602. [Google Scholar] [CrossRef]
- Lee, E.-J.; Jang, Y.; Kang, K.; Song, D.-H.; Kim, R.; Chang, H.-W.; Lee, D.E.; Song, C.K.-E.; Choi, B.; Kang, M.-J.; et al. Atrazine induces endoplasmic reticulum stress-mediated apoptosis of T lymphocytes via the caspase-8-dependent pathway. Environ. Toxicol. 2015, 31, 998–1008. [Google Scholar] [CrossRef]
- Tchounwou, P.B.; Wilson, B.A.; Ishaque, A.B.; Schneider, J. Atrazine potentiation of arsenic trioxide-induced cytotoxicity and gene expression in human liver carci-noma cells (HepG 2). In Molecular Mechanisms of Metal Toxicity and Carcinogenesis; Springer: Berlin/Heidelberg, Germany, 2001; pp. 49–59. [Google Scholar]
- Rochester, J.R. Bisphenol A and human health: A review of the literature. Reprod. Toxicol. 2013, 42, 132–155. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, L.S.; Oliveira, K.M.; Freitas, I.N.; Silva, J.A., Jr.; Silva, J.N.; Favero-Santos, B.C.; Bonfleur, M.L.; Carneiro, E.M.; Ribeiro, R.A. Bisphenol-A exposure worsens hepatic steatosis in ovariectomized mice fed on a high-fat diet: Role of endo-plasmic reticulum stress and fibrogenic pathways. Life Sci. 2020, 256, 118012. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Gómez-Toledano, R.; Sánchez-Esteban, S.; Cook, A.; Mínguez-Moratinos, M.; Ramírez-Carracedo, R.; Reventún, P.; Delgado-Marín, M.; Bosch, R.J.; Saura, M. Bisphenol A Induces Accelerated Cell Aging in Murine Endothelium. Biomolecules 2021, 11, 1429. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-F.; Liu, S.-H.; Su, C.-C.; Fang, K.-M.; Yen, C.-C.; Yang, C.-Y.; Tang, F.-C.; Liu, J.-M.; Wu, C.-C.; Lee, K.-I.; et al. Roles of ERK/Akt signals in mitochondria-dependent and endoplasmic reticulum stress-triggered neuronal cell apoptosis induced by 4-methyl-2,4-bis(4-hydroxyphenyl)pent-1-ene, a major active metabolite of bisphenol A. Toxicology 2021, 455, 152764. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Dai, Y.; Cui, Z.; Jiang, X.; Liu, W.; Han, F.; Lin, A.; Cao, J.; Liu, J. The regulation of cellular apoptosis by the ROS-triggered PERK/EIF2α/chop pathway plays a vital role in bisphenol A-induced male reproductive toxicity. Toxicol. Appl. Pharmacol. 2016, 314, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, S.V.; Huang, Y.; Snider, K.E.; Zhou, Y.; Pogash, T.J.; Russo, J. Expression and DNA methylation changes in human breast epithelial cells after bisphenol A exposure. Int. J. Oncol. 2012, 41, 369–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loganathan, N.; McIlwraith, E.K.; Belsham, D.D. Bisphenol A induces Agrp gene expression in hypothalamic neurons through a mechanism involving ATF3. Neuroendocrinology 2020. [Google Scholar] [CrossRef] [PubMed]
- Pu, Y.; Gingrich, J.D.; Steibel, J.P.; Veiga-Lopez, A. Sex-Specific Modulation of Fetal Adipogenesis by Gestational Bisphenol A and Bisphenol S Exposure. Endocrinology 2017, 158, 3844–3858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, G.-A.; Choi, K.-C.; Hwang, K.-A. Treatment with Phytoestrogens Reversed Triclosan and Bisphenol A-Induced Anti-Apoptosis in Breast Cancer Cells. Biomol. Ther. 2018, 26, 503–511. [Google Scholar] [CrossRef]
