
Carotenoids from Marine Sources as a New Approach in Neuroplasticity Enhancement

 , , , and
, , , and 
Abstract
:1. Introduction
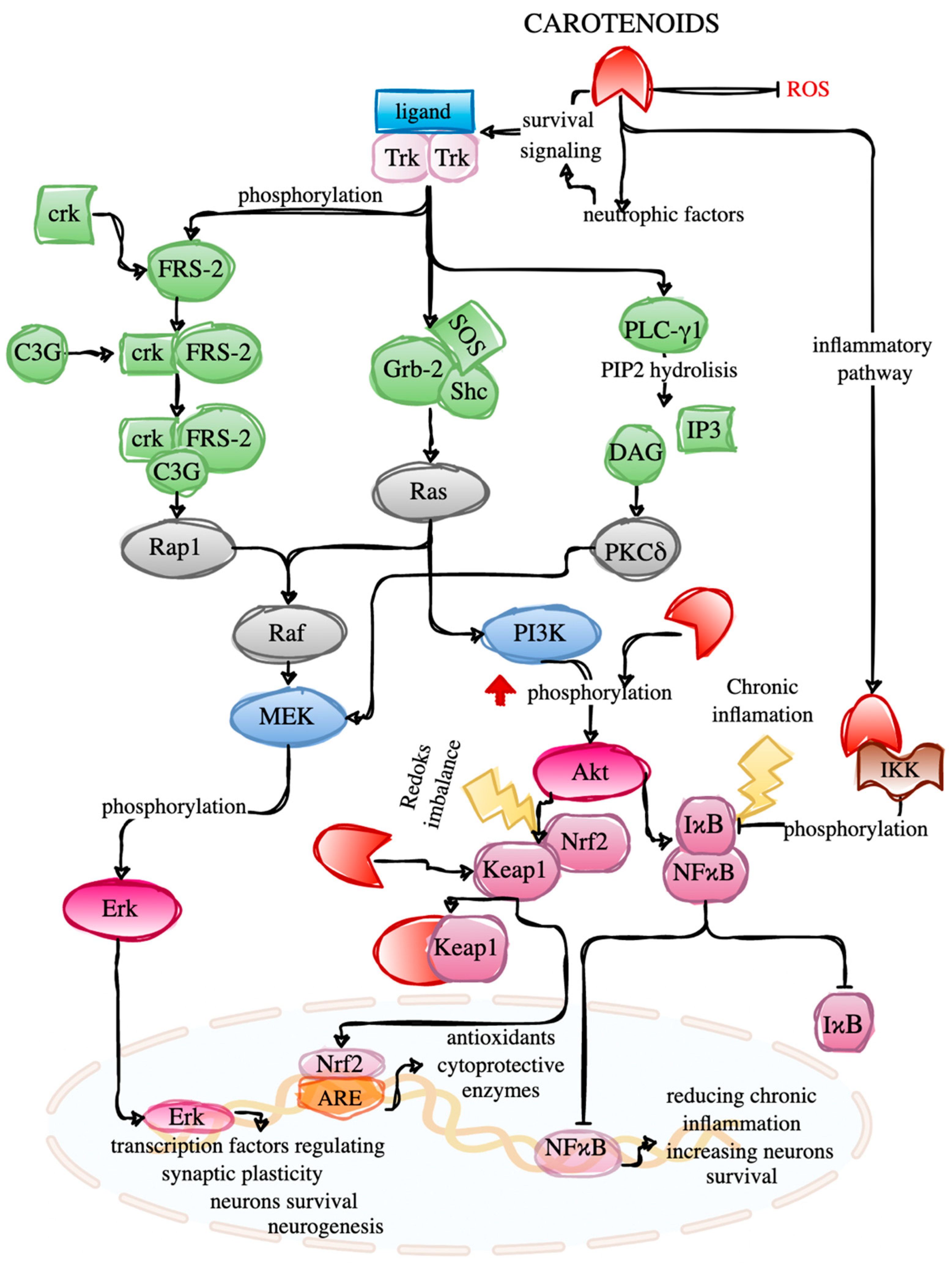
2. Neurorestorative Actions of Marine Carotenoids
3. Marine Carotenoids
3.1. Fucoxanthin
3.2. Astaxanthin
3.3. Siphonaxanthin
3.4. Mytiloxanthin
3.5. Saproxanthin and Myxol
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Davinelli, S.; Ali, S.; Solfrizzi, V.; Scapagnini, G.; Corbi, G. Carotenoids and Cognitive Outcomes: A Meta-Analysis of Randomized Intervention Trials. Antioxidants 2021, 10, 223. [Google Scholar] [CrossRef]
- Mateos-Aparicio, P.; Rodríguez-Moreno, A. The Impact of Studying Brain Plasticity. Front. Cell. Neurosci. 2019, 13, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, M.v.; Ishida, A.; Ishida, W.N.; Matsushita, H.B.; Nishimura, A.; Tsuji, M. Plasticity and Injury in the Developing Brain. Brain Dev. 2009, 31, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katsimpardi, L.; Lledo, P.M. Regulation of Neurogenesis in the Adult and Aging Brain. Curr. Opin. Neurobiol. 2018, 53, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Niklison-Chirou, M.V.; Agostini, M.; Amelio, I.; Melino, G. Regulation of Adult Neurogenesis in Mammalian Brain. Int. J. Mol. Sci. 2020, 21, 4869. [Google Scholar] [CrossRef]
- Isaev, N.K.; Stelmashook, E.v.; Genrikhs, E.E. Neurogenesis and Brain Aging. Rev. Neurosci. 2019, 30, 573–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourgeois, J.-P. Synaptogenèses et Épigenèses Cérébrales. Méd./Sci. 2005, 21, 428–433. [Google Scholar] [CrossRef] [PubMed]
- Kowiański, P.; Lietzau, G.; Czuba, E.; Waśkow, M.; Steliga, A.; Moryś, J. BDNF: A Key Factor with Multipotent Impact on Brain Signaling and Synaptic Plasticity. Cell. Mol. Neurobiol. 2018, 38, 579. [Google Scholar] [CrossRef]
- Dyer, A.H.; Vahdatpour, C.; Sanfeliu, A.; Tropea, D. The Role of Insulin-Like Growth Factor 1 (IGF-1) in Brain Development, Maturation and Neuroplasticity. Neuroscience 2016, 325, 89–99. [Google Scholar] [CrossRef]
- Zechel, S.; Werner, S.; Unsicker, K.; Halbach, O. von B. und Expression and Functions of Fibroblast Growth Factor 2 (FGF-2) in Hippocampal Formation. Neuroscientist 2010, 16, 357–373. [Google Scholar] [CrossRef]
- Keefe, K.M.; Sheikh, I.S.; Smith, G.M. Targeting Neurotrophins to Specific Populations of Neurons: NGF, BDNF, and NT-3 and Their Relevance for Treatment of Spinal Cord Injury. Int. J. Mol. Sci. 2017, 18, 548. [Google Scholar] [CrossRef] [PubMed]
- Carter, A.R.; Chen, C.; Schwartz, P.M.; Segal, R.A. Brain-Derived Neurotrophic Factor Modulates Cerebellar Plasticity and Synaptic Ultrastructure. J. Neurosci. 2002, 22, 1316. [Google Scholar] [CrossRef] [PubMed]
- Uren, R.T.; Turnley, A.M. Regulation of Neurotrophin Receptor (Trk) Signaling: Suppressor of Cytokine Signaling 2 (SOCS2) Is a New Player. Front. Mol. Neurosci. 2014, 7, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanisevic, L.; Saragovi, H.U. Neurotrophins. Handb. Biol. Act. Pept. 2013, 1639–1646. [Google Scholar] [CrossRef]
- Huang, E.J.; Reichardt, L.F. Trk Receptors: Roles in Neuronal Signal Transduction. Annu. Rev. Biochem. 2003, 72, 609–642. [Google Scholar] [CrossRef] [Green Version]
- Meeker, R.B.; Williams, K.S. The P75 Neurotrophin Receptor: At the Crossroad of Neural Repair and Death. Neural Regen. Res. 2015, 10, 721. [Google Scholar] [CrossRef]
