
Removal of Antimony in Wastewater by Antimony Tolerant Sulfate-Reducing Bacteria Isolated from Municipal Sludge

 , and
, and 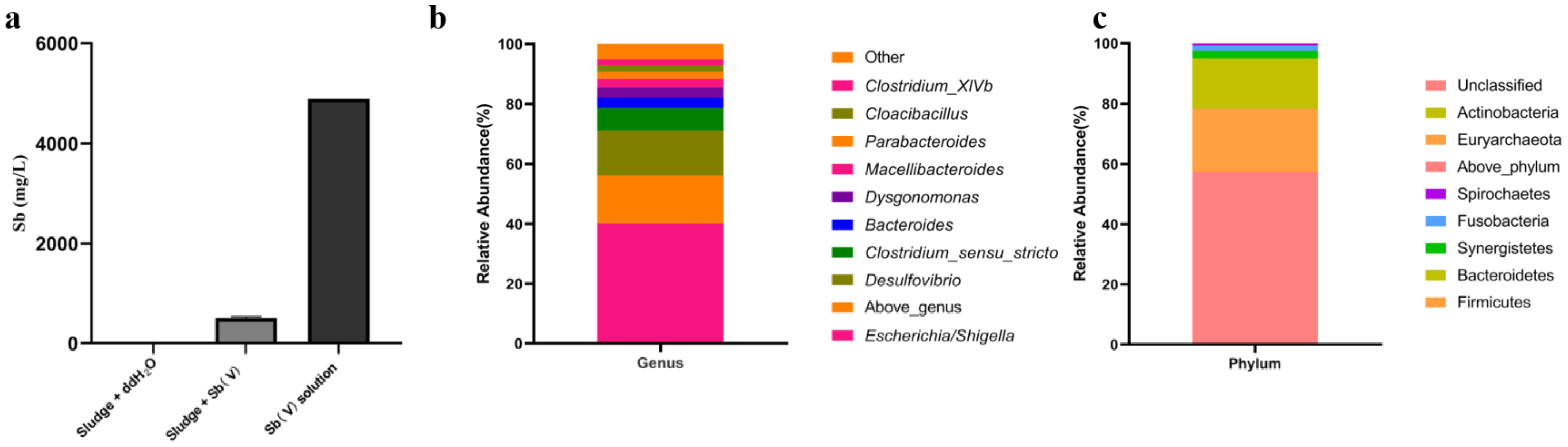{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Enrichment of SRB Community with Sb(V) Removal Activity and Its Community Analysis
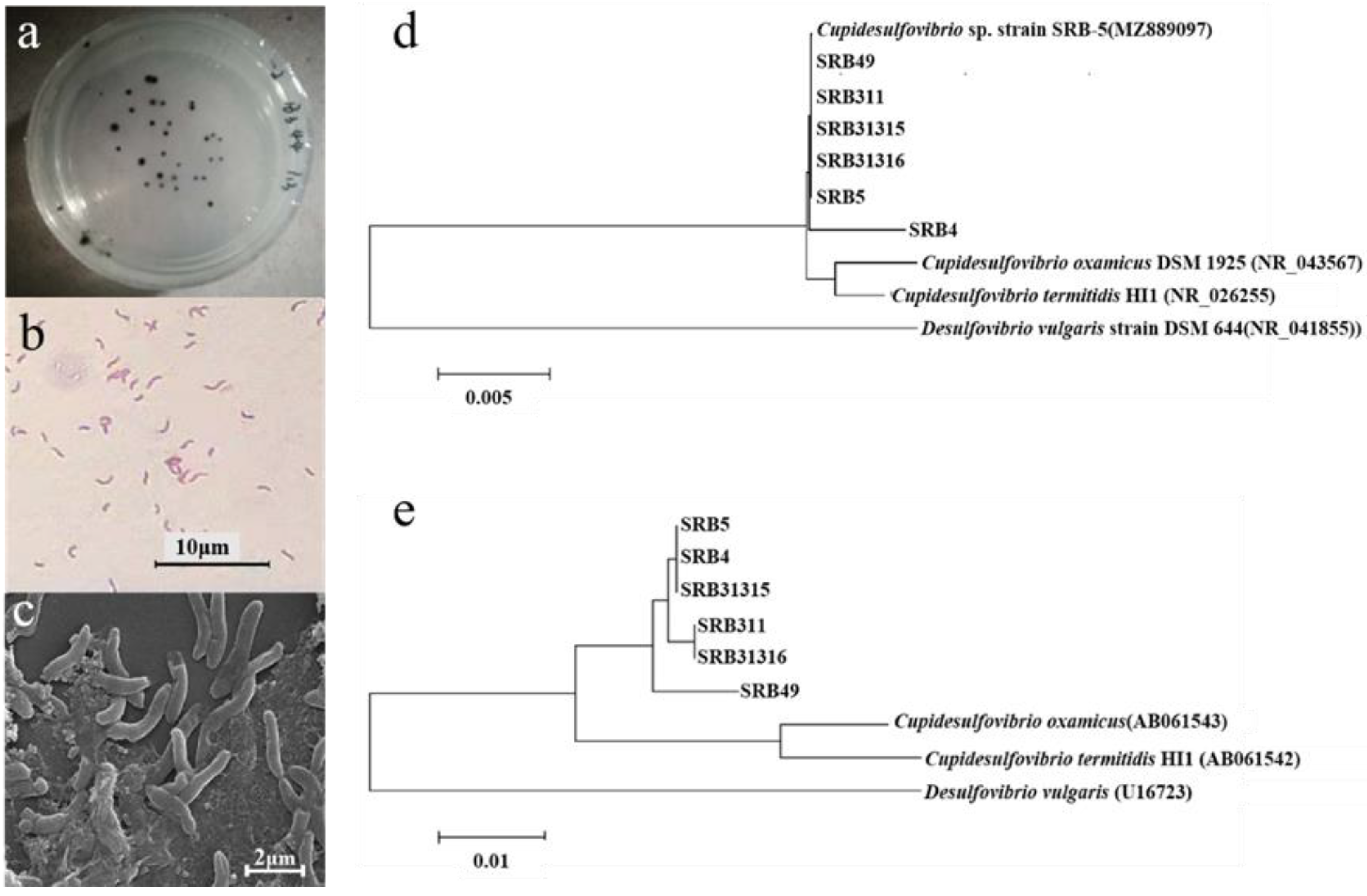
2.2. Isolation and Identification of Sb Tolerant SRB Strains
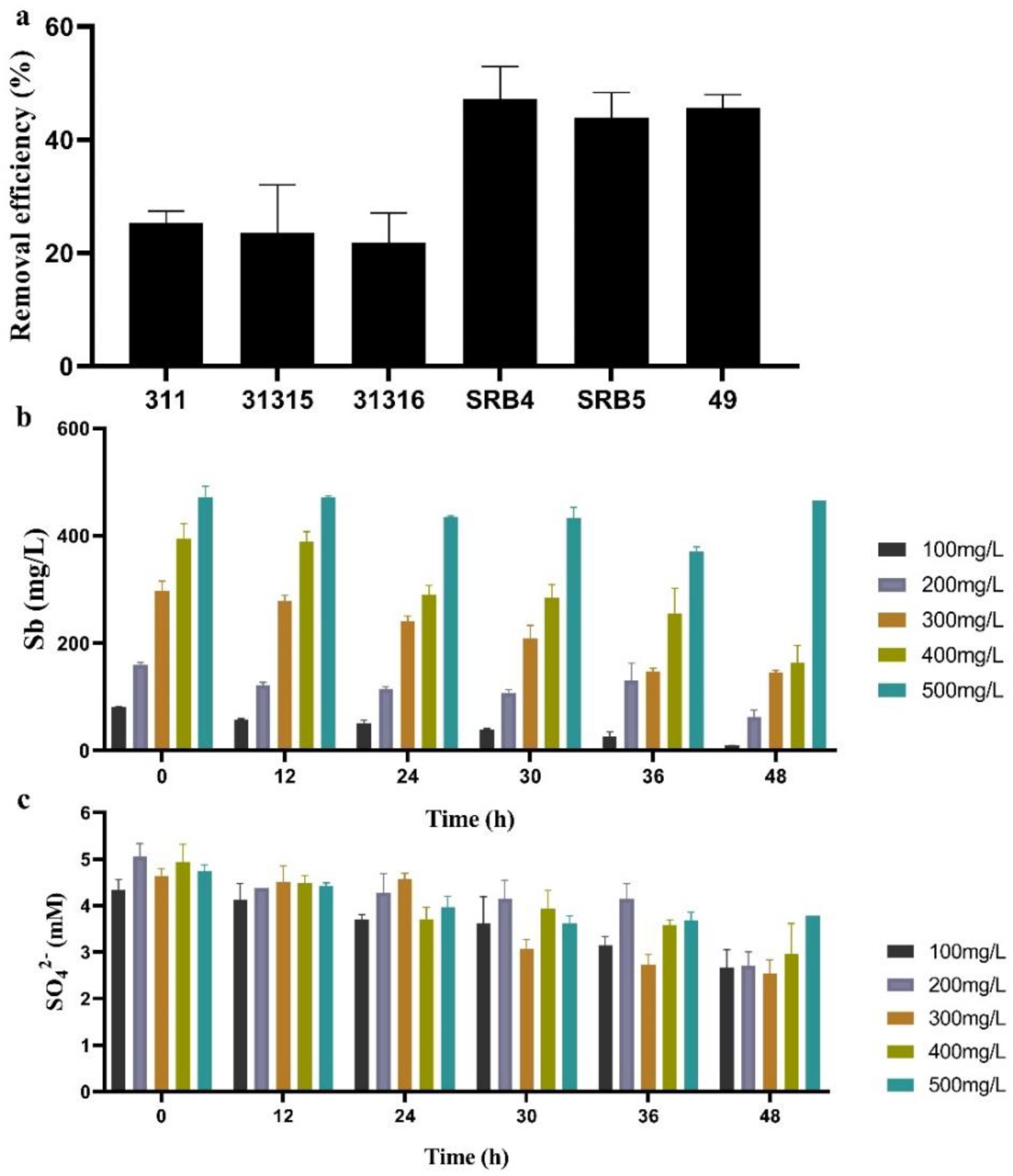
2.3. Sb(V) Removal by Pure Culture of SRB Strains
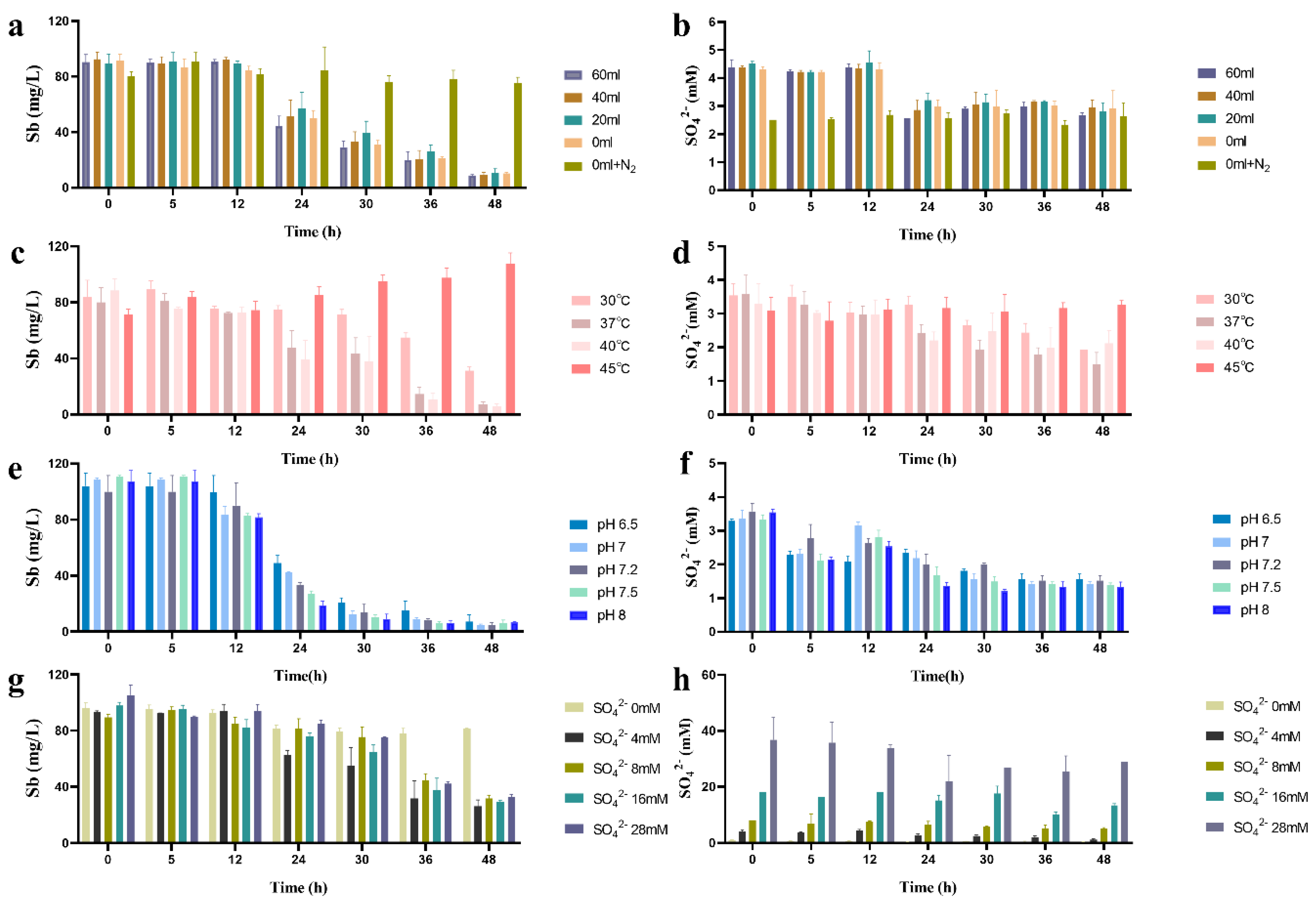
2.4. Effects of Different Culture Parameters on Sb(V) Removal by SRB49
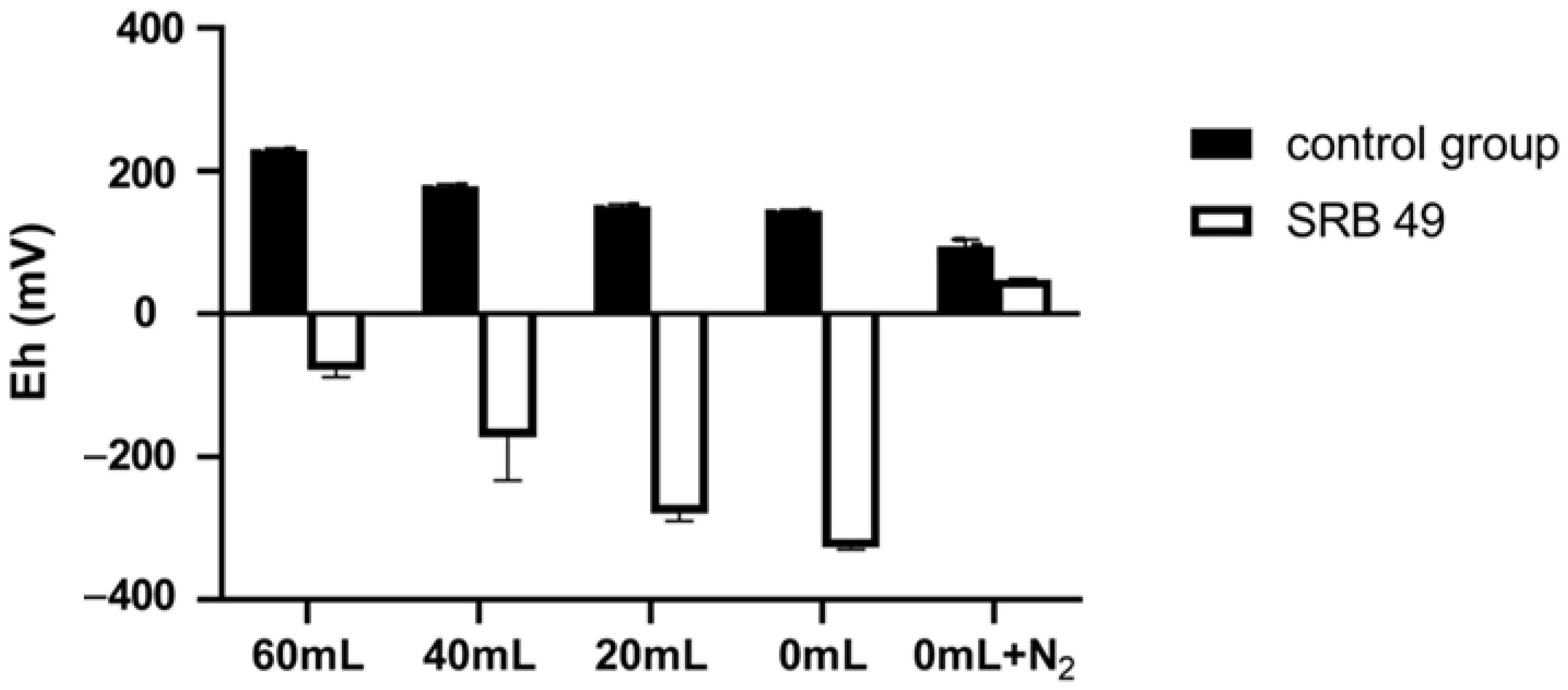
2.4.1. Effect of Redox Potential on the Removal of Sb(V)
2.4.2. Effect of Temperature, Initial pH, and Sulfate Concentration on the Removal of Sb(V)
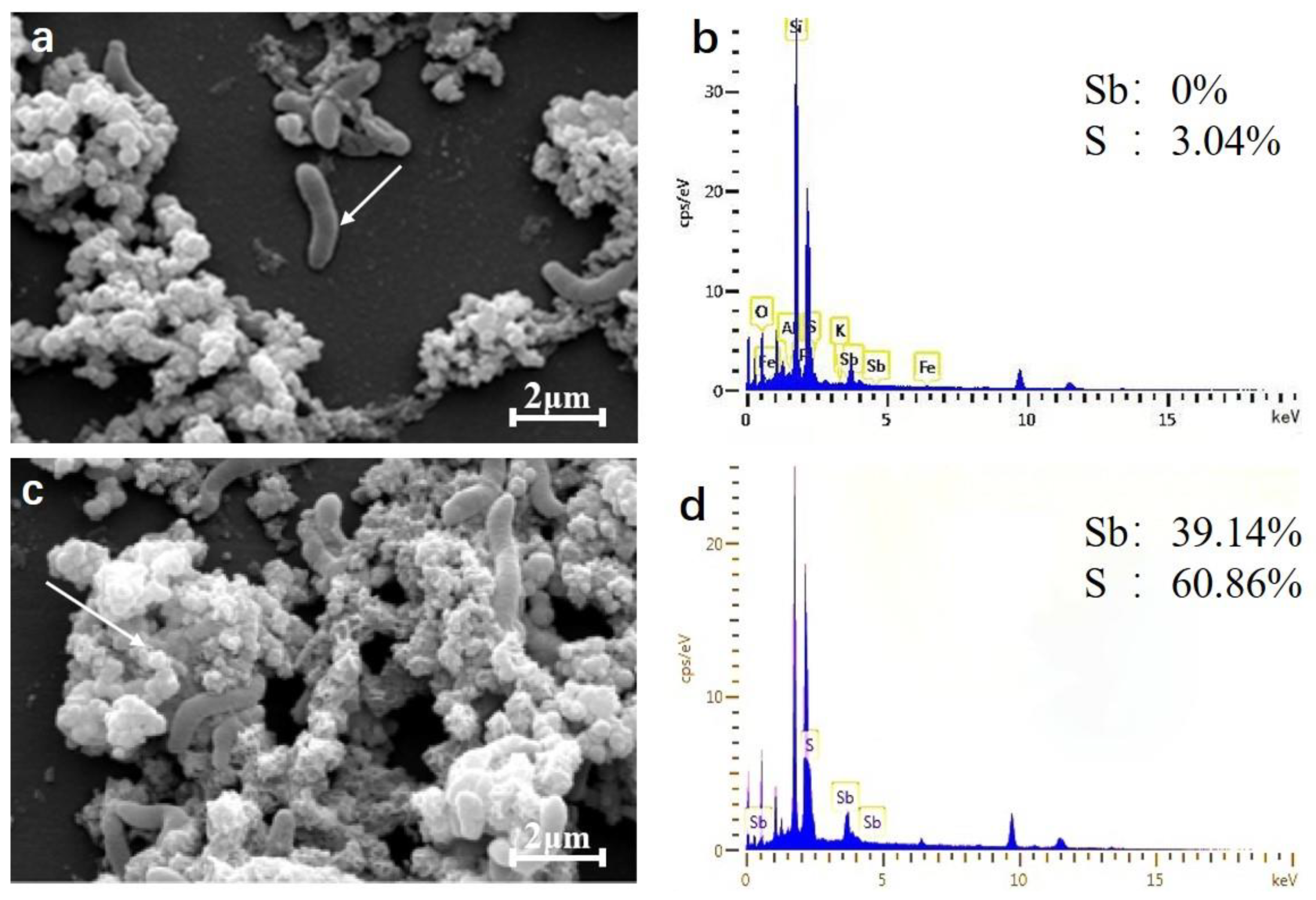
2.5. SEM-EDX Characterization Analysis
3. Discussion
3.1. Community Analysis of SRB Community Enrichment Culture
3.2. Sb Removal by Sb Tolerant SRB Strains
3.3. Influence of Different Culture Parameters on Sb(V) Removal of SRB49
4. Materials and Methods
4.1. Enrichment of Sb Tolerant Bacterial Community
4.2. Bacterial Community Analysis of SRB Enriched Culture by Illumina Sequencing of 16S rRNA Genes
4.3. Isolation and Molecular Identification of Sb Tolerant SRB Strains
