
The Protein Phosphatase GhAP2C1 Interacts Together with GhMPK4 to Synergistically Regulate the Immune Response to Fusarium oxysporum in Cotton

Abstract
:1. Introduction
2. Results
2.1. Enhanced Sensitivity of GhMPK4-Silenced Cotton to Fusarium oxysporum Increased
2.2. The Sensitivity of Plants to Fusarium oxysporum Increased after the Overexpression of GhMPK4
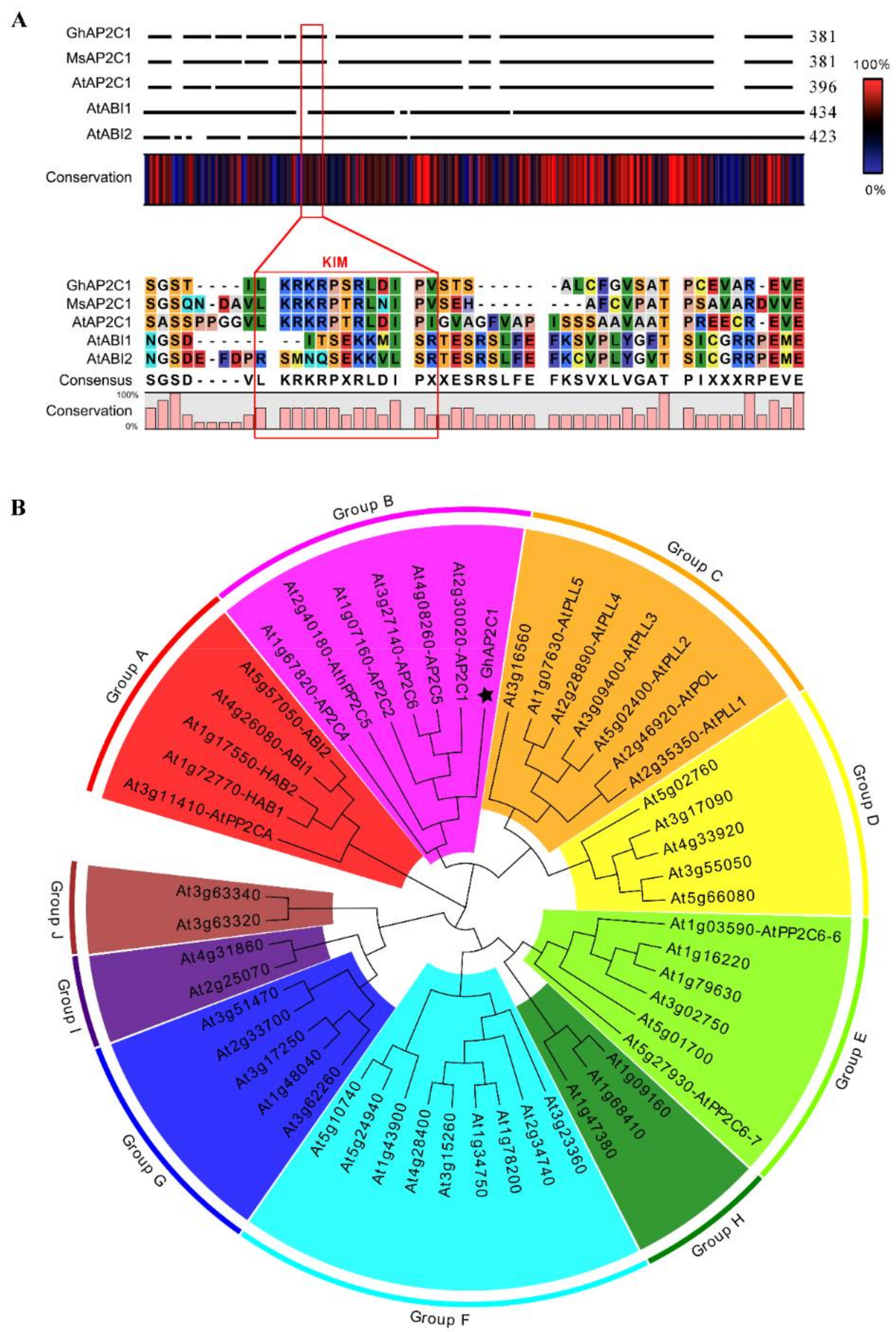
2.3. Characterization of GhAP2C1
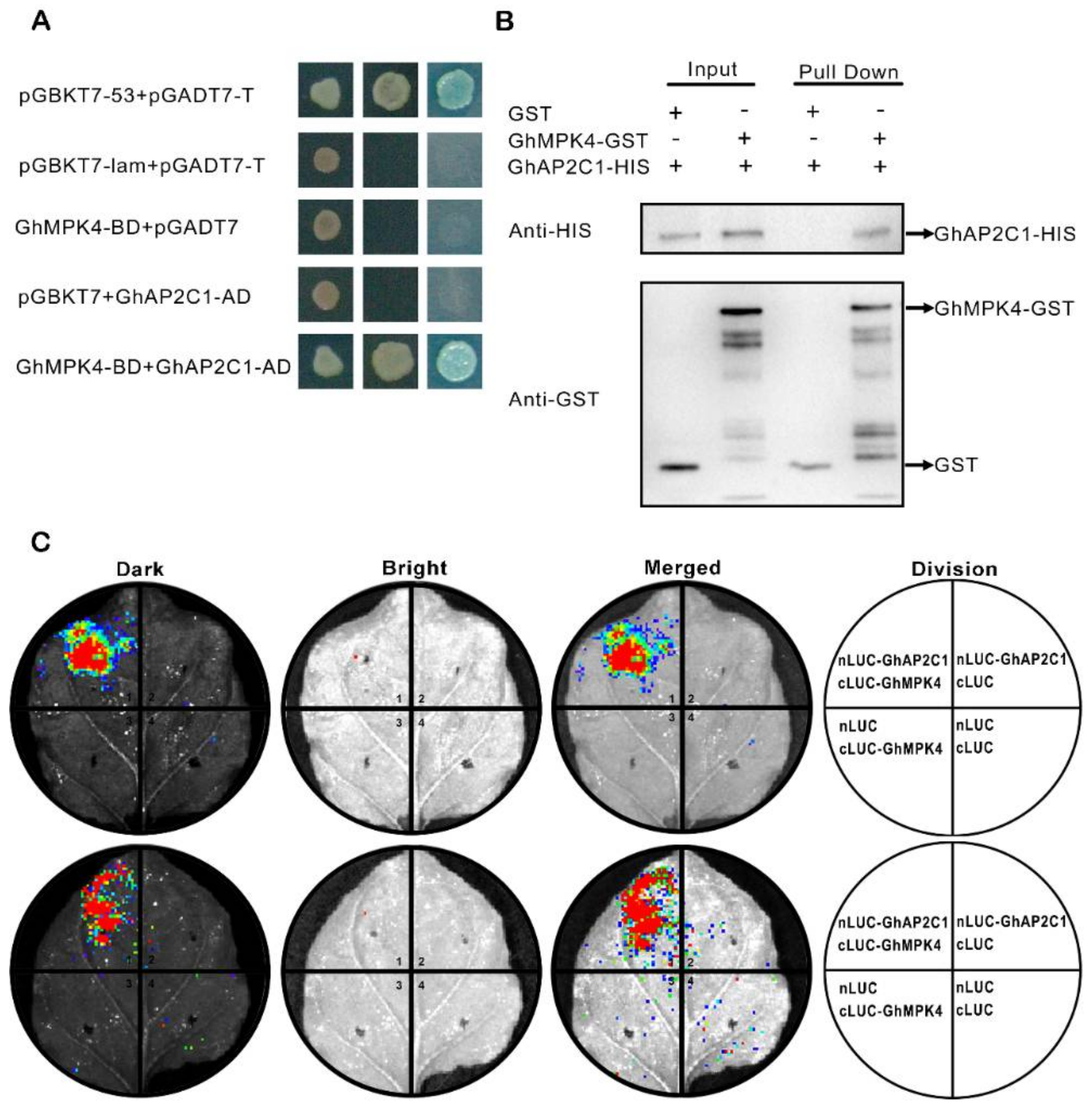
2.4. Verification of GhAP2C1 and GhMPK4 Interactions
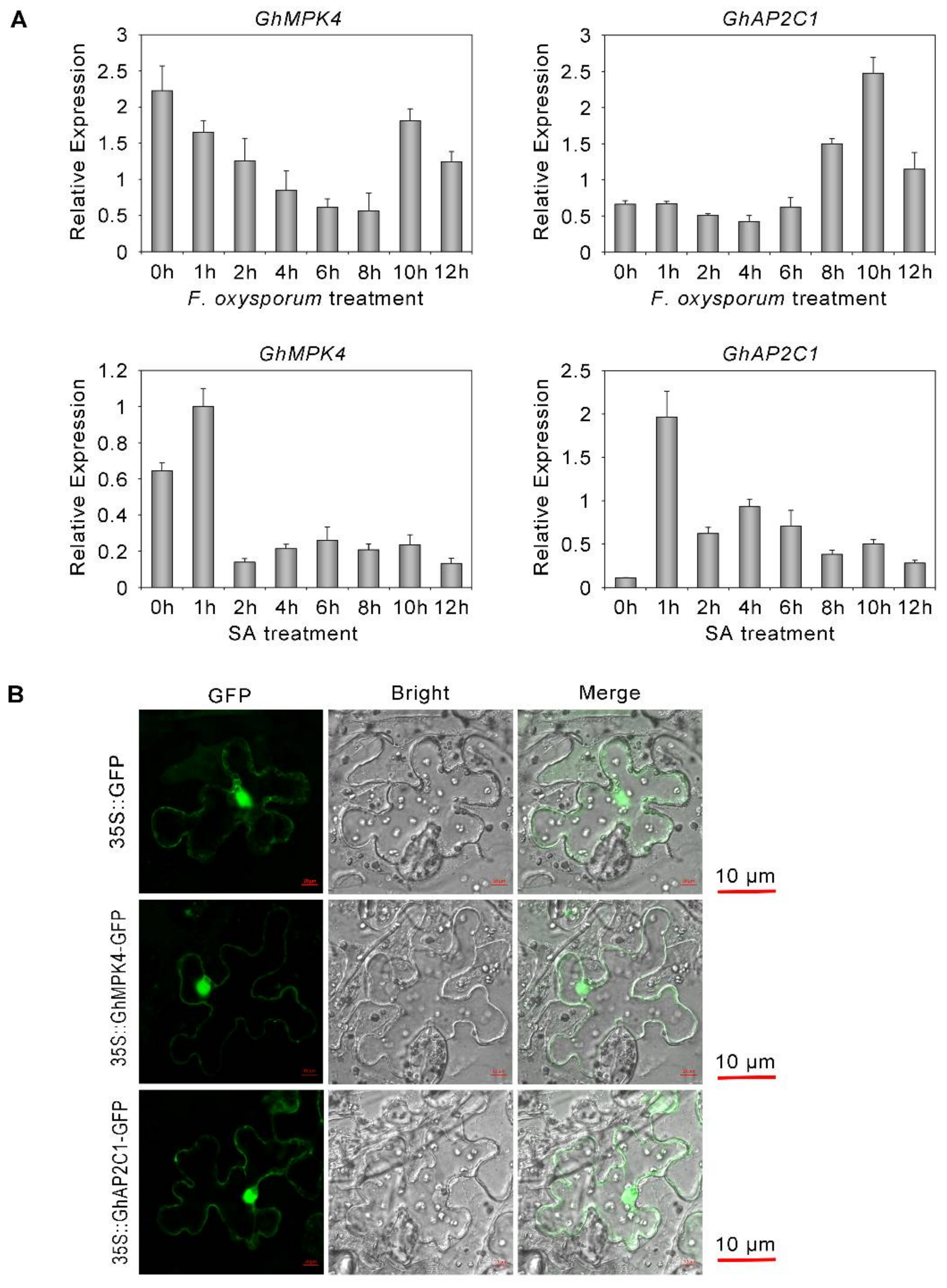
2.5. Expression Characteristics of GhAP2C1 and GhMPK4
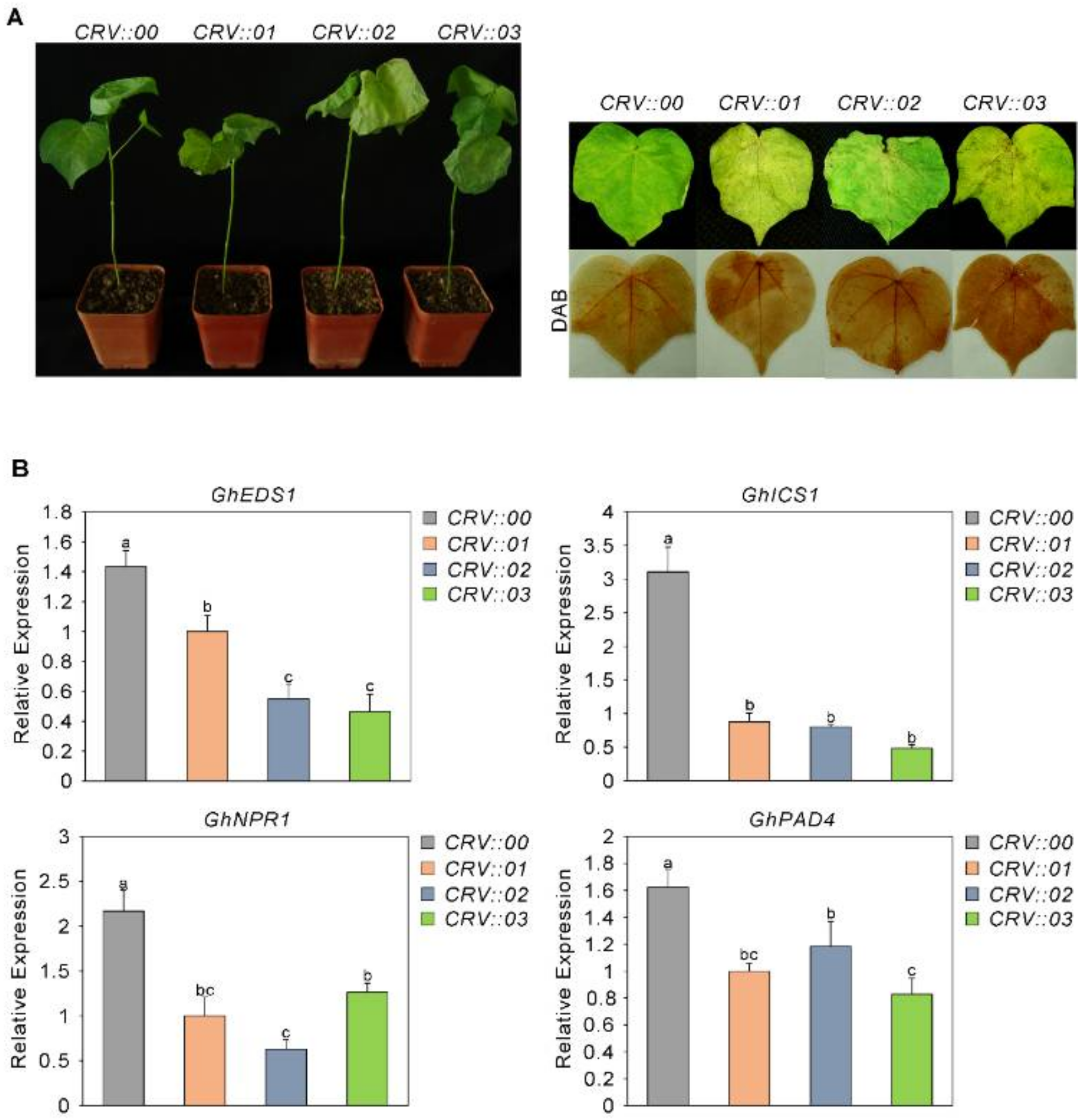
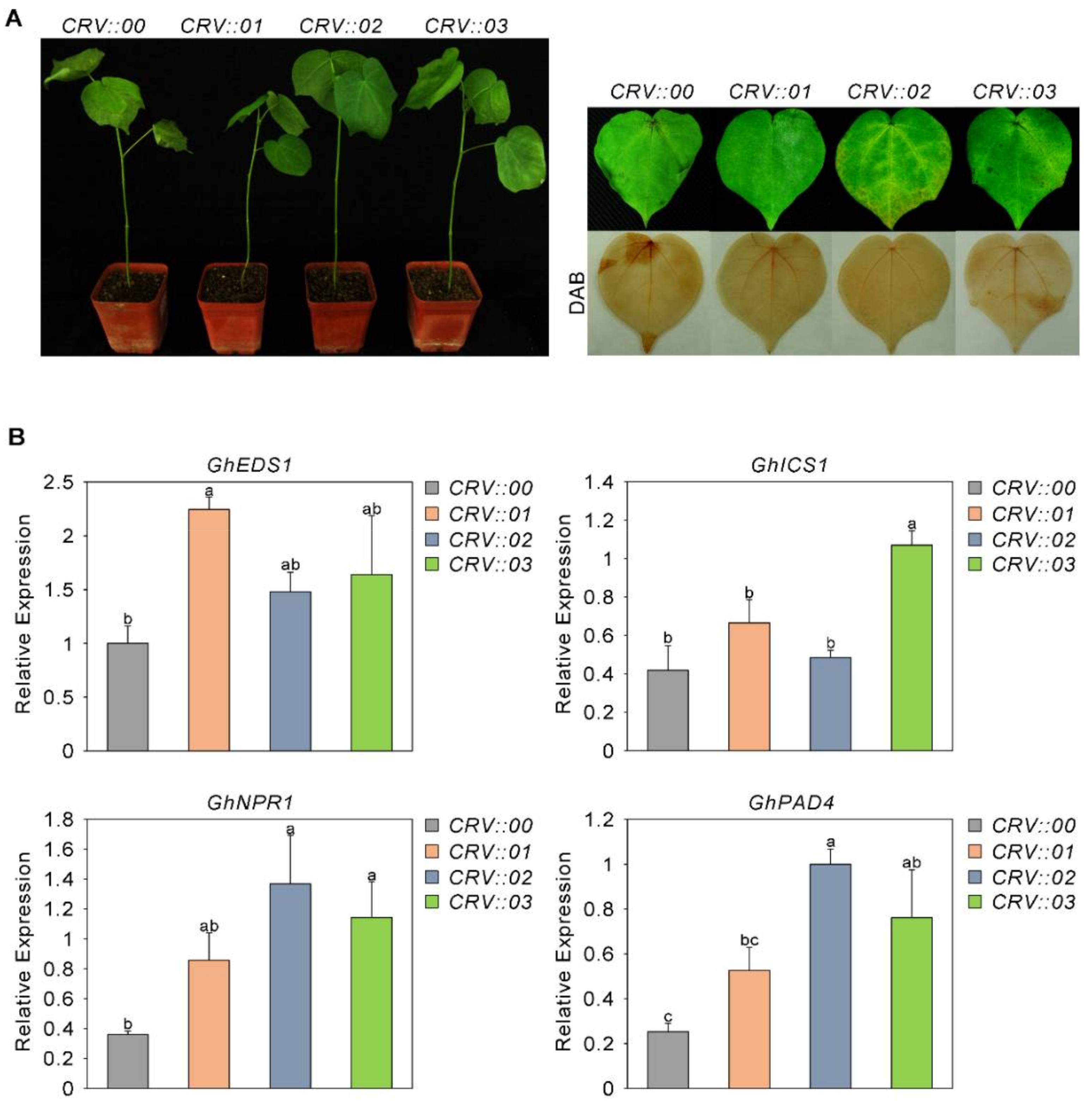
2.6. The Sensitivity of GhAP2C1-Silenced Cotton to Fusarium oxysporum Increased
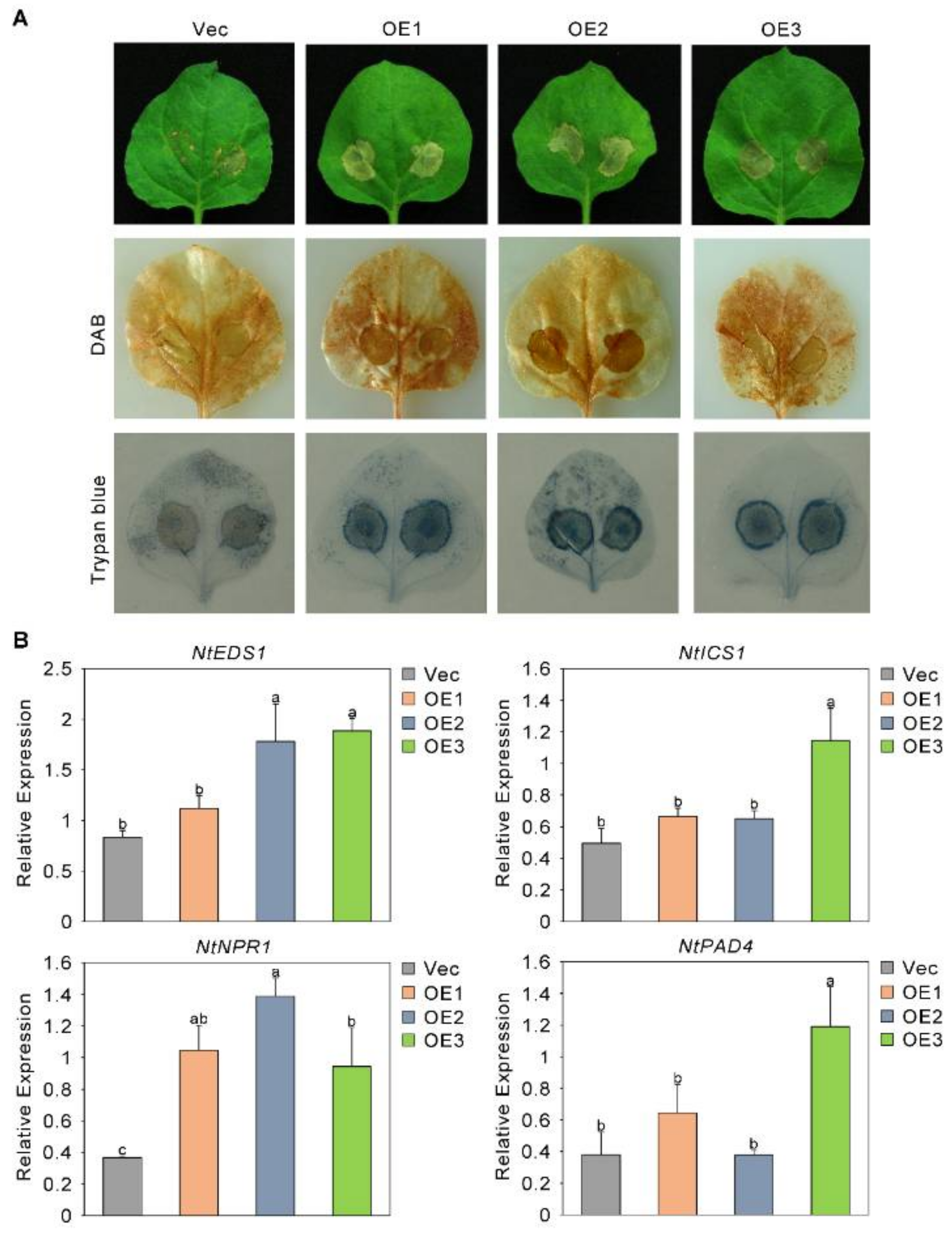
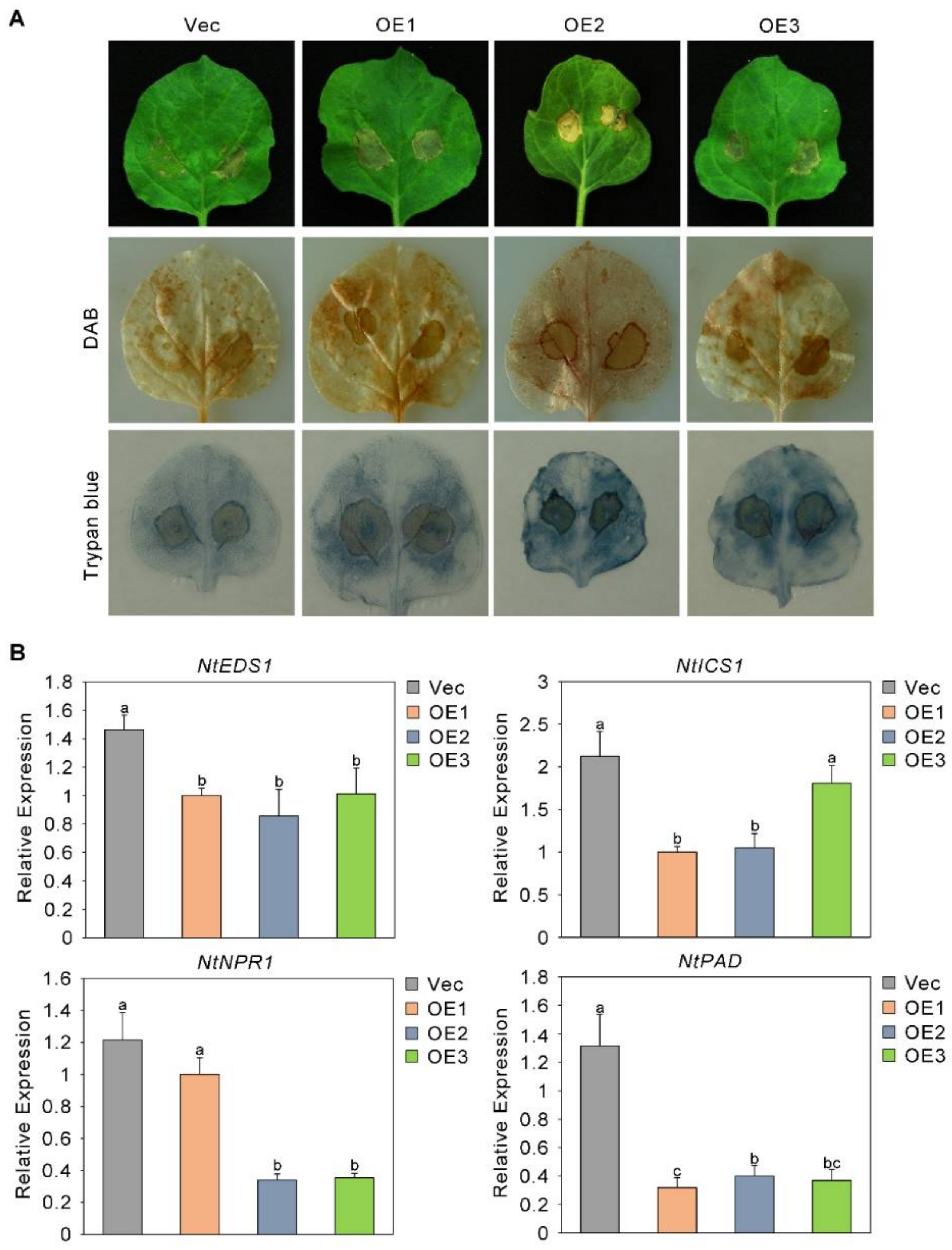
2.7. The Sensitivity of Plants to Fusarium oxysporum Increased after the Overexpression of GhAP2C1
3. Discussion
4. Materials and Methods
4.1. Plant Treatments
4.2. RNA Extraction and qRT-PCR
4.3. VIGS
4.4. Subcellular Localization of GhMPK4 and GhAP2C1
4.5. Yeast Two-Hybrid, LCI and Pull-Down Assays
4.6. Primers
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimura, M.T.; Dangl, J.L. Arabidopsis thaliana and the plant immune system. Plant J. 2010, 61, 1053–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, C.S.; Martin, G.B. Effector-triggered immunity mediated by the Pto kinase. Trends Plant Sci. 2011, 16, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.A.; Jones, J.D.; Dangl, J.L. Reactive oxygen species signaling in response to pathogens. Plant Physiol. 2006, 141, 373–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durrant, W.E.; Dong, X. Systemic acquired resistance. Annu. Rev. Phytopathol. 2004, 42, 185–209. [Google Scholar] [CrossRef] [PubMed]
- Naveed, Z.A.; Wei, X.; Chen, J.; Mubeen, H.; Ali, G.S. The PTI to ETI Continuum in Phytophthora-Plant Interactions. Front. Plant Sci. 2020, 11, 593905. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, K.; Katagiri, F. Comparing signaling mechanisms engaged in pattern-triggered and effector-triggered immunity. Curr. Opin. Plant Biol. 2010, 13, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Coaker, G.; Zhou, J.M.; Dong, X. Plant Immune Mechanisms: From Reductionistic to Holistic Points of View. Mol. Plant 2020, 13, 1358–1378. [Google Scholar] [CrossRef] [PubMed]
