
The iNKT Cell–Macrophage Axis in Homeostasis and Disease

 ,
,  and
and
Abstract
:1. Introduction
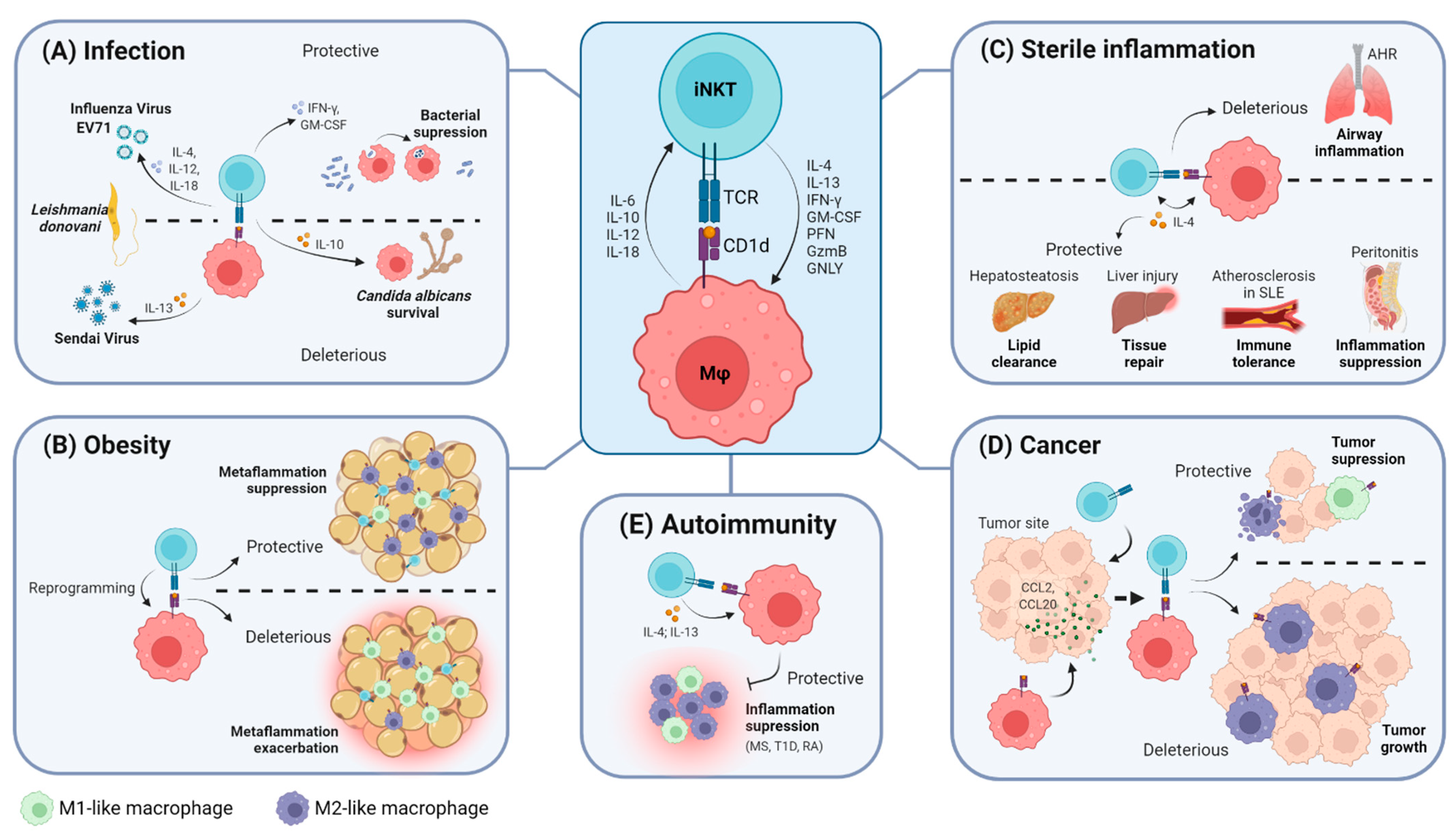
2. The iNKT Cell–Macrophage Axis
2.1. The iNKT Cell–Macrophage Axis in Homeostasis
2.2. The iNKT Cell–Macrophage Axis in Infection

{kind=link}
| Infection Model | Role | Readout | Ref. |
|---|---|---|---|
| CLP-induced sepsis | Protective | iNKT cells migrate from liver to peritoneum; iNKT cells increase macrophage phagocytic capacity. | [56] |
| CS-induced sepsis | Deleterious | PD-1-driven iNKT cells reduce macrophage activation. | [57] |
| CLP-induced sepsis | Deleterious | Lower numbers of hepatic iNKT cells; iNKT cells more activated; iNKT cells induce IL-6 secretion by hepatic macrophages. | [58] |
| Candida albicans | Deleterious | Activated iNKT cells correlated with lower survival rate; IL-10 production by iNKT cells. | [59] |
| Candida albicans post-sepsis | Deleterious | Impairment of macrophage phagocytic capacity by iNKT cells; Presence of iNKT cells associated with worst survival rate. | [60] |
| Leishmania donovani | Protective | Enhancement of hepatic iNKT cell activation in the presence of infected KCs; SIPRα-CD47 interaction enhances iNKT activation. | [61] |
| Leishmania donovani | Deleterious | Impairment of iNKT cell activation by infected splenic macrophages; GSPL-CD1d activates iNKT cells. | [62] |
| GSPL-TLR4 activates macrophages. | [63] | ||
| Borrelia burgdorferi | Protective | KCs induce CXCR3-mediated iNKT cell clustering; iNKT cells reduced bacteria dissemination. | [64] |
| Listeria monocytogenes | Protective | iNKT cells and macrophages ameliorate listeriosis; α-GalCer-driven activation of iNKT cells enhances bactericidal functions of peritoneal macrophages. | [65] |
| Mycobacterium tuberculosis | Protective | Monocytes pulsed with α-GalCer lead iNKT cells to restrict Mtb growth. | [66] |
| Mycobacterium tuberculosis | Protective | iNKT cells can be activated by Mtb-infected macrophages via CD1d, IL-12 and IL-18; suppression of bacterial replication. | [67] |
| Mycobacterium tuberculosis | Protective | CD1d-dependent suppression of bacterial growth; iNKT cells secrete GM-CSF in the presence of Mtb-infected macrophages; GM-CSF is sufficient to control Mtb growth. | [68] |
| Pseudomonas aeruginosa | Protective | α-GalCer-driven activation of iNKT cells enhances phagocytosis by alveolar macrophages. | [69] |
| Influenza virus | Protective | iNKT cell activation induced by α-GalCer enhances early innate immune response; possible migration of hepatic iNKT cells to the lungs. | [71] |
| Influenza virus | Protective | iNKT cells were associated with better survival rate in infected mice. | [72] |
| Influenza virus | Protective | NKT activation results in an early wave of IL-4 secretion upon their relocation and priming of nearby resident macrophages; induction of IL-4 requires both CD1d and IL-18. | [73] |
| Sendai Virus | Deleterious | Inflammation is potentiated by IL-13 from iNKT-activated macrophages. | [74] |
| Streptococcus pneumoniae | Protective | Mrp1 deficiencies in macrophages:
| [70] |
| Bacillus anthracis | Protective | α-GalCer administration delays bacterial systemic dissemination and increases survival rate; depletion of CD169+ macrophages reduces iNKT activation and increases bacterial dissemination. | [43] |
| Enterovirus 71 | Protective | iNKT cells increase survival rate; TLR3-dependent activation of iNKT cells by macrophages; iNKT activation also requires CD1d and IL-12. | [75] |
| Brucella suis | Protective | CD4+ iNKT cells impair bacterial intramacrophagic growth. | [76] |
2.3. The iNKT Cell–Macrophage Axis in Obesity
| Model | Role | Readout | Ref. |
|---|---|---|---|
| 60% HFD (4 days or 8 or 24 weeks) | Protective | M2 polarization of adipose tissue macrophages via IL-4 by activated iNKT cells. | [77,78] |
| 60% HFD (6 or 12 weeks) | Protective | Inverse correlation between numbers of adipose tissue iNKT cells and pro-inflammatory macrophages. | [79] |
| HFD (1 or 3 days and 1, 4 or 12 weeks) | Deleterious | iNKT-mediated recruitment of pro-inflammatory macrophages into adipose tissue. | [83] |
| 60% HFD (6, 8, 10 or 16 weeks) | Deleterious | iNKT activation by M1 macrophages exacerbated metaflammation and activation by M2-macrophage-ameliorated disease; M2-specific CD1d downregulation during obesity progression. | [81] |
2.4. The iNKT Cell–Macrophage Axis in Sterile Inflammation