4.4. Sb(V) Removal by SRB Enrichment and Pure Culture of SRB from Synthetic Wastewater
4.5. Effects of Different Culture Parameters on Sb(V) Removal by SRB Strain
4.6. Chemical Analysis and Statistical Analysis
4.7. SEM-EDX Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- He, M.; Wang, N.; Long, X.; Zhang, C.; Ma, C.; Zhong, Q.; Wang, A.; Wang, Y.; Pervaiz, A.; Shan, J. Antimony speciation in the environment: Recent advances in understanding the biogeochemical processes and ecological effects. J. Environ. Sci. (China) 2019, 75, 14–39. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Wang, X.; Wu, F.; Fu, Z. Antimony pollution in China. Sci. Total Environ. 2012, 421–422, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Ren, M.; Ding, S.; Fu, Z.; Yang, L.; Tang, W.; Tsang, D.C.W.; Wang, D.; Wang, Y. Seasonal antimony pollution caused by high mobility of antimony in sediments: In situ evidence and mechanical interpretation. J. Hazard. Mater. 2019, 367, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Nishad, P.A.; Bhaskarapillai, A. Antimony, a pollutant of emerging concern: A review on industrial sources and remediation technologies. Chemosphere 2021, 277, 130252. [Google Scholar] [CrossRef]
- Sundar, S.; Chakravarty, J. Antimony toxicity. Int. J. Environ Res. Public Health 2010, 7, 4267–4277. [Google Scholar] [CrossRef] [PubMed]
