
The Potential to Fight Obesity with Adipogenesis Modulating Compounds


Abstract
:1. Introduction
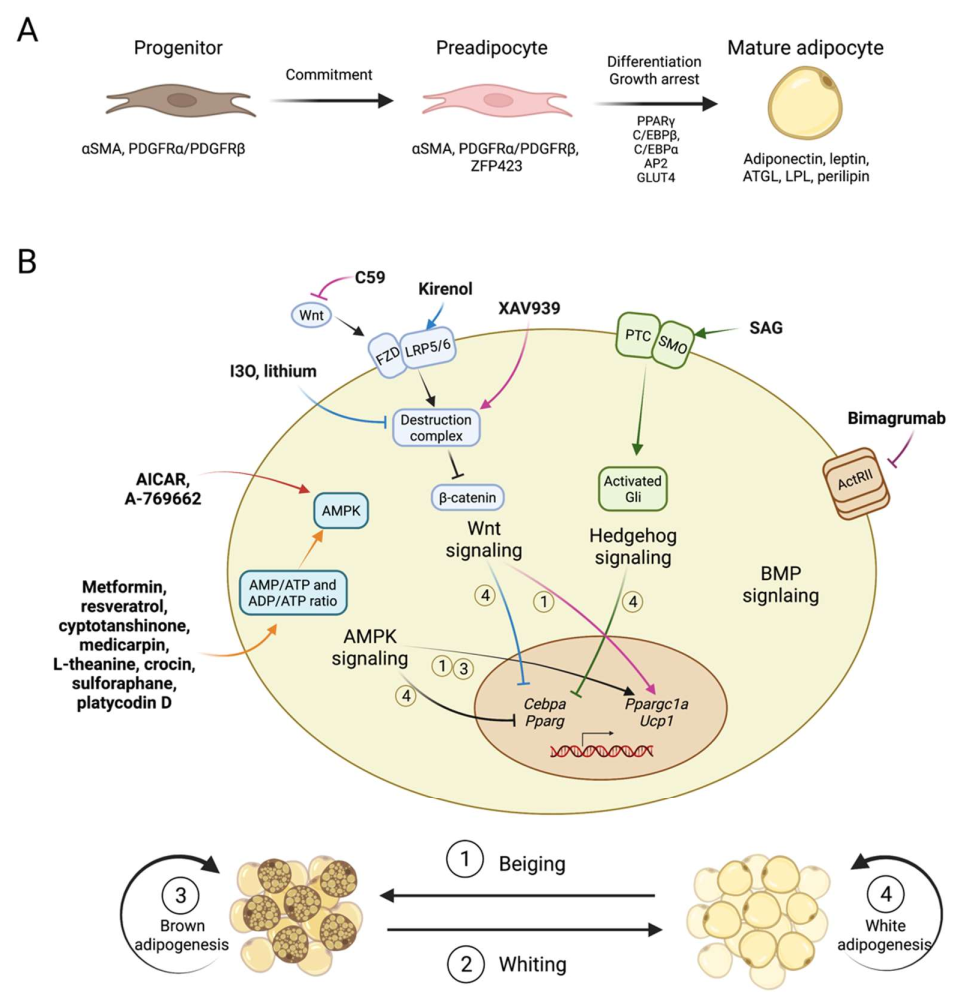
2. Mechanism of Adipogenesis
3. Compounds Modulating Adipogenic Signaling Pathways
3.1. AMPK Activators
3.2. Wnt Signaling Modulators
3.3. BMP Pathway Blockers
3.4. Hedgehog Pathways
3.5. Insulin and Other Pathways

{kind=link}
{kind=link}
| Compound | Structure | Pathways Involved | Experimental Conditions | Clinical Trials |
|---|---|---|---|---|
| AICAR |  | AMPK signaling pathway | In vitro: 1. Inhibit the differentiation of 3T3-L1 preadipocyte cell line [22,23]. 2. Promote brown adipogenesis in murine brown preadipocyte cell line [24]. In vivo: 1. Promote brown adipogenesis in murine WAT [24]. 2. Reduce adipose tissue content, improve glucose tolerance and insulin sensitivity in diet-induced DIO mice [23]. | NCT02322073, registered |
| A-769662 |  | AMPK signaling pathway | In vitro: 1. Inhibit the differentiation of 3T3-L1 preadipocyte cell line [26]. In vivo: 1. Promote brown adipogenesis in inguinal WAT, reduce body weight gain, improve glucose tolerance in HFD-fed mice [27]. | |
| Metformin |  | AMPK signaling pathway | In vivo: 1. Rescue the impaired brown adipogenesis in neonatal mice from obese mother through AMPK activation [28]. | NCT02274948, completed |
| Cyptotanshinone (from plant Salvia miltiorrhiza) |  | AMPK signaling pathway | In vitro: 1. Promote brown adipogenesis in C3H10T1/2 and 3T3-L1 cell line [35]. In vivo: 1. Reduce fat accumulation in ob/ob mice [36]. | |
| Medicarpin |  | AMPK signaling pathway | In vitro: 1. Promote brown adipogenesis in C3H10T1/2 mesenchymal stem cell [37]. | |
| L-Theanine (enriched in green tea) |  | AMPK signaling pathway | In vitro: 1. Promote brown adipogenesis of subcutaneous WAT in mice [38]. In vivo: 1. Improve glucose tolerance and insulin sensitivity, ameliorate obesity in HFD-fed mice [38]. | |
| Platycodin D (from plant Platycodon grandiflorum) |  | AMPK signaling pathway | In vitro: 1. Inhibit the differentiation of 3T3-L1 preadipocyte cell line [43]. In vivo: 1. Decrease adipogenic factors in WAT and promote thermogenic factors in BAT of db/db mice [42]. 2. Reduce body weight gain and white adipose tissue weight in db/db mice [42]. 3. Inhibit fat accumulation in HFD-induced obese mice [43]. | |
| Methyl cinnamate (from plant Zanthoxylum armatum) |  | AMPK signaling pathway | In vitro: 1. Inhibit the differentiation of 3T3-L1 preadipocyte cell line [45]. | |
| Sulforaphane (enriched in cruciferous vegetables like broccoli and cabbage) |  | AMPK signaling pathway | In vitro: 1. Inhibit the differentiation of 3T3-L1 preadipocyte cell line [40]. In vivo: 1. Reduce adipose mass gain and body weight gain in HFD-fed mice [41]. | NCT04364360, recruiting |
| Crocin (from plant saffron and gardenia) |  | AMPK signaling pathway | In vitro: 1. Inhibit the differentiation of 3T3-L1 preadipocyte cell line [39]. In vivo: 1. Inhibit adipose formation and reduce fat accumulation in db/db mice [39]. | |