2.5. The iNKT Cell–Macrophage Axis in Cancer
2.6. The iNKT Cell–Macrophage Axis in Autoimmunity
3. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bendelac, A.; Savage, P.B.; Teyton, L. The Biology of NKT Cells. Annu. Rev. Immunol. 2007, 25, 297–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godfrey, D.I.; Stankovic, S.; Baxter, A.G. Raising the NKT Cell Family. Nat. Immunol. 2010, 11, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.S.; Ribeiro, H.; Macedo, M.F. From Lysosomal Storage Diseases to NKT Cell Activation and Back. Int. J. Mol. Sci. 2017, 18, 502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porcelli, S.; Yockey, C.E.; Brenner, M.B.; Balk, S.P. Analysis of T Cell Antigen Receptor (TCR) Expression by Human Peripheral Blood CD4−8− α/β T Cells Demonstrates Preferential Use of Several Vβ Genes and an Invariant TCR α Chain. J. Exp. Med. 1993, 178, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lantz, O.; Bendelac, A. An Invariant T Cell Receptor α Chain Is Used by a Unique Subset of Major Histocompatibility Complex Class I-Specific CD4+ and CD4-8- T Cells in Mice and Humans. J. Exp. Med. 1994, 180, 1077–1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dellabona, B.P.; Padovan, E.; Casorati, G.; Brockhaus, M.; Lanzavecchia, A. An invariant Vα24-JαQ/Vβ11 T Cell Receptor Is Expressed in All Individuals by Clonally Expanded CD4-8- T cells. J. Exp. Med. 1994, 180, 1171–1175. [Google Scholar] [CrossRef] [PubMed]
- Watarai, H.; Nakagawa, R.; Omori-Miyake, M.; Dashtsoodol, N.; Taniguchi, M. Methods for Detection, Isolation and Culture of Mouse and Human Invariant NKT Cells. Nat. Protoc. 2008, 3, 70–78. [Google Scholar] [CrossRef]
- Exley, M.A.; Hou, R.; Shaulov, A.; Tonti, E.; Dellabona, P.; Casorati, G.; Akbari, O.; Akman, H.O.; Greenfield, E.A.; Gumperz, J.E.; et al. Selective Activation, Expansion, and Monitoring of Human INKT Cells with a Monoclonal Antibody Specific for the TCR α-Chain CDR3 Loop. Eur. J. Immunol. 2008, 38, 1756–1766. [Google Scholar] [CrossRef]
- Montoya, C.J.; Pollard, D.; Martinson, J.; Kumari, K.; Wasserfall, C.; Mulder, C.B.; Rugeles, M.T.; Atkinson, M.A.; Landay, A.L.; Wilson, S.B. Characterization of Human Invariant Natural Killer T Subsets in Health and Disease Using a Novel Invariant Natural Killer T Cell-Clonotypic Monoclonal Antibody, 6B11. Immunology 2007, 122, 1–14. [Google Scholar] [CrossRef]
- Crosby, C.M.; Kronenberg, M. Tissue-Specific Functions of Invariant Natural Killer T Cells. Nat. Rev. Immunol. 2018, 18, 559–574. [Google Scholar] [CrossRef]
- Pellicci, D.G.; Koay, H.F.; Berzins, S.P. Thymic Development of Unconventional T Cells: How NKT Cells, MAIT Cells and Γδ T Cells Emerge. Nat. Rev. Immunol. 2020, 20, 756–770. [Google Scholar] [CrossRef]
- Wolf, B.J.; Choi, J.E.; Exley, M.A. Novel Approaches to Exploiting Invariant NKT Cells in Cancer Immunotherapy. Front. Immunol. 2018, 9, 384. [Google Scholar] [CrossRef] [PubMed]
- Mori, L.; Lepore, M.; De Libero, G. The Immunology of CD1- and MR1-Restricted T Cells. Annu. Rev. Immunol. 2016, 34, 479–510. [Google Scholar] [CrossRef] [PubMed]
- Leite-de-Moraes, M.C.; Hameg, A.; Machavoine, F.; Koezuka, Y.; Herbelin, A.; Dy, M.; Schneider, E. A Distinct IL-18-Induced Pathway to Fully Activate NK T Lymphocytes Independently from TCR Engagement. J. Immunol. 1999, 163, 5871–5876. [Google Scholar] [PubMed]
- Kitamura, H.; Iwakabe, K.; Yahata, T.; Nishimura, S.I.; Ohta, A.; Ohmi, Y.; Sato, M.; Takeda, K.; Okumura, K.; Van Kaer, L.; et al. The Natural Killer T (NKT) Cell Ligand α-Galactosylceramide Demonstrates Its Immunopotentiating Effect by Inducing Interleukin (IL)-12 Production by Dendritic Cells and IL-12 Receptor Expression on NKT Cells. J. Exp. Med. 1999, 189, 1121–1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagarajan, N.A.; Kronenberg, M. Invariant NKT Cells Amplify the Innate Immune Response to Lipopolysaccharide. J. Immunol. 2007, 178, 2706–2713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reilly, E.C.; Wands, J.R.; Brossay, L. Cytokine Dependent and Independent INKT Cell Activation. Cytokine 2010, 51, 227–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gumperz, J.E.; Miyake, S.; Yamamura, T.; Brenner, M.B. Functionally Distinct Subsets of CD1d-Restricted Natural Killer T Cells Revealed by CD1d Tetramer Staining. J. Exp. Med. 2002, 195, 625–636. [Google Scholar] [CrossRef]