- Yamamura, S.; Iida, C.; Kobayashi, Y.; Watanabe, M.; Amachi, S. Production of two morphologically different antimony trioxides by a novel antimonate-reducing bacterium, Geobacter sp. SVR. J. Hazard. Mater. 2021, 411, 125100. [Google Scholar] [CrossRef] [PubMed]
- Xi, Y.; Lan, S.; Li, X.; Wu, Y.; Yuan, X.; Zhang, C.; Yunguo, L.; Huang, Y.; Quan, B.; Wu, S. Bioremediation of antimony from wastewater by sulfate-reducing bacteria: Effect of the coexisting ferrous ion. Int. Biodeterior. Biodegrad. 2020, 148, 104912. [Google Scholar] [CrossRef]
- Zhu, Y.; Wu, M.; Gao, N.; Chu, W.; An, N.; Wang, Q.; Wang, S. Removal of antimonate from wastewater by dissimilatory bacterial reduction: Role of the coexisting sulfate. J. Hazard. Mater. 2018, 341, 36–45. [Google Scholar] [CrossRef]
- Wang, H.; Chen, F.; Mu, S.; Zhang, D.; Pan, X.; Lee, D.-J.; Chang, J.-S. Removal of antimony (Sb (V)) from Sb mine drainage: Biological sulfate reduction and sulfide oxidation–precipitation. Bioresour. Technol. 2013, 146, 799–802. [Google Scholar] [CrossRef]
- Kulp, T.R.; Miller, L.G.; Braiotta, F.; Webb, S.M.; Kocar, B.D.; Blum, J.S.; Oremland, R.S. Microbiological Reduction of Sb(V) in Anoxic Freshwater Sediments. Environ. Sci. Technol. 2014, 48, 218–226. [Google Scholar] [CrossRef]