| Lithium | Wnt signaling pathway | In vitro: 1. Inhibit the differentiation of 3T3-L1 preadipocyte cell line [47]. | ||
| Indirubin-3’-oxime (I3O) |  | Wnt signaling pathway | In vitro: 1. Inhibit the differentiation of 3T3-L1 preadipocyte cell line [55]. In vivo: 1. Inhibit obesity development in HFD-fed mice [55]. 2. Improving metabolic disorders like hyperlipidemia and hyperglycemia [55]. | |
| Kirenol |  | Wnt signaling pathway | In vitro: 1. Inhibit the differentiation of 3T3-L1 preadipocyte cell line [57]. | |
| 13m |  | Wnt signaling pathway | In vitro: 1. Inhibit adipogenesis in 3T3-L1 and C3H10T1/2 cell line [56]. | |
| C59 |  | Wnt signaling pathway | In vitro: 1. Promote brown adipogenesis in stromal vascular fraction (SVF) from mouse inguinal white, most likely targeting beige precursor cells [58,59]. | |
| XAV939 |  | Wnt signaling pathway | ||
| Hedgehog activator smoothened agonist (SAG) |  | Hedgehog signaling pathway | In vivo: 1. Block white but not brown adipocyte differentiation in mice [71]. | |
| Bimagrumab | BMP signaling pathway | In vivo: 1. Lead to significant loss of total body fat mass and metabolic improvement in overweight patients with type 2 diabetes [64]. | NCT03005288, completed |
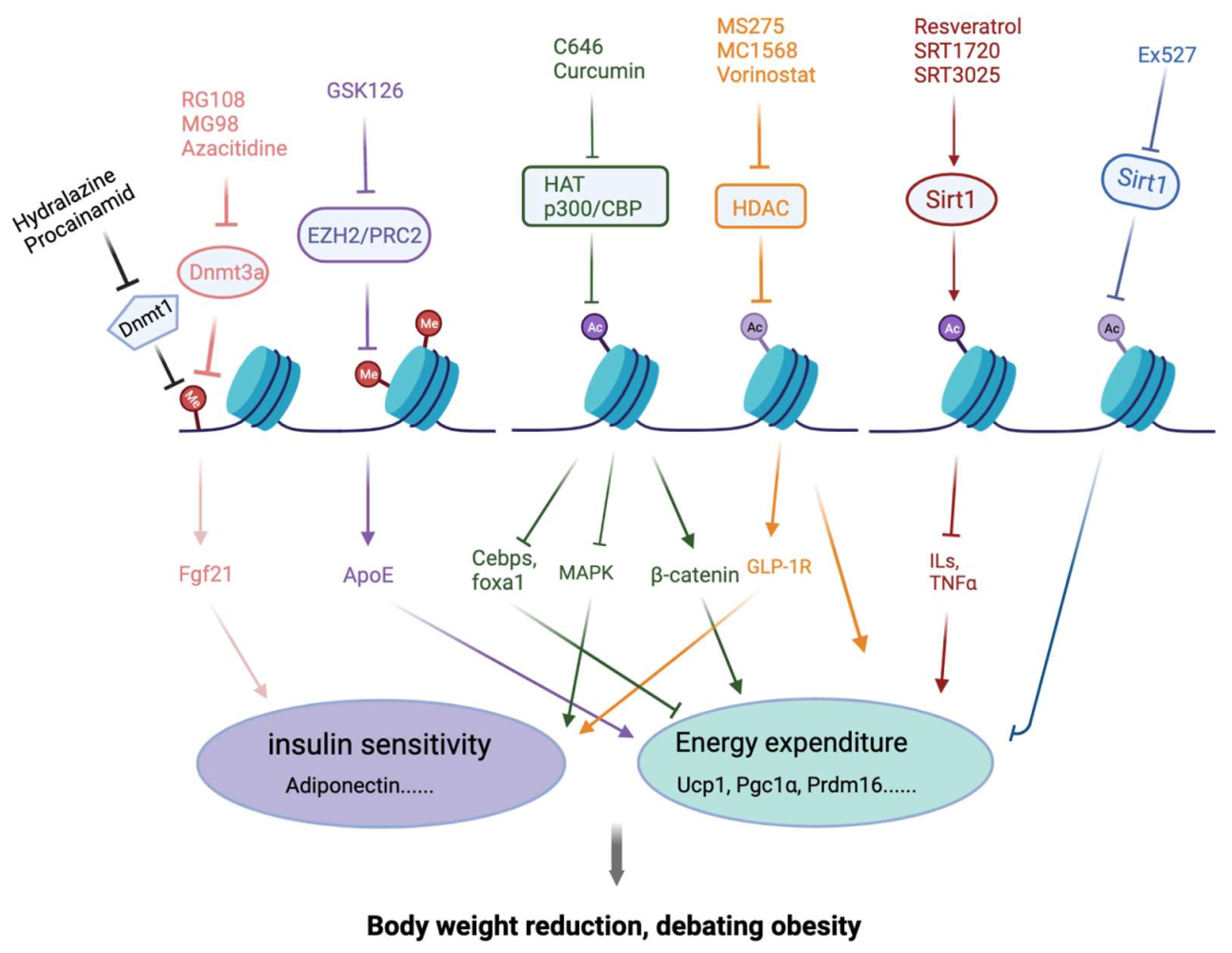
4. Epigenetic Modification Compounds Affecting Adipogenesis
4.1. Histone Acetylation Modulators
4.2. Histone Deacetylation Inhibitors
4.3. Sirtuin Activators
4.4. Histone Methylation Modulators
4.5. DNA Methylation Modulators
| Compound | Structure | Target | Experimental Conditions | NCT Numbers |
|---|---|---|---|---|
| Curcumin |  | HAT | In vitro: 1. induction of PPARγ [97,98] 2. anti-inflammation and antioxidant [99] 3. inhibits MAPK, activates the β-catenin and Wnt signaling [91,92,93] In vivo: 1. reduced epididymal adipose tissue and increased fatty acid β-oxidation [96] Clinical trials: 1. decreased serum triglyceride levels [100] | 1. NCT04723849, completed 2. NCT03864783, completed 3. NCT01975363, completed 4. NCT04315350, recruiting 5. NCT03542240, completed 6. NCT04595006, recruiting |
| C646 |  | p300/CBP | In vitro: 1. increase TIP60 and PCAF expression, promoting adipocyte diffentiation [103] 2. decrease CEBPB, CEBPD, FOXA1, and FOXA2, having an influence on energy expenditure [102] | |
| MS-275 |  | HDAC1–3 | In vitro: 1. enhances GLP-1R agonism to potentiate insulin secretion [110] 2. upregulation of Pgc1a and UCP1 expression [111] In vivo: 1. decreases body weight in DIO mice [110] | |
| MC1568 |  | HDAC4,5,7 and 9 | In vitro: 1. downregulation of Ucp1 [105] 2. attenuates the RAR- and PPARγ-mediated adipogenesis [112] | |
| Vorinostat |  | HDAC | In vitro: 1. less lipid accumulation and smaller lipid droplets [109] In vivo: 1. reduced body weight gain and increases the core temperature in lean and obese mice through Ucp1 upregulation [109] | |
