
Involvement of Histamine H3 Receptor Agonism in Premature Ejaculation Found by Studies in Rats

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. In Vivo Electrophysiological Recording from the Pelvic Nerve
2.1.1. Effects of Imetit on Mechanical Stimuli-Evoked Firing at the Peripheral Nerve
2.1.2. H3R Antagonism on the Suppressive Effect of Imetit on Mechanical Stimuli-Evoked Firing at the Peripheral Nerve
2.2. In Vivo Extracellular Recording of Neurons in the Deep Dorsal Horn
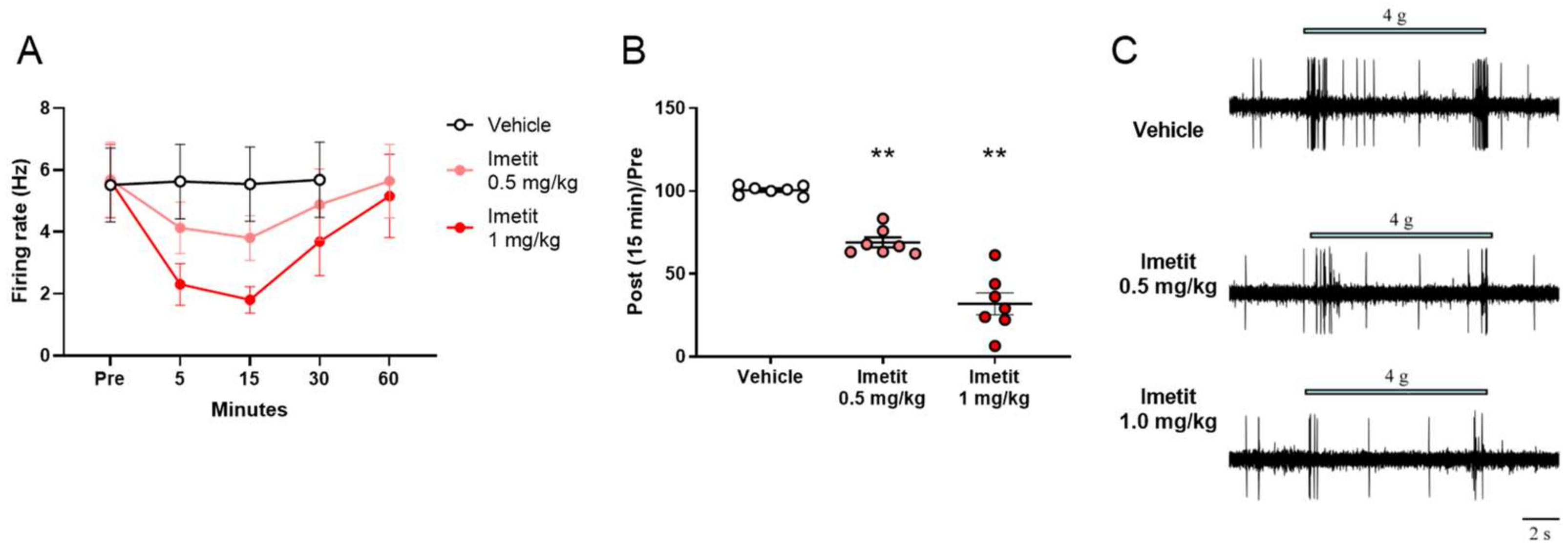
2.2.1. Effects of Imetit on Mechanical Stimuli-Evoked Firing of the Spinal Dorsal Horn
2.2.2. Dose-Dependent Effects of Imetit on Mechanical Stimuli-Evoked Firing of the Spinal Dorsal Horn
2.2.3. H3R and H4R Antagonism on the Suppressive Effects of Imetit on Mechanical Stimuli-Evoked Firing of the Spinal Dorsal Horn
2.3. Copulatory Behavior
2.3.1. Effects of Imetit on Ejaculation in Copulatory Behavior
2.3.2. H3R Antagonism on the Effect of Imetit to Prolong Ejaculation Latency
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Test Compounds
4.3. In Vivo Electrophysiological Recording from the Pelvic Nerve
4.4. In Vivo Extracellular Recording from the Deep Dorsal Horn Neurons
4.5. Copulatory Behavior Test
4.5.1. Procedure
4.5.2. Observation
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- ISSM Quick Reference Guide on Premature Ejaculation. Available online: https://www.issm.info/publications/clinical-guidelines (accessed on 1 December 2021).
