
New Function of Cholesterol Oxidation Products Involved in Osteoporosis Pathogenesis

 and
and 
Abstract
:1. Introduction
2. Molecular Mechanisms of OP
3. Anabolism of COPs
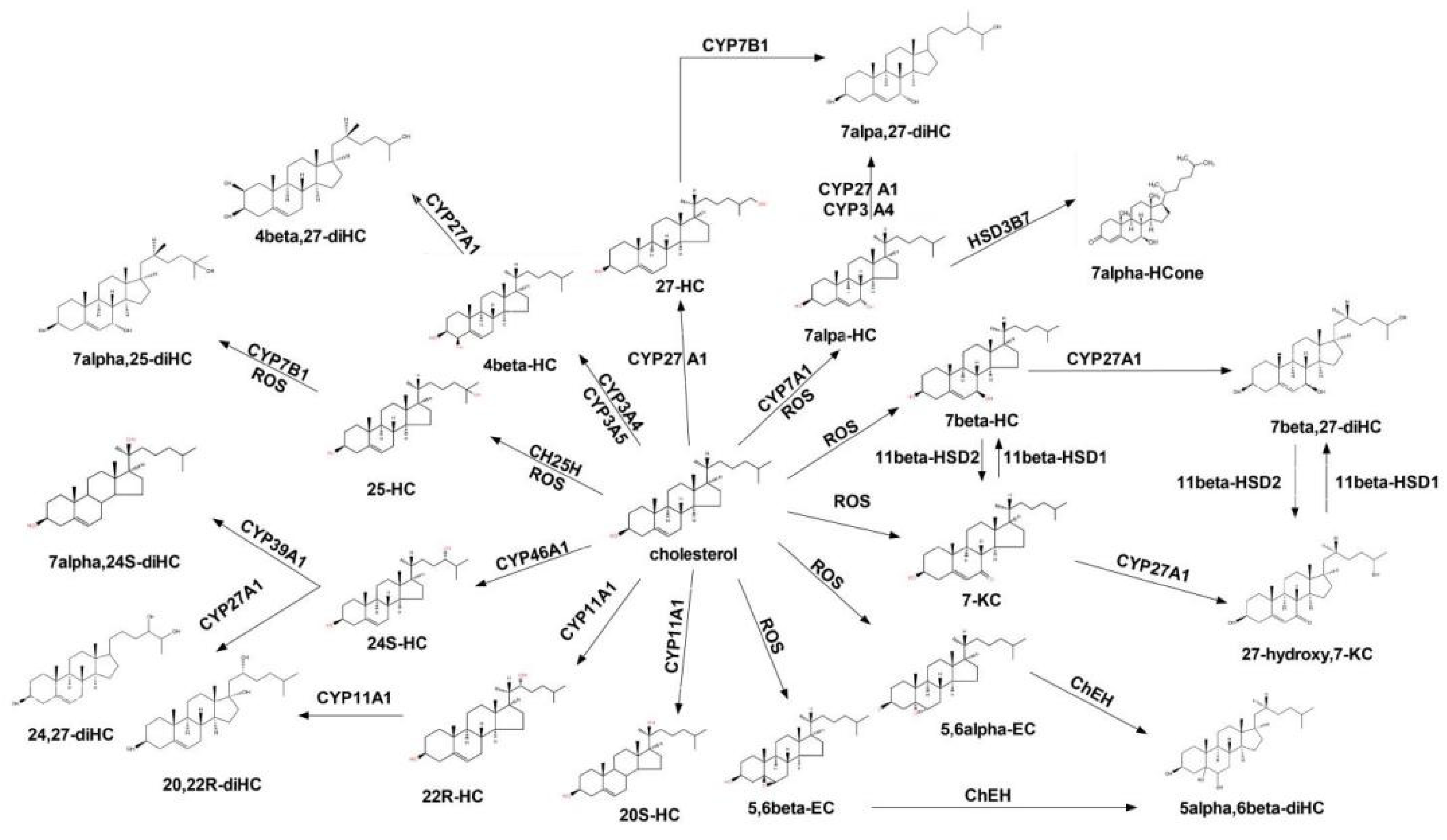
3.1. The Formation of COPs
3.1.1. Non-Enzymic Oxidation
3.1.2. Enzymic Oxidation
3.2. Dietary Sources of COPs
3.3. Metabolism of COPs
4. Biological Functions of COPs Involved in OP
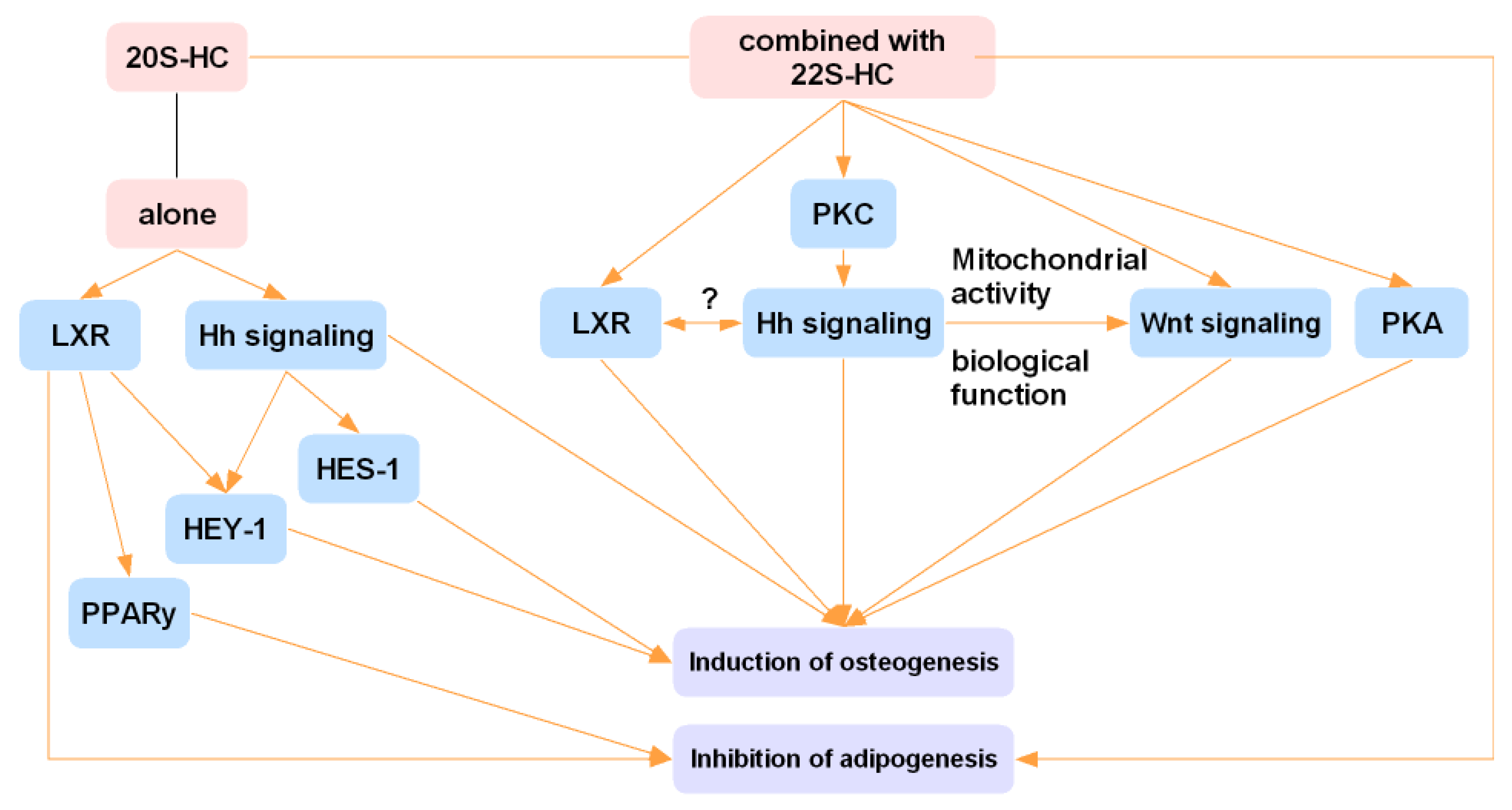
4.1. 20S-HC and 22S-HC
4.2. 27-HC
4.3. Other Cholesterol Oxidation Products
5. Research Challenges
5.1. COPs Standard Compounds
5.2. COPs Extraction in Biological Sample
5.3. Targeted Quantitation Approach
6. Conclusions and Perspectives
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
References
- Raisz, L.G. Pathogenesis of osteoporosis: Concepts, conflicts, and prospects. J. Clin. Investig. 2005, 115, 3318–3325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Xie, Z.; Sun, J.; Huang, S.; Chen, Y.; Li, C.; Sun, X.; Xia, B.; Tian, L.; Guo, C.; et al. Gut Microbiome Reveals Specific Dysbiosis in Primary Osteoporosis. Front. Cell. Infect. Microbiol. 2020, 10, 160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; Xu, S.; Zhang, B.; Xiao, C.; Chen, Z.; Si, F.; Fu, J.; Lin, X.; Zheng, G.; Yu, G.; et al. Gut microbiota and metabolite alterations associated with reduced bone mineral density or bone metabolic indexes in postmenopausal osteoporosis. Aging 2020, 12, 8583–8604. [Google Scholar] [CrossRef] [PubMed]
- Das, M.; Cronin, O.; Keohane, D.M.; Cormac, E.M.; Nugent, H.; Nugent, M.; Molloy, C.; O’Toole, P.W.; Shanahan, F.; Molloy, M.G.; et al. Gut microbiota alterations associated with reduced bone mineral density in older adults. Rheumatology 2019, 58, 2295–2304. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.L.; Shen, H.; Liu, A.; Dong, S.S.; Zhang, L.; Deng, F.Y.; Zhao, Q.; Deng, H.W. A road map for understanding molecular and genetic determinants of osteoporosis. Nat. Rev. Endocrinol. 2020, 16, 91–103. [Google Scholar] [CrossRef]
- Yin, W.; Li, Z.; Zhang, W. Modulation of Bone and Marrow Niche by Cholesterol. Nutrients 2019, 11, 1394. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.R.; Ko, N.Y.; Chen, K.H. Medical Treatment for Osteoporosis: From Molecular to Clinical Opinions. Int. J. Mol. Sci. 2019, 20, 2213. [Google Scholar] [CrossRef] [Green Version]
- Kha, H.T.; Basseri, B.; Shouhed, D.; Richardson, J.; Tetradis, S.; Hahn, T.J.; Parhami, F. Oxysterols regulate differentiation of mesenchymal stem cells: Pro-bone and anti-fat. J. Bone Miner. Res. 2004, 19, 830–840. [Google Scholar] [CrossRef] [Green Version]
- Lyons, M.A.; Samman, S.; Gatto, L.; Brown, A.J. Rapid hepatic metabolism of 7-ketocholesterol in vivo: Implications for dietary oxysterols. J. Lipid Res. 1999, 40, 1846–1857. [Google Scholar] [CrossRef]
- Brown, A.J.; Jessup, W. Oxysterols: Sources, cellular storage and metabolism, and new insights into their roles in cholesterol homeostasis. Mol. Aspects Med. 2009, 30, 111–122. [Google Scholar] [CrossRef]
- Griffiths, W.J.; Wang, Y. An update on oxysterol biochemistry: New discoveries in lipidomics. Biochem. Biophys. Res. Commun. 2018, 504, 617–622. [Google Scholar] [CrossRef] [PubMed]
- Kulig, W.; Cwiklik, L.; Jurkiewicz, P.; Rog, T.; Vattulainen, I. Cholesterol oxidation products and their biological importance. Chem. Phys. Lipids 2016, 199, 144–160. [Google Scholar] [CrossRef] [PubMed]
- Luu, W.; Sharpe, L.J.; Capell-Hattam, I.; Gelissen, I.C.; Brown, A.J. Oxysterols: Old Tale, New Twists. Annu. Rev. Pharmacol. Toxicol. 2016, 56, 447–467. [Google Scholar] [CrossRef] [PubMed]
- Sottero, B.; Rossin, D.; Staurenghi, E.; Gamba, P.; Poli, G.; Testa, G. Omics analysis of oxysterols to better understand their pathophysiological role. Free Radic. Biol. Med. 2019, 144, 55–71. [Google Scholar] [CrossRef]
- Gill, S.; Chow, R.; Brown, A.J. Sterol regulators of cholesterol homeostasis and beyond: The oxysterol hypothesis revisited and revised. Prog. Lipid Res. 2008, 47, 391–404. [Google Scholar] [CrossRef]
- Ma, L.; Nelson, E.R. Oxysterols and nuclear receptors. Mol. Cell. Endocrinol. 2019, 484, 42–51. [Google Scholar] [CrossRef]
- Zarrouk, A.; Vejux, A.; Mackrill, J.; O’Callaghan, Y.; Hammami, M.; O’Brien, N.; Lizard, G. Involvement of oxysterols in age-related diseases and ageing processes. Ageing Res. Rev. 2014, 18, 148–162. [Google Scholar] [CrossRef]
- Anderson, A.; Campo, A.; Fulton, E.; Corwin, A.; Jerome, W.G., 3rd; O’Connor, M.S. 7-Ketocholesterol in disease and aging. Redox Biol. 2020, 29, 101380. [Google Scholar] [CrossRef]
- Guillemot-Legris, O.; Mutemberezi, V.; Muccioli, G.G. Oxysterols in Metabolic Syndrome: From Bystander Molecules to Bioactive Lipids. Trends. Mol. Med. 2016, 22, 594–614. [Google Scholar] [CrossRef]