| Resveratrol |  | SIRT1 activator | In vitro: 1. upregulation of Pgc1a, prdm16 [128] In vivo: 1. reduce adipocyte size in rhesus monkeys fed a high-fat [123] Clinical trials: 1. protection against obesity-related comorbid conditions [123] 2. decreased the size of abdominal subcutaneous adipocytes [117] 3. decrease in the production of IL-6 , IL-6/IL-10, and TNF-α in patients with high cardiovascular risk [124,125] 4. reduce body weight and fat mass [118] 5. improve insulin sensitivity [119] | 1. NCT02247596, completed 2. NCT04723849, completed 3. NCT01446276, completed 4. NCT01150955, completed 5. NCT01412645, completed 6. NCT00998504, completed 7. NCT01714102, completed 8. NCT01717820, completed 9. NCT02419092, Completed 10. NCT00823381, completed 11. NCT01302639, completed 12. NCT02216552, completed 13. NCT02381145, completed 14. NCT02633150, completed 15. NCT03448094, completed 16. NCT01518764, completed 17. NCT02114892, completed 18. NCT02767869, completed |
| SRT1720 |  | SIRT1 activator | ||
| SRT3025 |  | In vitro: 1. increased Foxc2, Pgc1α, Dio2, Tfam, and Cyc1 expression [129] | ||
| Ex-527 |  | SIRT1 inhibitor | In vitro: 1. down-regulated UCP1 in C3HT101/2 cells [129] | |
| GSK126 |  | EZH2/PRC2 | In vitro: 1. promote lipid accumulation via inducing ApoE expression [140] | |
| Hydralazine |  | DNMT1 | In vivo: 1. decrease body fat in animals and humans, increase lipolysis in abdominal subcutaneous adipose tissue [149] | |
| Procainamid |  | DNMT1 | It reduces the affinity of DNMT1 to bind DNA and SAM | |
| RG108 |  | DNMT1 | In vitro: 1. rescue Dnmt3a-mediated insulin resistance through Fgf21 upregulation with azacytidine [150] | |
| Azacitidine |  | DNMT1 | In vitro: 1. rescue Dnmt3a-mediated insulin resistance through Fgf21 upregulation with RG108 [150] |
5. Concluding Remarks and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Demeulemeester, F.; de Punder, K.; van Heijningen, M.; van Doesburg, F. Obesity as a Risk Factor for Severe COVID-19 and Complications: A Review. Cells 2021, 10, 933. [Google Scholar] [CrossRef] [PubMed]
- Ambele, M.A.; Dhanraj, P.; Giles, R.; Pepper, M.S. Adipogenesis: A Complex Interplay of Multiple Molecular Determinants and Pathways. Int. J. Mol. Sci. 2020, 21, 4283. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Seale, W.W.P. Control of brown and beige fat development. Nat. Rev. Mol. Cell Biol. 2016, 17, 691–702. [Google Scholar] [CrossRef]
- Satpathy, A.T.; Granja, J.M.; Yost, K.E.; Qi, Y.; Meschi, F.; McDermott, G.P.; Olsen, B.N.; Mumbach, M.R.; Pierce, S.E.; Corces, M.R.; et al. Massively parallel single-cell chromatin landscapes of human immune cell development and intratumoral T cell exhaustion. Nat. Biotechnol. 2019, 37, 925–936. [Google Scholar] [CrossRef] [PubMed]
- Vishvanath, L.; Gupta, R.K. Contribution of adipogenesis to healthy adipose tissue expansion in obesity. J. Clin. Investig. 2019, 129, 4022–4031. [Google Scholar] [CrossRef]