- Asahi, J.; Kamo, H.; Baba, R.; Doi, Y.; Yamashita, A.; Murakami, D.; Hanada, A.; Hirano, T. Bisphenol A induces endoplasmic reticulum stress-associated apoptosis in mouse non-parenchymal hepatocytes. Life Sci. 2010, 87, 431–438. [Google Scholar] [CrossRef]
- Luo, G.; Wei, R.; Wang, S.; Wang, J. Paternal bisphenol a diet changes prefrontal cortex proteome and provokes behavioral dysfunction in male off-spring. Chemosphere 2017, 184, 720–729. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.-H.; Wu, Y.-K.; Liao, B.-Y.; Zhang, H.; Li, C.; Wang, J.-L.; Hu, L.-L.; Ma, B. Bisphenol A Exposure Disrupts Organelle Distribution and Functions During Mouse Oocyte Maturation. Front. Cell Dev. Biol. 2021, 9, 656. [Google Scholar] [CrossRef] [PubMed]
- Kita, K.; Jin, Y.H.; Sun, Z.; Chen, S.P.; Sumiya, Y.; Hongo, T.; Suzuki, N. Increase in the levels of chaperone proteins by exposure to β-estradiol, bisphenol A and 4-methoxyphenol in human cells transfected with estrogen receptor α cDNA. Toxicol. Vitr. 2009, 23, 728–735. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Lim, W.; Bazer, F.W.; Song, G. Butyl paraben promotes apoptosis in human trophoblast cells through increased oxidative stress-induced endo-plasmic reticulum stress. Environ. Toxicol. 2018, 33, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Anderson, F.L.; von Herrmann, K.M.; Young, A.L.; Havrda, M.C. Bbc3 Loss Enhances Survival and Protein Clearance in Neurons Exposed to the Organophosphate Pesticide Chlorpyrifos. Toxicol. Sci. 2021, 183, 378–392. [Google Scholar] [CrossRef]
- Reyna, L.; Martín, J.B.F.; Ridano, M.E.; Panzetta-Dutari, G.M.; Genti-Raimondi, S. Chlorpyrifos induces endoplasmic reticulum stress in JEG-3 cells. Toxicol. Vitr. 2017, 40, 88–93. [Google Scholar] [CrossRef] [Green Version]
- Maradonna, F.; Carnevali, O. Lipid Metabolism Alteration by Endocrine Disruptors in Animal Models: An Overview. Front. Endocrinol. 2018, 9, 654. [Google Scholar] [CrossRef] [Green Version]
- Hatch, E.E.; Nelson, J.W.; Qureshi, M.M.; Weinberg, J.; Moore, L.L.; Singer, M.; Webster, T.F. Association of urinary phthalate metabolite concentrations with body mass index and waist circumference: A cross-sectional study of NHANES data, 1999–2002. Environ. Health 2008, 7, 27. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Ling, X.; Liu, K.; Wang, Z.; Zou, P.; Gao, J.; Ao, L. The p-eIF2α/ATF4 pathway links endoplasmic reticulum stress to autophagy following the production of reactive oxygen species in mouse spermatocyte-derived cells exposed to dibutyl phthalate. Free Radic. Res. 2016, 50, 698–707. [Google Scholar] [CrossRef]
- Zhang, G.; Liu, K.; Ling, X.; Wang, Z.; Zou, P.; Wang, X.; Gao, J.; Yin, L.; Zhang, X.; Liu, J.; et al. DBP-induced endoplasmic reticulum stress in male germ cells causes autophagy, which has a cytoprotective role against apoptosis in vitro and in vivo. Toxicol. Lett. 2016, 245, 86–98. [Google Scholar] [CrossRef]
