
Functional Transient Receptor Potential Ankyrin 1 and Vanilloid 1 Ion Channels Are Overexpressed in Human Oral Squamous Cell Carcinoma

 , , ,
, , , 
 , and
, and 
Abstract
:1. Introduction
2. Results
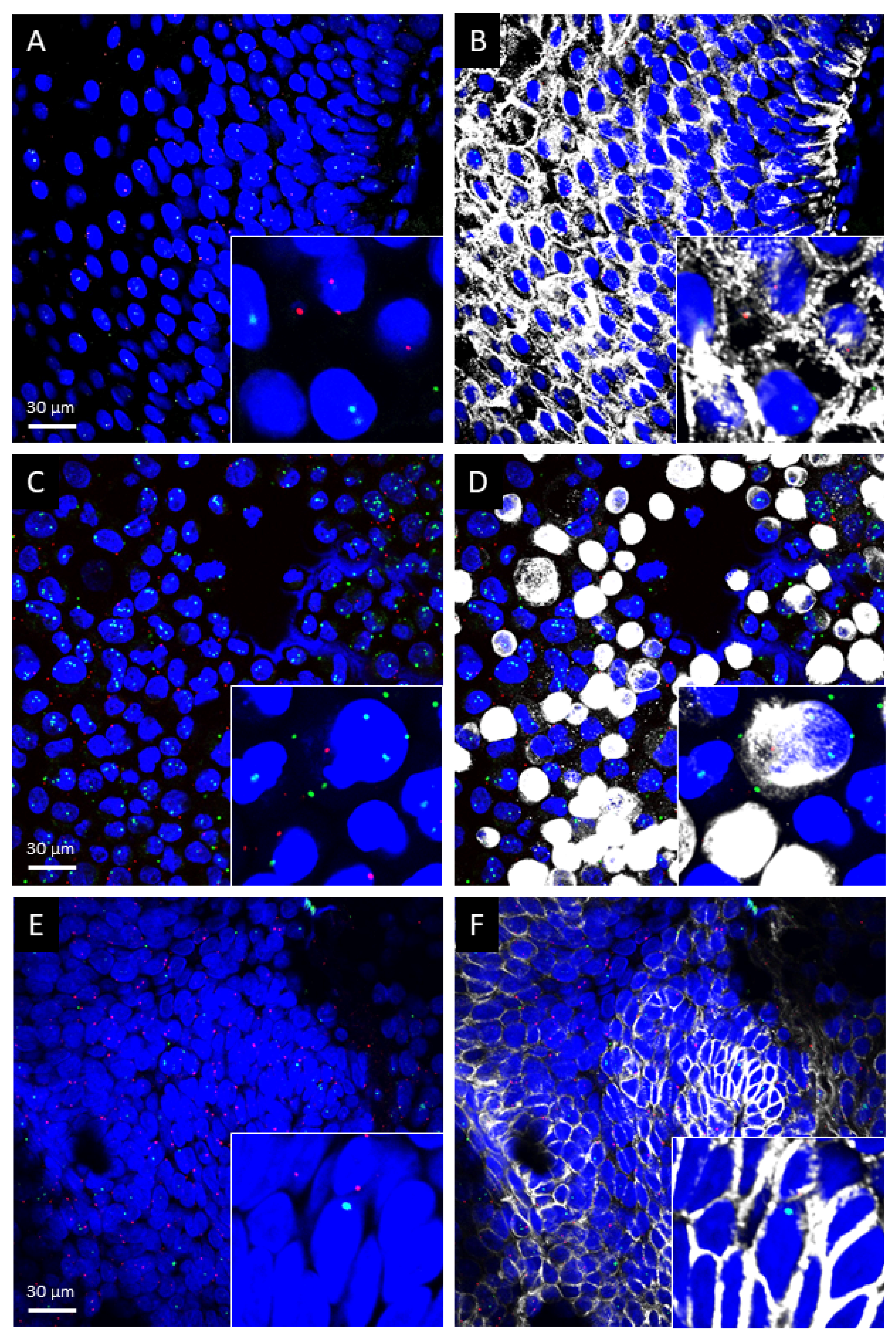
2.1. TRPA1 and TRPV1 mRNAs Are Present in Healthy Human Oral Mucosa Epithelial Cells, OSCC and PE/CA-PJ41 Cell Line
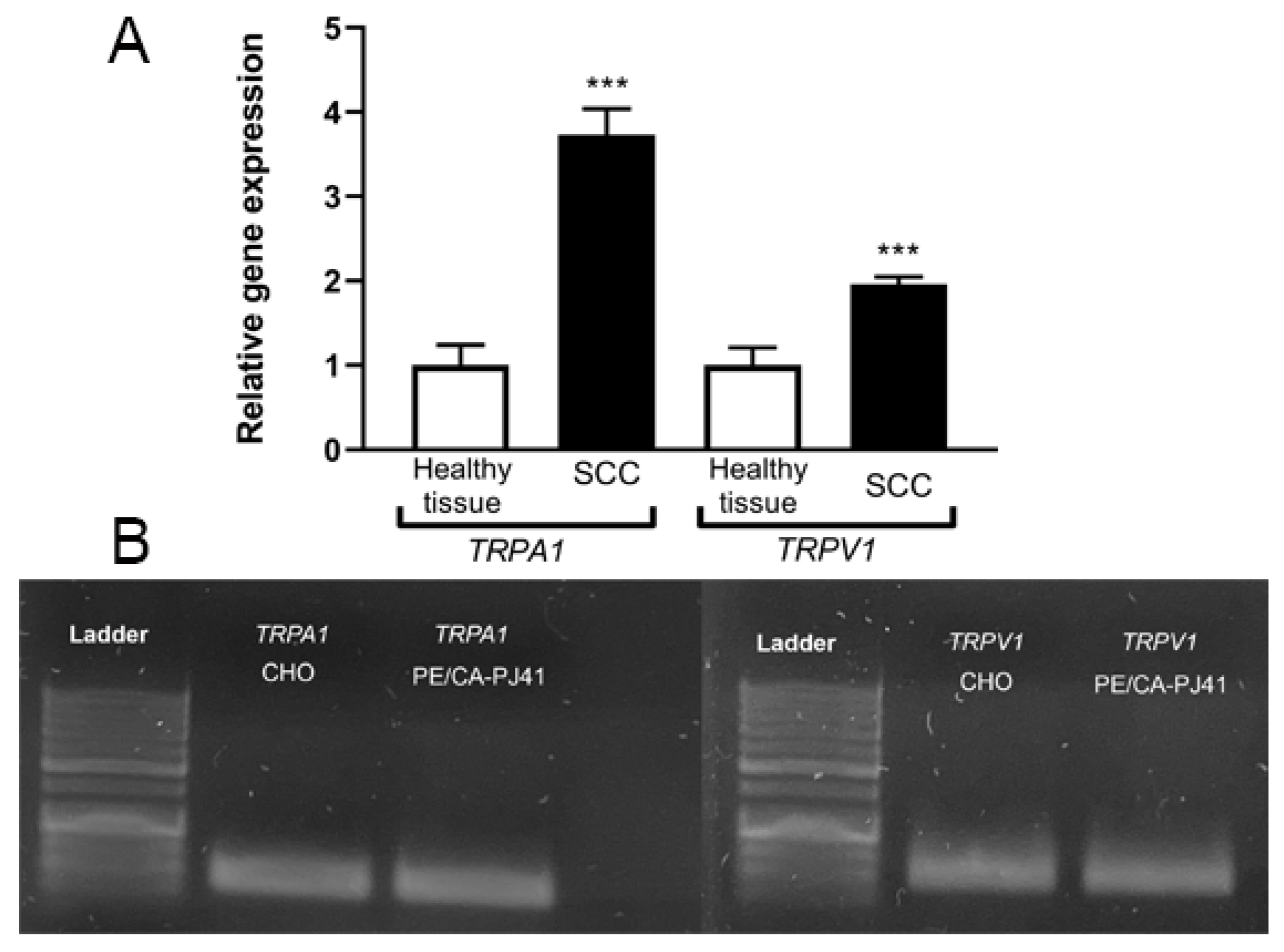
2.2. TRPA1 and TRPV1 mRNAs Are Significantly Upregulated in OSCC Samples
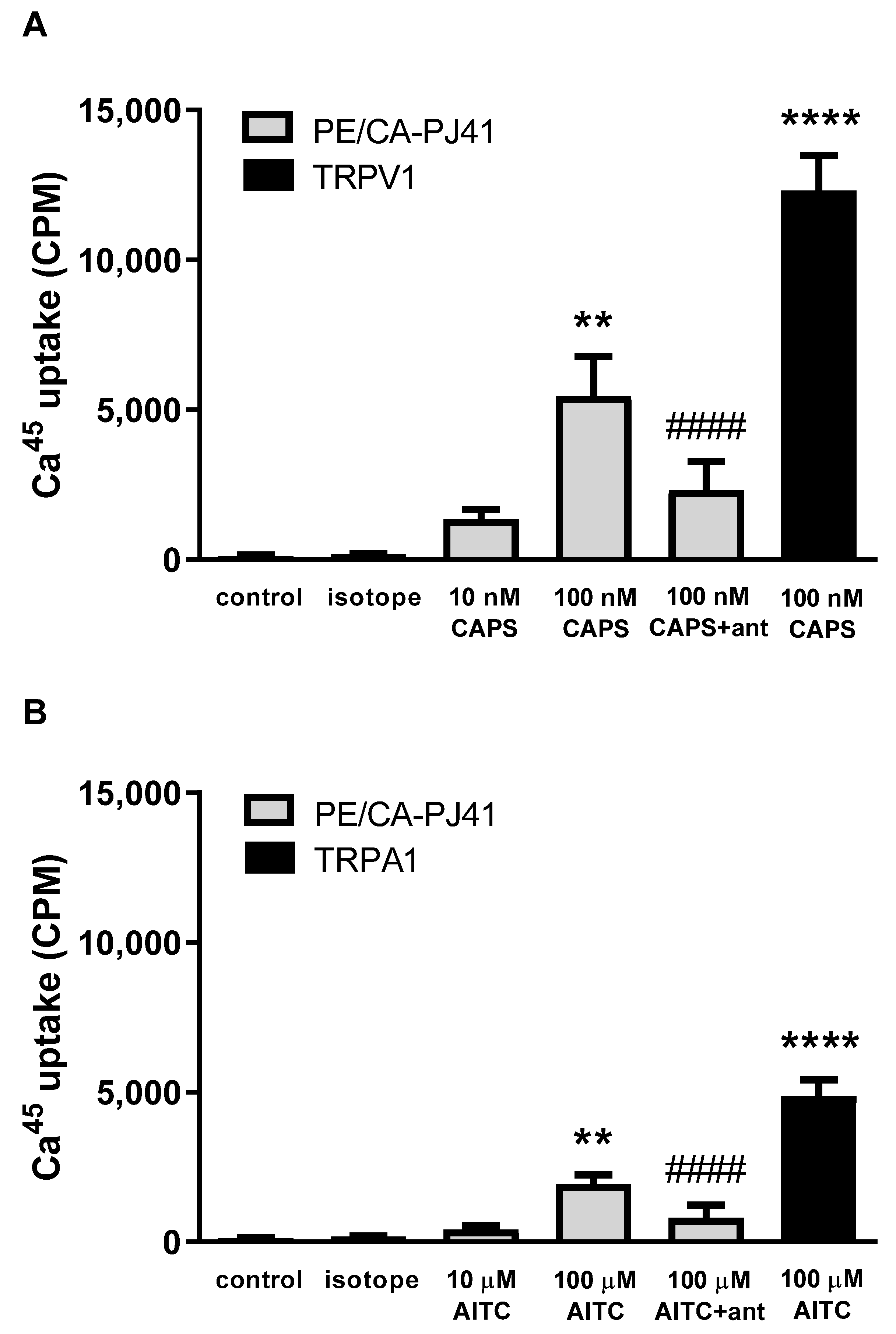
2.3. Both TRPA1 and TRPV1 Activations Induce Radioactive 45Ca2+ Uptake in PE/CA-PJ41 Cells
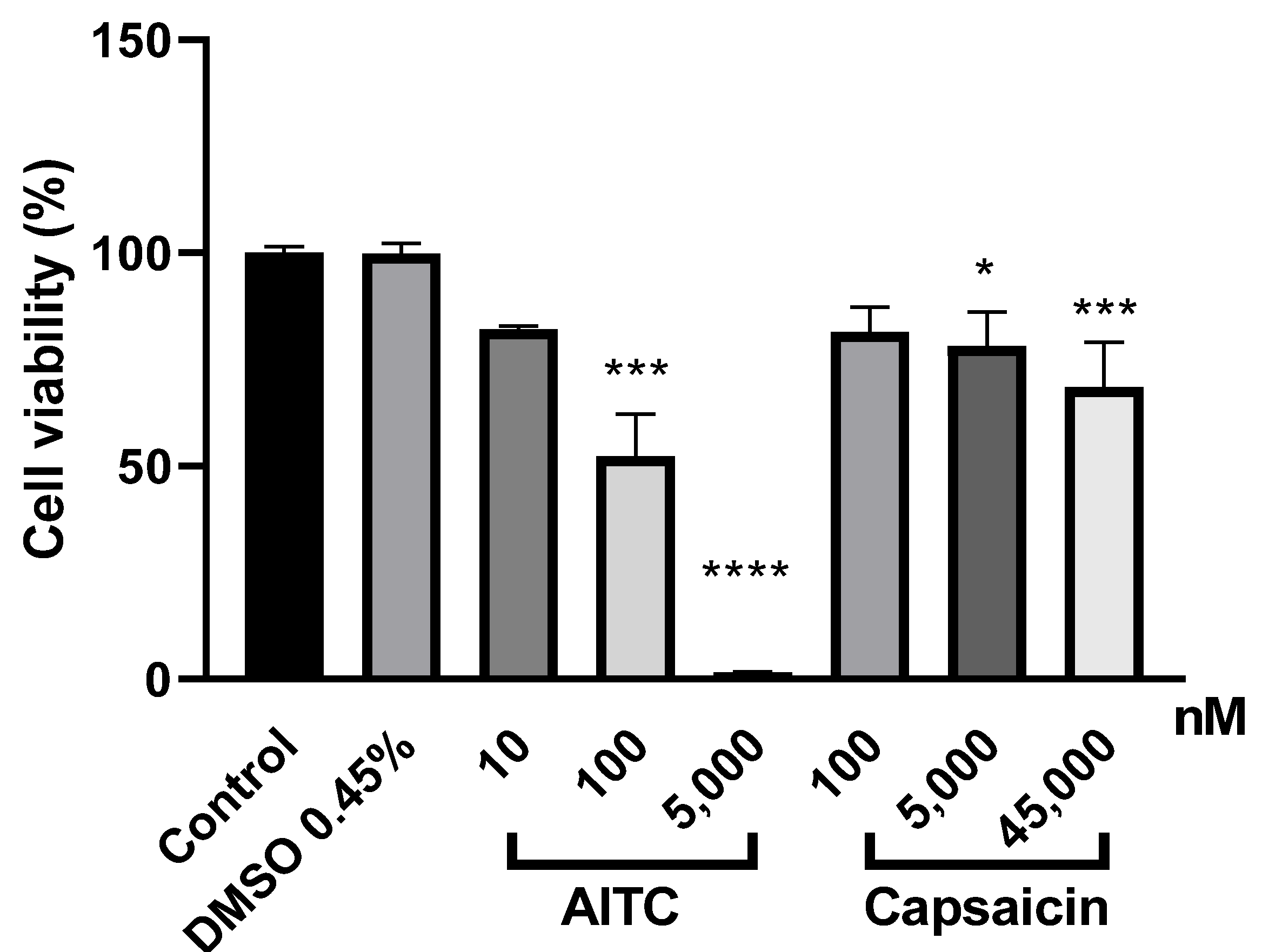
2.4. Both TRPA1 and TRPV1 Activations Reduce PE/CA-PJ41 Cell Viability
3. Discussion
4. Materials and Methods
4.1. Study Participants and Tissue Collection
4.2. Human Oral Squamous Cell Carcinoma Cell Line
4.3. RNA Isolation and Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR)
4.4. Tissue Collection and Sample Preparation for RNAscope Study
4.5. Immunohistochemistry and Image Analysis
4.6. RNAscope In Situ Hybridization Combined with Immunolabeling
4.7. Radioactive Calcium-45 Uptake Experiments on PE/CA-PJ41 Cells and TRPV1 or TRPA1 Receptor-Expressing CHO Cell Lines
4.8. Cell Viability Assay
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, T.; Wakai, K.; Matsuo, K.; Hirose, K.; Ito, H.; Kuriki, K.; Sato, S.; Ueda, R.; Hasegawa, Y.; Tajima, K. Effect of dietary antioxidants and risk of oral, pharyngeal and laryngeal squamous cell carcinoma according to smoking and drinking habits. Cancer Sci. 2006, 97, 760–767. [Google Scholar] [CrossRef] [PubMed]
- Humans, I. Alcohol drinking. Epidemiological studies of cancer in humans. IARC Monogr. Eval. Carcinog. Risks Hum. 1988, 44, 153–250. [Google Scholar]
- Hashibe, M.; Brennan, P.; Benhamou, S.; Castellsague, X.; Chen, C.; Curado, M.P.; Dal Maso, L.; Daudt, A.W.; Fabianova, E.; Fernandez, L.; et al. Alcohol drinking in never users of tobacco, cigarette smoking in never drinkers, and the risk of head and neck cancer: Pooled analysis in the International Head and Neck Cancer Epidemiology Consortium. J. Natl. Cancer Inst. 2007, 99, 777–789. [Google Scholar] [CrossRef]
- Lewin, F.; Norell, S.E.; Johansson, H.; Gustavsson, P.; Wennerberg, J.; Biörklund, A.; Rutqvist, L.E. Smoking tobacco, oral snuff, and alcohol in the etiology of squamous cell carcinoma of the head and neck: A population-based case-referent study in Sweden. Cancer 1998, 82, 1367–1375. [Google Scholar] [CrossRef]