- Simanshu, D.K.; Nissley, D.v.; McCormick, F. RAS Proteins and Their Regulators in Human Disease. Cell 2017, 170, 17. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Alegría, K.; Flores-León, M.; Avila-Muñoz, E.; Rodríguez-Corona, N.; Arias, C. PI3K Signaling in Neurons: A Central Node for the Control of Multiple Functions. Int. J. Mol. Sci. 2018, 19, 3725. [Google Scholar] [CrossRef] [Green Version]
- Hemmings, B.A.; Restuccia, D.F. PI3K-PKB/Akt Pathway. Cold Spring Harb. Perspect. Biol. 2012, 4, a011189. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-ΚB Signaling in Inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Cantley, L.C. The Phosphoinositide 3-Kinase Pathway. Science 2002, 296, 1655–1657. [Google Scholar] [CrossRef] [PubMed]
- Vetter, M.L.; Martin-Zanca, D.; Parada, L.F.; Bishop, J.M.; Kaplan, D.R. Nerve Growth Factor Rapidly Stimulates Tyrosine Phosphorylation of Phospholipase C-Gamma 1 by a Kinase Activity Associated with the Product of the Trk Protooncogene. Proc. Natl. Acad. Sci. USA 1991, 88, 5650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reichardt, L.F. Neurotrophin-Regulated Signalling Pathways. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 1545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbit, K.C.; Foster, D.A.; Rosner, M.R. Protein Kinase Cδ Mediates Neurogenic but Not Mitogenic Activation of Mitogen-Activated Protein Kinase in Neuronal Cells. Mol. Cell. Biol. 1999, 19, 4209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Britton, G. Structure and Properties of Carotenoids in Relation to Function. FASEB J. 1995, 9, 1551–1558. [Google Scholar] [CrossRef]
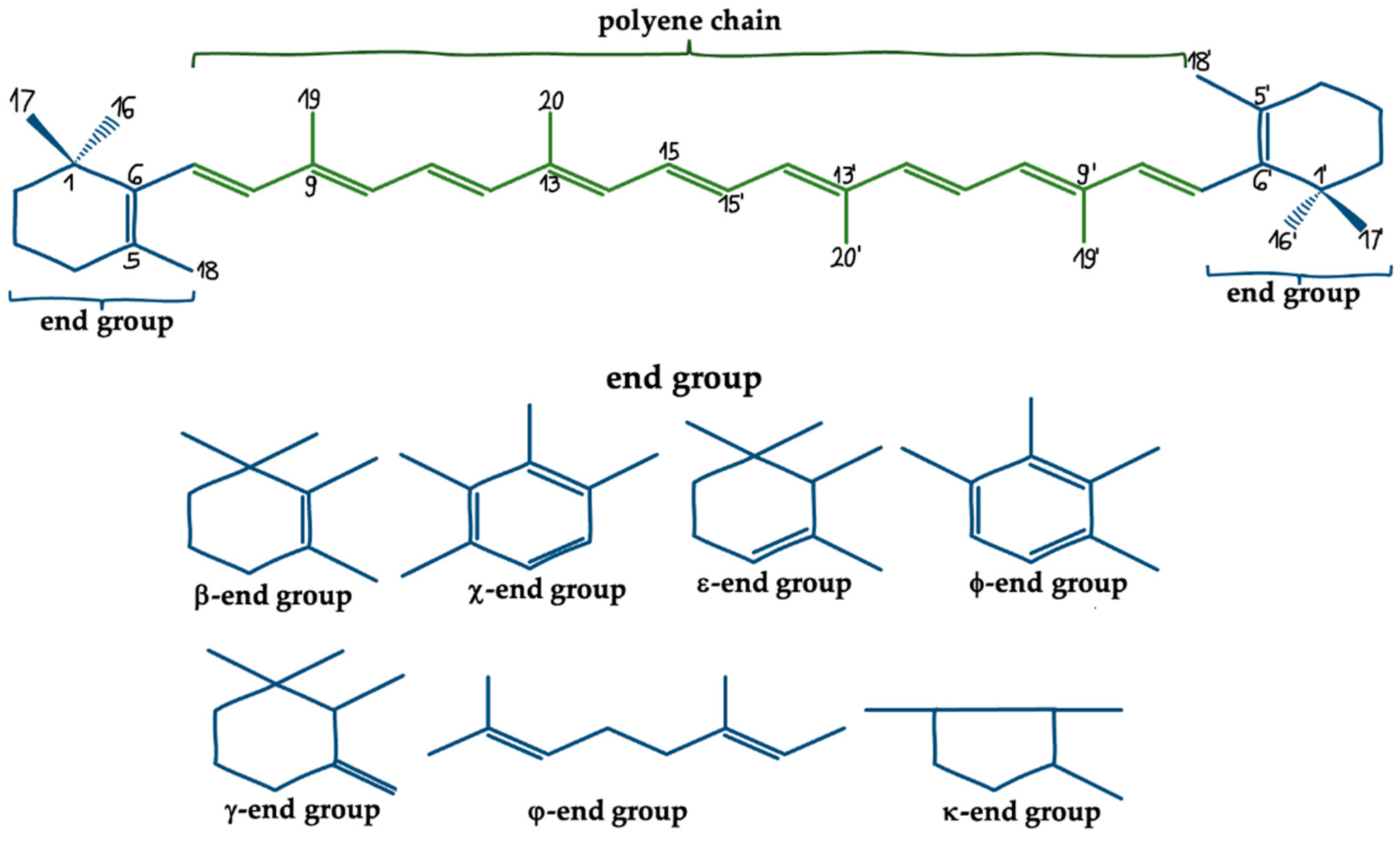
- Meléndez-Martínez, A.J.; Mapelli-Brahm, P.; Hornero-Méndez, D.; Vicario, I.M. CHAPTER 1 Structures, Nomenclature and General Chemistry of Carotenoids and Their Esters. Food Chem. Funct. Anal. 2019, 1–50. [Google Scholar] [CrossRef]
- Katayama, S.; Ogawa, H.; Nakamura, S. Apricot Carotenoids Possess Potent Anti-Amyloidogenic Activity in Vitro. J. Agric. Food Chem. 2011, 59, 12691–12696. [Google Scholar] [CrossRef]
- Grimmig, B.; Kim, S.H.; Nash, K.; Bickford, P.C.; Douglas Shytle, R. Neuroprotective Mechanisms of Astaxanthin: A Potential Therapeutic Role in Preserving Cognitive Function in Age and Neurodegeneration. GeroScience 2017, 39, 19. [Google Scholar] [CrossRef]
- Chisté, R.C.; Freitas, M.; Mercadante, A.Z.; Fernandes, E. Carotenoids Inhibit Lipid Peroxidation and Hemoglobin Oxidation, but Not the Depletion of Glutathione Induced by ROS in Human Erythrocytes. Life Sci. 2014, 99, 52–60. [Google Scholar] [CrossRef]
- Shaish, A.; Harari, A.; Kamari, Y.; Soudant, E.; Harats, D.; Ben-Amotz, A. A Carotenoid Algal Preparation Containing Phytoene and Phytofluene Inhibited LDL Oxidation In Vitro. Plant Foods Hum. Nutr. 2008, 63, 83–86. [Google Scholar] [CrossRef]
- Pérez-Gálvez, A.; Mínguez-Mosquera, M.I. Esterification of Xanthophylls and Its Effect on Chemical Behavior and Bioavailability of Carotenoids in the Human. Nutr. Res. 2005, 25, 631–640. [Google Scholar] [CrossRef]
- Papandreou, M.A.; Kanakis, C.D.; Polissiou, M.G.; Efthimiopoulos, S.; Cordopatis, P.; Margarity, M.; Lamari, F.N. Inhibitory Activity on Amyloid-β Aggregation and Antioxidant Properties of Crocus Sativus Stigmas Extract and Its Crocin Constituents. J. Agric. Food Chem. 2006, 54, 8762–8768. [Google Scholar] [CrossRef] [PubMed]