- Farmer, S.R. Transcriptional control of adipocyte formation. Cell Metab. 2006, 4, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Tao, C.; Jiang, L.; Shao, M.; Ye, R.; Zhu, Y.; Gordillo, R.; Ali, A.; Lian, Y.; Holland, W.L.; et al. Distinct regulatory mechanisms governing embryonic versus adult adipocyte maturation. Nat. Cell Biol. 2015, 17, 1099–1111. [Google Scholar] [CrossRef]
- Wang, W.; Kissig, M.; Rajakumari, S.; Huang, L.; Lim, H.-W.; Won, K.-J.; Seale, P. Ebf2 is a selective marker of brown and beige adipogenic precursor cells. Proc. Natl. Acad. Sci. USA 2014, 111, 14466–14471. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-E.; Schmidt, H.; Lai, B.; Ge, K. Transcriptional and Epigenomic Regulation of Adipogenesis. Mol. Cell. Biol. 2019, 39. [Google Scholar] [CrossRef] [Green Version]
- Green, H.; Meuth, M. An established pre-adipose cell line and its differentiation in culture. Cell 1974, 3, 127–133. [Google Scholar] [CrossRef]
- Müller, T.D.; Blüher, M.; Tschöp, M.H.; DiMarchi, R.D. Anti-obesity drug discovery: Advances and challenges. Nat. Rev. Drug Discov. 2021, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Cohen, P.; Kajimura, S. The cellular and functional complexity of thermogenic fat. Nat. Rev. Mol. Cell Biol. 2021, 22, 393–409. [Google Scholar] [CrossRef] [PubMed]
- Herzig, S.; Shaw, R.J. AMPK: Guardian of metabolism and mitochondrial homeostasis. Nat. Rev. Mol. Cell Biol. 2018, 19, 121–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woods, A.; Johnstone, S.R.; Dickerson, K.; Leiper, F.C.; Fryer, L.G.D.; Neumann, D.; Schlattner, U.; Wallimann, T.; Carlson, M.; Carling, D. LKB1 Is the Upstream Kinase in the AMP-Activated Protein Kinase Cascade. Curr. Biol. 2003, 13, 2004–2008. [Google Scholar] [CrossRef] [Green Version]
- Hurley, R.L.; Anderson, K.A.; Franzone, J.M.; Kemp, B.; Means, A.R.; Witters, L.A. The Ca2+/Calmodulin-dependent Protein Kinase Kinases Are AMP-activated Protein Kinase Kinases. J. Biol. Chem. 2005, 280, 29060–29066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardie, D.G. Regulation of AMP-activated protein kinase by natural and synthetic activators. Acta Pharm. Sin. B 2015, 6, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, B.; Serpell, C.J.; Fong, I.L.; Wong, E.H. Molecular Mechanisms of Adipogenesis: The Anti-adipogenic Role of AMP-Activated Protein Kinase. Front. Mol. Biosci. 2020, 7, 76. [Google Scholar] [CrossRef]
- Hardie, D.G.; Ross, F.A.; Hawley, S.A. AMPK: A nutrient and energy sensor that maintains energy homeostasis. Nat. Rev. Mol. Cell Biol. 2012, 13, 251–262. [Google Scholar] [CrossRef] [Green Version]
- Carling, D.; Zammit, V.A.; Hardie, G. A common bicyclic protein kinase cascade inactivates the regulatory enzymes of fatty acid and cholesterol biosynthesis. FEBS Lett. 1987, 223, 217–222. [Google Scholar] [CrossRef] [Green Version]