- Wingender, G.; Krebs, P.; Beutler, B.; Kronenberg, M. Antigen-Specific Cytotoxicity by Invariant NKT Cells In Vivo Is CD95/CD178-Dependent and Is Correlated with Antigenic Potency. J. Immunol. 2010, 185, 2721–2729. [Google Scholar] [CrossRef] [Green Version]
- Cortesi, F.; Delfanti, G.; Grilli, A.; Calcinotto, A.; Gorini, F.; Pucci, F.; Lucianò, R.; Grioni, M.; Recchia, A.; Benigni, F.; et al. Bimodal CD40/Fas-Dependent Crosstalk between INKT Cells and Tumor-Associated Macrophages Impairs Prostate Cancer Progression. Cell Rep. 2018, 22, 3006–3020. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Basabe, A.; Strati, F.; Facciotti, F. License to Kill: When Inkt Cells Are Granted the Use of Lethal Cytotoxicity. Int. J. Mol. Sci. 2020, 21, 3909. [Google Scholar] [CrossRef] [PubMed]
- Carnaud, C.; Lee, D.; Donnars, O.; Park, S.H.; Beavis, A.; Koezuka, Y.; Bendelac, A. Cutting Edge: Cross-Talk between Cells of the Innate Immune System: NKT Cells Rapidly Activate NK Cells. J. Immunol. 1999, 163, 4647–4650. [Google Scholar]
- Eberl, G.; MacDonald, H.R. Selective Induction of NK Cell Proliferation and Cytotoxicity by Activated NKT Cells. Eur. J. Immunol. 2000, 30, 985–992. [Google Scholar] [CrossRef]
- Nakamura, T.; Sonoda, K.-H.; Faunce, D.E.; Gumperz, J.; Yamamura, T.; Miyake, S.; Stein-Streilein, J. CD4 + NKT Cells, But Not Conventional CD4 + T Cells, Are Required to Generate Efferent CD8 + T Regulatory Cells Following Antigen Inoculation in an Immune-Privileged Site. J. Immunol. 2003, 171, 1266–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, H.; Nieda, M.; Rozenkov, V.; Nicol, A.J. Analysis of the Effect of Different NKT Cell Subpopulations on the Activation of CD4 and CD8 T Cells, NK Cells, and B Cells. Exp. Hematol. 2006, 34, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The Chemokine System in Diverse Forms of Macrophage Activation and Polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Martinez, F.O.; Gordon, S. The M1 and M2 Paradigm of Macrophage Activation: Time for Reassessment. F1000Prime Rep. 2014, 6, 13. [Google Scholar] [CrossRef] [Green Version]
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.A.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage Plasticity, Polarization, and Function in Health and Disease. J. Cell. Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef]
- Xue, J.; Schmidt, S.V.; Sander, J.; Draffehn, A.; Krebs, W.; Quester, I.; DeNardo, D.; Gohel, T.D.; Emde, M.; Schmidleithner, L.; et al. Transcriptome-Based Network Analysis Reveals a Spectrum Model of Human Macrophage Activation. Immunity 2014, 40, 274–288. [Google Scholar] [CrossRef] [Green Version]
- Murray, P.J.; Allen, J.E.; Biswas, S.K.; Fisher, E.A.; Gilroy, D.W.; Goerdt, S.; Gordon, S.; Hamilton, J.A.; Ivashkiv, L.B.; Lawrence, T.; et al. Macrophage Activation and Polarization: Nomenclature and Experimental Guidelines. Immunity 2014, 41, 14–20. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.; Yakala, G.K.; van den Hil, F.E.; Cochrane, A.; Mummery, C.L.; Orlova, V.V. Differentiation and Functional Comparison of Monocytes and Macrophages from HiPSCs with Peripheral Blood Derivatives. Stem Cell Rep. 2019, 12, 1282–1297. [Google Scholar] [CrossRef] [PubMed]
- Verschoor, C.P.; Puchta, A.; Bowdish, D. The macrophage. In Leucocytes. Methods in Molecular Biology (Methods and Protocols); Ashman, R., Ed.; Springer: New York, NY, USA, 2012; Volume 844. [Google Scholar] [CrossRef]
- Bertani, F.R.; Mozetic, P.; Fioramonti, M.; Iuliani, M.; Ribelli, G.; Pantano, F.; Santini, D.; Tonini, G.; Trombetta, M.; Businaro, L.; et al. Classification of M1/M2-Polarized Human Macrophages by Label-Free Hyperspectral Reflectance Confocal Microscopy and Multivariate Analysis. Sci. Rep. 2017, 7, 8965. [Google Scholar] [CrossRef] [PubMed]
- Auffray, C.; Sieweke, M.H.; Geissmann, F. Blood Monocytes: Development, Heterogeneity, and Relationship with Dendritic Cells. Annu. Rev. Immunol. 2009, 27, 669–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varol, C.; Mildner, A.; Jung, S. Macrophages: Development and tissue specialization. Annu. Rev. Immunol. 2015, 33, 643–675. [Google Scholar] [CrossRef]
- Murray, P.J.; Wynn, T.A. Protective and Pathogenic Functions of Macrophage Subsets. Nat. Rev. Immunol. 2011, 11, 723–737. [Google Scholar] [CrossRef]
- Fujii, S.I.; Liu, K.; Smith, C.; Bonito, A.J.; Steinman, R.M. The Linkage of Innate to Adaptive Immunity via Maturing Dendritic Cells in vivo Requires CD40 Ligation in Addition to Antigen Presentation and CD80/86 Costimulation. J. Exp. Med. 2004, 199, 1607–1618. [Google Scholar] [CrossRef]