- Li, Y.C.; Xu, Z.; Wu, J.X.; Mo, P. Efficiency and mechanisms of antimony removal from wastewater using mixed cultures of iron-oxidizing bacteria and sulfate-reducing bacteria based on scrap iron. Sep. Purif. Technol. 2020, 246, 116756. [Google Scholar] [CrossRef]
- Wang, L.; Ye, L.; Yu, Y.; Jing, C. Antimony Redox Biotransformation in the Subsurface: Effect of Indigenous Sb(V) Respiring Microbiota. Environ. Sci. Technol. 2018, 52, 1200–1207. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Ouyang, X.; Li, H.; Fu, Z.; Chen, J. Bioremoval of antimony from contaminated waters by a mixed batch culture of sulfate-reducing bacteria. Int. Biodeterior. Biodegrad. 2016, 115, 148–155. [Google Scholar] [CrossRef]
- Xiao, E.Z.; Krumins, V.; Tang, S.; Xiao, T.F.; Ning, Z.P.; Lan, X.L.; Sun, W.M. Correlating microbial community profiles with geochemical conditions in a watershed heavily contaminated by an antimony tailing pond. Environ. Pollut. 2016, 215, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Baek, Y.-W.; An, Y.-J. Microbial toxicity of metal oxide nanoparticles (CuO, NiO, ZnO, and Sb2O3) to Escherichia coli, Bacillus subtilis, and Streptococcus aureus. Sci. Total Environ. 2011, 409, 1603–1608. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Chen, H.; Jing, C. Sulfate-Reducing Bacteria Mobilize Adsorbed Antimonate by Thioantimonate Formation. Environ. Sci. Technol. Lett. 2019, 6, 418–422. [Google Scholar] [CrossRef]