- Wu, N.; Zheng, B.; Shaywitz, A.; Dagon, Y.; Tower, C.; Bellinger, G.; Shen, C.-H.; Wen, J.; Asara, J.; McGraw, T.E.; et al. AMPK-Dependent Degradation of TXNIP upon Energy Stress Leads to Enhanced Glucose Uptake via GLUT1. Mol. Cell 2013, 49, 1167–1175. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, J.E.; Brocklehurst, K.J.; Marley, A.E.; Carey, F.; Carling, D.; Beri, R.K. Inhibition of lipolysis and lipogenesis in isolated rat adipocytes with AICAR, a cell-permeable activator of AMP-activated protein kinase. FEBS Lett. 1994, 353, 33–36. [Google Scholar] [CrossRef] [Green Version]
- Habinowski, S.A.; Witters, L.A. The Effects of AICAR on Adipocyte Differentiation of 3T3-L1 Cells. Biochem. Biophys. Res. Commun. 2001, 286, 852–856. [Google Scholar] [CrossRef]
- Giri, S.; Rattan, R.; Haq, E.; Khan, M.; Yasmin, R.; Won, J.-S.; Key, L.; Singh, A.K.; Singh, I. AICAR inhibits adipocyte differentiation in 3T3L1 and restores metabolic alterations in diet-induced obesity mice model. Nutr. Metab. 2006, 3, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vila-Bedmar, R.; Lorenzo, M.; Fernández-Veledo, S. Adenosine 5′-Monophosphate-Activated Protein Kinase-Mammalian Target of Rapamycin Cross Talk Regulates Brown Adipocyte Differentiation. Endocrinology 2010, 151, 980–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cool, B.; Zinker, B.; Chiou, W.; Kifle, L.; Cao, N.; Perham, M.; Dickinson, R.; Adler, A.; Gagne, G.; Iyengar, R.; et al. Identification and characterization of a small molecule AMPK activator that treats key components of type 2 diabetes and the metabolic syndrome. Cell Metab. 2006, 3, 403–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Wang, D.; Zhu, Q.; Gao, X.; Yang, S.; Xu, A.; Wu, D. Inhibitory Effects of A-769662, a Novel Activator of AMP-Activated Protein Kinase, on 3T3-L1 Adipogenesis. Biol. Pharm. Bull. 2009, 32, 993–998. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Zhang, L.; Li, B.; Jiang, H.; Duan, Y.; Xie, Z.; Shuai, L.; Li, J.; Li, J. AMP-Activated Protein Kinase (AMPK) Regulates Energy Metabolism through Modulating Thermogenesis in Adipose Tissue. Front. Physiol. 2018, 9, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Liang, X.; Sun, X.; Zhang, L.; Fu, X.; Rogers, C.J.; Berim, A.; Zhang, S.; Wang, S.; Wang, B.; et al. AMPK/α-Ketoglutarate Axis Dynamically Mediates DNA Demethylation in the Prdm16 Promoter and Brown Adipogenesis. Cell Metab. 2016, 24, 542–554. [Google Scholar] [CrossRef] [Green Version]
- Ning, H.-H.; Le, J.; Wang, Q.; Young, C.A.; Deng, B.; Gao, P.-X.; Zhang, H.-Q.; Qin, S.-L. The effects of metformin on simple obesity: A meta-analysis. Endocrine 2018, 62, 528–534. [Google Scholar] [CrossRef]
- Lentferink, Y.E.; Knibbe, C.A.J.; Van Der Vorst, M.M.J. Efficacy of Metformin Treatment with Respect to Weight Reduction in Children and Adults with Obesity: A Systematic Review. Drugs 2018, 78, 1887–1901. [Google Scholar] [CrossRef] [Green Version]
- Turner, R. Effect of intensive blood-glucose control with metformin on complications in overweight patients with type 2 diabetes (UKPDS 34). Lancet 1998, 352, 854–865. [Google Scholar] [CrossRef]