- Bigeard, J.; Colcombet, J.; Hirt, H. Signaling mechanisms in pattern-triggered immunity (PTI). Mol. Plant 2015, 8, 521–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chisholm, S.T.; Coaker, G.; Day, B.; Staskawicz, B.J. Host-microbe interactions: Shaping the evolution of the plant immune response. Cell 2006, 124, 803–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes Fischer, N.; Naseer, N.; Shin, S.; Brodsky, I.E. Effector-triggered immunity and pathogen sensing in metazoans. Nat. Microbiol. 2020, 5, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Zavaliev, R.; Mohan, R.; Chen, T.; Dong, X. Formation of NPR1 Condensates Promotes Cell Survival during the Plant Immune Response. Cell 2020, 182, 1093–1108.e1018. [Google Scholar] [CrossRef] [PubMed]
- Ngou, B.P.M.; Ahn, H.K.; Ding, P.; Jones, J.D.G. Mutual potentiation of plant immunity by cell-surface and intracellular receptors. Nature 2021, 592, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Jiang, Z.; Bi, G.; Nomura, K.; Liu, M.; Wang, Y.; Cai, B.; Zhou, J.M.; He, S.Y.; Xin, X.F. Pattern-recognition receptors are required for NLR-mediated plant immunity. Nature 2021, 592, 105–109. [Google Scholar] [CrossRef]
- Zhou, J.M.; Zhang, Y. Plant Immunity: Danger Perception and Signaling. Cell 2020, 181, 978–989. [Google Scholar] [CrossRef]
- Wu, J.; Hettenhausen, C.; Meldau, S.; Baldwin, I.T. Herbivory rapidly activates MAPK signaling in attacked and unattacked leaf regions but not between leaves of Nicotiana attenuata. Plant Cell 2007, 19, 1096–1122. [Google Scholar] [CrossRef] [Green Version]
- Bi, G.; Zhou, Z.; Wang, W.; Li, L.; Rao, S.; Wu, Y.; Zhang, X.; Menke, F.L.H.; Chen, S.; Zhou, J.M. Receptor-Like Cytoplasmic Kinases Directly Link Diverse Pattern Recognition Receptors to the Activation of Mitogen-Activated Protein Kinase Cascades in Arabidopsis thaliana. Plant Cell 2018, 30, 1543–1561. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, K.; Yamada, K.; Kawasaki, T. Receptor-like cytoplasmic kinases are pivotal components in pattern recognition receptor-mediated signaling in plant immunity. Plant Signal. Behav. 2013, 8, e25662. [Google Scholar] [CrossRef] [Green Version]
- Adachi, H.; Nakano, T.; Miyagawa, N.; Ishihama, N.; Yoshioka, M.; Katou, Y.; Yaeno, T.; Shirasu, K.; Yoshioka, H. WRKY Transcription Factors Phosphorylated by MAPK Regulate a Plant Immune NADPH Oxidase in Nicotiana benthamiana. Plant Cell 2015, 27, 2645–2663. [Google Scholar] [CrossRef] [Green Version]
- Lang, J.; Colcombet, J. Sustained Incompatibility between MAPK Signaling and Pathogen Effectors. Int. J. Mol. Sci. 2020, 21, 7954. [Google Scholar] [CrossRef]