- Liu, F.; Zhang, G.; Liu, S.; Fu, Z.; Chen, J.; Ma, C. Bioremoval of arsenic and antimony from wastewater by a mixed culture of sulfate-reducing bacteria using lactate and ethanol as carbon sources. Int. Biodeterior. Biodegrad. 2018, 126, 152–159. [Google Scholar] [CrossRef]
- Abin, C.A.; Hollibaugh, J.T. Desulfuribacillus stibiiarsenatis sp. nov., an obligately anaerobic, dissimilatory antimonate- and arsenate-reducing bacterium isolated from anoxic sediments, and emended description of the genus Desulfuribacillus. Int. J. Syst. Evol. Microbiol. 2017, 67, 1011–1017. [Google Scholar] [CrossRef]
- Lai, C.Y.; Wen, L.L.; Zhang, Y.; Luo, S.S.; Wang, Q.Y.; Luo, Y.H.; Chen, R.; Yang, X.E.; Rittmann, B.E.; Zhao, H.P. Autotrophic antimonate bio-reduction using hydrogen as the electron donor. Water Res. 2016, 88, 467–474. [Google Scholar] [CrossRef]
- Muyzer, G.; Stams, A.J.M. The ecology and biotechnology of sulphate-reducing bacteria. Nat. Rev. Microbiol. 2008, 6, 441–454. [Google Scholar] [CrossRef]
- Li, X.; Xiao, H.; Zhang, W.; Li, Y.; Tang, X.; Duan, J.; Yang, Z.; Wang, J.; Guan, F.; Ding, G. Analysis of cultivable aerobic bacterial community composition and screening for facultative sulfate-reducing bacteria in marine corrosive steel. J. Oceanol. Limnol. 2019, 37, 600–614. [Google Scholar] [CrossRef]