- Stahl, W.; Sies, H. Bioactivity and Protective Effects of Natural Carotenoids. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2005, 1740, 101–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaulmann, A.; Bohn, T. Carotenoids, Inflammation, and Oxidative Stress—Implications of Cellular Signaling Pathways and Relation to Chronic Disease Prevention. Nutr. Res. 2014, 34, 907–929. [Google Scholar] [CrossRef]
- Woodall, A.A.; Lee, S.W.M.; Weesie, R.J.; Jackson, M.J.; Britton, G. Oxidation of Carotenoids by Free Radicals: Relationship between Structure and Reactivity. Biochim. Biophys. Acta (BBA)-Gen. Subj. 1997, 1336, 33–42. [Google Scholar] [CrossRef]
- Ben-Dor, A.; Steiner, M.; Gheber, L.; Danilenko, M.; Dubi, N.; Linnewiel, K.; Zick, A.; Sharoni, Y.; Levy, J. Carotenoids Activate the Antioxidant Response Element Transcription System. Mol. Cancer Ther. 2005, 4, 177–186. [Google Scholar]
- Palozza, P.; Serini, S.; Torsello, A.; di Nicuolo, F.; Piccioni, E.; Ubaldi, V.; Pioli, C.; Wolf, F.I.; Calviello, G. β-Carotene Regulates NF-ΚB DNA-Binding Activity by a Redox Mechanism in Human Leukemia and Colon Adenocarcinoma Cells. J. Nutr. 2003, 133, 381–388. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Cha, Y.N.; Surh, Y.J. A Protective Role of Nuclear Factor-Erythroid 2-Related Factor-2 (Nrf2) in Inflammatory Disorders. Mutat. Res./Fundam. Mol. Mech. Mutagenes. 2010, 690, 12–23. [Google Scholar] [CrossRef]
- Kansanen, E.; Kuosmanen, S.M.; Leinonen, H.; Levonenn, A.L. The Keap1-Nrf2 Pathway: Mechanisms of Activation and Dysregulation in Cancer. Redox Biol. 2013, 1, 45–49. [Google Scholar] [CrossRef] [Green Version]
- Jung, K.A.; Kwak, M.K. The Nrf2 System as a Potential Target for the Development of Indirect Antioxidants. Molecules 2010, 15, 7266–7291. [Google Scholar] [CrossRef] [Green Version]
- Piovan, A.; Filippini, R.; Corbioli, G.; Costa, V.D.; Giunco, E.M.V.; Burbello, G.; Pagetta, A.; Giusti, P.; Zusso, M. Carotenoid Extract Derived from Euglena Gracilis Overcomes Lipopolysaccharide-Induced Neuroinflammation in Microglia: Role of NF-ΚB and Nrf2 Signaling Pathways. Mol. Neurobiol. 2021, 58, 3515. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Gan, D.; Fan, C.; Wen, C.; Li, A.; Li, Q.; Zhao, J.; Wang, Z.; Zhu, L.; Lu, D. The Secretion from Neural Stem Cells Pretreated with Lycopene Protects against Tert-Butyl Hydroperoxide-Induced Neuron Oxidative Damage. Oxidative Med. Cell. Longev. 2018, 2018, 5490218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.A.; Hayden, M.M.; Bannerman, S.; Jansen, J.; Crowe-White, K.M. Anti-Apoptotic Effects of Carotenoids in Neurodegeneration. Molecules 2020, 25, 3453. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Ren, B.; Guo, R.; Zhang, W.; Ma, S.; Yao, Y.; Yuan, T.; Liu, Z.; Liu, X. Supplementation of Lycopene Attenuates Oxidative Stress Induced Neuroinflammation and Cognitive Impairment via Nrf2/NF-ΚB Transcriptional Pathway. Food Chem. Toxicol. 2017, 109, 505–516. [Google Scholar] [CrossRef]
- Thomas, G.M.; Huganir, R.L. MAPK Cascade Signalling and Synaptic Plasticity. Nat. Rev. Neurosci. 2004, 5, 173–183. [Google Scholar] [CrossRef]
- Besnard, A.; Galan-Rodriguez, B.; Vanhoutte, P.; Caboche, J. Elk-1 a Transcription Factor with Multiple Facets in the Brain. Front. Neurosci. 2011, 5, 35. [Google Scholar] [CrossRef] [Green Version]
- Cattaneo, A.; Cattane, N.; Begni, V.; Pariante, C.M.; Riva, M.A. The Human BDNF Gene: Peripheral Gene Expression and Protein Levels as Biomarkers for Psychiatric Disorders. Transl. Psychiatry 2016, 6, e958. [Google Scholar] [CrossRef]
- Dincheva, I.; Lynch, N.B.; Lee, F.S. The Role of BDNF in the Development of Fear Learning. Depress. Anxiety 2016, 33, 907. [Google Scholar] [CrossRef] [Green Version]