- Sankaranarayanan, R.; Masuyer, E.; Swaminathan, R.; Ferlay, J.; Whelan, S. Head and neck cancer: A global perspective on epidemiology and prognosis. Anticancer Res. 1998, 18, 4779–4786. [Google Scholar]
- Ouadid-Ahidouch, H.; Dhennin-Duthille, I.; Gautier, M.; Sevestre, H.; Ahidouch, A. TRP channels: Diagnostic markers and therapeutic targets for breast cancer? Trends Mol. Med. 2013, 19, 117–124. [Google Scholar] [CrossRef]
- Weber, L.V.; Al-Refae, K.; Wölk, G.; Bonatz, G.; Altmüller, J.; Becker, C.; Gisselmann, G.; Hatt, H. Expression and functionality of TRPV1 in breast cancer cells. Breast Cancer Targets Ther. 2016, 8, 243–252. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.T.; Peters, A.A.; Tan, P.T.; Roberts-Thomson, S.J.; Monteith, G.R. Consequences of activating the calcium-permeable ion channel TRPV1 in breast cancer cells with regulated TRPV1 expression. Cell Calcium 2014, 56, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Gao, N.; Yang, F.; Chen, S.; Wan, H.; Zhao, X.; Dong, H. The role of TRPV1 ion channels in the suppression of gastric cancer development. J. Exp. Clin. Cancer Res. 2020, 39, 206. [Google Scholar] [CrossRef]
- Fonseca, B.M.; Correia-da-Silva, G.; Teixeira, N.A. Cannabinoid-induced cell death in endometrial cancer cells: Involvement of TRPV1 receptors in apoptosis. J. Physiol. Biochem. 2018, 74, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Chinigò, G.; Castel, H.; Chever, O.; Gkika, D. TRP Channels in Brain Tumors. Front. Cell Dev. Biol. 2021, 9, 617801. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, E.A.; Stohr, S.; Meister, M.; Aigner, A.; Gudermann, T.; Buech, T.R. Stimulation of the chemosensory TRPA1 cation channel by volatile toxic substances promotes cell survival of small cell lung cancer cells. Biochem. Pharmacol. 2013, 85, 426–438. [Google Scholar] [CrossRef] [PubMed]
- Van Haute, C.; De Ridder, D.; Nilius, B. TRP Channels in Human Prostate. Sci. World J. 2010, 10, 832469. [Google Scholar] [CrossRef] [Green Version]
- Sakakibara, A.; Sakakibara, S.; Kusumoto, J.; Takeda, D.; Hasegawa, T.; Akashi, M.; Minamikawa, T.; Hashikawa, K.; Terashi, H.; Komori, T. Upregulated Expression of Transient Receptor Potential Cation Channel Subfamily V Receptors in Mucosae of Patients with Oral Squamous Cell Carcinoma and Patients with a History of Alcohol Consumption or Smoking. PLoS ONE 2017, 12, e0169723. [Google Scholar] [CrossRef]