- Zhang, G.; Yang, W.; Jiang, F.; Zou, P.; Zeng, Y.; Ling, X.; Ao, L. PERK regulates Nrf2/ARE antioxidant pathway against dibutyl phthalate-induced mitochondrial damage and apoptosis dependent of reactive oxygen species in mouse spermatocyte-derived cells. Toxicol. Lett. 2019, 308, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Li, J.; Qu, Z.; Qian, H.; Zhang, J.; Wang, H.; Xu, X.; Liu, S. Intrauterine exposure to low-dose DBP in the mice induces obesity in offspring via suppression of UCP1 mediated ER stress. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, X.; Qi, Y.; Huang, D.; Zhang, Y. 2,4-dichlorophenol induces ER stress-mediated apoptosis via e IF 2α dephosphorylation in vitro. Environ. Toxicol. 2016, 31, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Zhang, X.; Chen, P.; Zhang, Y. Endoplasmic reticulum stress is involved in 2,4-dichlorophenol-induced hepatotoxicity. J. Toxicol. Sci. 2016, 41, 745–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavlikova, N.; Sramek, J.; Jelinek, M.; Halada, P.; Kovar, J. Markers of acute toxicity of DDT exposure in pancreatic beta-cells determined by a proteomic approach. PLoS ONE 2020, 15, e0229430. [Google Scholar] [CrossRef] [PubMed]
- Stahlhut, R.W.; van Wijngaarden, E.; Dye, T.D.; Cook, S.; Swan, S.H. Concentrations of urinary phthalate metabolites are associated with increased waist circumference and insu-lin resistance in adult US males. Environ. Health Perspect. 2007, 115, 876–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Du, Z.-H.; Talukder, M.; Lin, J.; Li, X.-N.; Zhang, C.; Li, J.-L. Crosstalk between unfolded protein response and Nrf2-mediated antioxidant defense in Di-(2-ethylhexyl) phthalate-induced renal injury in quail (Coturnix japonica). Environ. Pollut. 2018, 242, 1871–1879. [Google Scholar] [CrossRef]
- Pan, J.; Yao, Y.; Guo, X.; Kong, F.; Zhou, J.; Meng, X. Endoplasmic reticulum stress, a novel significant mechanism responsible for DEHP-induced increased distance be-tween seminiferous tubule of mouse testis. J. Cell. Physiol. 2019, 234, 19807–19823. [Google Scholar] [CrossRef]
- Sun, X.; Lin, Y.; Huang, Q.; Shi, J.; Qiu, L.; Kang, M.; Chen, Y.; Fang, C.; Ye, T.; Dong, S. Di(2-ethylhexyl) phthalate-induced apoptosis in rat INS -1 cells is dependent on activation of endoplasmic reticulum stress and suppression of antioxidant protection. J. Cell. Mol. Med. 2014, 19, 581–594. [Google Scholar] [CrossRef]
- Amara, I.; Ontario, M.; Scuto, M.; Dico, G.L.; Sciuto, S.; Greco, V.; Abid-Essefi, S.; Signorile, A.; Salinaro, A.; Calabrese, V. Moringa oleifera Protects SH-SY5YCells from DEHP-Induced Endoplasmic Reticulum Stress and Apoptosis. Antioxidants 2021, 10, 532. [Google Scholar] [CrossRef]
- Peropadre, A.; Freire, P.F.; Martín, J.M.P.; Herrero, Ó.; Hazen, M.J. Endoplasmic reticulum stress as a novel cellular response to di (2-ethylhexyl) phthalate exposure. Toxicol. Vitr. 2015, 30, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Dallegrave, E.; Mantese, F.D.; Oliveira, R.T.; Andrade, A.J.M.; Dalsenter, P.R.; Langeloh, A. Pre- and postnatal toxicity of the commercial glyphosate formulation in Wistar rats. Arch. Toxicol. 2007, 81, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Yang, X.; Lu, J.; Xie, Q.; Ye, A.; Sun, W. The endoplasmic reticulum stress and related signal pathway mediated the glyphosate-induced testosterone synthesis inhibition in TM3 cells. Environ. Pollut. 2020, 260, 113949. [Google Scholar] [CrossRef] [PubMed]