- Cortesi, F.; Delfanti, G.; Casorati, G.; Dellabona, P. The Pathophysiological Relevance of the INKT Cell/Mononuclear Phagocyte Crosstalk in Tissues. Front. Immunol. 2018, 9, 2375. [Google Scholar] [CrossRef] [Green Version]
- Brennan, P.J.; Brigl, M.; Brenner, M.B. Invariant Natural Killer T Cells: An Innate Activation Scheme Linked to Diverse Effector Functions. Nat. Rev. Immunol. 2013, 13, 101–117. [Google Scholar] [CrossRef]
- Wang, H.; Breed, E.R.; Lee, Y.J.; Qian, L.J.; Jameson, S.C.; Hogquist, K.A. Myeloid Cells Activate INKT Cells to Produce IL-4 in the Thymic Medulla. Proc. Natl. Acad. Sci. USA 2019, 116, 22262–22268. [Google Scholar] [CrossRef]
- Barral, P.; Polzella, P.; Bruckbauer, A.; Van Rooijen, N.; Besra, G.S.; Cerundolo, V.; Batista, F.D. CD169+ Macrophages Present Lipid Antigens to Mediate Early Activation of INKT Cells in Lymph Nodes. Nat. Immunol. 2010, 11, 303–312. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, N.; Vela, J.L.; Nycholat, C.M.; Rademacher, C.; Khurana, A.; Van Rooijen, N.; Crocker, P.R.; Kronenberg, M.; Paulson, J.C. Targeted Delivery of Lipid Antigen to Macrophages via the CD169/Sialoadhesin Endocytic Pathway Induces Robust Invariant Natural Killer T Cell Activation. Proc. Natl. Acad. Sci. USA 2013, 110, 7826–7831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Gars, M.; Haustant, M.; Klezovich-Bénard, M.; Paget, C.; Trottein, F.; Goossens, P.L.; Tournier, J.-N. Mechanisms of Invariant NKT Cell Activity in Restraining Bacillus Anthracis Systemic Dissemination. J. Immunol. 2016, 197, 3225–3232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barral, P.; Sénchez-Nieo, M.D.; Van Rooijen, N.; Cerundolo, V.; Batista, F.D. The Location of Splenic NKT Cells Favours Their Rapid Activation by Blood-Borne Antigen. EMBO J. 2012, 31, 2378–2390. [Google Scholar] [CrossRef] [PubMed]
- King, I.L.; Amiel, E.; Tighe, M.; Mohrs, K.; Veerapen, N.; Besra, G.; Mohrs, M.; Leadbetter, E.A. The Mechanism of Splenic Invariant NKT Cell Activation Dictates Localization In Vivo. J. Immunol. 2013, 191, 572–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Covarrubias, R.; Wilhelm, A.J.; Major, A.S. Specific Deletion of LDL Receptor-Related Protein on Macrophages Has Skewed in vivo Effects on Cytokine Production by Invariant Natural Killer T Cells. PLoS ONE 2014, 9, e102236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Middendorp, S.; Nieuwenhuis, E.E.S. NKT Cells in Mucosal Immunity. Mucosal Immunol. 2009, 2, 393–402. [Google Scholar] [CrossRef] [Green Version]
- Dowds, C.M.; Blumberg, R.S.; Zeissig, S. Control of Intestinal Homeostasis through Crosstalk between Natural Killer T Cells and the Intestinal Microbiota. Clin. Immunol. 2014, 159, 128–133. [Google Scholar] [CrossRef] [Green Version]
- Sáez de Guinoa, J.; Jimeno, R.; Gaya, M.; Kipling, D.; Garzón, M.J.; Dunn-Walters, D.; Ubeda, C.; Barral, P. CD 1d-mediated Lipid Presentation by CD11c+ Cells Regulates Intestinal Homeostasis. EMBO J. 2018, 37, e97537. [Google Scholar] [CrossRef]
- Gensollen, T.; Lin, X.; Zhang, T.; Pyzik, M.; See, P.; Glickman, J.N.; Ginhoux, F.; Waldor, M.; Salmi, M.; Rantakari, P.; et al. Embryonic Macrophages Function during Early Life to Determine Invariant Natural Killer T Cell Levels at Barrier Surfaces. Nat. Immunol. 2021, 22, 699–710. [Google Scholar] [CrossRef]
- Davies, L.C.; Jenkins, S.J.; Allen, J.E.; Taylor, P.R. Tissue-Resident Macrophages. Nat. Immunol. 2013, 14, 986–995. [Google Scholar] [CrossRef]
- Macrophages, R.; Elchaninov, A.V.; Fatkhudinov, T.K.; Vishnyakova, P.A. Phenotypical and Functional Polymorphism of Liver. Cells 2019, 8, 1032. [Google Scholar]
- Schmieg, J.; Yang, G.; Franck, R.W.; Van Rooijen, N.; Tsuji, M. Glycolipid Presentation to Natural Killer T Cells Differs in an Organ-Dependent Fashion. Proc. Natl. Acad. Sci. USA 2005, 102, 1127–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, L.; Michelet, X.; Zhang, S.; Brennan, P.J.; Moseman, A.; Lester, C.; Besra, G.; Vomhof-Dekrey, E.E.; Tighe, M.; Koay, H.F.; et al. Regulatory INKT Cells Lack Expression of the Transcription Factor PLZF and Control the Homeostasis of T Reg Cells and Macrophages in Adipose Tissue. Nat. Immunol. 2015, 16, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.; Jeong, D.; Oh, S.J.; Ahn, J.; Lee, S.H.; Chung, D.H. GM-CSF and IL-4 Produced by NKT Cells Inversely Regulate IL-1β Production by Macrophages. Immunol. Lett. 2017, 182, 50–56. [Google Scholar] [CrossRef]