- Bautista, D.M.; Pellegrino, M.; Tsunozaki, M. TRPA1: A gatekeeper for inflammation. Annu. Rev. Physiol. 2013, 75, 181–200. [Google Scholar] [CrossRef] [Green Version]
- Radresa, O.; Dahllöf, H.; Nyman, E.; Nolting, A.; Albert, J.S.; Raboisson, P. Roles of TRPA1 in Pain Pathophysiology and Implications for the Development of a New Class of Analgesic Drugs. Open Pain J. 2013, 6, 137–153. [Google Scholar] [CrossRef] [Green Version]
- Bautista, D.M.; Jordt, S.E.; Nikai, T.; Tsuruda, P.R.; Read, A.J.; Poblete, J.; Yamoah, E.N.; Basbaum, A.I.; Julius, D. TRPA1 mediates the inflammatory actions of environmental irritants and proalgesic agents. Cell 2006, 124, 1269–1282. [Google Scholar] [CrossRef] [Green Version]
- Karashima, Y.; Talavera, K.; Everaerts, W.; Janssens, A.; Kwan, K.Y.; Vennekens, R.; Nilius, B.; Voets, T. TRPA1 acts as a cold sensor in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2009, 106, 1273–1278. [Google Scholar] [CrossRef] [Green Version]
- Story, G.M.; Peier, A.M.; Reeve, A.J.; Eid, S.R.; Mosbacher, J.; Hricik, T.R.; Earley, T.J.; Hergarden, A.C.; Andersson, D.A.; Hwang, S.W.; et al. ANKTM1, a TRP-like channel expressed in nociceptive neurons, is activated by cold temperatures. Cell 2003, 112, 819–829. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.F.; Chen, J.; Faltynek, C.R.; Moreland, R.B.; Neelands, T.R. Transient receptor potential A1 mediates an osmotically activated ion channel. Eur. J. Neurosci. 2008, 27, 605–611. [Google Scholar] [CrossRef] [PubMed]
- López-González, M.J.; Luis, E.; Fajardo, O.; Meseguer, V.; Gers-Barlag, K.; Niñerola, S.; Viana, F. TRPA1 Channels Mediate Human Gingival Fibroblast Response to Phenytoin. J. Dent. Res. 2017, 96, 832–839. [Google Scholar] [CrossRef] [PubMed]
- Trevisani, M.; Siemens, J.; Materazzi, S.; Bautista, D.M.; Nassini, R.; Campi, B.; Imamachi, N.; Andrè, E.; Patacchini, R.; Cottrell, G.S.; et al. 4-Hydroxynonenal, an endogenous aldehyde, causes pain and neurogenic inflammation through activation of the irritant receptor TRPA1. Proc. Natl. Acad. Sci. USA 2007, 104, 13519–13524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obata, K.; Katsura, H.; Mizushima, T.; Yamanaka, H.; Kobayashi, K.; Dai, Y.; Fukuoka, T.; Tokunaga, A.; Tominaga, M.; Noguchi, K. TRPA1 induced in sensory neurons contributes to cold hyperalgesia after inflammation and nerve injury. J. Clin. Investig. 2005, 115, 2393–2401. [Google Scholar] [CrossRef] [Green Version]