- Nelson, E.R.; Wardell, S.E.; McDonnell, D.P. The molecular mechanisms underlying the pharmacological actions of estrogens, SERMs and oxysterols: Implications for the treatment and prevention of osteoporosis. Bone 2013, 53, 42–50. [Google Scholar] [CrossRef] [Green Version]
- Nelson, E.R.; DuSell, C.D.; Wang, X.; Howe, M.K.; Evans, G.; Michalek, R.D.; Umetani, M.; Rathmell, J.C.; Khosla, S.; Gesty-Palmer, D.; et al. The oxysterol, 27-hydroxycholesterol, links cholesterol metabolism to bone homeostasis through its actions on the estrogen and liver X receptors. Endocrinology 2011, 152, 4691–4705. [Google Scholar] [CrossRef] [PubMed]
- Aghaloo, T.L.; Amantea, C.M.; Cowan, C.M.; Richardson, J.A.; Wu, B.M.; Parhami, F.; Tetradis, S. Oxysterols enhance osteoblast differentiation in vitro and bone healing in vivo. J. Orthop. Res. 2007, 25, 1488–1497. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Li, Z.; Hu, Y. Prevalence of osteoporosis in China: A meta-analysis and systematic review. BMC Public Health 2016, 16, 1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuggle, N.R.; Curtis, E.M.; Ward, K.A.; Harvey, N.C.; Dennison, E.M.; Cooper, C. Fracture prediction, imaging and screening in osteoporosis. Nat. Rev. Endocrinol. 2019, 15, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Compston, J.E.; McClung, M.R.; Leslie, W.D. Osteoporosis. Lancet 2019, 393, 364–376. [Google Scholar] [CrossRef]
- Deeks, E.D. Denosumab: A Review in Postmenopausal Osteoporosis. Drugs Aging. 2018, 35, 163–173. [Google Scholar] [CrossRef]
- Alonso-Bouzon, C.; Duque, G. Senile osteoporosis: An update. Rev. Esp. Geriatr. Gerontol. 2011, 46, 223–229. [Google Scholar] [CrossRef]
- Stein, E.; Shane, E. Secondary osteoporosis. Endocrinol. Metab. Clin. N. Am. 2003, 32, 115–134. [Google Scholar] [CrossRef]
- Kwan, P. Osteoporosis: From osteoscience to neuroscience and beyond. Mech. Ageing Dev. 2015, 145, 26–38. [Google Scholar] [CrossRef]
- Yang, D.; Wan, Y. Molecular determinants for the polarization of macrophage and osteoclast. Semin. Immunopathol. 2019, 41, 551–563. [Google Scholar] [CrossRef]
- Udagawa, N.; Koide, M.; Nakamura, M.; Nakamichi, Y.; Yamashita, T.; Uehara, S.; Kobayashi, Y.; Furuya, Y.; Yasuda, H.; Fukuda, C.; et al. Osteoclast differentiation by RANKL and OPG signaling pathways. J. Bone. Miner. Metab. 2021, 39, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Nagy, V.; Penninger, J.M. The RANKL-RANK Story. Gerontology 2015, 61, 534–542. [Google Scholar] [CrossRef] [PubMed]
- Wada, T.; Nakashima, T.; Hiroshi, N.; Penninger, J.M. RANKL-RANK signaling in osteoclastogenesis and bone disease. Trends Mol. Med. 2006, 12, 17–25. [Google Scholar] [CrossRef]
- Tanaka, S. Signaling axis in osteoclast biology and therapeutic targeting in the RANKL/RANK/OPG system. Am. J. Nephrol. 2007, 27, 466–478. [Google Scholar] [CrossRef] [PubMed]
- Takayanagi, H. RANKL as the master regulator of osteoclast differentiation. J. Bone Miner. Metab. 2021, 39, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Yin, C.; Zhao, F.; Ali, A.; Ma, J.; Qian, A. Mesenchymal Stem Cells: Cell Fate Decision to Osteoblast or Adipocyte and Application in Osteoporosis Treatment. Int. J. Mol. Sci. 2018, 19, 360. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Jin, D.; Xie, W.; Wen, L.; Chen, W.; Xu, J.; Ding, J.; Ren, D. PPAR-gamma and Wnt Regulate the Differentiation of MSCs into Adipocytes and Osteoblasts Respectively. Curr. Stem Cell Res. Ther. 2018, 13, 185–192. [Google Scholar] [CrossRef]
- Takada, I.; Kouzmenko, A.P.; Kato, S. Wnt and PPARgamma signaling in osteoblastogenesis and adipogenesis. Nat. Rev. Rheumatol. 2009, 5, 442–447. [Google Scholar] [CrossRef]
- Baron, R.; Rawadi, G.; Roman-Roman, S. Wnt signaling: A key regulator of bone mass. Curr. Top. Dev. Biol. 2006, 76, 103–127. [Google Scholar]
- Day, T.F.; Yang, Y. Wnt and hedgehog signaling pathways in bone development. J. Bone Joint Surg. Am. 2008, 90 (Suppl. 1), 19–24. [Google Scholar] [CrossRef]
- Rodda, S.J.; McMahon, A.P. Distinct roles for Hedgehog and canonical Wnt signaling in specification, differentiation and maintenance of osteoblast progenitors. Development 2006, 133, 3231–3244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suh, J.M.; Gao, X.; McKay, J.; McKay, R.; Salo, Z.; Graff, J.M. Hedgehog signaling plays a conserved role in inhibiting fat formation. Cell Metab. 2006, 3, 25–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Jie, Q.; Zhang, H.; Zhao, Y.; Lin, Y.; Du, J.; Shi, J.; Wang, L.; Guo, K.; Li, Y.; et al. Disturbed MEK/ERK signaling increases osteoclast activity via the Hedgehog-Gli pathway in postmenopausal osteoporosis. Prog. Biophys. Mol. Biol. 2016, 122, 101–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, G.L.; Hankenson, K.D. Integration of BMP, Wnt, and notch signaling pathways in osteoblast differentiation. J. Cell. Biochem. 2011, 112, 3491–3501. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Wang, Z.; Duan, N.; Zhu, G.; Schwarz, E.M.; Xie, C. Osteoblast-osteoclast interactions. Connect. Tissue Res. 2018, 59, 99–107. [Google Scholar] [CrossRef]