- Meng, X.; Zhang, S. MAPK cascades in plant disease resistance signaling. Annu. Rev. Phytopathol. 2013, 51, 245–266. [Google Scholar] [CrossRef]
- Singh, R.; Jwa, N.S. The rice MAPKK-MAPK interactome: The biological significance of MAPK components in hormone signal transduction. Plant Cell Rep. 2013, 32, 923–931. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Su, J.; Zhang, Y.; Xu, J.; Zhang, S. Conveying endogenous and exogenous signals: MAPK cascades in plant growth and defense. Curr. Opin. Plant Biol. 2018, 45, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Danquah, A.; de Zelicourt, A.; Colcombet, J.; Hirt, H. The role of ABA and MAPK signaling pathways in plant abiotic stress responses. Biotechnol. Adv. 2014, 32, 40–52. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, S. Mitogen-activated protein kinase cascades in signaling plant growth and development. Trends Plant Sci. 2015, 20, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Pinaud, L.; Sansonetti, P.J.; Phalipon, A. Host Cell Targeting by Enteropathogenic Bacteria T3SS Effectors. Trends Microbiol. 2018, 26, 266–283. [Google Scholar] [CrossRef] [PubMed]
- Hettenhausen, C.; Schuman, M.C.; Wu, J. MAPK signaling: A key element in plant defense response to insects. Insect Sci. 2015, 22, 157–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitzschke, A.; Djamei, A.; Bitton, F.; Hirt, H. A major role of the MEKK1-MKK1/2-MPK4 pathway in ROS signalling. Mol. Plant 2009, 2, 120–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Liu, J.; Yin, C.; de Souza Vespoli, L.; Ge, D.; Huang, Y.; Feng, B.; Xu, G.; Manhães, A.; Dou, S.; et al. RNA Interference-Based Screen Reveals Concerted Functions of MEKK2 and CRCK3 in Plant Cell Death Regulation. Plant Physiol. 2020, 183, 331–344. [Google Scholar] [CrossRef] [Green Version]
- Gao, M.; Liu, J.; Bi, D.; Zhang, Z.; Cheng, F.; Chen, S.; Zhang, Y. MEKK1, MKK1/MKK2 and MPK4 function together in a mitogen-activated protein kinase cascade to regulate innate immunity in plants. Cell Res. 2008, 18, 1190–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, S.; Tischer, S.V.; Wunschel, C.; Christmann, A.; Grill, E. Abscisic acid sensor RCAR7/PYL13, specific regulator of protein phosphatase coreceptors. Proc. Natl. Acad. Sci. USA 2014, 111, 5741–5746. [Google Scholar] [CrossRef] [Green Version]
- Farooq, A.; Zhou, M.M. Structure and regulation of MAPK phosphatases. Cell Signal. 2004, 16, 769–779. [Google Scholar] [CrossRef] [PubMed]