- Rocco, M.L.; Soligo, M.; Manni, L.; Aloe, L. Nerve Growth Factor: Early Studies and Recent Clinical Trials. Curr. Neuropharmacol. 2018, 16, 1455. [Google Scholar] [CrossRef]
- Stringham, N.T.; Holmes, P.V.; Stringham, J.M. Lutein Supplementation Increases Serum Brain-Derived Neurotrophic Factor (BDNF) in Humans. FASEB J. 2016, 30, 689.3. [Google Scholar] [CrossRef]
- Prakash, A.; Kumar, A. Implicating the Role of Lycopene in Restoration of Mitochondrial Enzymes and BDNF Levels in β-Amyloid Induced Alzheimer’s Disease. Eur. J. Pharmacol. 2014, 741, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Sahin, K.; Orhan, C.; Tuzcu, M.; Sahin, N.; Juturu, V. Regular Exercise Training with Lutein/Zeaxanthin Isomers Regulates Brain Transcription Factors and Neurotrophic and Synaptic Proteins in Rats (P06-020-19). Curr. Dev. Nutr. 2019, 3. [Google Scholar] [CrossRef]
- Dong, Y.L.; Pu, K.J.; Duan, W.J.; Chen, H.C.; Chen, L.X.; Wang, Y.M. Involvement of Akt/CREB Signaling Pathways in the Protective Effect of EPA against Interleukin-1β-Induced Cytotoxicity and BDNF down-Regulation in Cultured Rat Hippocampal Neurons. BMC Neurosci. 2018, 19, 52. [Google Scholar] [CrossRef] [PubMed]
- Putcha, G.v.; Moulder, K.L.; Golden, J.P.; Bouillet, P.; Adams, J.A.; Strasser, A.; Johnson, E.M. Induction of BIM, a Proapoptotic BH3-Only BCL-2 Family Member, Is Critical for Neuronal Apoptosis. Neuron 2001, 29, 615–628. [Google Scholar] [CrossRef] [Green Version]
- Biswas, S.C.; Greene, L.A. Nerve Growth Factor (NGF) Down-Regulates the Bcl-2 Homology 3 (BH3) Domain-Only Protein Bim and Suppresses Its Proapoptotic Activity by Phosphorylation. J. Biol. Chem. 2002, 277, 49511–49516. [Google Scholar] [CrossRef] [Green Version]
- Jomova, K.; Valko, M. Health Protective Effects of Carotenoids and Their Interactions with Other Biological Antioxidants. Eur. J. Med. Chem. 2013, 70, 102–110. [Google Scholar] [CrossRef]
- Wen, X.; Huang, A.; Hu, J.; Zhong, Z.; Liu, Y.; Li, Z.; Pan, X.; Liu, Z. Neuroprotective Effect of Astaxanthin against Glutamate-Induced Cytotoxicity in HT22 Cells: Involvement of the Akt/GSK-3β Pathway. Neuroscience 2015, 303, 558–568. [Google Scholar] [CrossRef]
- Xu, L.; Zhu, J.; Yin, W.; Ding, X. Astaxanthin Improves Cognitive Deficits from Oxidative Stress, Nitric Oxide Synthase and Inflammation through Upregulation of PI3K/Akt in Diabetes Rat. Int. J. Clin. Exp. Pathol. 2015, 8, 6083. [Google Scholar]
- Lu, Y.; Wang, X.; Feng, J.; Xie, T.; Si, P.; Wang, W. Neuroprotective Effect of Astaxanthin on Newborn Rats Exposed to Prenatal Maternal Seizures. Brain Res. Bull. 2019, 148, 63–69. [Google Scholar] [CrossRef]
- Xiang, S.; Liu, F.; Lin, J.; Chen, H.; Huang, C.; Chen, L.; Zhou, Y.; Ye, L.; Zhang, K.; Jin, J.; et al. Fucoxanthin Inhibits β-Amyloid Assembly and Attenuates β-Amyloid Oligomer-Induced Cognitive Impairments. J. Agric. Food Chem. 2017, 65, 4092–4102. [Google Scholar] [CrossRef]
- Johnson, E.J.; Vishwanathan, R.; Schalch, W.; Poon, L.; Wittwer, J.; Johnson, M.A.; Hausman, D.; Davey, A.; Green, R.; Gearing, M.; et al. Brain Levels of Lutein (L) and Zeaxanthin (Z) Are Related to Cognitive Function in Centenarians. FASEB J. 2011, 25, 975.21. [Google Scholar] [CrossRef]