- Kim, J.M.; Lin, C.; Stavre, Z.; Greenblatt, M.B.; Shim, J.H. Osteoblast-Osteoclast Communication and Bone Homeostasis. Cells 2020, 9, 2073. [Google Scholar] [CrossRef]
- Zhao, H.; Li, Y.; Zhang, M.; Qi, L.; Tang, Y. Blood lipid levels in patients with osteopenia and osteoporosis:a systematic review and meta-analysis. J. Bone Miner. Metab. 2021, 39, 510–520. [Google Scholar] [CrossRef]
- Li, K.; Xiu, C.; Zhou, Q.; Ni, L.I.; Du, J.; Gong, T.; Li, M.; Yang, H.; Chen, J. A dual role of cholesterol in osteogenic differentiation of bone marrow stromal cells. J. Cell. Physiol. 2019, 234, 2058–2066. [Google Scholar] [CrossRef]
- Dias, I.H.K.; Milic, I.; Lip, G.Y.H.; Devitt, A.; Polidori, M.C.; Griffiths, H.R. Simvastatin reduces circulating oxysterol levels in men with hypercholesterolaemia. Redox Biol. 2018, 16, 139–145. [Google Scholar] [CrossRef]
- Griffiths, W.J.; Wang, Y. Oxysterols as lipid mediators: Their biosynthetic genes, enzymes and metabolites. Prostaglandins Other Lipid Mediat. 2020, 147, 106381. [Google Scholar] [CrossRef]
- Schweizer, R.A.; Zurcher, M.; Balazs, Z.; Dick, B.; Odermatt, A. Rapid hepatic metabolism of 7-ketocholesterol by 11beta-hydroxysteroid dehydrogenase type 1: Species-specific differences between the rat, human, and hamster enzyme. J. Biol. Chem. 2004, 279, 18415–18424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjorkhem, I.; Diczfalusy, U.; Lovgren-Sandblom, A.; Starck, L.; Jonsson, M.; Tallman, K.; Schirmer, H.; Ousager, L.B.; Crick, P.J.; Wang, Y.; et al. On the formation of 7-ketocholesterol from 7-dehydrocholesterol in patients with CTX and SLO. J. Lipid Res. 2014, 55, 1165–1172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Yutuc, E.; Griffiths, W.J. Neuro-oxysterols and neuro-sterols as ligands to nuclear receptors, GPCRs, ligand-gated ion channels and other protein receptors. Br. J. Pharmacol. 2021, 178, 3176–3193. [Google Scholar] [CrossRef] [PubMed]
- Mutemberezi, V.; Guillemot-Legris, O.; Muccioli, G.G. Oxysterols: From cholesterol metabolites to key mediators. Prog. Lipid Res. 2016, 64, 152–169. [Google Scholar] [CrossRef]
- Hahn, C.; Reichel, C.; von Bergmann, K. Serum concentration of 7 alpha-hydroxycholesterol as an indicator of bile acid synthesis in humans. J. Lipid Res. 1995, 36, 2059–2066. [Google Scholar] [CrossRef]
- Meaney, S.; Hassan, M.; Sakinis, A.; Lutjohann, D.; von Bergmann, K.; Wennmalm, A.; Diczfalusy, U.; Bjorkhem, I. Evidence that the major oxysterols in human circulation originate from distinct pools of cholesterol: A stable isotope study. J. Lipid Res. 2001, 42, 70–78. [Google Scholar] [CrossRef]
- Schroepfer, G.J., Jr. Oxysterols: Modulators of cholesterol metabolism and other processes. Physiol. Rev. 2000, 80, 361–554. [Google Scholar] [CrossRef]
- Bodin, K.; Andersson, U.; Rystedt, E.; Ellis, E.; Norlin, M.; Pikuleva, I.; Eggertsen, G.; Bjorkhem, I.; Diczfalusy, U. Metabolism of 4 beta -hydroxycholesterol in humans. J. Biol. Chem. 2002, 277, 31534–31540. [Google Scholar] [CrossRef] [Green Version]
- Heverin, M.; Meaney, S.; Lutjohann, D.; Diczfalusy, U.; Wahren, J.; Bjorkhem, I. Crossing the barrier: Net flux of 27-hydroxycholesterol into the human brain. J. Lipid Res. 2005, 46, 1047–1052. [Google Scholar] [CrossRef] [Green Version]
- Bjorkhem, I.; Lutjohann, D.; Diczfalusy, U.; Stahle, L.; Ahlborg, G.; Wahren, J. Cholesterol homeostasis in human brain: Turnover of 24S-hydroxycholesterol and evidence for a cerebral origin of most of this oxysterol in the circulation. J. Lipid Res. 1998, 39, 1594–1600. [Google Scholar] [CrossRef]
- Nelson, J.A.; Steckbeck, S.R.; Spencer, T.A. Biosynthesis of 24,25-epoxycholesterol from squalene 2,3;22,23-dioxide. J. Biol. Chem. 1981, 256, 1067–1068. [Google Scholar] [CrossRef]
- Spencer, T.A.; Gayen, A.K.; Phirwa, S.; Nelson, J.A.; Taylor, F.R.; Kandutsch, A.A.; Erickson, S.K. 24(S),25-Epoxycholesterol. Evidence consistent with a role in the regulation of hepatic cholesterogenesis. J. Biol. Chem. 1985, 260, 13391–13394. [Google Scholar] [CrossRef]
- Griffiths, W.J.; Abdel-Khalik, J.; Crick, P.J.; Yutuc, E.; Wang, Y. New methods for analysis of oxysterols and related compounds by LC-MS. J. Steroid Biochem. Mol. Biol. 2016, 162, 4–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otaegui-Arrazola, A.; Menendez-Carreno, M.; Ansorena, D.; Astiasaran, I. Oxysterols: A world to explore. Food Chem. Toxicol. 2010, 48, 3289–3303. [Google Scholar] [CrossRef] [PubMed]
- Leonarduzzi, G.; Sottero, B.; Poli, G. Oxidized products of cholesterol: Dietary and metabolic origin, and proatherosclerotic effects(review). J. Nutr. Biochem. 2002, 13, 700–710. [Google Scholar] [CrossRef]