- Gorell, J.; Johnson, C.C.; Rybicki, B.A.; Peterson, E.L.; Kortsha, G.X.; Brown, G.G.; Richardson, R.J. Occupational exposure to manganese, copper, lead, iron, mercury and zinc and the risk of Parkinson’s disease. Neurotoxicology 1999, 20, 239–247. [Google Scholar] [PubMed]
- Gao, K.; Zhang, C.; Tian, Y.; Naeem, S.; Zhang, Y.; Qi, Y. The role of endoplasmic reticulum stress in lead (Pb)-induced mitophagy of HEK293 cells. Toxicol. Ind. Health 2020, 36, 1002–1009. [Google Scholar] [CrossRef]
- Qian, Y.; Tiffany-Castiglioni, E. Lead-Induced Endoplasmic Reticulum (ER) Stress Responses in the Nervous System. Neurochem. Res. 2003, 28, 153–162. [Google Scholar] [CrossRef]
- Lepretti, M.; Paolella, G.; Giordano, D.; Marabotti, A.; Gay, F.; Capaldo, A.; Esposito, C.; Caputo, I.; Lepretti, M.; Paolella, G.; et al. 4-Nonylphenol reduces cell viability and induces apoptosis and ER-stress in a human epithelial intestinal cell line. Toxicol. Vitr. 2015, 29, 1436–1444. [Google Scholar] [CrossRef]
- Li, S.; Jiang, Z.; Chai, W.; Xu, Y.; Wang, Y. Autophagy activation alleviates nonylphenol-induced apoptosis in cultured cortical neurons. Neurochem. Int. 2018, 122, 73–84. [Google Scholar] [CrossRef]
- Paolella, G.; Romanelli, A.M.; Martucciello, S.; Sposito, S.; Lepretti, M.; Esposito, C.; Caputo, I. The mechanism of cytotoxicity of 4-nonylphenol in a human hepatic cell line involves ER-stress, apoptosis, and mitochondrial dysfunction. J. Biochem. Mol. Toxicol. 2021, 35, e22780. [Google Scholar] [CrossRef]
- Sasaya, H.; Yasuzumi, K.; Maruoka, H.; Fujita, A.; Kato, Y.; Waki, T.; Shimoke, K.; Ikeuchi, T. Apoptosis-inducing activity of endocrine-disrupting chemicals in cultured PC12 cells. Adv. Biol. Chem. 2012, 2, 92–105. [Google Scholar] [CrossRef] [Green Version]
- Gong, Y.; Wu, J.; Huang, Y.; Shen, S.; Han, X. Nonylphenol induces apoptosis in rat testicular Sertoli cells via endoplasmic reticulum stress. Toxicol. Lett. 2009, 186, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Kusunoki, T.; Shimoke, K.; Komatsubara, S.; Kishi, S.; Ikeuchi, T. p-Nonylphenol induces endoplasmic reticulum stress-mediated apoptosis in neuronally differentiated PC12 cells. Neurosci. Lett. 2007, 431, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Hayes, T. More feedback on whether atrazine is a potent endocrine disruptor chemical. Environ. Sci. Technol. 2009, 43, 6115. [Google Scholar] [CrossRef] [PubMed]
- Laws, S.C.; Ferrell, J.M.; Stoker, T.; Cooper, R.L. Pubertal Development in Female Wistar Rats following Exposure to Propazine and Atrazine Biotransformation By-Products, Diamino-S-Chlorotriazine and Hydroxyatrazine. Toxicol. Sci. 2003, 76, 190–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La Merrill, M.A.; Vandenberg, L.N.; Smith, M.T.; Goodson, W.; Browne, P.; Patisaul, H.B.; Guyton, K.Z.; Kortenkamp, A.; Cogliano, V.J.; Woodruff, T.J.; et al. Consensus on the key characteristics of endocrine-disrupting chemicals as a basis for hazard identification. Nat. Rev. Endocrinol. 2019, 16, 45–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janesick, A.; Blumberg, B. Obesogens: An emerging threat to public health. Am. J. Obstet. Gynecol. 2016, 214, 559–565. [Google Scholar] [CrossRef] [Green Version]