- Brock, A.K.; Willmann, R.; Kolb, D.; Grefen, L.; Lajunen, H.M.; Bethke, G.; Lee, J.; Nürnberger, T.; Gust, A.A. The Arabidopsis thaliana mitogen-activated protein kinase phosphatase PP2C5 affects seed germination, stomatal aperture, and abscisic acid-inducible gene expression. Plant Physiol. 2010, 153, 1098–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schweighofer, A.; Kazanaviciute, V.; Scheikl, E.; Teige, M.; Doczi, R.; Hirt, H.; Schwanninger, M.; Kant, M.; Schuurink, R.; Mauch, F.; et al. The PP2C-type phosphatase AP2C1, which negatively regulates MPK4 and MPK6, modulates innate immunity, jasmonic acid, and ethylene levels in Arabidopsis. Plant Cell 2007, 19, 2213–2224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galletti, R.; Ferrari, S.; De Lorenzo, G. Arabidopsis thaliana MPK3 and MPK6 play different roles in basal and oligogalacturonide- or flagellin-induced resistance against Botrytis cinerea. Plant Physiol. 2011, 157, 804–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shubchynskyy, V.; Boniecka, J.; Schweighofer, A.; Simulis, J.; Kvederaviciute, K.; Stumpe, M.; Mauch, F.; Balazadeh, S.; Mueller-Roeber, B.; Boutrot, F.; et al. Protein phosphatase AP2C1 negatively regulates basal resistance and defense responses to Pseudomonas syringae. J. Exp. Bot. 2017, 68, 1169–1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segonzac, C.; Feike, D.; Gimenez-Ibanez, S.; Hann, D.R.; Zipfel, C.; Rathjen, J.P. Hierarchy and roles of pathogen-associated molecular pattern-induced responses in Nicotiana benthamiana. Plant Physiol. 2011, 156, 687–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, K.; Pan, S. Maize protein phosphatase gene family: Identification and molecular characterization. BMC Genomics 2014, 15, 773. [Google Scholar] [CrossRef] [Green Version]
- Schweighofer, A.; Hirt, H.; Meskiene, I. Plant PP2C phosphatases: Emerging functions in stress signaling. Trends Plant Sci. 2004, 9, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Umbrasaite, J.; Schweighofer, A.; Kazanaviciute, V.; Magyar, Z.; Ayatollahi, Z.; Unterwurzacher, V.; Choopayak, C.; Boniecka, J.; Murray, J.A.; Bogre, L.; et al. MAPK phosphatase AP2C3 induces ectopic proliferation of epidermal cells leading to stomata development in Arabidopsis. PLoS ONE 2010, 5, e15357. [Google Scholar] [CrossRef] [Green Version]