- Han, Y.; Xie, H.; Liu, Y.; Gao, P.; Yang, X.; Shen, Z. Effect of metformin on all-cause and cardiovascular mortality in patients with coronary artery diseases: A systematic review and an updated meta-analysis. Cardiovasc. Diabetol. 2019, 18, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pernicova, I.; Korbonits, M. Metformin—mode of action and clinical implications for diabetes and cancer. Nat. Rev. Endocrinol. 2014, 10, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Liang, X.; Yang, Q.; Fu, X.; Zhu, M.; Rodgers, B.D.; Jiang, Q.; Dodson, M.V.; Du, M. Resveratrol enhances brown adipocyte formation and function by activating AMP-activated protein kinase (AMPK) α1 in mice fed high-fat diet. Mol. Nutr. Food Res. 2016, 61, 1600746. [Google Scholar] [CrossRef] [Green Version]
- Imran, K.M.; Rahman, N.; Yoon, D.; Jeon, M.; Lee, B.-T.; Kim, Y.-S. Cryptotanshinone promotes commitment to the brown adipocyte lineage and mitochondrial biogenesis in C3H10T1/2 mesenchymal stem cells via AMPK and p38-MAPK signaling. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2017, 1862, 1110–1120. [Google Scholar] [CrossRef]
- Kim, E.J.; Jung, S.-N.; Son, K.H.; Kim, S.R.; Ha, T.Y.; Park, M.G.; Jo, I.G.; Park, J.G.; Choe, W.; Ha, J. Antidiabetes and Antiobesity Effect of Cryptotanshinone via Activation of AMP-Activated Protein Kinase. Mol. Pharmacol. 2007, 72, 62–72. [Google Scholar] [CrossRef] [Green Version]
- Imran, K.M.; Yoon, D.; Kim, Y.-S. A pivotal role of AMPK signaling in medicarpin-mediated formation of brown and beige. BioFactors 2017, 44, 168–179. [Google Scholar] [CrossRef]
- Peng, W.-Q.; Xiao, G.; Li, B.-Y.; Guo, Y.-Y.; Guo, L.; Tang, Q.-Q. l-Theanine Activates the Browning of White Adipose Tissue Through the AMPK/α-Ketoglutarate/Prdm16 Axis and Ameliorates Diet-Induced Obesity in Mice. Diabetes 2021, 70, 1458–1472. [Google Scholar] [CrossRef]
- Gu, M.; Luo, L.; Fang, K. Crocin inhibits obesity via AMPK-dependent inhibition of adipocyte differentiation and promotion of lipolysis. Biosci. Trends 2018, 12, 587–594. [Google Scholar] [CrossRef] [Green Version]
- Choi, K.-M.; Lee, Y.-S.; Sin, D.-M.; Lee, S.; Lee, M.K.; Lee, Y.-M.; Hong, J.-T.; Yun, Y.-P.; Yoo, H.-S. Sulforaphane Inhibits Mitotic Clonal Expansion During Adipogenesis Through Cell Cycle Arrest. Obesity 2012, 20, 1365–1371. [Google Scholar] [CrossRef]
- Choi, K.-M.; Lee, Y.-S.; Kim, W.; Kim, S.J.; Shin, K.-O.; Yu, J.-Y.; Lee, M.K.; Lee, Y.-M.; Hong, J.T.; Yun, Y.-P.; et al. Sulforaphane attenuates obesity by inhibiting adipogenesis and activating the AMPK pathway in obese mice. J. Nutr. Biochem. 2013, 25, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-L.; Park, J.; Jung, Y.; Ahn, K.S.; Um, J.