- Heffernan, D.S.; Monaghan, S.F.; Thakkar, R.K.; Tran, M.L.; Chung, C.S.; Gregory, S.H.; Cioffi, W.G.; Ayala, A. Inflammatory Mechanisms in Sepsis: Elevated Invariant Natural Killer T-Cell Numbers in Mouse and Their Modulatory Effect on Macrophage Function. Shock 2013, 40, 122–128. [Google Scholar] [CrossRef] [Green Version]
- Fallon, E.A.; Chun, T.T.; Young, W.A.; Gray, C.; Ayala, A.; Heffernan, D.S. Program Cell Death Receptor-1-Mediated Invariant Natural Killer T-Cell Control of Peritoneal Macrophage Modulates Survival in Neonatal Sepsis. Front. Immunol. 2017, 8, 1469. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.K.; Venet, F.; Heffernan, D.S.; Wang, Y.L.; Horner, B.; Huang, X.; Chung, C.-S.; Gregory, S.H.; Ayala, A. The Role of Hepatic Invariant NKT Cells in Systemic/Local Inflammation and Mortality during Polymicrobial Septic Shock. J. Immunol. 2009, 182, 2467–2475. [Google Scholar] [CrossRef] [Green Version]
- Haraguchi, N.; Kikuchi, N.; Morishima, Y.; Matsuyama, M.; Sakurai, H.; Shibuya, A.; Shibuya, K.; Taniguchi, M.; Ishii, Y. Activation of Murine Invariant NKT Cells Promotes Susceptibility to Candidiasis by IL-10 Induced Modulation of Phagocyte Antifungal Activity. Eur. J. Immunol. 2016, 46, 1691–1703. [Google Scholar] [CrossRef]
- Kim, E.Y.; Ner-Gaon, H.; Varon, J.; Cullen, A.M.; Guo, J.; Choi, J.; Barragan-Bradford, D.; Higuera, A.; Pinilla-Vera, M.; Short, S.A.P.; et al. Post-Sepsis Immunosuppression Depends on NKT Cell Regulation of MTOR/IFN-γ in NK Cells. J. Clin. Investig. 2020, 130, 3238–3252. [Google Scholar] [CrossRef] [Green Version]
- Beattie, L.; Svensson, M.; Bune, A.; Brown, N.; Maroof, A.; Zubairi, S.; Smith, K.R.; Kaye, P.M. Leishmania Donovani-Induced Expression of Signal Regulatory Protein α on Kupffer Cells Enhances Hepatic Invariant NKT-Cell Activation. Eur. J. Immunol. 2010, 40, 117–123. [Google Scholar] [CrossRef] [Green Version]
- Karmakar, S.; Paul, J.; De, T. Leishmania Donovani Glycosphingolipid Facilitates Antigen Presentation by Inducing Relocation of CD1d into Lipid Rafts in Infected Macrophages. Eur. J. Immunol. 2011, 41, 1376–1387. [Google Scholar] [CrossRef] [PubMed]
- Karmakar, S.; Bhaumik, S.K.; Paul, J.; De, T. TLR4 and NKT Cell Synergy in Immunotherapy against Visceral Leishmaniasis. PLoS Pathog. 2012, 8, e1002646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, W.Y.; Moriarty, T.J.; Wong, C.H.Y.; Zhou, H.; Strieter, R.M.; Van Rooijen, N.; Chaconas, G.; Kubes, P. An Intravascular Immune Response to Borrelia Burgdorferi Involves Kupffer Cells and INKT Cells. Nat. Immunol. 2010, 11, 295–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emoto, M.; Yoshida, T.; Fukuda, T.; Kawamura, I.; Mitsuyama, M.; Kita, E.; Hurwitz, R.; Kaufmann, S.H.E.; Emoto, Y. α-Galactosylceramide Promotes Killing of Listeria Monocytogenes within the Macrophage Phagosome through Invariant NKT-Cell Activation. Infect. Immun. 2010, 78, 2667–2676. [Google Scholar] [CrossRef] [Green Version]
- Gansert, J.L.; Kieβler, V.; Engele, M.; Wittke, F.; Röllinghoff, M.; Krensky, A.M.; Porcelli, S.A.; Modlin, R.L.; Stenger, S. Human NKT Cells Express Granulysin and Exhibit Antimycobacterial Activity. J. Immunol. 2003, 170, 3154–3161. [Google Scholar] [CrossRef] [Green Version]
- Sada-Ovalle, I.; Chiba, A.; Gonzales, A.; Brenner, M.B.; Behar, S.M. Innate Invariant NKT Cells Recognize Mycobacterium Tuberculosis-Infected Macrophages, Produce Interferon-γ, and Kill Intracellular Bacteria. PLoS Pathog. 2008, 4, e1000239. [Google Scholar] [CrossRef]
- Rothchild, A.C.; Jayaraman, P.; Nunes-Alves, C.; Behar, S.M. INKT Cell Production of GM-CSF Controls Mycobacterium Tuberculosis. PLoS Pathog. 2014, 10, e1003805. [Google Scholar] [CrossRef]
- Nieuwenhuis, E.E.S. CD1d-Dependent, Macrophage-Mediated Clearance of Pseudomonas Aeruginosa from Lung. Biomed. Pharmacother. 2002, 56, 423. [Google Scholar] [CrossRef]
- Chandra, S.; Gray, J.; Kiosses, W.B.; Khurana, A.; Hitomi, K.; Crosby, C.M.; Chawla, A.; Fu, Z.; Zhao, M.; Veerapen, N.; et al. Mrp1 Is Involved in Lipid Presentation and INKT Cell Activation by Streptococcus Pneumoniae. Nat. Commun. 2018, 9, 4279. [Google Scholar] [CrossRef]
- Ho, L.P.; Denny, L.; Luhn, K.; Teoh, D.; Clelland, C.; McMichael, A.J. Activation of Invariant NKT Cells Enhances the Innate Immune Response and Improves the Disease Course in Influenza A Virus Infection. Eur. J. Immunol. 2008, 38, 1913–1922. [Google Scholar] [CrossRef]