- Bjorkhem, I.; Reihner, E.; Angelin, B.; Ewerth, S.; Akerlund, J.E.; Einarsson, K. On the possible use of the serum level of 7 alpha-hydroxycholesterol as a marker for increased activity of the cholesterol 7 alpha-hydroxylase in humans. J. Lipid Res. 1987, 28, 889–894. [Google Scholar] [CrossRef]
- Zhang, L.; Lv, Y.; Xian, G.; Lin, Y. 25-hydroxycholesterol promotes RANKL-induced osteoclastogenesis through coordinating NFATc1 and Sp1 complex in the transcription of miR-139–5p. Biochem. Biophys. Res. Commun. 2017, 485, 736–741. [Google Scholar] [CrossRef]
- Smith, L.L. Cholesterol autoxidation 1981–1986. Chem. Phys. Lipids 1987, 44, 87–125. [Google Scholar] [CrossRef]
- Norlin, M.; Toll, A.; Bjorkhem, I.; Wikvall, K. 24-hydroxycholesterol is a substrate for hepatic cholesterol 7alpha-hydroxylase(CYP7A). J. Lipid Res. 2000, 41, 1629–1639. [Google Scholar] [CrossRef]
- Duane, W.C.; Javitt, N.B. 27-hydroxycholesterol: Production rates in normal human subjects. J. Lipid Res. 1999, 40, 1194–1199. [Google Scholar] [CrossRef]
- Cao, Q.; Liu, Z.; Xiong, Y.; Zhong, Z.; Ye, Q. Multiple Roles of 25-Hydroxycholesterol in Lipid Metabolism, Antivirus Process, Inflammatory Response, and Cell Survival. Oxid. Med. Cell. Longev. 2020, 2020, 8893305. [Google Scholar] [CrossRef] [PubMed]
- Pandak, W.M.; Kakiyama, G. The acidic pathway of bile acid synthesis: Not just an alternative pathway. Liver Res. 2019, 3, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Yamamuro, D.; Yamazaki, H.; Osuga, J.I.; Okada, K.; Wakabayashi, T.; Takei, A.; Takei, S.; Takahashi, M.; Nagashima, S.; Holleboom, A.G.; et al. Esterification of 4beta-hydroxycholesterol and other oxysterols in human plasma occurs independently of LCAT. J. Lipid Res. 2020, 61, 1287–1299. [Google Scholar] [CrossRef]
- Lee, J.S.; Kim, E.; Han, S.; Kang, K.L.; Heo, J.S. Evaluating the oxysterol combination of 22(S)-hydroxycholesterol and 20(S)-hydroxycholesterol in periodontal regeneration using periodontal ligament stem cells and alveolar bone healing models. Stem Cell. Res. Ther. 2017, 8, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenhill, C. Bone: High 27-hydroxycholesterol decreases BMD. Nat. Rev. Endocrinol. 2010, 6, 474. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.K.; Meliton, V.; Tetradis, S.; Weinmaster, G.; Hahn, T.J.; Carlson, M.; Nelson, S.F.; Parhami, F. Osteogenic oxysterol, 20(S)-hydroxycholesterol, induces notch target gene expression in bone marrow stromal cells. J. Bone Miner. Res. 2010, 25, 782–795. [Google Scholar] [CrossRef] [Green Version]
- Yalom, A.; Hokugo, A.; Sorice, S.; Li, A.; Segovia Aguilar, L.A.; Zuk, P.; Jarrahy, R. In vitro osteoinductive effects of hydroxycholesterol on human adipose-derived stem cells are mediated through the hedgehog signaling pathway. Plast. Reconstr. Surg. 2014, 134, 960–968. [Google Scholar] [CrossRef]
- Kim, W.K.; Meliton, V.; Amantea, C.M.; Hahn, T.J.; Parhami, F. 20(S)-hydroxycholesterol inhibits PPARgamma expression and adipogenic differentiation of bone marrow stromal cells through a hedgehog-dependent mechanism. J. Bone Miner. Res. 2007, 22, 1711–1719. [Google Scholar] [CrossRef]
- Adhikari, R.; Chen, C.; Kim, W.K. Effect of 20(S)-Hydroxycholesterol on Multilineage Differentiation of Mesenchymal Stem Cells Isolated from Compact Bones in Chicken. Genes. 2020, 11, 1360. [Google Scholar] [CrossRef]
- Richardson, J.A.; Amantea, C.M.; Kianmahd, B.; Tetradis, S.; Lieberman, J.R.; Hahn, T.J.; Parhami, F. Oxysterol-induced osteoblastic differentiation of pluripotent mesenchymal cells is mediated through a PKC- and PKA-dependent pathway. J. Cell. Biochem. 2007, 100, 1131–1145. [Google Scholar] [CrossRef]
- Moseti, D.; Regassa, A.; Chen, C.; O, K.; Kim, W.K. 25-Hydroxycholesterol Inhibits Adipogenic Differentiation of C3H10T1/2 Pluripotent Stromal Cells. Int. J. Mol. Sci. 2020, 21, 412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shouhed, D.; Kha, H.T.; Richardson, J.A.; Amantea, C.M.; Hahn, T.J.; Parhami, F. Osteogenic oxysterols inhibit the adverse effects of oxidative stress on osteogenic differentiation of marrow stromal cells. J. Cell. Biochem. 2005, 95, 1276–1283. [Google Scholar] [CrossRef] [PubMed]
- Dwyer, J.R.; Sever, N.; Carlson, M.; Nelson, S.F.; Beachy, P.A.; Parhami, F. Oxysterols are novel activators of the hedgehog signaling pathway in pluripotent mesenchymal cells. J. Biol. Chem. 2007, 282, 8959–8968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, I.K.; Lee, S.C.; Hwang, Y.S.; Heo, J.S. Mitochondrial function contributes to oxysterol-induced osteogenic differentiation in mouse embryonic stem cells. Biochim. Biophys. Acta 2015, 1853, 561–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakshi, R.; Hokugo, A.; Zhou, S.; Zhang, Z.; Wang, L.; Rezzadeh, K.; Segovia, L.A.; Jarrahy, R. Application of Hydroxycholesterols for Alveolar Cleft Osteoplasty in a Rodent Model. Plast. Reconstr. Surg. 2019, 143, 1385–1395. [Google Scholar] [CrossRef] [PubMed]