- Jordt, S.E.; Bautista, D.M.; Chuang, H.H.; McKemy, D.D.; Zygmunt, P.M.; Högestätt, E.D.; Meng, I.D.; Julius, D. Mustard oils and cannabinoids excite sensory nerve fibres through the TRP channel ANKTM1. Nature 2004, 427, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Earley, S. TRPA1 channels in the vasculature. Br. J. Pharmacol. 2012, 167, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Nilius, B.; Mahieu, F. A road map for TR(I)Ps. Mol. Cell 2006, 22, 297–307. [Google Scholar] [CrossRef]
- Geppetti, P.; Trevisani, M. Activation and sensitisation of the vanilloid receptor: Role in gastrointestinal inflammation and function. Br. J. Pharmacol. 2004, 141, 1313–1320. [Google Scholar] [CrossRef]
- Nassini, R.; Pedretti, P.; Moretto, N.; Fusi, C.; Carnini, C.; Facchinetti, F.; Viscomi, A.R.; Pisano, A.R.; Stokesberry, S.; Brunmark, C.; et al. Transient receptor potential ankyrin 1 channel localized to non-neuronal airway cells promotes non-neurogenic inflammation. PLoS ONE 2012, 7, e42454. [Google Scholar] [CrossRef] [Green Version]
- Engel, M.A.; Leffler, A.; Niedermirtl, F.; Babes, A.; Zimmermann, K.; Filipović, M.R.; Izydorczyk, I.; Eberhardt, M.; Kichko, T.I.; Mueller-Tribbensee, S.M.; et al. TRPA1 and substance P mediate colitis in mice. Gastroenterology 2011, 141, 1346–1358. [Google Scholar] [CrossRef]
- Csekő, K.; Beckers, B.; Keszthelyi, D.; Helyes, Z. Role of TRPV1 and TRPA1 Ion Channels in Inflammatory Bowel Diseases: Potential Therapeutic Targets? Pharmaceuticals 2019, 12, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cojocaru, F.; Şelescu, T.; Domocoş, D.; Măruţescu, L.; Chiritoiu, G.; Chelaru, N.R.; Dima, S.; Mihăilescu, D.; Babes, A.; Cucu, D. Functional expression of the transient receptor potential ankyrin type 1 channel in pancreatic adenocarcinoma cells. Sci. Rep. 2021, 11, 2018. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.T.; Yen, S.L.; Li, C.F.; Chan, T.C.; Chen, T.J.; Lee, S.W.; He, H.L.; Chang, I.W.; Hsing, C.H.; Shiue, Y.L. Overexpression of Transient Receptor Protein Cation Channel Subfamily A Member 1, Confers an Independent Prognostic Indicator in Nasopharyngeal Carcinoma. J. Cancer 2016, 7, 1181–1188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, K.; Shen, B.; Jiang, F.; Xia, L.; Fan, T.; Qin, M.; Yang, L.; Guo, J.; Li, Y.; Zhu, M.; et al. TRPP2 Enhances Metastasis by Regulating Epithelial-Mesenchymal Transition in Laryngeal Squamous Cell Carcinoma. Cell. Physiol. Biochem. 2016, 39, 2203–2215. [Google Scholar] [CrossRef]