- Kabuto, H.; Amakawa, M.; Shishibori, T. Exposure to bisphenol A during embryonic/fetal life and infancy increases oxidative inju-ry and causes underdevelopment of the brain and testis in mice. Life Sci. 2004, 74, 2931–2940. [Google Scholar] [CrossRef]
- Liu, S.-H.; Su, C.C.; Lee, K.I.; Chen, Y.W. Effects of bisphenol a metabolite 4-Methyl-2, 4-bis (4-hydroxyphenyl) pent-1-ene on lung function and type 2 pul-monary alveolar epithelial cell growth. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Van Winkle, L.S.; Murphy, S.R.; Boetticher, M.V.; Vandevoort, C.A. Fetal Exposure of Rhesus Macaques to Bisphenol A Alters Cellular Development of the Conducting Airway by Changing Epithelial Secretory Product Expression. Environ. Health Perspect. 2013, 121, 912–918. [Google Scholar] [CrossRef] [Green Version]
- Midoro-Horiuti, T.; Tiwari, R.; Watson, C.S.; Goldblum, R.M. Maternal Bisphenol A Exposure Promotes the Development of Experimental Asthma in Mouse Pups. Environ. Health Perspect. 2010, 118, 273–277. [Google Scholar] [CrossRef] [Green Version]
- Baker, M.E.; Chandsawangbhuwana, C. 3D models of MBP, a biologically active metabolite of bisphenol A, in human estrogen receptor α and estrogen receptor β. PLoS ONE 2012, 7, e46078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoshiba, K.; Nagai, A. Oxidative stress, cell death, and other damage to alveolar epithelial cells induced by cigarette smoke. Tob. Induc. Dis. 2003, 1, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Sun, L.; Hu, Y.; Jiao, J.; Hu, J. Inverse antagonist activities of parabens on human oestrogen-related receptor γ (ERRγ): In vitro and in silico studies. Toxicol. Appl. Pharmacol. 2013, 270, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Naughton, S.X.; Terry, A.V. Neurotoxicity in acute and repeated organophosphate exposure. Toxicology 2018, 408, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Rauh, V.; Arunajadai, S.; Horton, M.; Perera, F.; Hoepner, L.; Barr, D.B.; Whyatt, R. Seven-Year Neurodevelopmental Scores and Prenatal Exposure to Chlorpyrifos, a Common Agricultural Pesticide. Environ. Health Perspect. 2011, 119, 1196–1201. [Google Scholar] [CrossRef] [PubMed]
- Rauh, V.A.; Garfinkel, R.; Perera, F.P.; Andrews, H.F.; Hoepner, L.; Barr, D.B.; Whyatt, R.W. Impact of prenatal chlorpyrifos exposure on neurodevelopment in the first 3 years of life among inner-city children. Pediatrics 2006, 118, e1845–e1859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Cock, M.; van de Bor, M. Obesogenic effects of endocrine disruptors, what do we know from animal and human studies? Environ. Int. 2014, 70, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Feng, Q.; He, Q.; Huang, Y.; Zhang, Y.; Jiang, G.; Xu, Z. Sources, atmospheric transport and deposition mechanism of organochlorine pesticides in soils of the Tibetan Plateau. Sci. Total Environ. 2017, 577, 405–412. [Google Scholar] [CrossRef]
- Ploteau, S.; Antignac, J.-P.; Volteau, C.; Marchand, P.; Vénisseau, A.; Vacher, V.; Le Bizec, B. Distribution of persistent organic pollutants in serum, omental, and parietal adipose tissue of French women with deep infiltrating endometriosis and circulating versus stored ratio as new marker of exposure. Environ. Int. 2016, 97, 125–136. [Google Scholar] [CrossRef]