- Kok, W.L.; Denney, L.; Benam, K.; Cole, S.; Clelland, C.; McMichael, A.J.; Ho, L.-P. Pivotal Advance: Invariant NKT Cells Reduce Accumulation of Inflammatory Monocytes in the Lungs and Decrease Immune-Pathology during Severe Influenza A Virus Infection. J. Leukoc. Biol. 2012, 91, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Gaya, M.; Barral, P.; Burbage, M.; Aggarwal, S.; Montaner, B.; Warren Navia, A.; Aid, M.; Tsui, C.; Maldonado, P.; Nair, U.; et al. Initiation of Antiviral B Cell Immunity Relies on Innate Signals from Spatially Positioned NKT Cells. Cell 2018, 172, 517–533.e20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.Y.; Battaile, J.T.; Patel, A.C.; You, Y.; Agapov, E.; Grayson, M.H.; Benoit, L.A.; Byers, D.E.; Alevy, Y.; Tucker, J.; et al. Persistent Activation of an Innate Immune Response Translates Respiratory Viral Infection into Chronic Lung Disease. Nat. Med. 2008, 14, 633–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, K.; Yang, J.; Luo, K.; Yang, C.; Zhang, N.; Xu, R. TLR3 Signaling in Macrophages Is Indispensable for the Protective Immunity of Invariant Natural Killer T Cells against Enterovirus 71 Infection. PLoS Pathog. 2015, 11, e1004613. [Google Scholar] [CrossRef] [Green Version]
- Bessoles, S.; Dudal, S.; Besra, G.S.; Sanchez, F.; Lafont, V. Human CD4+ Invariant NKT Cells Are Involved in Antibacterial Immunity against Brucella Suis through CD1d-Dependent but CD4-Independent Mechanisms. Eur. J. Immunol. 2009, 39, 1025–1035. [Google Scholar] [CrossRef]
- Ji, Y.; Sun, S.; Xu, A.; Bhargava, P.; Yang, L.; Lam, K.S.L.; Gao, B.; Lee, C.H.; Kersten, S.; Qi, L. Activation of Natural Killer T Cells Promotes M2 Macrophage Polarization in Adipose Tissue and Improves Systemic Glucose Tolerance via Interleukin-4 (IL-4)/STAT6 Protein Signaling Axis in Obesity. J. Biol. Chem. 2012, 287, 13561–13571. [Google Scholar] [CrossRef] [Green Version]
- Ji, Y.; Sun, S.; Xia, S.; Yang, L.; Li, X.; Qi, L. Short Term High Fat Diet Challenge Promotes Alternative Macrophage Polarization in Adipose Tissue via Natural Killer T Cells and Interleukin-4. J. Biol. Chem. 2012, 287, 24378–24386. [Google Scholar] [CrossRef] [Green Version]
- Lynch, L.; Nowak, M.; Varghese, B.; Clark, J.; Hogan, A.E.; Toxavidis, V.; Balk, S.P.; O’Shea, D.; O’Farrelly, C.; Exley, M.A. Adipose Tissue Invariant NKT Cells Protect against Diet-Induced Obesity and Metabolic Disorder through Regulatory Cytokine Production. Immunity 2012, 37, 574–587. [Google Scholar] [CrossRef] [Green Version]
- Lynch, L.; O’Shea, D.; Winter, D.C.; Geoghegan, J.; Doherty, D.G.; O’Farrelly, C. Invariant NKT Cells and CD1d+ Cells Amass in Human Omentum and Are Depleted in Patients with Cancer and Obesity. Eur. J. Immunol. 2009, 39, 1893–1901. [Google Scholar] [CrossRef]
- Zhang, H.; Xue, R.; Zhu, S.; Fu, S.; Chen, Z.; Zhou, R.; Tian, Z.; Bai, L. M2-Specific Reduction of CD1d Switches NKT Cell-Mediated Immune Responses and Triggers Metaflammation in Adipose Tissue. Cell. Mol. Immunol. 2018, 15, 506–517. [Google Scholar] [CrossRef]
- Ohmura, K.; Ishimori, N.; Ohmura, Y.; Tokuhara, S.; Nozawa, A.; Horii, S.; Andoh, Y.; Fujii, S.; Iwabuchi, K.; Onoé, K.; et al. Natural Killer T Cells Are Involved in Adipose Tissues Inflammation and Glucose Intolerance in Diet-Induced Obese Mice. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Parekh, V.V.; Gabriel, C.L.; Bracy, D.P.; Marks-Shulman, P.A.; Tamboli, R.A.; Kim, S.; Mendez-Fernandez, Y.V.; Besra, G.S.; Lomenick, J.P.; et al. Activation of Invariant Natural Killer T Cells by Lipid Excess Promotes Tissue Inflammation, Insulin Resistance, and Hepatic Steatosis in Obese Mice. Proc. Natl. Acad. Sci. USA 2012, 109, E1143–E1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantell, B.S.; Stefanovic-Racic, M.; Yang, X.; Dedousis, N.; Sipula, I.J.; O’Doherty, R.M. Mice Lacking NKT Cells but with a Complete Complement of CD8+ T-Cells Are Not Protected against the Metabolic Abnormalities of Diet-Induced Obesity. PLoS ONE 2011, 6, e19831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liew, P.X.; Lee, W.Y.; Kubes, P. INKT Cells Orchestrate a Switch from Inflammation to Resolution of Sterile Liver Injury. Immunity 2017, 47, 752–765.e5. [Google Scholar] [CrossRef] [Green Version]