- Meskiene, I.; Baudouin, E.; Schweighofer, A.; Liwosz, A.; Jonak, C.; Rodriguez, P.L.; Jelinek, H.; Hirt, H. Stress-induced protein phosphatase 2C is a negative regulator of a mitogen-activated protein kinase. J. Biol. Chem. 2003, 278, 18945–18952. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Chen, H.; Hou, Z.; Li, Y.; Yang, C.; Wang, D.; Song, C.P. Screening of abiotic stress-responsive cotton genes using a cotton full-length cDNA overexpressing Arabidopsis thaliana library. J. Integr. Plant Biol. 2020, 62, 998–1016. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Zhu, L.; Wassan, G.M.; Wang, Y.; Miao, Y.; Shaban, M.; Hu, H.; Sun, H.; Zhang, X. GhJAZ2 attenuates cotton resistance to biotic stresses via the inhibition of the transcriptional activity of GhbHLH171. Mol. Plant Pathol. 2018, 19, 896–908. [Google Scholar] [CrossRef]
- Gaspar, Y.M.; McKenna, J.A.; McGinness, B.S.; Hinch, J.; Poon, S.; Connelly, A.A.; Anderson, M.A.; Heath, R.L. Field resistance to Fusarium oxysporum and Verticillium dahliae in transgenic cotton expressing the plant defensin NaD1. J. Exp. Bot. 2014, 65, 1541–1550. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zhao, Y.L.; Zhao, J.H.; Wang, S.; Jin, Y.; Chen, Z.Q.; Fang, Y.Y.; Hua, C.L.; Ding, S.W.; Guo, H.S. Cotton plants export microRNAs to inhibit virulence gene expression in a fungal pathogen. Nat. Plants 2016, 2, 16153. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Guo, H.; He, X.; Zhang, S.; Wang, J.; Wang, L.; Guo, D.; Guo, X. Scaffold protein GhMORG1 enhances the resistance of cotton to Fusarium oxysporum by facilitating the MKK6-MPK4 cascade. Plant Biotechnol. J. 2020, 18, 1421–1433. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; He, X.; Wang, X.; Zhang, S.; Guo, X. ghr-miR5272a-mediated regulation of GhMKK6 gene transcription contributes to the immune response in cotton. J. Exp. Bot. 2017, 68, 5895–5906. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Xu, R.; Li, Y. Molecular Networks of Seed Size Control in Plants. Annu. Rev. Plant Biol. 2019, 70, 435–463. [Google Scholar] [CrossRef] [PubMed]
- Fiil, B.K.; Petersen, K.; Petersen, M.; Mundy, J. Gene regulation by MAP kinase cascades. Curr. Opin. Plant Biol. 2009, 12, 615–621. [Google Scholar] [CrossRef] [PubMed]
- Asai, T.; Tena, G.; Plotnikova, J.; Willmann, M.R.; Chiu, W.L.; Gomez-Gomez, L.; Boller, T.; Ausubel, F.M.; Sheen, J. MAP kinase signalling cascade in Arabidopsis thaliana innate immunity. Nature 2002, 415, 977–983. [Google Scholar] [CrossRef]
- Hua, Z.M.; Yang, X.; Fromm, M.E. Activation of the NaCl- and drought-induced RD29A and RD29B promoters by constitutively active Arabidopsis thaliana MAPKK or MAPK proteins. Plant Cell Environ. 2006, 29, 1761–1770. [Google Scholar] [CrossRef] [PubMed]