- Kotani, T. Protein kinase A activity and Hedgehog signaling pathway. Vitam. Horm. 2012, 88, 273–291. [Google Scholar]
- Warner, M.; Nilsson, S.; Gustafsson, J.A. The estrogen receptor family. Curr. Opin. Obstet. Gynecol. 1999, 11, 249–254. [Google Scholar] [CrossRef]
- Robinson, J.A.; Harris, S.A.; Riggs, B.L.; Spelsberg, T.C. Estrogen regulation of human osteoblastic cell proliferation and differentiation. Endocrinology 1997, 138, 2919–2927. [Google Scholar] [CrossRef]
- Pakdel, F. Molecular Pathways of Estrogen Receptor Action. Int. J. Mol. Sci. 2018, 19, 2591. [Google Scholar] [CrossRef] [Green Version]
- Jia, M.; Dahlman-Wright, K.; Gustafsson, J.A. Estrogen receptor alpha and beta in health and disease. Best. Pract. Res. Clin. Endocrinol. Metab. 2015, 29, 557–568. [Google Scholar] [CrossRef]
- Umetani, M.; Domoto, H.; Gormley, A.K.; Yuhanna, I.S.; Cummins, C.L.; Javitt, N.B.; Korach, K.S.; Shaul, P.W.; Mangelsdorf, D.J. 27-Hydroxycholesterol is an endogenous SERM that inhibits the cardiovascular effects of estrogen. Nat. Med. 2007, 13, 1185–1192. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, M.; Liu, J.; Li, X.; Yang, M.; Su, B.; Lin, Y. 27-Hydroxycholesterol enhanced osteoclastogenesis in lung adenocarcinoma microenvironment. J. Cell. Physiol. 2019, 234, 12692–12700. [Google Scholar] [CrossRef] [PubMed]
- DuSell, C.D.; Nelson, E.R.; Wang, X.; Abdo, J.; Modder, U.I.; Umetani, M.; Gesty-Palmer, D.; Javitt, N.B.; Khosla, S.; McDonnell, D.P. The endogenous selective estrogen receptor modulator 27-hydroxycholesterol is a negative regulator of bone homeostasis. Endocrinology 2010, 151, 3675–3685. [Google Scholar] [CrossRef] [Green Version]
- Chang, P.Y.; Feldman, D.; Stefanick, M.L.; McDonnell, D.P.; Thompson, B.M.; McDonald, J.G.; Lee, J.S. 27-Hydroxycholesterol, an Endogenous SERM, and Risk of Fracture in Postmenopausal Women: A Nested Case-Cohort Study in the Women’s Health Initiative. J. Bone Miner. Res. 2019, 34, 59–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nevius, E.; Pinho, F.; Dhodapkar, M.; Jin, H.; Nadrah, K.; Horowitz, M.C.; Kikuta, J.; Ishii, M.; Pereira, J.P. Oxysterols and EBI2 promote osteoclast precursor migration to bone surfaces and regulate bone mass homeostasis. J. Exp. Med. 2015, 212, 1931–1946. [Google Scholar] [CrossRef] [PubMed]
- Paz, J.L.; Levy, D.; Oliveira, B.A.; de Melo, T.C.; de Freitas, F.A.; Reichert, C.O.; Rodrigues, A.; Pereira, J.; Bydlowski, S.P. 7-Ketocholesterol Promotes Oxiapoptophagy in Bone Marrow Mesenchymal Stem Cell from Patients with Acute Myeloid Leukemia. Cells 2019, 8, 482. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Ishihara, N.; Nagayama, D.; Saiki, A.; Tatsuno, I. 7-ketocholesterol induces apoptosis of MC3T3-E1 cells associated with reactive oxygen species generation, endoplasmic reticulum stress and caspase-3/7 dependent pathway. Mol. Genet. Metab. Rep. 2017, 10, 56–60. [Google Scholar] [CrossRef]
- Sul, O.J.; Li, G.; Kim, J.E.; Kim, E.S.; Choi, H.S. 7-ketocholesterol enhances autophagy via the ROS-TFEB signaling pathway in osteoclasts. J. Nutr. Biochem. 2021, 96, 108783. [Google Scholar] [CrossRef]
- Jiang, X.; Sidhu, R.; Porter, F.D.; Yanjanin, N.M.; Speak, A.O.; te Vruchte, D.T.; Platt, F.M.; Fujiwara, H.; Scherrer, D.E.; Zhang, J.; et al. A sensitive and specific LC-MS/MS method for rapid diagnosis of Niemann-Pick C1 disease from human plasma. J. Lipid Res. 2011, 52, 1435–1445. [Google Scholar] [CrossRef] [Green Version]
- Reunert, J.; Lotz-Havla, A.S.; Polo, G.; Kannenberg, F.; Fobker, M.; Griese, M.; Mengel, E.; Muntau, A.C.; Schnabel, P.; Sommerburg, O.; et al. Niemann-Pick Type C-2 Disease: Identification by Analysis of Plasma Cholestane-3beta,5alpha,6beta-Triol and Further Insight into the Clinical Phenotype. JIMD Rep. 2015, 23, 17–26. [Google Scholar]
- Liu, H.; Yuan, L.; Xu, S.; Wang, K.; Zhang, T. Cholestane-3beta,5alpha,6beta-triol inhibits osteoblastic differentiation and promotes apoptosis of rat bone marrow stromal cells. J. Cell. Biochem. 2005, 96, 198–208. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Qian, C.; Chen, X.Z. A highly stereoselective synthesis of C-24 and C-25 oxysterols from desmosterol. Steroids 2016, 109, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, S.; Kakiyama, G.; Muto, A.; Hosoda, A.; Mitamura, K.; Ikegawa, S.; Hofmann, A.F.; Iida, T. A facile synthesis of C-24 and C-25 oxysterols by in situ generated ethyl(trifluoromethyl)dioxirane. Steroids 2009, 74, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.R.; Chai, D.; Wright, D. Synthesis of(25R)-26-hydroxycholesterol. Steroids 2002, 67, 1041–1044. [Google Scholar] [CrossRef]
- Offei, S.D.; Arman, H.D.; Yoshimoto, F.K. Chemical synthesis of 7alpha-hydroxycholest-4-en-3-one, a biomarker for irritable bowel syndrome and bile acid malabsorption. Steroids 2019, 151, 108449. [Google Scholar] [CrossRef]