- McKee, R.H.; Butala, J.H.; David, R.M.; Gans, G. NTP center for the evaluation of risks to human reproduction reports on phthalates: Addressing the data gaps. Reprod. Toxicol. 2004, 18, 1–22. [Google Scholar] [CrossRef]
- Paluselli, A.; Fauvelle, V.; Schmidt, N.; Galgani, F.; Net, S.; Sempéré, R. Distribution of phthalates in Marseille Bay (NW Mediterranean Sea). Sci. Total Environ. 2018, 621, 578–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, D.-J.; Gao, B.; Zhu, Z.-J.; Liu, G.; Yang, B. [Correlation between di-2-ethylhexyl phthalate and idiopathic oligoasthenospermia]. Zhonghua Nan Ke Xue = Natl. J. Androl. 2014, 20, 702–705. [Google Scholar]
- Piepenbrink, M.S.; Hussain, I.; Marsh, J.A.; Dietert, R.R.; Waller, S.K.; Waller, L. Developmental Immunotoxicology of Di-(2-Ethylhexyl)phthalate (DEHP): Age-Based Assessment in the Female Rat. J. Immunotoxicol. 2005, 2, 21–31. [Google Scholar] [CrossRef] [Green Version]
- Guvenius, D.M.; Aronsson, A.; Ekman-Ordeberg, G.; Bergman, A.; Norén, K. Human prenatal and postnatal exposure to polybrominated diphenyl ethers, polychlorinated biphenyls, polychlorobiphenylols, and pentachlorophenol. Environ. Health Perspect. 2003, 111, 1235–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arcadi, F.; Costa, C.; Imperatore, C.; Marchese, A.; Rapisarda, A.; Salemi, M.; Trimarchi, G.; Costa, G. Oral Toxicity of Bis(2-Ethylhexyl) Phthalate During Pregnancy and Suckling in the Long–Evans Rat. Food Chem. Toxicol. 1998, 36, 963–970. [Google Scholar] [CrossRef]
- Doyle, T.J.; Bowman, J.L.; Windell, V.L.; McLean, D.J.; Kim, K.H. Transgenerational Effects of Di-(2-ethylhexyl) Phthalate on Testicular Germ Cell Associations and Spermatogonial Stem Cells in Mice1. Biol. Reprod. 2013, 88, 112. [Google Scholar] [CrossRef]
- Do, R.P.; Stahlhut, R.W.; Ponzi, D.; Vom Saal, F.S.; Taylor, J.A. Non-monotonic dose effects of in utero exposure to di (2-ethylhexyl) phthalate (DEHP) on testicular and serum testosterone and anogenital distance in male mouse fetuses. Reprod. Toxicol. 2012, 34, 614–621. [Google Scholar] [CrossRef] [Green Version]
- Grande, S.W.; Andrade, A.J.; Talsness, C.E.; Grote, K.; Golombiewski, A.; Sterner-Kock, A.; Chahoud, I. A dose–response study following in utero and lactational exposure to di-(2-ethylhexyl) phthalate (DEHP): Reproductive effects on adult female offspring rats. Toxicology 2007, 229, 114–122. [Google Scholar] [CrossRef]
- Jones, S.; Boisvert, A.; Francois, S.; Zhang, L.; Culty, M. In Utero Exposure to Di-(2-Ethylhexyl) Phthalate Induces Testicular Effects in Neonatal Rats That Are Antagonized by Genistein Cotreatment1. Biol. Reprod. 2015, 93, 92. [Google Scholar] [CrossRef]
- Svensson, K.; Hernández-Ramírez, R.U.; Burguete-García, A.; Cebrián, M.E.; Calafat, A.M.; Needham, L.L.; Claudio, L.; López-Carrillo, L. Phthalate exposure associated with self-reported diabetes among Mexican women. Environ. Res. 2011, 111, 792–796. [Google Scholar] [CrossRef] [Green Version]