- Desikan, R.; Hancock, J.T.; Ichimura, K.; Shinozaki, K.; Neill, S.J. Harpin induces activation of the Arabidopsis thaliana mitogen-activated protein kinases AtMPK4 and AtMPK6. Plant Physiol. 2001, 126, 1579–1587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Li, Y.; Lu, W.; Meng, F.; Wu, C.A.; Guo, X. Cotton GhMKK5 affects disease resistance, induces HR-like cell death, and reduces the tolerance to salt and drought stress in transgenic Nicotiana benthamiana. J. Exp. Bot. 2012, 63, 3935–3951. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.J.; Yang, W.X. Kinesins in MAPK cascade: How kinesin motors are involved in the MAPK pathway? Gene 2019, 684, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Boudsocq, M.; Danquah, A.; de Zélicourt, A.; Hirt, H.; Colcombet, J. Plant MAPK cascades: Just rapid signaling modules? Plant Signal. Behav. 2015, 10, e1062197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, Q.; Qu, N.; Gao, M.; Zhang, Z.; Ding, X.; Yang, F.; Li, Y.; Dong, O.X.; Chen, S.; Li, X.; et al. The MEKK1-MKK1/MKK2-MPK4 kinase cascade negatively regulates immunity mediated by a mitogen-activated protein kinase kinase kinase in Arabidopsis thaliana. Plant Cell 2012, 24, 2225–2236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Mi, X.; Chen, C.; Wang, H.; Guo, W. Identification on mitogen-activated protein kinase signaling cascades by integrating protein interaction with transcriptional profiling analysis in cotton. Sci. Rep. 2018, 8, 8178. [Google Scholar] [CrossRef] [Green Version]
- Xiang, L.; Stephen Sze, C.W.; Ng, T.B.; Tong, Y.; Shaw, P.C.; Sydney Tang, C.W.; Kalin Zhang, Y.B. Polysaccharides of Dendrobium officinale inhibit TNF-α-induced apoptosis in A-253 cell line. Inflamm. Res. 2013, 62, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Zhao, F.; Chen, L.; Pan, Y.; Sun, L.; Bao, N.; Zhang, T.; Cui, C.X.; Qiu, Z.; Zhang, Y.; et al. Jasmonate-mediated wound signalling promotes plant regeneration. Nat. Plants 2019, 5, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Ji, Z.L.; Yu, M.H.; Ding, Y.Y.; Li, J.; Zhu, F.; He, J.X.; Yang, L.N. Coiled-Coil N21 of Hpa1 in Xanthomonas oryzae pv. oryzae Promotes Plant Growth, Disease Resistance and Drought Tolerance in Non-Hosts via Eliciting HR and Regulation of Multiple Defense Response Genes. Int. J. Mol. Sci. 2020, 22, 203. [Google Scholar] [CrossRef]