- Weber, D.; Ni, Z.X.; Vetter, D.; Hoffmann, R.; Fedorova, M. Electrochemical oxidation of cholesterol: An easy way to generate numerous oxysterols in short reaction times. Eur. J. Lipid. Sci. Tech. 2016, 118, 325–331. [Google Scholar] [CrossRef]
- Georgiou, C.A.; Constantinou, M.S.; Kapnissi-Christodoulou, C.P. Sample preparation: A critical step in the analysis of cholesterol oxidation products. Food Chem. 2014, 145, 918–926. [Google Scholar] [CrossRef]
- Karuna, R.; Christen, I.; Sailer, A.W.; Bitsch, F.; Zhang, J. Detection of dihydroxycholesterols in human plasma using HPLC-ESI-MS/MS. Steroids 2015, 99, 131–138. [Google Scholar] [CrossRef]
- Ulmer, C.Z.; Jones, C.M.; Yost, R.A.; Garrett, T.J.; Bowden, J.A. Optimization of Folch, Bligh-Dyer, and Matyash sample-to-extraction solvent ratios for human plasma-based lipidomics studies. Anal. Chim. Acta 2018, 1037, 351–357. [Google Scholar] [CrossRef]
- Brouwers, J.F.; Boerke, A.; Silva, P.F.; Garcia-Gil, N.; van Gestel, R.A.; Helms, J.B.; van de Lest, C.H.; Gadella, B.M. Mass spectrometric detection of cholesterol oxidation in bovine sperm. Biol. Reprod. 2011, 85, 128–136. [Google Scholar] [CrossRef] [Green Version]
- Adamek, M.; Davies, J.; Beck, A.; Jordan, L.; Becker, A.M.; Mojzesz, M.; Rakus, K.; Rumiac, T.; Collet, B.; Brogden, G.; et al. Antiviral Actions of 25-Hydroxycholesterol in Fish Vary with the Virus-Host Combination. Front. Immunol. 2021, 12, 581786. [Google Scholar] [CrossRef] [PubMed]
- Mendiara, I.; Domeno, C.; Nerin, C.; Geurts, A.M.; Osada, J.; Martinez-Beamonte, R. Determination of total plasma oxysterols by enzymatic hydrolysis, solid phase extraction and liquid chromatography coupled to mass-spectrometry. J. Pharm. Biomed. Anal. 2018, 150, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.; Siegmund, W.; Oswald, S. Rapid LC-MS/MS method for the determination of 4-hydroxycholesterol/cholesterol ratio in serum as endogenous biomarker for CYP3A activity in human and foals. J. Chromatogr. B 2016, 1033–1034, 193–199. [Google Scholar] [CrossRef]
- Sidhu, R.; Jiang, H.; Farhat, N.Y.; Carrillo-Carrasco, N.; Woolery, M.; Ottinger, E.; Porter, F.D.; Schaffer, J.E.; Ory, D.S.; Jiang, X. A validated LC-MS/MS assay for quantification of 24(S)-hydroxycholesterol in plasma and cerebrospinal fluid. J. Lipid Res. 2015, 56, 1222–1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceglarek, U.; Dittrich, J.; Leopold, J.; Helmschrodt, C.; Becker, S.; Staab, H.; Richter, O.; Rohm, S.; Aust, G. Free cholesterol, cholesterol precursor and plant sterol levels in atherosclerotic plaques are independently associated with symptomatic advanced carotid artery stenosis. Atherosclerosis 2020, 295, 18–24. [Google Scholar] [CrossRef]
- Fellows Maxwell, K.; Bhattacharya, S.; Bodziak, M.L.; Jakimovski, D.; Hagemeier, J.; Browne, R.W.; Weinstock-Guttman, B.; Zivadinov, R.; Ramanathan, M. Oxysterols and apolipoproteins in multiple sclerosis: A 5 year follow-up study. J. Lipid Res. 2019, 60, 1190–1198. [Google Scholar] [CrossRef]
- Huang, X.; Sterling, N.W.; Du, G.; Sun, D.; Stetter, C.; Kong, L.; Zhu, Y.; Neighbors, J.; Lewis, M.M.; Chen, H.; et al. Brain cholesterol metabolism and Parkinson’s disease. Mov. Disord. 2019, 34, 386–395. [Google Scholar] [CrossRef]
- Lu, D.L.; Le Cornet, C.; Sookthai, D.; Johnson, T.S.; Kaaks, R.; Fortner, R.T. Circulating 27-Hydroxycholesterol and Breast Cancer Risk: Results From the EPIC-Heidelberg Cohort. J. Natl. Cancer Inst. 2019, 111, 365–371. [Google Scholar] [CrossRef]
- Zerbinati, C.; Caponecchia, L.; Puca, R.; Ciacciarelli, M.; Salacone, P.; Sebastianelli, A.; Pastore, A.; Palleschi, G.; Petrozza, V.; Porta, N.; et al. Mass spectrometry profiling of oxysterols in human sperm identifies 25-hydroxycholesterol as a marker of sperm function. Redox Biol. 2017, 11, 111–117. [Google Scholar] [CrossRef] [Green Version]
- Nury, T.; Zarrouk, A.; Ragot, K.; Debbabi, M.; Riedinger, J.M.; Vejux, A.; Aubourg, P.; Lizard, G. 7-Ketocholesterol is increased in the plasma of X-ALD patients and induces peroxisomal modifications in microglial cells: Potential roles of 7-ketocholesterol in the pathophysiology of X-ALD. J. Steroid Biochem. Mol. Biol. 2017, 169, 123–136. [Google Scholar] [CrossRef]
- Marcello, A.; Civra, A.; Milan Bonotto, R.; Nascimento Alves, L.; Rajasekharan, S.; Giacobone, C.; Caccia, C.; Cavalli, R.; Adami, M.; Brambilla, P.; et al. The cholesterol metabolite 27-hydroxycholesterol inhibits SARS-CoV-2 and is markedly decreased in COVID-19 patients. Redox Biol. 2020, 36, 101682. [Google Scholar] [CrossRef] [PubMed]
- Hole, K.; Heiberg, P.L.; Gjestad, C.; Mehus, L.L.; Ro, O.; Molden, E. Elevated 4beta-hydroxycholesterol/cholesterol ratio in anorexia nervosa patients. Pharmacol. Res. Perspect. 2018, 6, e00430. [Google Scholar] [CrossRef] [PubMed]