- Zhou, L.; Ouyang, L.; Lin, S.; Chen, S.; Liu, Y.J.; Zhou, W.; Wang, X. Protective Role of β-Carotene against Oxidative Stress and Neuroinflammation in a Rat Model of Spinal Cord Injury. Int. Immunopharmacol. 2018, 61, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.; Xu, N.; Ma, T.; Liu, Y.; Xu, H.; Lu, Y. Anti-Inflammatory Effect of Lycopene on Experimental Spinal Cord Ischemia Injury via Cyclooxygenase-2 Suppression. Neuroimmunomodulation 2019, 26, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Rhim, H. Therapeutic Implication of Autophagy in Neurodegenerative Diseases. BMB Rep. 2017, 50, 345. [Google Scholar] [CrossRef] [Green Version]
- Fung, F.K.C.; Law, B.Y.K.; Lo, A.C.Y. Lutein Attenuates Both Apoptosis and Autophagy upon Cobalt (II) Chloride-Induced Hypoxia in Rat Műller Cells. PLoS ONE 2016, 11, e0167828. [Google Scholar] [CrossRef]
- Chang, C.J.; Lin, J.F.; Hsiao, C.Y.; Chang, H.H.; Li, H.J.; Chang, H.H.; Lee, G.A.; Hung, C.F. Lutein Induces Autophagy via Beclin-1 Upregulation in IEC-6 Rat Intestinal Epithelial Cells. Am. J. Chin. Med. 2017, 45, 1273–1291. [Google Scholar] [CrossRef]
- Zeng, C.; Li, H.; Fan, Z.; Zhong, L.; Guo, Z.; Guo, Y.; Xi, Y. Crocin-Elicited Autophagy Rescues Myocardial Ischemia/Reperfusion Injury via Paradoxical Mechanisms. Am. J. Chin. Med. 2016, 44, 515–530. [Google Scholar] [CrossRef]
- Shen, M.; Chen, K.; Lu, J.; Cheng, P.; Xu, L.; Dai, W.; Wang, F.; He, L.; Zhang, Y.; Chengfen, W.; et al. Protective Effect of Astaxanthin on Liver Fibrosis through Modulation of TGF- β 1 Expression and Autophagy. Mediat. Inflamm. 2014, 2014, 954502. [Google Scholar] [CrossRef] [Green Version]
- Buyuklu, M.; Kandemir, F.M.; Ozkaraca, M.; Set, T.; Bakirci, E.M.; Topal, E.; Ileriturk, M.; Turkmen, K. Benefical Effects of Lycopene against Contrast Medium-Induced Oxidative Stress, Inflammation, Autophagy, and Apoptosis in Rat Kidney. Hum. Exp. Toxicol. 2015, 34, 487–496. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, H.; Fan, Y.; Gao, Y.; Li, X.; Hu, Z.; Ding, K.; Wang, Y.; Wang, X. Fucoxanthin Provides Neuroprotection in Models of Traumatic Brain Injury via the Nrf2-ARE and Nrf2-Autophagy Pathways. Sci. Rep. 2017, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Maoka, T. Carotenoids as Natural Functional Pigments. J. Nat. Med. 2020, 74, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Xu, Y.; Sun, X.; Wang, S.; Wang, J.; Zhu, J.; Wang, D.; Zhao, L. Stability, Bioactivity, and Bioaccessibility of Fucoxanthin in Zein-Caseinate Composite Nanoparticles Fabricated at Neutral PH by Antisolvent Precipitation. Food Hydrocoll. 2018, 84, 379–388. [Google Scholar] [CrossRef]
- Huang, Z.; Xu, L.; Zhu, X.; Hu, J.; Peng, H.; Zeng, Z.; Xiong, H. Stability and Bioaccessibility of Fucoxanthin in Nanoemulsions Prepared from Pinolenic Acid-Contained Structured Lipid. Int. J. Food Eng. 2017, 13. [Google Scholar] [CrossRef] [Green Version]
- Effect of an Antioxidants Mix on Cognitive Performance and Well Being: The Bacopa, Licopene, Astaxantina, Vitamin B12-Full Text View-ClinicalTrials.Gov. Available online: https://clinicaltrials.gov/ct2/show/NCT03825042?term=astaxanthin&draw=3&rank=23 (accessed on 8 November 2021).
- Effect of Astaxanthin on the Patients with Alzheimer Disease-Full Text View-ClinicalTrials.Gov. Available online: https://clinicaltrials.gov/ct2/show/NCT05015374?term=astaxanthin&draw=2&rank=1 (accessed on 8 November 2021).
- A Randomised, Double-Blind, Placebo-Controlled, Parallel Study of the Effect of BrainPhyt on Cognitive Function in Healthy Older Subjects-Full Text View-ClinicalTrials.Gov. Available online: https://clinicaltrials.gov/ct2/show/NCT04832412?term=fucoxanthin&draw=2&rank=5 (accessed on 8 November 2021).