- Serefoglu, E.C.; McMahon, C.G.; Waldinger, M.D.; Althof, S.E.; Shindel, A.; Adaikan, G.; Becher, E.F.; Dean, J.; Giuliano, F.; Hellstrom, W.J.; et al. An evidence-based unified definition of lifelong and acquired premature ejaculation: Report of the second international society for sexual medicine ad hoc committee for the definition of premature ejaculation. Sex. Med. 2014, 2, 41–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Althof, S.E.; McMahon, C.G.; Waldinger, M.D.; Serefoglu, E.C.; Shindel, A.W.; Adaikan, P.G.; Becher, E.; Dean, J.; Giuliano, F.; Hellstrom, W.J.; et al. An update of the International Society of Sexual Medicine’s guidelines for the diagnosis and treatment of premature ejaculation (PE). J. Sex. Med. 2014, 11, 1392–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondaini, N.; Fusco, F.; Cai, T.; Benemei, S.; Mirone, V.; Bartoletti, R. Dapoxetine treatment in patients with lifelong premature ejaculation: The reasons of a “Waterloo”. Urology 2013, 82, 620–624. [Google Scholar] [CrossRef] [PubMed]
- Waldinger, M.D. Pharmacotherapy for premature ejaculation. Expert Opin. Pharmacother. 2015, 16, 2615–2624. [Google Scholar] [CrossRef]
- Saleh, R.; Majzoub, A.; Abu El-Hamd, M. An update on the treatment of premature ejaculation: A systematic review. Arab J. Urol. 2021, 19, 281–302. [Google Scholar] [CrossRef] [PubMed]
- Althof, S.E. Prevalence, characteristics and implications of premature ejaculation/rapid ejaculation. J. Urol. 2006, 175, 842–848. [Google Scholar] [CrossRef]
- Nieto-Alamilla, G.; Marquez-Gomez, R.; Garcia-Galvez, A.M.; Morales-Figueroa, G.E.; Arias-Montano, J.A. The Histamine H3 Receptor: Structure, Pharmacology, and Function. Mol. Pharmacol. 2016, 90, 649–673. [Google Scholar] [CrossRef] [Green Version]
- Leurs, R.; Bakker, R.A.; Timmerman, H.; de Esch, I.J. The histamine H3 receptor: From gene cloning to H3 receptor drugs. Nat. Rev. Drug Discov. 2005, 4, 107–120. [Google Scholar] [CrossRef]
- Cannon, K.E.; Chazot, P.L.; Hann, V.; Shenton, F.; Hough, L.B.; Rice, F.L. Immunohistochemical localization of histamine H3 receptors in rodent skin, dorsal root ganglia, superior cervical ganglia, and spinal cord: Potential antinociceptive targets. Pain 2007, 129, 76–92. [Google Scholar] [CrossRef]
- Hough, L.B.; Rice, F.L. H3 receptors and pain modulation: Peripheral, spinal, and brain interactions. J. Pharmacol. Exp. Ther. 2011, 336, 30–37. [Google Scholar] [CrossRef] [Green Version]
- Cara, A.M.; Lopes-Martins, R.A.; Antunes, E.; Nahoum, C.R.; De Nucci, G. The role of histamine in human penile erection. Br. J. Urol. 1995, 75, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.C.; Davies, M.G.; Lee, T.H.; Hagen, P.O.; Carson, C.C., 3rd. Characterization and function of histamine receptors in corpus cavernosum. J. Urol. 1995, 153, 506–510. [Google Scholar] [CrossRef] [PubMed]
- Meston, C.M.; Frohlich, P.F. The neurobiology of sexual function. Arch. Gen. Psychiatry 2000, 57, 1012–1030. [Google Scholar] [CrossRef] [PubMed]
- Garbarg, M.; Arrang, J.M.; Rouleau, A.; Ligneau, X.; Tuong, M.D.; Schwartz, J.C.; Ganellin, C.R. S-[2-(4-imidazolyl)ethyl]isothiourea, a highly specific and potent histamine H3 receptor agonist. J. Pharmacol. Exp. Ther. 1992, 263, 304–310. [Google Scholar] [PubMed]