- Kremer, M.; Thomas, E.; Milton, R.J.; Perry, A.W.; Van Rooijen, N.; Wheeler, M.D.; Zacks, S.; Fried, M.; Rippe, R.A.; Hines, I.N. Kupffer Cell and Interleukin-12-Dependent Loss of Natural Killer T Cells in Hepatosteatosis. Hepatology 2010, 51, 130–141. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Li, L.; Li, Y.; Li, Y.; Sha, Y.; Wen, S.; You, Q.; Liu, L.; Shi, M.; Zhou, H. Intravital Imaging of Interactions between INKT and Kupffer Cells to Clear Free Lipids during Steatohepatitis. Theranostics 2021, 11, 2149–2169. [Google Scholar] [CrossRef]
- Zeng, M.Y.; Pham, D.; Bagaitkar, J.; Liu, J.; Otero, K.; Shan, M.; Wynn, T.A.; Brombacher, F.; Brutkiewicz, R.R.; Kaplan, M.H.; et al. An Efferocytosis-Induced, IL-4-Dependent Macrophage-INKT Cell Circuit Suppresses Sterile Inflammation and Is Defective in Murine CGD. Blood 2013, 121, 3473–3483. [Google Scholar] [CrossRef] [Green Version]
- Brailey, P.M.; Lebrusant-Fernandez, M.; Barral, P. NKT Cells and the Regulation of Intestinal Immunity: A Two-Way Street. FEBS J. 2020, 287, 1686–1699. [Google Scholar] [CrossRef]
- Heller, F.; Fuss, I.J.; Nieuwenhuis, E.E.; Blumberg, R.S.; Strober, W. Oxazolone Colitis, a Th2 Colitis Model Resembling Ulcerative Colitis, Is Mediated by IL-13-Producing NK-T Cells. Immunity 2002, 17, 629–638. [Google Scholar] [CrossRef] [Green Version]
- Iyer, S.S.; Gensollen, T.; Gandhi, A.; Oh, S.F.; Neves, J.F.; Collin, F.; Lavin, R.; Serra, C.; Glickman, J.; de Silva, P.S.A.; et al. Dietary and Microbial Oxazoles Induce Intestinal Inflammation by Modulating Aryl Hydrocarbon Receptor Responses. Cell 2018, 173, 1123–1134.e11. [Google Scholar] [CrossRef] [Green Version]
- Lezmi, G.; Leite-de-Moraes, M. Invariant Natural Killer T and Mucosal-Associated Invariant T Cells in Asthmatic Patients. Front. Immunol. 2018, 9, 1766. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Chang, Y.J.; Subramanian, S.; Lee, H.H.; Albacker, L.A.; Matangkasombut, P.; Savage, P.B.; McKenzie, A.N.J.; Smith, D.E.; Rottman, J.B.; et al. Innate Lymphoid Cells Responding to IL-33 Mediate Airway Hyperreactivity Independently of Adaptive Immunity. J. Allergy Clin. Immunol. 2012, 129, 216–227.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakai, Y.; Iwabuchi, K.; Fujii, S.; Ishimori, N.; Dashtsoodol, N.; Watano, K.; Mishima, T.; Iwabuchi, C.; Tanaka, S.; Bezbradica, J.S.; et al. Natural Killer T Cells Accelerate Atherogenesis in Mice. Blood 2004, 104, 2051–2059. [Google Scholar] [CrossRef]
- Tupin, E.; Nicoletti, A.; Elhage, R.; Rudling, M.; Ljunggren, H.G.; Hansson, G.K.; Berne, G.P. CD1d-Dependent Activation of NKT Cells Aggravates Atherosclerosis. J. Exp. Med. 2004, 199, 417–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, E.; Croca, S.; Waddington, K.E.; Sofat, R.; Griffin, M.; Nicolaides, A.; Isenberg, D.A.; Torra, I.P.; Rahman, A.; Jury, E.C. Cross-Talk between INKT Cells and Monocytes Triggers an Atheroprotective Immune Response in SLE Patients with Asymptomatic Plaque. Sci. Immunol. 2016, 1, eaah4081. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Ara, T.; Wu, H.W.; Woo, C.W.; Reynolds, C.P.; Seeger, R.C.; DeClerck, Y.A.; Thiele, C.J.; Sposto, R.; Metelitsa, L.S. Oncogene MYCN Regulates Localization of NKT Cells to the Site of Disease in Neuroblastoma. J. Clin. Investig. 2007, 117, 2702–2712. [Google Scholar] [CrossRef] [Green Version]
- Metelitsa, L.S.; Wu, H.W.; Wang, H.; Yang, Y.; Warsi, Z.; Asgharzadeh, S.; Groshen, S.; Wilson, S.B.; Seeger, R.C. Natural Killer T Cells Infiltrate Neuroblastomas Expressing the Chemokine CCL2. J. Exp. Med. 2004, 199, 1213–1221. [Google Scholar] [CrossRef] [Green Version]
- Bottazzi, B.; Polentarutti, N.; Acero, R.; Balsari, A.; Boraschi, D.; Ghezzi, P.; Salmona, M.; Mantovani, A. Regulation of the Macrophage Content of Neoplasms by Chemoattractants. Science 1983, 220, 210–212. [Google Scholar] [CrossRef]
- Matsushima, K.; Larsen, C.G.; DuBois, G.C.; Oppenheim, J.J. Purification and Characterization of a Novel Monocyte Chemotactic and Activating Factor Produced by a Human Myelomonocytic Cell Line. J. Exp. Med. 1989, 169, 1485–1490. [Google Scholar] [CrossRef] [Green Version]
- Song, L.; Asgharzadeh, S.; Salo, J.; Engell, K.; Wu, H.W.; Sposto, R.; Ara, T.; Silverman, A.M.; DeClerck, Y.A.; Seeger, R.C.; et al. Vα24-Invariant NKT Cells Mediate Antitumor Activity via Killing of Tumor-Associated Macrophages. J. Clin. Investig. 2009, 119, 1524–1536. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Song, L.; Wei, J.; Courtney, A.N.; Gao, X.; Marinova, E.; Guo, L.; Heczey, A.; Asgharzadeh, S.; Kim, E.; et al. IL-15 Protects NKT Cells from Inhibition by Tumor-Associated Macrophages and Enhances Antimetastatic Activity. J. Clin. Investig. 2012, 122, 2221–2233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schutyser, E.; Struyf, S.; Van Damme, J. The CC Chemokine CCL20 and Its Receptor CCR6. Cytokine Growth Factor Rev. 2003, 14, 409–426. [Google Scholar] [CrossRef]