- Szallasi, A.; Cortright, D.N.; Blum, C.A.; Eid, S.R. The vanilloid receptor TRPV1: 10 years from channel cloning to antagonist proof-of-concept. Nat. Rev. Drug Discov. 2007, 6, 357–372. [Google Scholar] [CrossRef]
- Birder, L.A.; Kanai, A.J.; de Groat, W.C.; Kiss, S.; Nealen, M.L.; Burke, N.E.; Dineley, K.E.; Watkins, S.; Reynolds, I.J.; Caterina, M.J. Vanilloid receptor expression suggests a sensory role for urinary bladder epithelial cells. Proc. Natl. Acad. Sci. USA 2001, 98, 13396–13401. [Google Scholar] [CrossRef] [Green Version]
- McGarvey, L.P.; Butler, C.A.; Stokesberry, S.; Polley, L.; McQuaid, S.; Abdullah, H.; Ashraf, S.; McGahon, M.K.; Curtis, T.M.; Arron, J.; et al. Increased expression of bronchial epithelial transient receptor potential vanilloid 1 channels in patients with severe asthma. J. Allergy Clin. Immunol. 2014, 133, 704–712. [Google Scholar] [CrossRef] [Green Version]
- Tóth, B.I.; Géczy, T.; Griger, Z.; Dózsa, A.; Seltmann, H.; Kovács, L.; Nagy, L.; Zouboulis, C.C.; Paus, R.; Bíró, T. Transient receptor potential vanilloid-1 signaling as a regulator of human sebocyte biology. J. Investig. Dermatol. 2009, 129, 329–339. [Google Scholar] [CrossRef]
- Marincsák, R.; Tóth, B.I.; Czifra, G.; Márton, I.; Rédl, P.; Tar, I.; Tóth, L.; Kovács, L.; Bíró, T. Increased expression of TRPV1 in squamous cell carcinoma of the human tongue. Oral Dis. 2009, 15, 328–335. [Google Scholar] [CrossRef]
- Virk, H.S.; Rekas, M.Z.; Biddle, M.S.; Wright, A.K.A.; Sousa, J.; Weston, C.A.; Chachi, L.; Roach, K.M.; Bradding, P. Validation of antibodies for the specific detection of human TRPA1. Sci. Rep. 2019, 9, 18500. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Danjo, A.; Kajiya, H.; Okabe, K.; Kido, M.A. Oral epithelial cells are activated via TRP channels. J. Dent. Res. 2011, 90, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Wang, F.; Yang, Y.; Ma, W.; Lin, Z.; Cheng, N.; Long, Y.; Deng, S.; Li, Z. Recurrent activations of transient receptor potential vanilloid-1 and vanilloid-4 promote cellular proliferation and migration in esophageal squamous cell carcinoma cells. FEBS Open Bio 2019, 9, 206–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.Y.; Liu, X.Y.; Zhuo, D.X.; Huang, H.B.; Zhang, F.B.; Liao, S.F. Decreased expression of TRPV1 in renal cell carcinoma: Association with tumor Fuhrman grades and histopathological subtypes. Cancer Manag. Res. 2018, 10, 1647–1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozano, C.; Córdova, C.; Marchant, I.; Zúñiga, R.; Ochova, P.; Ramírez-Barrantes, R.; González-Arriagada, W.A.; Rodriguez, B.; Olivero, P. Intracellular aggregated TRPV1 is associated with lower survival in breast cancer patients. Breast Cancer Targets Ther. 2018, 10, 161–168. [Google Scholar] [CrossRef] [Green Version]