- Lim, H.D.; van Rijn, R.M.; Ling, P.; Bakker, R.A.; Thurmond, R.L.; Leurs, R. Evaluation of histamine H1-, H2-, and H3-receptor ligands at the human histamine H4 receptor: Identification of 4-methylhistamine as the first potent and selective H4 receptor agonist. J. Pharmacol. Exp. Ther. 2005, 314, 1310–1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ligneau, X.; Lin, J.; Vanni-Mercier, G.; Jouvet, M.; Muir, J.L.; Ganellin, C.R.; Stark, H.; Elz, S.; Schunack, W.; Schwartz, J. Neurochemical and behavioral effects of ciproxifan, a potent histamine H3-receptor antagonist. J. Pharmacol. Exp. Ther. 1998, 287, 658–666. [Google Scholar]
- Thurmond, R.L.; Desai, P.J.; Dunford, P.J.; Fung-Leung, W.P.; Hofstra, C.L.; Jiang, W.; Nguyen, S.; Riley, J.P.; Sun, S.; Williams, K.N.; et al. A potent and selective histamine H4 receptor antagonist with anti-inflammatory properties. J. Pharmacol. Exp. Ther. 2004, 309, 404–413. [Google Scholar] [CrossRef] [Green Version]
- Tohei, A.; Watanabe, G.; Taya, K. Effects of thyroidectomy or thiouracil treatment on copulatory behavior in adult male rats. J. Vet. Med. Sci. 1998, 60, 281–285. [Google Scholar] [CrossRef] [Green Version]
- Noguchi, M.; Kimura, T. Screening Method of Curative Medicine for Premature Ejaculation. J.P. Patent 2013024626, 4 February 2013. [Google Scholar]
- de Jong, T.R.; Pattij, T.; Veening, J.G.; Dederen, P.J.; Waldinger, M.D.; Cools, A.R.; Olivier, B. Citalopram combined with WAY 100635 inhibits ejaculation and ejaculation-related Fos immunoreactivity. Eur. J. Pharmacol. 2005, 509, 49–59. [Google Scholar] [CrossRef]
- Pellegrini-Quarantotti, B.; Paglietti, E.; Bonanni, A.; Petta, M.; Gessa, G.L. Naloxone shortens ejaculation latency in male rats. Experientia 1979, 35, 524–525. [Google Scholar] [CrossRef]
- Powell, W.S.; Dominguez, J.M.; Hull, E.M. An NMDA antagonist impairs copulation and the experience-induced enhancement of male sexual behavior in the rat. Behav. Neurosci. 2003, 117, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Giuliano, F.; Clement, P. Physiology of ejaculation: Emphasis on serotonergic control. Eur. Urol. 2005, 48, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Vignoli, G.C. Premature ejaculation: New electrophysiologic approach. Urology 1978, 11, 81–82. [Google Scholar] [CrossRef]
- Xin, Z.C.; Chung, W.S.; Choi, Y.D.; Seong, D.H.; Choi, Y.J.; Choi, H.K. Penile sensitivity in patients with primary premature ejaculation. J. Urol. 1996, 156, 979–981. [Google Scholar] [CrossRef]
- Halata, Z.; Munger, B.L. The neuroanatomical basis for the protopathic sensibility of the human glans penis. Brain Res. 1986, 371, 205–230. [Google Scholar] [CrossRef]
- Tiligada, E.; Kyriakidis, K.; Chazot, P.L.; Passani, M.B. Histamine pharmacology and new CNS drug targets. CNS Neurosci. Ther. 2011, 17, 620–628. [Google Scholar] [CrossRef]
- Obara, I.; Telezhkin, V.; Alrashdi, I.; Chazot, P.L. Histamine, histamine receptors, and neuropathic pain relief. Br. J. Pharmacol. 2020, 177, 580–599. [Google Scholar] [CrossRef]
- Cannon, K.E.; Hough, L.B. Inhibition of chemical and low-intensity mechanical nociception by activation of histamine H3 receptors. J. Pain Off. J. Am. Pain Soc. 2005, 6, 193–200. [Google Scholar] [CrossRef]
- Taylor, D.C.; Korf, H.W.; Pierau, F.K. Distribution of sensory neurons of the pudendal nerve in the dorsal root ganglia and their projection to the spinal cord. Horseradish-peroxidase studies in the rat. Cell Tissue Res. 1982, 226, 555–564. [Google Scholar] [CrossRef]