- Chen, W.; Xiang, F.; Fu, J.; Wang, Q.; Wang, W.; Zeng, Q.; Yu, L. Identification and Phylogenetic Analysis of New Sulfate-Reducing Bacteria Isolated from Oilfield Samples. Z. Nat. C 2009, 64, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Mogensen, G.L.; Kjeldsen, K.U.; Ingvorsen, K. Desulfovibrio aerotolerans sp nov., an oxygen tolerant sulphatereducing bacterium isolated from activated sludge. Anaerobe 2005, 11, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Okkenhaug, G.; Zhu, Y.-G.; He, J.; Li, X.; Luo, L.; Mulder, J. Antimony (Sb) and Arsenic (As) in Sb Mining Impacted Paddy Soil from Xikuangshan, China: Differences in Mechanisms Controlling Soil Sequestration and Uptake in Rice. Environ. Sci. Technol. 2012, 46, 3155–3162. [Google Scholar] [CrossRef]
- Ňancucheo, I.; Rowe, O.F.; Hedrich, S.; Johnson, D.B. Solid and liquid media for isolating and cultivating acidophilic and acid-tolerant sulfate-reducing bacteria. FEMS Microbiol. Lett. 2016, 363, fnw083. [Google Scholar] [CrossRef] [Green Version]
- Geets, J.; Borrernans, B.; Diels, L.; Springael, D.; Vangronsveld, J.; van der Lelie, D.; Vanbroekhoven, K. DsrB gene-based DGGE for community and diversity surveys of sulfate-reducing bacteria. J. Microbiol. Methods 2006, 66, 194–205. [Google Scholar] [CrossRef]