- Sun, X.; Gilroy, E.M.; Chini, A.; Nurmberg, P.L.; Hein, I.; Lacomme, C.; Birch, P.R.; Hussain, A.; Yun, B.W.; Loake, G.J. ADS1 encodes a MATE-transporter that negatively regulates plant disease resistance. New Phytol. 2011, 192, 471–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suarez-Rodriguez, M.C.; Adams-Phillips, L.; Liu, Y.; Wang, H.; Su, S.H.; Jester, P.J.; Zhang, S.; Bent, A.F.; Krysan, P.J. MEKK1 is required for flg22-induced MPK4 activation in Arabidopsis thaliana plants. Plant Physiol. 2007, 143, 661–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, M.; Bai, M.Y.; Kim, J.G.; Wang, T.; Oh, E.; Chen, L.; Park, C.H.; Son, S.H.; Kim, S.K.; Mudgett, M.B.; et al. The bHLH transcription factor HBI1 mediates the trade-off between growth and pathogen-associated molecular pattern-triggered immunity in Arabidopsis thaliana. Plant Cell 2014, 26, 828–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sidonskaya, E.; Schweighofer, A.; Shubchynskyy, V.; Kammerhofer, N.; Hofmann, J.; Wieczorek, K.; Meskiene, I. Plant resistance against the parasitic nematode Heterodera schachtii is mediated by MPK3 and MPK6 kinases, which are controlled by the MAPK phosphatase AP2C1 in Arabidopsis thaliana. J. Exp. Bot. 2016, 67, 107–118. [Google Scholar] [CrossRef] [Green Version]
- Gu, Z.; Huang, C.; Li, F.; Zhou, X. A versatile system for functional analysis of genes and microRNAs in cotton. Plant Biotechnol. J. 2014, 12, 638–649. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Bi, G.; Zhou, J.M. Luciferase Complementation Assay for Protein-Protein Interactions in Plants. Curr. Protoc. Plant Biol. 2018, 3, 42–50. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Primer Sequence (5′→3′) | Annotation |
|---|---|---|
| K4-QC-5 | ATGAAAAAGGAAATGGGGAGTAC | The full-length cDNA primers |
| K4-QC-3 | CTTAATGGACTGGATCTGGATTG | |
| AP2C1-QC-5 | ATGTCGTGTTCGGTCGCTGTATG | |
| AP2C1-QC-3 | AATATAGCGGCCCAGTTGAATC | |
| GhEDS1-5 | GCAGCAACAGCTCCTCTACCTCAA | Primers used in qRT-PCR |
| GhEDS1-3 | GGCAGACCAAGACGCTACAGATACA | |
| GhICS1-5 | ATGGATGAATGGGTGCGAAGG | |
| GhICS1-3 | AAGAATGCCAGAGGTAAGAGGAGGA | |
| GhPAD4-5 | GGATGGAAGAATGGAAAGAAATGAA | |
| GhPAD4-3 | GAACTAGGAAAGCAGACTAAGGAACCA | |
| GhNPR1-5 | GCGAATCGTTGCTTTCTTCTTCA | |
| GhNPR1-3 | CACGTGGTGCTGTTGTTGTTACTG | |
| NbEDS1-5 | TCTGGATAGGCTGAAAGCAC | |
| NbEDS1-3 | CCATACAAGCAAAGCAGTTCC | |
| NbICS1-5 | CAATTCCGCCATCTCTCACT | |
| NbICS1-3 | TGAGCATGAAGCCACTCAAG | |
| NbPAD4-5 | GGACTCACACTCCAGCGTTT | |
| NbPAD4-3 | GGCAACTCATCCTCTTCCTG | |
| NbNPR1-5 | GCAGCAGACGATGTAATGATGG | |