- Galasso, C.; Corinaldesi, C.; Sansone, C. Carotenoids from Marine Organisms: Biological Functions and Industrial Applications. Antioxidants 2017, 6, 96. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, E.; Mariutti, L.R.B.; Mercadante, A.Z. Scavenging Capacity of Marine Carotenoids against Reactive Oxygen and Nitrogen Species in a Membrane-Mimicking System. Mar. Drugs 2012, 10, 1784. [Google Scholar] [CrossRef]
- Riccioni, G.; D’Orazio, N.; Franceschelli, S.; Speranza, L. Marine Carotenoids and Cardiovascular Risk Markers. Mar. Drugs 2011, 9, 1166. [Google Scholar] [CrossRef] [Green Version]
- D’Orazio, N.; Gammone, M.A.; Gemello, E.; de Girolamo, M.; Cusenza, S.; Riccioni, G. Marine Bioactives: Pharmacological Properties and Potential Applications against Inflammatory Diseases. Mar. Drugs 2012, 10, 812. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.H.; Shin, H.Y.; Park, J.H.; Koo, S.Y.; Kim, S.M.; Yang, S.H. Fucoxanthin from Microalgae Phaeodactylum Tricornutum Inhibits Pro-Inflammatory Cytokines by Regulating Both NF-ΚB and NLRP3 Inflammasome Activation. Sci. Rep. 2021, 11, 1–12. [Google Scholar] [CrossRef]
- Ha, A.W.; Na, S.J.; Kim, W.K. Antioxidant Effects of Fucoxanthin Rich Powder in Rats Fed with High Fat Diet. Nutr. Res. Pract. 2013, 7, 475. [Google Scholar] [CrossRef] [Green Version]
- Rengarajan, T.; Rajendran, P.; Nandakumar, N.; Balasubramanian, M.P.; Nishigaki, I. Cancer Preventive Efficacy of Marine Carotenoid Fucoxanthin: Cell Cycle Arrest and Apoptosis. Nutrients 2013, 5, 4978. [Google Scholar] [CrossRef] [Green Version]
- Das, S.K.; Hashimoto, T.; Kanazawa, K. Growth Inhibition of Human Hepatic Carcinoma HepG2 Cells by Fucoxanthin Is Associated with Down-Regulation of Cyclin D. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2008, 1780, 743–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Miyashita, K. Dietary Combination of Fucoxanthin and Fish Oil Attenuates the Weight Gain of White Adipose Tissue and Decreases Blood Glucose in Obese/Diabetic KK-A y Mice. J. Agric. Food Chem. 2007, 55, 7701–7706. [Google Scholar] [CrossRef] [PubMed]
- Mayer, C.; Côme, M.; Blanckaert, V.; Zittelli, G.C.; Faraloni, C.; Nazih, H.; Ouguerram, K.; Mimouni, V.; Chénais, B. Effect of Carotenoids from Phaeodactylum Tricornutum on Palmitate-Treated HepG2 Cells. Molecules 2020, 25, 2845. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.M.; Li, K.L.; Lin, Y.C. Fucoidan–Fucoxanthin Ameliorated Cardiac Function via IRS1/GRB2/SOS1, GSK3β/CREB Pathways and Metabolic Pathways in Senescent Mice. Mar. Drugs 2019, 17, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, L.; Chen, W.; Tian, F.; Yuan, C.; Wang, H.; Yue, H. Neuroprotective Role of Fucoxanthin against Cerebral Ischemic/Reperfusion Injury through Activation of Nrf2/HO-1 Signaling. Biomed. Pharmacother. 2018, 106, 1484–1489. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Han, H.; Liu, J.; Tang, M.; Wu, X.; Cao, X.; Zhao, T.; Lu, Y.; Niu, T.; Chen, J.; et al. Fucoxanthin Prevents 6-OHDA-Induced Neurotoxicity by Targeting Keap1. Oxidative Med. Cell. Longev. 2021, 2021, 6688708. [Google Scholar] [CrossRef]
- Zhao, D.; Kwon, S.H.; Chun, Y.S.; Gu, M.Y.; Yang, H.O. Anti-Neuroinflammatory Effects of Fucoxanthin via Inhibition of Akt/NF-ΚB and MAPKs/AP-1 Pathways and Activation of PKA/CREB Pathway in Lipopolysaccharide-Activated BV-2 Microglial Cells. Neurochem. Res. 2016, 42, 667–677. [Google Scholar] [CrossRef]
- Lin, J.; Huang, L.; Yu, J.; Xiang, S.; Wang, J.; Zhang, J.; Yan, X.; Cui, W.; He, S.; Wang, Q. Fucoxanthin, a Marine Carotenoid, Reverses Scopolamine-Induced Cognitive Impairments in Mice and Inhibits Acetylcholinesterase in Vitro. Mar. Drugs 2016, 14. [Google Scholar] [CrossRef] [Green Version]
- Higuera-Ciapara, I.; Félix-Valenzuela, L.; Goycoolea, F.M. Astaxanthin: A Review of Its Chemistry and Applications. Crit. Rev. Food Sci. Nutr. 2007, 46, 185–196. [Google Scholar] [CrossRef]
- Brotosudarmo, T.H.P.; Limantara, L.; Setiyono, E. Heriyanto Structures of Astaxanthin and Their Consequences for Therapeutic Application. Int. J. Food Sci. 2020, 2020, 2156582. [Google Scholar] [CrossRef]
- Ambati, R.R.; Moi, P.S.; Ravi, S.; Aswathanarayana, R.G. Astaxanthin: Sources, Extraction, Stability, Biological Activities and Its Commercial Applications—A Review. Mar. Drugs 2014, 12, 128. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Kumar, R.; Kumari, A.; Panwar, A. Astaxanthin: A Super Antioxidant from Microalgae and Its Therapeutic Potential. J. Basic Microbiol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Raza, S.H.A.; Naqvi, S.R.Z.; Abdelnour, S.A.; Schreurs, N.; Mohammedsaleh, Z.M.; Khan, I.; Shater, A.F.; Abd El-Hack, M.E.; Khafaga, A.F.; Quan, G.; et al. Beneficial Effects and Health Benefits of Astaxanthin Molecules on Animal Production: A Review. Res. Vet. Sci. 2021, 138, 69–78. [Google Scholar] [CrossRef]
- Aoi, W.; Naito, Y.; Sakuma, K.; Kuchide, M.; Tokuda, H.; Maoka, T.; Toyokuni, S.; Oka, S.; Yasuhara, M.; Yoshikawa, T. Astaxanthin Limits Exercise-Induced Skeletal and Cardiac Muscle Damage in Mice. Antioxid. Redox Signal. 2004, 5, 139–144. [Google Scholar] [CrossRef]