-Y. Platycodin D, a novel activator of AMP-activated protein kinase, attenuates obesity in db/db mice via regulation of adipogenesis and thermogenesis. Phytomedicine 2018, 52, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.J.; Kang, M.; Kim, Y.S. Platycodin D Inhibits Lipogenesis through AMPKα-PPARγ2 in 3T3-L1 Cells and Modulates Fat Accumulation in Obese Mice. Planta Med. 2012, 78, 1536–1542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Ojeda, F.J.; Rupérez, A.I.; Gomez-Llorente, C.; Gil, A.; Aguilera, C.M. Cell Models and Their Application for Studying Adipogenic Differentiation in Relation to Obesity: A Review. Int. J. Mol. Sci. 2016, 17, 1040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.-Y.; Lee, M.-H.; Hsu, C.-C.; Wei, C.-L.; Tsai, Y.-C. Methyl Cinnamate Inhibits Adipocyte Differentiation via Activation of the CaMKK2–AMPK Pathway in 3T3-L1 Preadipocytes. J. Agric. Food Chem. 2012, 60, 955–963. [Google Scholar] [CrossRef]
- Nusse, R.; Clevers, H. Wnt/β-Catenin Signaling, Disease, and Emerging Therapeutic Modalities. Cell 2017, 169, 985–999. [Google Scholar] [CrossRef]
- Ross, S.E.; Hemati, N.; Longo, K.A.; Bennett, C.N.; Lucas, P.C.; Erickson, R.L.; MacDougald, O.A. Inhibition of Adipogenesis by Wnt Signaling. Science 2000, 289, 950–953. [Google Scholar] [CrossRef]
- Schunk, S.J.; Floege, J.; Fliser, D.; Speer, T. WNT–β-catenin signalling—A versatile player in kidney injury and repair. Nat. Rev. Nephrol. 2020, 17, 172–184. [Google Scholar] [CrossRef]
- Grainger, S.; Traver, D.; Willert, K. Wnt Signaling in Hematological Malignancies. Prog. Mol. Biol. Transl. Sci. 2017, 153, 321–341. [Google Scholar] [CrossRef]
- Gao, J.; Liao, Y.; Qiu, M.; Shen, W. Wnt/β-Catenin Signaling in Neural Stem Cell Homeostasis and Neurological Diseases. Neuroscientist 2020, 27, 58–72. [Google Scholar] [CrossRef]
- Jridi, I.; Canté-Barrett, K.; Pike-Overzet, K.; Staal, F.J.T. Inflammation and Wnt Signaling: Target for Immunomodulatory Therapy? Front. Cell Dev. Biol. 2021, 8. [Google Scholar] [CrossRef]
- Guo, Y.; Xiao, L.; Sun, L.; Liu, F. Wnt/β-Catenin Signaling: A Promising New Target for Fibrosis Diseases. Physiol. Res. 2012, 61, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Kahn, M. Can we safely target the WNT pathway? Nat. Rev. Drug Discov. 2014, 13, 513–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, P.S.; Melton, D.A. A molecular mechanism for the effect of lithium on development. Proc. Natl. Acad. Sci. USA 1996, 93, 8455–8459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, O.M.; Cho, Y.-H.; Choi, S.; Lee, S.-H.; Seo, S.H.; Kim, H.-Y.; Han, G.; Min, D.S.; Park, T.; Choi, K.Y. The small molecule indirubin-3′-oxime activates Wnt/β-catenin signaling and inhibits adipocyte differentiation and obesity. Int. J. Obes. 2013, 38, 1044–1052. [Google Scholar] [CrossRef] [Green Version]
- Rajan, S.; Puri, S.; Kumar, D.; Babu, M.H.; Shankar, K.; Varshney, S.; Srivastava, A.; Gupta, A.; Reddy, M.S.; Gaikwad, A.N. Novel indole and triazole based hybrid molecules exhibit potent anti-adipogenic and antidyslipidemic activity by activating Wnt3a/β-catenin pathway. Eur. J. Med. Chem. 2018, 143, 1345–1360. [Google Scholar] [CrossRef]
- Kim, M.-B.; Song, Y.; Kim, C.; Hwang, J.-K. Kirenol inhibits adipogenesis through activation of the Wnt/β-catenin signaling pathway in 3T3-L1 adipocytes. Biochem. Biophys. Res. Commun. 2014, 445, 433–438. [Google Scholar] [CrossRef]