| NbNPR1-3 | TCCACAAGCCTAGTGAGCCTC | |
| AP2C1-5 | GGAAGCGTTGAACTTGTCTTTG | |
| AP2C1-3 | GCCGAAACCCCAAAACACAA | |
| K4-CRV-5 | ACTAGTCATCAAGGACATCATTCGACC | Primers used in VIGS |
| K4-CRV-3 | TTAATTAAGATGAACGTAATCTTTGCCTGG | |
| AP2C1-CRV-5 | ATCAAGGCACGTCTGGGTCTCTTTTGTCAAC | |
| AP2C1-CRV-3 | TTAATTAACTCCACTCCTGTCTCCTCTTC | |
| K4-BD-5 | CATATGAAAAAGGAAATGGGGAGTAC | Primers used in Yeast two-hybrid |
| K4-BD-3 | GTCGACTTAATGGACTGGATCTGGATTG | |
| AP2C1-AD-5 | CATATGATGTCGTGTTCGGTCGCTG | |
| AP2C1-AD-3 | GAGCTCAATATAGCGGCCCAGTTGAATC | |
| AP2C1-GFP-5 | TCTAGAATGTCGTGTTCGGTCGCTG | Primers used in Subcellular localization |
| AP2C1-GFP-3 | GGTACCAATATAGCGGCCCAGTTGAATC | |
| K4-GFP-5 | TCTAGAATGAAAAAGGAAATGGGG | |
| K4-GFP-3 | GGATCCATGGACTGGATCTGGAT | |
| K4-LUC-5 | GGATCCATGAAAAAGGAAATGGGG | Primers used in LCI |
| K4-LUC-3 | GTCGACATGGACTGGATCTGGAT | |
| AP2C-LUC-5 | GGATCCATGTCGTGTTCGGTCGCTG | |
| AP2C-LUC-5 | GTCGACAATATAGCGGCCCAGTTGAATC | |
| AP2C-HIS-5 | GGATCCATGTCGTGTTCGGTCGCTGTATG | Primers used in Pull-down |
| AP2C-HIS-3 | GAGCTCCTTGTCGTCATCGTCTTTGTAGTC | |
| K4-GST-5 | GTCGACATGAAAAAGGAAATGGGG | |
| K4-GST-3 | TCTAGACATGGACTGGATCTGGAT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, D.; Hao, C.; Hou, J.; Zhao, G.; Shan, W.; Guo, H.; Wang, C.; Guo, X. The Protein Phosphatase GhAP2C1 Interacts Together with GhMPK4 to Synergistically Regulate the Immune Response to Fusarium oxysporum in Cotton. Int. J. Mol. Sci. 2022, 23, 2014. https://doi.org/10.3390/ijms23042014
Guo D, Hao C, Hou J, Zhao G, Shan W, Guo H, Wang C, Guo X. The Protein Phosphatase GhAP2C1 Interacts Together with GhMPK4 to Synergistically Regulate the Immune Response to Fusarium oxysporum in Cotton. International Journal of Molecular Sciences. 2022; 23(4):2014. https://doi.org/10.3390/ijms23042014
Chicago/Turabian StyleGuo, Dezheng, Cuihong Hao, Junbin Hou, Guangdong Zhao, Wenlu Shan, Huijuan Guo, Chen Wang, and Xingqi Guo. 2022. "The Protein Phosphatase GhAP2C1 Interacts Together with GhMPK4 to Synergistically Regulate the Immune Response to Fusarium oxysporum in Cotton" International Journal of Molecular Sciences 23, no. 4: 2014. https://doi.org/10.3390/ijms23042014
APA StyleGuo, D., Hao, C., Hou, J., Zhao, G., Shan, W., Guo, H., Wang, C., & Guo, X. (2022). The Protein Phosphatase GhAP2C1 Interacts Together with GhMPK4 to Synergistically Regulate the Immune Response to Fusarium oxysporum in Cotton. International Journal of Molecular Sciences, 23(4), 2014. https://doi.org/10.3390/ijms23042014