- Camera, E.; Mastrofrancesco, A.; Fabbri, C.; Daubrawa, F.; Picardo, M.; Sies, H.; Stahl, W. Astaxanthin, Canthaxanthin and β-Carotene Differently Affect UVA-Induced Oxidative Damage and Expression of Oxidative Stress-Responsive Enzymes. Exp. Dermatol. 2009, 18, 222–231. [Google Scholar] [CrossRef]
- Focsan, A.L.; Polyakov, N.E.; Kispert, L.D. Photo Protection of Haematococcus Pluvialis Algae by Astaxanthin: Unique Properties of Astaxanthin Deduced by EPR, Optical and Electrochemical Studies. Antioxidants 2017, 6, 80. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Wang, X.; Xiang, Q.; Meng, X.; Peng, Y.; Du, N.; Liu, Z.; Sun, Q.; Wang, C.; Liu, X. Astaxanthin Alleviates Brain Aging in Rats by Attenuating Oxidative Stress and Increasing BDNF Levels. Food Funct. 2013, 5, 158–166. [Google Scholar] [CrossRef]
- Aslankoc, R.; Ozmen, O.; Yalcın, A. Astaxanthin Ameliorates Damage to the Cerebral Cortex, Hippocampus and Cerebellar Cortex Caused by Methotrexate. Biotech. Histochem. 2021, 1–12. [Google Scholar] [CrossRef]
- Zhao, L.; Tao, X.; Song, T. Astaxanthin Alleviates Neuropathic Pain by Inhibiting the MAPKs and NF-ΚB Pathways. Eur. J. Pharmacol. 2021, 912, 174575. [Google Scholar] [CrossRef]
- Emad El-Agamy, S.; Kamal Abdel-Aziz, A.; Wahdan, S.; Esmat, A.; Azab, S.S. Astaxanthin Ameliorates Doxorubicin-Induced Cognitive Impairment (Chemobrain) in Experimental Rat Model: Impact on Oxidative, Inflammatory, and Apoptotic Machineries. Mol. Neurobiol. 2018, 55, 5727–5740. [Google Scholar] [CrossRef]
- Zhu, N.; Liang, X.; Zhang, M.; Yin, X.; Yang, H.; Zhi, Y.; Ying, G.; Zou, J.; Chen, L.; Yao, X.; et al. Astaxanthin Protects Cognitive Function of Vascular Dementia. Behav. Brain Funct. 2020, 16, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Loganathan, C.; Sakayanathan, P.; Thayumanavan, P. Astaxanthin-s-Allyl Cysteine Diester against High Glucose-Induced Neuronal Toxicity in Vitro and Diabetes-Associated Cognitive Decline in Vivo: Effect on P53, Oxidative Stress and Mitochondrial Function. NeuroToxicology 2021, 86, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Katagiri, M.; Satoh, A.; Tsuji, S.; Shirasawa, T. Effects of Astaxanthin-Rich Haematococcus Pluvialis Extract on Cognitive Function: A Randomised, Double-Blind, Placebo-Controlled Study. J. Clin. Biochem. Nutr. 2012, 51, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ata Yaseen Abdulqader, Y.; Abdel Kawy, H.S.; Mohammed Alkreathy, H.; Abdullah Rajeh, N. The Potential Antiepileptic Activity of Astaxanthin in Epileptic Rats Treated with Valproic Acid. Saudi Pharm. J. SPJ 2021, 29, 418. [Google Scholar] [CrossRef] [PubMed]
- Ricketts, T.R. The Structures of Siphonein and Siphonaxanthin from Codium Fragile. Phytochemistry 1971, 10, 155–160. [Google Scholar] [CrossRef]
- Sugawara, T.; Ganesan, P.; Li, Z.; Manabe, Y.; Hirata, T. Siphonaxanthin, a Green Algal Carotenoid, as a Novel Functional Compound. Mar. Drugs 2014, 12, 3660. [Google Scholar] [CrossRef] [Green Version]
- Dambeck, M.; Sandmann, G. Antioxidative Activities of Algal Keto Carotenoids Acting as Antioxidative Protectants in the Chloroplast. Photochem. Photobiol. 2014, 90, 814–819. [Google Scholar] [CrossRef]
- Manabe, Y.; Takii, Y.; Sugawara, T. Siphonaxanthin, a Carotenoid from Green Algae, Suppresses Advanced Glycation End Product-Induced Inflammatory Responses. J. Nat. Med. 2019, 74, 127–134. [Google Scholar] [CrossRef]
- Ganesan, P.; Noda, K.; Manabe, Y.; Ohkubo, T.; Tanaka, Y.; Maoka, T.; Sugawara, T.; Hirata, T. Siphonaxanthin, a Marine Carotenoid from Green Algae, Effectively Induces Apoptosis in Human Leukemia (HL-60) Cells. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2011, 1810, 497–503. [Google Scholar] [CrossRef]
- Ganesan, P.; Matsubara, K.; Sugawara, T.; Hirata, T. Marine Algal Carotenoids Inhibit Angiogenesis by Down-Regulating FGF-2-Mediated Intracellular Signals in Vascular Endothelial Cells. Mol. Cell. Biochem. 2013, 380, 1–9. [Google Scholar] [CrossRef]
- Ganesan, P.; Matsubara, K.; Ohkubo, T.; Tanaka, Y.; Noda, K.; Sugawara, T.; Hirata, T. Anti-Angiogenic Effect of Siphonaxanthin from Green Alga, Codium Fragile. Phytomedicine 2010, 17, 1140–1144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.; Manabe, Y.; Sugawara, T. Siphonaxanthin, a Carotenoid from Green Algae Codium Cylindricum, Protects Ob/Ob Mice Fed on a High-Fat Diet against Lipotoxicity by Ameliorating Somatic Stresses and Restoring Anti-Oxidative Capacity. Nutr. Res. 2020, 77, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Li, Z.; Manabe, Y.; Kim, M.; Goto, T.; Kawada, T.; Sugawara, T. Siphonaxanthin, a Carotenoid From Green Algae, Inhibits Lipogenesis in Hepatocytes via the Suppression of Liver X Receptor α Activity. Lipids 2018, 53, 41–52. [Google Scholar] [CrossRef]
- Manabe, Y.; Tomonaga, N.; Maoka, T.; Sugawara, T. Multivariate Analysis Reveals That Unsubstituted β-Ring and C8-Keto Structures Are Important Factors for Anti-Inflammatory Activity of Carotenoids. Nutrients 2021, 13, 3699. [Google Scholar] [CrossRef]