- Wagner, M.; Roger, A.J.; Flax, J.L.; Brusseau, G.A.; Stahl, D.A. Phylogeny of dissimilatory sulfite reductases supports an early origin of sulfate respiration. J. Bacteriol. 1998, 180, 2975–2982. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Wu, S.S.; Zhao, L.H.; Ma, Q.L.; Li, X.; Ni, M.Y.; Zhou, T.; Zhu, H.L. Culture-dependent and-independent analysis of bacterial community structure in Jiangshui, a traditional Chinese fermented vegetable food. LWT-Food Sci. Technol. 2018, 96, 244–250. [Google Scholar]
- Althobiti, R.A.; Beauchemin, D. Pragmatic method based on on-line leaching and inductively coupled plasma mass spectrometry for risk assessment of the impact of short-term pollution. J. Anal. At. Spectrom. 2021, 36, 622–629. [Google Scholar] [CrossRef]
- Kolmert, A.; Wikstrom, P.; Hallberg, K.B. A fast and simple turbidimetric method for the determination of sulfate in sulfate-reducing bacterial cultures. J. Microbiol. Methods 2000, 41, 179–184. [Google Scholar] [CrossRef]
- Castillo, U.; Myers, S.; Browne, L.; Strobel, G.; Hess, W.M.; Hanks, J.; Reay, D. Scanning electron microscopy of some endophytic streptomycetes in Snakevine—Kennedia nigricans. Scanning 2005, 27, 305–311. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Fei, Y.; Xue, S.; Zhang, G.; Bian, Z.; Guo, F.; Wang, L.; Chai, R.; Zhang, S.; Cui, Z.; et al. Removal of Antimony in Wastewater by Antimony Tolerant Sulfate-Reducing Bacteria Isolated from Municipal Sludge. Int. J. Mol. Sci. 2022, 23, 1594. https://doi.org/10.3390/ijms23031594
Li H, Fei Y, Xue S, Zhang G, Bian Z, Guo F, Wang L, Chai R, Zhang S, Cui Z, et al. Removal of Antimony in Wastewater by Antimony Tolerant Sulfate-Reducing Bacteria Isolated from Municipal Sludge. International Journal of Molecular Sciences. 2022; 23(3):1594. https://doi.org/10.3390/ijms23031594
Chicago/Turabian StyleLi, He, Yue Fei, Shuwen Xue, Gege Zhang, Ziqi Bian, Fanfan Guo, Li Wang, Ruiqing Chai, Shuqi Zhang, Zhenyu Cui, and et al. 2022. "Removal of Antimony in Wastewater by Antimony Tolerant Sulfate-Reducing Bacteria Isolated from Municipal Sludge" International Journal of Molecular Sciences 23, no. 3: 1594. https://doi.org/10.3390/ijms23031594
APA StyleLi, H., Fei, Y., Xue, S., Zhang, G., Bian, Z., Guo, F., Wang, L., Chai, R., Zhang, S., Cui, Z., Wang, S., & Zhang, J. (2022). Removal of Antimony in Wastewater by Antimony Tolerant Sulfate-Reducing Bacteria Isolated from Municipal Sludge. International Journal of Molecular Sciences, 23(3), 1594. https://doi.org/10.3390/ijms23031594