- Vancauwenberghe, E.; Noyer, L.; Derouiche, S.; Lemonnier, L.; Gosset, P.; Sadofsky, L.R.; Mariot, P.; Warnier, M.; Bokhobza, A.; Slomianny, C.; et al. Activation of mutated TRPA1 ion channel by resveratrol in human prostate cancer associated fibroblasts (CAF). Mol. Carcinog. 2017, 56, 1851–1867. [Google Scholar] [CrossRef]
- Takahashi, N.; Matsuda, Y.; Yamada, H.; Tabeta, K.; Nakajima, T.; Murakami, S.; Yamazaki, K. Epithelial TRPV1 signaling accelerates gingival epithelial cell proliferation. J. Dent. Res. 2014, 93, 1141–1147. [Google Scholar] [CrossRef]
- Kemény, Á.; Kodji, X.; Horváth, S.; Komlódi, R.; Szőke, É.; Sándor, Z.; Perkecz, A.; Gyömörei, C.; Sétáló, G.; Kelemen, B.; et al. TRPA1 Acts in a Protective Manner in Imiquimod-Induced Psoriasiform Dermatitis in Mice. J. Investig. Dermatol. 2018, 138, 1774–1784. [Google Scholar] [CrossRef] [Green Version]
- Park, M.; Naidoo, A.A.; Burns, A.; Choi, J.K.; Gatfield, K.M.; Vidgeon-Hart, M.; Bae, I.H.; Lee, C.S.; Choi, G.; Powell, A.J.; et al. Do TRPV1 antagonists increase the risk for skin tumourigenesis? A collaborative in vitro and in vivo assessment. Cell Biol. Toxicol. 2018, 34, 143–162. [Google Scholar] [CrossRef]
- Qin, G.; Li, P.; Xue, Z. Effect of allyl isothiocyanate on the viability and apoptosis of the human cervical cancer HeLa cell line in vitro. Oncol. Lett. 2018, 15, 8756–8760. [Google Scholar] [CrossRef]
- Sávio, A.L.; da Silva, G.N.; Salvadori, D.M. Inhibition of bladder cancer cell proliferation by allyl isothiocyanate (mustard essential oil). Mutat. Res. 2015, 771, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Meral, O.; Alpay, M.; Kismali, G.; Kosova, F.; Cakir, D.U.; Pekcan, M.; Yigit, S.; Sel, T. Capsaicin inhibits cell proliferation by cytochrome c release in gastric cancer cells. Tumour Biol. 2014, 35, 6485–6492. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Behray, M.; Wang, Q.; Wang, W.; Zhou, Z.; Chao, Y.; Bao, Y. Anti-cancer activities of allyl isothiocyanate and its conjugated silicon quantum dots. Sci. Rep. 2018, 8, 1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thejass, P.; Kuttan, G. Inhibition of endothelial cell differentiation and proinflammatory cytokine production during angiogenesis by allyl isothiocyanate and phenyl isothiocyanate. Integr. Cancer Ther. 2007, 6, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Bo, P.; Lien, J.C.; Chen, Y.Y.; Yu, F.S.; Lu, H.F.; Yu, C.S.; Chou, Y.C.; Yu, C.C.; Chung, J.G. Allyl Isothiocyanate Induces Cell Toxicity by Multiple Pathways in Human Breast Cancer Cells. Am. J. Chin. Med. 2016, 44, 415–437. [Google Scholar] [CrossRef] [PubMed]
- Kiss, F.; Pohóczky, K.; Szállási, A.; Helyes, Z. Transient Receptor Potential (TRP) Channels in Head-and-Neck Squamous Cell Carcinomas: Diagnostic, Prognostic, and Therapeutic Potentials. Int. J. Mol. Sci. 2020, 21, 6374. [Google Scholar] [CrossRef]
- Czaja, K.; Burns, G.A.; Ritter, R.C. Capsaicin-induced neuronal death and proliferation of the primary sensory neurons located in the nodose ganglia of adult rats. Neuroscience 2008, 154, 621–630. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, S.F.; Stock, C. Ion channels and transporters in cancer: Pathophysiology, regulation, and clinical potential. Cancer Res. 2013, 73, 1658–1661. [Google Scholar] [CrossRef] [Green Version]
- Berndt, A.; Hyckel, P.; Könneker, A.; Katenkamp, D.; Kosmehl, H. Oral squamous cell carcinoma invasion is associated with a laminin-5 matrix re-organization but independent of basement membrane and hemidesmosome formation. clues from an in vitro invasion model. Invasion Metastasis 1997, 17, 251–258. [Google Scholar]
- Iyer, G.; Wang, A.R.; Brennan, S.R.; Bourgeois, S.; Armstrong, E.; Shah, P.; Harari, P.M. Identification of stable housekeeping genes in response to ionizing radiation in cancer research. Sci. Rep. 2017, 7, 43763. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Localization | Sex | Age | Comorbidities | Smoking | Alcohol |
|---|---|---|---|---|---|---|
| OSCC | tongue, floor of oral cavity | F | 69 | HT | yes | no |
| OSCC | floor of oral cavity | M | 65 | No | yes | yes |
| OSCC | bucca | F | 57 | RA, OP | no | no |
| OSCC | floor of oral cavity | M | 92 | DM | no | no |
| OSCC | tongue | M | 67 | CLL, HT | yes | yes |
| OSCC | tongue | M | 67 | pancreatitis | yes | yes |
| OSCC | floor of oral cavity | F | 56 | no | yes | yes |
| OSCC | tongue | M | 53 | epilepsy | yes | yes |
| OSCC | lower lip | M | 58 | HT, paraplegia | no | no |
| OSCC | floor of oral cavity | F | 68 | no | yes | no |
| OSCC | gingiva | F | 63 | no | yes | no |