- Wuolikainen, A.; Acimovic, J.; Lovgren-Sandblom, A.; Parini, P.; Andersen, P.M.; Bjorkhem, I. Cholesterol, oxysterol, triglyceride, and coenzyme Q homeostasis in ALS. Evidence against the hypothesis that elevated 27-hydroxycholesterol is a pathogenic factor. PLoS ONE 2014, 9, e113619. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.C.; Joaquim, H.P.G.; Nunes, V.S.; Kerr, D.S.; Ferreira, G.S.; Forlenza, O.V.; Gattaz, W.F.; Talib, L.L. Donepezil effects on cholesterol and oxysterol plasma levels of Alzheimer’s disease patients. Eur. Arch. Psychiatry. Clin. Neurosci. 2018, 268, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Aksu, N.; Samadi, A.; Yalcinkaya, A.; Cetin, T.; Eser, B.; Lay, I.; Ozis, T.N.; Oztas, Y.; Sabuncuoglu, S. Evaluation of oxysterol levels of patients with silicosis by LC-MS/MS method. Mol. Cell. Biochem. 2020, 467, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Pataj, Z.; Liebisch, G.; Schmitz, G.; Matysik, S. Quantification of oxysterols in human plasma and red blood cells by liquid chromatography high-resolution tandem mass spectrometry. J. Chromatogr. A 2016, 1439, 82–88. [Google Scholar] [CrossRef]
- Klinke, G.; Rohrbach, M.; Giugliani, R.; Burda, P.; Baumgartner, M.R.; Tran, C.; Gautschi, M.; Mathis, D.; Hersberger, M. LC-MS/MS based assay and reference intervals in children and adolescents for oxysterols elevated in Niemann-Pick diseases. Clin. Biochem. 2015, 48, 596–602. [Google Scholar] [CrossRef]
- Samadi, A.; Gurlek, A.; Sendur, S.N.; Karahan, S.; Akbiyik, F.; Lay, I. Oxysterol species: Reliable markers of oxidative stress in diabetes mellitus. J. Endocrinol. Investig. 2019, 42, 7–17. [Google Scholar] [CrossRef]
- Samadi, A.; Isikhan, S.Y.; Tinkov, A.A.; Lay, I.; Dosa, M.D.; Skalny, A.V.; Skalnaya, M.G.; Chirumbolo, S.; Bjorklund, G. Zinc, copper, and oxysterol levels in patients with type 1 and type 2 diabetes mellitus. Clin. Nutr. 2020, 39, 1849–1856. [Google Scholar] [CrossRef]
- Honda, A.; Yamashita, K.; Hara, T.; Ikegami, T.; Miyazaki, T.; Shirai, M.; Xu, G.; Numazawa, M.; Matsuzaki, Y. Highly sensitive quantification of key regulatory oxysterols in biological samples by LC-ESI-MS/MS. J. Lipid Res. 2009, 50, 350–357. [Google Scholar] [CrossRef] [Green Version]
- Hirayama, T.; Ikegami, T.; Honda, A.; Miyazaki, T.; Yara, S.I.; Kohjima, M.; Nakamuta, M.; Matsuzaki, Y. Differences in the Serum 4beta-hydroxycholesterol Levels of Patients with Chronic Hepatitis C Virus (HCV) Infection: A Possible Impact on the Efficacy and Safety of Interferon(IFN)-free Treatment. Intern. Med. 2018, 57, 1219–1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marelli, C.; Lamari, F.; Rainteau, D.; Lafourcade, A.; Banneau, G.; Humbert, L.; Monin, M.L.; Petit, E.; Debs, R.; Castelnovo, G.; et al. Plasma oxysterols: Biomarkers for diagnosis and treatment in spastic paraplegia type 5. Brain 2018, 141, 72–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikegami, T.; Honda, A.; Miyazaki, T.; Kohjima, M.; Nakamuta, M.; Matsuzaki, Y. Increased serum oxysterol concentrations in patients with chronic hepatitis C virus infection. Biochem. Biophys. Res. Commun. 2014, 446, 736–740. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, W.J.; Hornshaw, M.; Woffendin, G.; Baker, S.F.; Lockhart, A.; Heidelberger, S.; Gustafsson, M.; Sjovall, J.; Wang, Y. Discovering oxysterols in plasma: A window on the metabolome. J. Proteome Res. 2008, 7, 3602–3612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raselli, T.; Hearn, T.; Wyss, A.; Atrott, K.; Peter, A.; Frey-Wagner, I.; Spalinger, M.R.; Maggio, E.M.; Sailer, A.W.; Schmitt, J.; et al. Elevated oxysterol levels in human and mouse livers reflect nonalcoholic steatohepatitis. J. Lipid Res. 2019, 60, 1270–1283. [Google Scholar] [CrossRef] [Green Version]
- Crick, P.J.; Griffiths, W.J.; Zhang, J.; Beibel, M.; Abdel-Khalik, J.; Kuhle, J.; Sailer, A.W.; Wang, Y. Reduced Plasma Levels of 25-Hydroxycholesterol and Increased Cerebrospinal Fluid Levels of Bile Acid Precursors in Multiple Sclerosis Patients. Mol. Neurobiol. 2017, 54, 8009–8020. [Google Scholar] [CrossRef] [Green Version]
- Yutuc, E.; Angelini, R.; Baumert, M.; Mast, N.; Pikuleva, I.; Newton, J.; Clench, M.R.; Skibinski, D.O.F.; Howell, O.W.; Wang, Y.J.; et al. Localization of sterols and oxysterols in mouse brain reveals distinct spatial cholesterol metabolism. Proc. Natl. Acad. Sci. USA 2020, 117, 5749–5760. [Google Scholar] [CrossRef] [Green Version]
- Lutjohann, D.; Bjorkhem, I.; Friedrichs, S.; Kerksiek, A.; Geilenkeuser, W.J.; Lovgren-Sandblom, A.; Ansorena, D.; Astiasaran, I.; Baila-Rueda, L.; Barriuso, B.; et al. International descriptive and interventional survey for oxycholesterol determination by gas- and liquid-chromatographic methods. Biochimie 2018, 153, 26–32. [Google Scholar] [CrossRef]