- James-Todd, T.; Stahlhut, R.; Meeker, J.D.; Powell, S.G.; Hauser, R.; Huang, T.; Rich-Edwards, J. Urinary phthalate metabolite concentrations and diabetes among women in the National Health and Nutri-tion Examination Survey (NHANES) 2001–2008. Environ. Health Perspect. 2012, 120, 1307–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajesh, P.; Sathish, S.; Srinivasan, C.; Selvaraj, J.; Balasubramanian, K. Diethyl Hexyl Phthalate (DEHP) is associated with insulin resistance in adipose tissue of male rat: Protective role of antioxidant vitamins (C & E). J. Cell. Biochem. 2012, 114, 558–569. [Google Scholar] [CrossRef]
- Srinivasan, C.; Khan, A.I.; Balaji, V.; Selvaraj, J.; Balasubramanian, K. Diethyl hexyl phthalate-induced changes in insulin signaling molecules and the protective role of antioxidant vitamins in gastrocnemius muscle of adult male rat. Toxicol. Appl. Pharmacol. 2011, 257, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Dzhekova-Stojkova, S.; Bogdanska, J.; Stojkova, Z. Peroxisome Proliferators: Their Biological and Toxicological Effects. Clin. Chem. Lab. Med. (CCLM) 2001, 39. [Google Scholar] [CrossRef] [PubMed]
- Hohmeier, H.E.; Newgard, C.B. Cell lines derived from pancreatic islets. Mol. Cell. Endocrinol. 2004, 228, 121–128. [Google Scholar] [CrossRef]
- Kristensen, D.M.; Skalkam, M.L.; Audouze, K.; Lesné, L.; Desdoits-Lethimonier, C.; Frederiksen, H.; Brunak, S.; Skakkebæk, N.E.; Jégou, B.; Hansen, J.B.; et al. Many Putative Endocrine Disruptors Inhibit Prostaglandin Synthesis. Environ. Health Perspect. 2011, 119, 534–541. [Google Scholar] [CrossRef] [Green Version]
- Mesnage, R.; Renney, G.; Séralini, G.-E.; Ward, M.; Antoniou, M.N. Multiomics reveal non-alcoholic fatty liver disease in rats following chronic exposure to an ultra-low dose of Roundup herbicide. Sci. Rep. 2017, 7, 39328. [Google Scholar] [CrossRef] [Green Version]
- Cattani, D.; Cavalli, V.L.D.L.O.; Rieg, C.E.H.; Domingues, J.T.; Dal-Cim, T.; Tasca, C.I.; Silva, F.R.M.B.; Zamoner, A. Mechanisms underlying the neurotoxicity induced by glyphosate-based herbicide in immature rat hippocampus: Involvement of glutamate excitotoxicity. Toxicology 2014, 320, 34–45. [Google Scholar] [CrossRef] [Green Version]
- Gidlow, D. Lead toxicity. Occup. Med. 2004, 54, 76–81. [Google Scholar] [CrossRef] [Green Version]
- Hamasaki, M.; Furuta, N.; Matsuda, A.; Nezu, A.; Yamamoto, A.; Fujita, N.; Oomori, H.; Noda, T.; Haraguchi, T.; Hiraoka, Y.; et al. Autophagosomes form at ER–mitochondria contact sites. Nature 2013, 495, 389–393. [Google Scholar] [CrossRef]
- Acir, I.-H.; Guenther, K. Endocrine-disrupting metabolites of alkylphenol ethoxylates—A critical review of analytical methods, environmental occurrences, toxicity, and regulation. Sci. Total Environ. 2018, 635, 1530–1546. [Google Scholar] [CrossRef] [PubMed]
- Dobrzyńska, M.M. DNA damage in organs of female and male mice exposed to nonylphenol, as a single agent or in combination with ionizing irradiation: A comet assay study. Mutat. Res. Toxicol. Environ. Mutagen. 2014, 772, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhou, L.; Ni, Y.; Wang, A.; Hu, M.; Lin, Y.; Hong, C.; Wan, J.; Chen, B.; Fang, L.; et al. Nonylphenol induces pancreatic damage in rats through mitochondrial dysfunction and oxidative stress. Toxicol. Res. 2017, 6, 353–360. [Google Scholar] [CrossRef] [Green Version]