| OSCC | sublingual region, tongue | M | 63 | no | yes | yes |
| OSCC | tongue | F | 82 | HT | no | no |
| OSCC | hard palate | M | 57 | HT | yes | no |
| OSCC | bucca | M | 54 | no | yes | yes |
| normal tissue | bucca | M | 64 | |||
| normal tissue | bucca | F | 33 | |||
| normal tissue | lower lip | F | 45 | |||
| normal tissue | gingiva | F | 59 | |||
| normal tissue | bucca | F | 70 | |||
| normal tissue | gingiva | M | 65 | |||
| normal tissue | lower lip | F | 47 | |||
| normal tissue | lower lip | M | 52 | |||
| normal tissue | tongue | F | 53 | |||
| normal tissue | tongue | F | 69 |
| Type | Localization | Sex | Age |
|---|---|---|---|
| OSCC | tongue | M | 80 |
| OSCC | gingiva | M | 53 |
| OSCC | gingiva | M | 56 |
| OSCC | tongue | M | 68 |
| OSCC | hard palate | M | 57 |
| OSCC | tongue | F | 72 |
| normal tissue | tongue | M | 66 |
| normal tissue | gingiva | F | 22 |
| normal tissue | gingiva | M | 29 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiss, F.; Kormos, V.; Szőke, É.; Kecskés, A.; Tóth, N.; Steib, A.; Szállási, Á.; Scheich, B.; Gaszner, B.; Kun, J.; et al. Functional Transient Receptor Potential Ankyrin 1 and Vanilloid 1 Ion Channels Are Overexpressed in Human Oral Squamous Cell Carcinoma. Int. J. Mol. Sci. 2022, 23, 1921. https://doi.org/10.3390/ijms23031921
Kiss F, Kormos V, Szőke É, Kecskés A, Tóth N, Steib A, Szállási Á, Scheich B, Gaszner B, Kun J, et al. Functional Transient Receptor Potential Ankyrin 1 and Vanilloid 1 Ion Channels Are Overexpressed in Human Oral Squamous Cell Carcinoma. International Journal of Molecular Sciences. 2022; 23(3):1921. https://doi.org/10.3390/ijms23031921
Chicago/Turabian StyleKiss, Fruzsina, Viktória Kormos, Éva Szőke, Angéla Kecskés, Norbert Tóth, Anita Steib, Árpád Szállási, Bálint Scheich, Balázs Gaszner, József Kun, and et al. 2022. "Functional Transient Receptor Potential Ankyrin 1 and Vanilloid 1 Ion Channels Are Overexpressed in Human Oral Squamous Cell Carcinoma" International Journal of Molecular Sciences 23, no. 3: 1921. https://doi.org/10.3390/ijms23031921
APA StyleKiss, F., Kormos, V., Szőke, É., Kecskés, A., Tóth, N., Steib, A., Szállási, Á., Scheich, B., Gaszner, B., Kun, J., Fülöp, G., Pohóczky, K., & Helyes, Z. (2022). Functional Transient Receptor Potential Ankyrin 1 and Vanilloid 1 Ion Channels Are Overexpressed in Human Oral Squamous Cell Carcinoma. International Journal of Molecular Sciences, 23(3), 1921. https://doi.org/10.3390/ijms23031921