- Lutjohann, D.; Bjorkhem, I.; Friedrichs, S.; Kerksiek, A.; Lovgren-Sandblom, A.; Geilenkeuser, W.J.; Ahrends, R.; Andrade, I.; Ansorena, D.; Astiasaran, I.; et al. First international descriptive and interventional survey for cholesterol and non-cholesterol sterol determination by gas- and liquid-chromatography-Urgent need for harmonisation of analytical methods. J. Steroid. Biochem. Mol. Biol. 2019, 190, 115–125. [Google Scholar] [CrossRef]
- Li, L.; Ren, W.; Kong, H.; Zhao, C.; Zhao, X.; Lin, X.; Lu, X.; Xu, G. An alignment algorithm for LC-MS-based metabolomics dataset assisted by MS/MS information. Anal. Chim. Acta 2017, 990, 96–102. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| NO. | Analytical Aspects | Preparatiom Aspects | Analytes | Content | Population Sex | Population Age (Year) | Ref |
|---|---|---|---|---|---|---|---|
| 1 | HPLC-ESI-MS | PP Derivatization to N, N-dimethylglycine ester | 7-KC a | 0.050 ± 0.003 (µmol/L) | - | 43.6 ± 9.6 | [127] |
| C-triol a | 0.033 ± 0.004 (µmol/L) | ||||||
| 2 | HPLC-APCI-MS | LLE | 4β-HC a | 30.4 ± 19.9 ng/mL | - | 22–27 | [115] |
| 3 | HPLC-ESI-MS | LLE Derivatization to picolinyl ester | 4β-HC a | 77 ± 40 ng/mL | - | - | [132] |
| 7α-HC a | 145 ± 82 ng/mL | ||||||
| 22R-HC a | 10 ± 18 ng/mL | ||||||
| 24S-HC a | 51 ± 12 ng/mL | ||||||
| 25-HC a | 31 ± 11 ng/mL | ||||||
| 27-HC a | 117 ± 35 ng/mL | ||||||
| 24S,25-EC a | 2 ± 2 ng/mL | ||||||
| 4 | HPLC-ESI-MS | LLE Derivatization to N, N-dimethylglycine ester | 24-HC a | 38.1 ng/mL | - | - | [128] |
| 25-HC a | 29.7 ng/mL | ||||||
| 27-HC a | 74.1 ng/mL | ||||||
| 4β-HC a | 17.8 ng/mL | ||||||
| 7α-HC a | 29.3 ng/mL | ||||||
| 7β-HC a | 6.9 ng/mL | ||||||
| 7-KC a | 18.8 ng/mL | ||||||
| 5 | HPLC-ESI-MS/MS | PP Derivatization to N, N-dimethylglycine ester | 7-KC b | 3–100 ng/mL | - | 0–18 | [129] |
| C-triol b | 3–60 ng/mL | ||||||
| 6 | UFLC-ESI-MS | LLE Derivatization to picolinyl ester | 4β-HC a | 59.0 ± 3.79 ng/mL | female | - | [133] |
| 43.7 ± 2.31 ng/mL | male | - | |||||
| 7 | GC-MS/MS | LLE Derivatization to trimethylsilyl esters | 7α-HC a | 0.1 µmol/L | - | 19–48 | [124] |
| 7β-HC a | 0.1 µmol/L | ||||||
| 7β-HC a | 0.1 µmol/L | ||||||
| 8 | UPLC-ESI-MS/MS | LLE Derivatization to picolinyl ester | 24S-HC a | 65.7 ± 60.6 nmol/L | - | 29 ± 10 | [134] |
| 25-HC a | 15.3 ± 17.5 nmol/L | ||||||
| 27-HC a | 139.3 ± 76.3 nmol/L | ||||||
| 9 | GC-MS/MS | SPE Derivatization to trimethylsiyl ethers | 24S-HC a | 61.9 ± 14.1 ng/mL | female | - | [125] |
| 25-HC a | 6.4 ±2.2 ng/mL | ||||||
| 27-HC a | 0.14 ± 0.03 µg/mL | ||||||
| 7α-HCone a | 37.6 ± 27.8 nmol/mL | ||||||
| 24S-HC a | 61.3 ± 12.6 ng/mL | male | |||||
| 25-HC a | 7.5 ± 2.6 ng/mL | ||||||
| 27-HC a | 0.2 ± 0.05 µg/mL | ||||||
| 7α-HCone a | 68.3 ± 92.9 nmol/mL | ||||||
| 10 | HPLC-APCI-MS/MS | LLE | 7-HC b | 1.7–3.3 mg/L | - | average age 34 | [117] |
| 7-KC b | 3.0–6.9 mg/L | ||||||
| 5,6α-EC b | 0.9–1.9 mg/L | ||||||
| 5,6β-EC b | 3.5 ± 0.3 mg/mL | ||||||
| 11 | LC-ESI-MS/MS | PP Derivatization to N, N-dimethylglycine ester | 7-KC b | 18.33 ± 3.76 ng/mL | - | under the age of 40 | [130] |
| C-triol b | 9.39 ± 3.17 ng/mL | ||||||
| 7-KC b | 19.72 ± 2.47 ng/mL | - | over the age of 40 | ||||
| C-triol b | 10.62 ± 2.77 ng/mL | ||||||
| 12 | UFLC-ESI-MS | LLE Derivatization to picolinyl ester | 24S-HC a | 64.4 ± 1.8 ng/mL | - | average age 52.3 | [135] |
| 25-HC a | 14.9 ± 1.1 ng/mL | ||||||
| 27-HC a | 139.0 ± 4.7 ng/mL | ||||||
| 7α-HC a | 136.5 ± 12 ng/mL | ||||||
| 4β-HC a | 51.9 ± 2.4 ng/mL | ||||||
| 13 | HPLC-APCI-MS | SPE | 24-HC a | 67 ng/mL | - | average age 39 | [118] |
| 25-HC a | 12 ng/mL | ||||||
| 7α-HC a | 55 ng/ml | ||||||
| 27-HC a | 355 ng/mL | ||||||
| 7-KC a | 11 ng/mL | ||||||
| 27-HC b | 57 ng/mL | ||||||
| 14 | GC-MS/MS | LLE Derivatization to trimethylsilyl esters | 24S-HC a | 60.30 ± 14.24 ng/mL | - | 68.22 ± 9.17 | [123] |
| 25-HC a | 8.52 ± 2.58 ng/mL |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Che, Y.; Yang, J.; Tang, F.; Wei, Z.; Chao, Y.; Li, N.; Li, H.; Wu, S.; Dong, X. New Function of Cholesterol Oxidation Products Involved in Osteoporosis Pathogenesis. Int. J. Mol. Sci. 2022, 23, 2020. https://doi.org/10.3390/ijms23042020
Che Y, Yang J, Tang F, Wei Z, Chao Y, Li N, Li H, Wu S, Dong X. New Function of Cholesterol Oxidation Products Involved in Osteoporosis Pathogenesis. International Journal of Molecular Sciences. 2022; 23(4):2020. https://doi.org/10.3390/ijms23042020
Chicago/Turabian StyleChe, Yanting, Jingzhi Yang, Fen Tang, Ziheng Wei, Yufan Chao, Na Li, Henghui Li, Si Wu, and Xin Dong. 2022. "New Function of Cholesterol Oxidation Products Involved in Osteoporosis Pathogenesis" International Journal of Molecular Sciences 23, no. 4: 2020. https://doi.org/10.3390/ijms23042020
APA StyleChe, Y., Yang, J., Tang, F., Wei, Z., Chao, Y., Li, N., Li, H., Wu, S., & Dong, X. (2022). New Function of Cholesterol Oxidation Products Involved in Osteoporosis Pathogenesis. International Journal of Molecular Sciences, 23(4), 2020. https://doi.org/10.3390/ijms23042020