- Kourouma, A.; Keita, H.; Duan, P.; Quan, C.; Bilivogui, K.K.; Qi, S.; Yang, K. Effects of 4-nonylphenol on oxidant/antioxidant balance system inducing hepatic steatosis in male rat. Toxicol. Rep. 2015, 2, 1423–1433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magnifico, M.C.; Xhani, M.; Popov, M.; Saso, L.; Sarti, P.; Arese, M. Nonylphenol and Octylphenol Differently Affect Cell Redox Balance by Modulating the Nitric Oxide Signaling. Oxidative Med. Cell. Longev. 2018, 2018, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Chemical | Uses | Classification | Upregulating ER Stress Regulator | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| ATF6 | PERK | p-PERK | eIF2α | p-eIF2α | CHOP | Ire1α | GRP78 | |||
| Atrazine | Herbicide | Hepatotoxicity damage [30], potentially toxic to immune system [9] | [30,31] | [31] | [32] | [32] | [32] | [30,32,33] | ||
| Bisphenol A | Plastic, flame retardant | Adverse effect on spermatogenesis [34], hepatic fibrosis [35] | [35,36,37,38,39,40] | [36,37,38,41] | [37,38] | [37] | [37,38,42] | [36,37,38,40,42,43,44] | [36,41] | [29,36,37,38,43,44,45,46] |
| Butyl-paraben | Cosmetic, food, pharmaceutical product | Reproductive toxicant [6,7] | [47] | [47] | [47] | [47] | ||||
| Chloropyrifos | Pesticide | Alzheimer’s disease, Parkinson’s disease [48] | [49] | [48] | [49] | [49] | ||||
| Dibutyl phthalate (DBP) | Plasticizer | Lipid metabolism alteration [50,51] | [52,53] | [54] | [52,53] | [52,53,55] | [52,53,55] | |||
| 2,4-Dichlorophenol (2,4-DCP) | Herbicide | Endocrine disruptor [8] | [56] | [56,57] | [56,57] | [56,57] | ||||
| Dichloro-diphenyl Trichloro-ethane (DDT) | Insecticide | Pancreatic β cell damage [58] | [58] | [58] | ||||||
| Di(2-ethylhexyl)phthalate (DEHP) | Plasticizer | Type II diabetes [59] | [60,61] | [60,62] | [62] | [62] | [61,62] | [61,62,63,64] | [60,61] | [60,61,62,63,64] |
| Glyphosate | Herbicide | Reproducible toxicity [65] | [66] | [66] | [66] | [66] | [66] | |||
| Lead | Heavy metal | Possible carcinogen [13] Parkinson‘s disease [67] | [68] | [68] | [68] | [68,69] | ||||
| 4-Nonyl phenol | Surfactant Detergent Plasticizer | Intestine damage [70] | [71,72,73] | [70,71,72,73,74,75] | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, K.; Kwon, J.-S.; Ahn, C.; Jeung, E.-B. Endocrine-Disrupting Chemicals and Their Adverse Effects on the Endoplasmic Reticulum. Int. J. Mol. Sci. 2022, 23, 1581. https://doi.org/10.3390/ijms23031581
Kim K, Kwon J-S, Ahn C, Jeung E-B. Endocrine-Disrupting Chemicals and Their Adverse Effects on the Endoplasmic Reticulum. International Journal of Molecular Sciences. 2022; 23(3):1581. https://doi.org/10.3390/ijms23031581
Chicago/Turabian StyleKim, Kangmin, Jin-Sook Kwon, Changhwan Ahn, and Eui-Bae Jeung. 2022. "Endocrine-Disrupting Chemicals and Their Adverse Effects on the Endoplasmic Reticulum" International Journal of Molecular Sciences 23, no. 3: 1581. https://doi.org/10.3390/ijms23031581
APA StyleKim, K., Kwon, J. -S., Ahn, C., & Jeung, E. -B. (2022). Endocrine-Disrupting Chemicals and Their Adverse Effects on the Endoplasmic Reticulum. International Journal of Molecular Sciences, 23(3), 1581. https://doi.org/10.3390/ijms23031581