- Thomas, S.Y.; Hou, R.; Boyson, J.E.; Means, T.K.; Hess, C.; Olson, D.P.; Strominger, J.L.; Brenner, M.B.; Gumperz, J.E.; Wilson, S.B.; et al. CD1d-Restricted NKT Cells Express a Chemokine Receptor Profile Indicative of Th1-Type Inflammatory Homing Cells. J. Immunol. 2003, 171, 2571–2580. [Google Scholar] [CrossRef] [Green Version]
- Paul, S.; Chhatar, S.; Mishra, A.; Lal, G. Natural Killer T Cell Activation Increases INOS+CD206− M1 Macrophage and Controls the Growth of Solid Tumor. J. Immunother. Cancer 2019, 7, 208. [Google Scholar] [CrossRef] [Green Version]
- Janakiram, N.B.; Mohammed, A.; Bryant, T.; Ritchie, R.; Stratton, N.; Jackson, L.; Lightfoot, S.; Benbrook, D.M.; Asch, A.S.; Lang, M.L.; et al. Loss of Natural Killer T Cells Promotes Pancreatic Cancer in LSL-KrasG12D/+ Mice. Immunology 2017, 152, 36–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Sedimbi, S.; Löfbom, L.; Singh, A.K.; Porcelli, S.A.; Cardell, S.L. Unique Invariant Natural Killer T Cells Promote Intestinal Polyps by Suppressing TH1 Immunity and Promoting Regulatory T Cells. Mucosal Immunol. 2018, 11, 131–143. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Bhave, M.S.; Yagita, H.; Cardell, S.L. Natural Killer T-Cell Agonist α-Galactosylceramide and PD-1 Blockade Synergize to Reduce Tumor Development in a Preclinical Model of Colon Cancer. Front. Immunol. 2020, 11, 581301. [Google Scholar] [CrossRef]
- Gorini, F.; Azzimonti, L.; Delfanti, G.; Scarfo, L.; Scielzo, C.; Bertilaccio, M.T.; Ranghetti, P.; Gulino, A.; Doglioni, C.; Di Napoli, A.; et al. Invariant NKT Cells Contribute to Chronic Lymphocytic Leukemia Surveillance and Prognosis. Blood 2017, 129, 3440–3451. [Google Scholar] [CrossRef]
- Denney, L.; Kok, W.L.; Cole, S.L.; Sanderson, S.; McMichael, A.J.; Ho, L.-P. Activation of Invariant NKT Cells in Early Phase of Experimental Autoimmune Encephalomyelitis Results in Differentiation of Ly6C Hi Inflammatory Monocyte to M2 Macrophages and Improved Outcome. J. Immunol. 2012, 189, 551–557. [Google Scholar] [CrossRef] [Green Version]
- Lehuen, A.; Diana, J.; Zaccone, P.; Cooke, A. Immune Cell Crosstalk in Type 1 Diabetes. Nat. Rev. Immunol. 2010, 10, 501–513. [Google Scholar] [CrossRef]
- Ghazarian, L.; Diana, J.; Beaudoin, L.; Larsson, P.G.; Puri, R.K.; Van Rooijen, N.; Flodström-Tullberg, M.; Lehuen, A. Protection against Type 1 Diabetes upon Coxsackievirus B4 Infection and INKT-Cell Stimulation: Role of Suppressive Macrophages. Diabetes 2013, 62, 3785–3796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drennan, M.B.; Aspeslagh, S.; Elewaut, D. Invariant Natural Killer T Cells in Rheumatic Disease: A Joint Dilemma. Nat. Rev. Rheumatol. 2010, 6, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Miellot-Gafsou, A.; Biton, J.; Bourgeois, E.; Herbelin, A.; Boissier, M.C.; Bessis, N. Early Activation of Invariant Natural Killer T Cells in a Rheumatoid Arthritis Model and Application to Disease Treatment. Immunology 2010, 130, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Basri, F.; Jung, S.; Park, S.H.; Park, S.H. CD1d Deficiency Limits Tolerogenic Properties of Peritoneal Macrophages. BMB Rep. 2021, 54, 209–214. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cruz, M.S.; Loureiro, J.P.; Oliveira, M.J.; Macedo, M.F. The iNKT Cell–Macrophage Axis in Homeostasis and Disease. Int. J. Mol. Sci. 2022, 23, 1640. https://doi.org/10.3390/ijms23031640
Cruz MS, Loureiro JP, Oliveira MJ, Macedo MF. The iNKT Cell–Macrophage Axis in Homeostasis and Disease. International Journal of Molecular Sciences. 2022; 23(3):1640. https://doi.org/10.3390/ijms23031640
Chicago/Turabian StyleCruz, Mariana S., José Pedro Loureiro, Maria J. Oliveira, and Maria Fatima Macedo. 2022. "The iNKT Cell–Macrophage Axis in Homeostasis and Disease" International Journal of Molecular Sciences 23, no. 3: 1640. https://doi.org/10.3390/ijms23031640
APA StyleCruz, M. S., Loureiro, J. P., Oliveira, M. J., & Macedo, M. F. (2022). The iNKT Cell–Macrophage Axis in Homeostasis and Disease. International Journal of Molecular Sciences, 23(3), 1640. https://doi.org/10.3390/ijms23031640