- Eswarakumar, V.P.; Lax, I.; Schlessinger, J. Cellular Signaling by Fibroblast Growth Factor Receptors. Cytokine Growth Factor Rev. 2005, 16, 139–149. [Google Scholar] [CrossRef]
- Khare, A.; Moss, G.P.; Weedon, B.C.L. Mytiloxanthin and Isomytiloxanthin, Two Novel Acetylenic Carotenoids. Tetrahedron Lett. 1973, 14, 3921–3924. [Google Scholar] [CrossRef]
- Maoka, T.; Nishino, A.; Yasui, H.; Yamano, Y.; Wada, A. Anti-Oxidative Activity of Mytiloxanthin, a Metabolite of Fucoxanthin in Shellfish and Tunicates. Mar. Drugs 2016, 14, 93. [Google Scholar] [CrossRef] [Green Version]
- Shindo, K.; Kikuta, K.; Suzuki, A.; Katsuta, A.; Kasai, H.; Yasumoto-Hirose, M.; Matsuo, Y.; Misawa, N.; Takaichi, S. Rare Carotenoids, (3R)-Saproxanthin and (3R,2′S)-Myxol, Isolated from Novel Marine Bacteria (Flavobacteriaceae) and Their Antioxidative Activities. Appl. Microbiol. Biotechnol. 2007, 74, 1350–1357. [Google Scholar] [CrossRef]
- Subczynski, W.K.; Markowska, E.; Sielewiesiuk, J. Effect of Polar Carotenoids on the Oxygen Diffusion-Concentration Product in Lipid Bilayers. An EPR Spin Label Study. Biochim. Biophys. Acta (BBA)-Biomembr. 1991, 1068, 68–72. [Google Scholar] [CrossRef]
- Woodall, A.A.; Britton, G.; Jackson, M.J. Carotenoids and Protection of Phospholipids in Solution or in Liposomes against Oxidation by Peroxyl Radicals: Relationship between Carotenoid Structure and Protective Ability. Biochim. Biophys. Acta (BBA)-Gen. Subj. 1997, 1336, 575–586. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Carotenoid | Effect | Model | Bioactive Concentration | Target | Ref. |
|---|---|---|---|---|---|
| Fucoxanthin | neuroprotection | rat cortical neurons | 5, 10 and 20 μM | Nrf2 signaling | [88] |
| neuroprotection | PC12 cells | 0.5, 1, 2 and 5 μM | Nrf2 signaling | [89] | |
| anti-neuroinflammation | BV-2 microglial cells | 5, 10, and 20 μM | MAPKs and NF-κB signaling | [90] | |
| anti-neuroinflammation | bone marrow-derived macrophages, bone marrow-derived dendritic cells, astrocytes | 40 μM | NF-κB and NLRP3 inflammasome signaling | [81] | |
| Astaxanthin | anti-neuroinflammation | BV2 cells, PC12 cells, primary astrocytes | 5 or 10 μM | MAPKs and NF-κB signaling | [102] |
| neuronal viability | human neuronal cell line SH-SY5Y | 5, 10 and 15 μM | pro-apoptotic proteins | [105] | |
| Siphonaxanthin | anti-neuroinflammation | human monocytic cells | 1 μM for 24 h | NF-κB signaling | [117] |
| neuron survival synaptic plasticity | human endothelial cells | 0.1 and 0.5 μM for 6 h | FGF-2 signaling | [113] | |
| anti-proliferative | human leukemia cells | 20 μM | Bcl-2, CASP 3 | [112] | |
| Saproxanthin and Myxol | neuroprotection | embryonic rat retinal neuron hybrid cells | 3.1 and 8.1 μM, respectively | L-glutamate toxicity | [121] |
| Carotenoid | Effect | Model | Bioactive Concentration | Target | Ref. |
|---|---|---|---|---|---|
| Fucoxanthin | neuroprotection | rat stroke | 30, 60 and 90 mg/kg | Nrf2 signaling | [88] |
| neuroprotection | zebrafish | 6.25, 12.5, 25 and 50 μg/mL | Nrf2 signaling | [89] | |
| neuroprotection | traumatic brain injury mice | 50, 100 and 200 mg/kg | Nrf2/ARE signaling | [70] | |
| cognitive impairments attenuation | Alzheimer’s Disease mice | 50, 100 and 200 mg/kg | AChE, BDNF | [91] | |
| Astaxanthin | antioxidation anti-neuroinflammation, neuroregeneration | rats’ brain | 0.02% of daily diet, 3 times a week | antioxidant enzymes COX2, BDNF | [100] |
| anti-apoptotic, anti-inflammation, oxidative stress alleviation | rats | 100 mg/kg for 7 days | MBP, CASP 3, iNOS | [101] | |
| neuropathic pain alleviation | C57BL/6 mice | 5 or 10 mg/kg for 23 days | MAPKs and NF-κB signaling | [102] | |
| neuroprotection | rats | 25 mg/kg 5 times a week for 25 days | AChE | [103] | |
| oxidative stress alleviation | vascular dementia mice | 50, 100 and 200 mg/kg for 30 days | SOD, MDA, IL-4, IL-1β | [104] | |
| psychomotor speed improvement | people with mild memory impairment | 6 and 12 mg/day for 12 weeks | – | [106] | |
| Antiepileptic anti-inflammation | epileptic rats | 100 mg/kg | – | [107] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pietrasik, S.; Cichon, N.; Bijak, M.; Gorniak, L.; Saluk-Bijak, J. Carotenoids from Marine Sources as a New Approach in Neuroplasticity Enhancement. Int. J. Mol. Sci. 2022, 23, 1990. https://doi.org/10.3390/ijms23041990
Pietrasik S, Cichon N, Bijak M, Gorniak L, Saluk-Bijak J. Carotenoids from Marine Sources as a New Approach in Neuroplasticity Enhancement. International Journal of Molecular Sciences. 2022; 23(4):1990. https://doi.org/10.3390/ijms23041990
Chicago/Turabian StylePietrasik, Sylwia, Natalia Cichon, Michal Bijak, Leslaw Gorniak, and Joanna Saluk-Bijak. 2022. "Carotenoids from Marine Sources as a New Approach in Neuroplasticity Enhancement" International Journal of Molecular Sciences 23, no. 4: 1990. https://doi.org/10.3390/ijms23041990
APA StylePietrasik, S., Cichon, N., Bijak, M., Gorniak, L., & Saluk-Bijak, J. (2022). Carotenoids from Marine Sources as a New Approach in Neuroplasticity Enhancement. International Journal of Molecular Sciences, 23(4), 1990. https://doi.org/10.3390/ijms23041990