- Carro-Juarez, M.; Rodriguez-Manzo, G. The spinal pattern generator for ejaculation. Brain Res. Rev. 2008, 58, 106–120. [Google Scholar] [CrossRef]
- Hubscher, C.H. Ascending spinal pathways from sexual organs: Effects of chronic spinal lesions. Prog. Brain Res. 2006, 152, 401–414. [Google Scholar] [CrossRef]
- Todd, A.J. Neuronal circuitry for pain processing in the dorsal horn. Nat. Rev. Neurosci. 2010, 11, 823–836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uta, D.; Oti, T.; Sakamoto, T.; Sakamoto, H. In Vivo Electrophysiology of Peptidergic Neurons in Deep Layers of the Lumbar Spinal Cord after Optogenetic Stimulation of Hypothalamic Paraventricular Oxytocin Neurons in Rats. Int. J. Mol. Sci. 2021, 22, 3400. [Google Scholar] [CrossRef]
- Uta, D.; Kato, G.; Doi, A.; Andoh, T.; Kume, T.; Yoshimura, M.; Koga, K. Animal models of chronic pain increase spontaneous glutamatergic transmission in adult rat spinal dorsal horn in vitro and in vivo. Biochem. Biophys. Res. Commun. 2019, 512, 352–359. [Google Scholar] [CrossRef]
- Uta, D.; Koga, K.; Furue, H.; Imoto, K.; Yoshimura, M. L-bupivacaine Inhibition of Nociceptive Transmission in Rat Peripheral and Dorsal Horn Neurons. Anesthesiology 2021, 134, 88–102. [Google Scholar] [CrossRef] [PubMed]
- Uta, D.; Tsuboshima, K.; Nishijo, H.; Mizumura, K.; Taguchi, T. Neuronal Sensitization and Synaptic Facilitation in the Superficial Dorsal Horn of a Rat Reserpine-induced Pain Model. Neuroscience 2021, 479, 125–139. [Google Scholar] [CrossRef] [PubMed]
- Takanami, K.; Uta, D.; Matsuda, K.I.; Kawata, M.; Carstens, E.; Sakamoto, T.; Sakamoto, H. Estrogens influence female itch sensitivity via the spinal gastrin-releasing peptide receptor neurons. Proc. Natl. Acad. Sci. USA 2021, 118. [Google Scholar] [CrossRef] [PubMed]
- Oti, T.; Satoh, K.; Uta, D.; Nagafuchi, J.; Tateishi, S.; Ueda, R.; Takanami, K.; Young, L.J.; Galione, A.; Morris, J.F.; et al. Oxytocin Influences Male Sexual Activity via Non-synaptic Axonal Release in the Spinal Cord. Curr. Biol. 2021, 31, 103–114 e105. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiyohara, K.; Uta, D.; Nagaoka, Y.; Kino, Y.; Nonaka, H.; Ninomiya-Baba, M.; Fujita, T. Involvement of Histamine H3 Receptor Agonism in Premature Ejaculation Found by Studies in Rats. Int. J. Mol. Sci. 2022, 23, 2291. https://doi.org/10.3390/ijms23042291
Kiyohara K, Uta D, Nagaoka Y, Kino Y, Nonaka H, Ninomiya-Baba M, Fujita T. Involvement of Histamine H3 Receptor Agonism in Premature Ejaculation Found by Studies in Rats. International Journal of Molecular Sciences. 2022; 23(4):2291. https://doi.org/10.3390/ijms23042291
Chicago/Turabian StyleKiyohara, Kazuhiro, Daisuke Uta, Yuuya Nagaoka, Yurika Kino, Hideki Nonaka, Midori Ninomiya-Baba, and Takuya Fujita. 2022. "Involvement of Histamine H3 Receptor Agonism in Premature Ejaculation Found by Studies in Rats" International Journal of Molecular Sciences 23, no. 4: 2291. https://doi.org/10.3390/ijms23042291
APA StyleKiyohara, K., Uta, D., Nagaoka, Y., Kino, Y., Nonaka, H., Ninomiya-Baba, M., & Fujita, T. (2022). Involvement of Histamine H3 Receptor Agonism in Premature Ejaculation Found by Studies in Rats. International Journal of Molecular Sciences, 23(4), 2291. https://doi.org/10.3390/ijms23042291