- Kang, S.; Bajnok, L.; Longo, K.A.; Petersen, R.K.; Hansen, J.; Kristiansen, K.; MacDougald, O. Effects of Wnt Signaling on Brown Adipocyte Differentiation and Metabolism Mediated by PGC-1α. Mol. Cell. Biol. 2005, 25, 1272–1282. [Google Scholar] [CrossRef] [Green Version]
- Lo, K.A.; Ng, P.Y.; Kabiri, Z.; Virshup, D.M.; Sun, L. Wnt inhibition enhances browning of mouse primary white adipocytes. Adipocyte 2016, 5, 224–231. [Google Scholar] [CrossRef] [Green Version]
- Jia, S.; Meng, A. TGFβ family signaling and development. Development 2021, 148, dev188490. [Google Scholar] [CrossRef]
- Zaragosi, L.-E.; Wdziekonski, B.; Villageois, P.; Keophiphath, M.; Maumus, M.; Tchkonia, T.; Bourlier, V.; Mohsen-Kanson, T.; Ladoux, A.; Elabd, C.; et al. Activin A Plays a Critical Role in Proliferation and Differentiation of Human Adipose Progenitors. Diabetes 2010, 59, 2513–2521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Q.-Q.; Otto, T.C.; Lane, M.D. Commitment of C3H10T1/2 pluripotent stem cells to the adipocyte lineage. Proc. Natl. Acad. Sci. USA 2004, 101, 9607–9611. [Google Scholar] [CrossRef] [Green Version]
- Tseng, Y.-H.; Kokkotou, E.; Schulz, T.J.; Huang, T.L.; Winnay, J.N.; Taniguchi, C.M.; Tran, T.T.; Suzuki, R.; Espinoza, D.O.; Yamamoto, Y.; et al. New role of bone morphogenetic protein 7 in brown adipogenesis and energy expenditure. Nature 2008, 454, 1000–1004. [Google Scholar] [CrossRef]
- Lach-Trifilieff, E.; Minetti, G.C.; Sheppard, K.; Ibebunjo, C.; Feige, J.N.; Hartmann, S.; Brachat, S.; Rivet, H.; Koelbing, C.; Morvan, F.; et al. An Antibody Blocking Activin Type II Receptors Induces Strong Skeletal Muscle Hypertrophy and Protects from Atrophy. Mol. Cell. Biol. 2014, 34, 606–618. [Google Scholar] [CrossRef] [Green Version]
- Heymsfield, S.B.; Coleman, L.A.; Miller, R.; Rooks, D.S.; Laurent, D.; Petricoul, O.; Praestgaard, J.; Swan, T.; Wade, T.; Perry, R.G.; et al. Effect of Bimagrumab vs. Placebo on Body Fat Mass Among Adults with Type 2 Diabetes and Obesity A Phase 2 Randomized Clinical Trial. JAMA Netw. Open 2021, 4, e2033457. [Google Scholar] [CrossRef]
- Fournier, B.; Murray, B.; Gutzwiller, S.; Marcaletti, S.; Marcellin, D.; Bergling, S.; Brachat, S.; Persohn, E.; Pierrel, E.; Bombard, F.; et al. Blockade of the Activin Receptor IIB Activates Functional Brown Adipogenesis and Thermogenesis by Inducing Mitochondrial Oxidative Metabolism. Mol. Cell. Biol. 2012, 32, 2871–2879. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-J. Regulation of Muscle Mass by Myostatin. Annu. Rev. Cell Dev. Biol. 2004, 20, 61–86. [Google Scholar] [CrossRef]
- Nüsslein-Volhard, C.; Wieschaus, E. Mutations affecting segment number and polarity in Drosophila. Nature 1980, 287, 795–801. [Google Scholar] [CrossRef]
- Briscoe, J.; Therond, P.P. The mechanisms of Hedgehog signalling and its roles in development and disease. Nat. Rev. Mol. Cell Biol. 2013, 14, 416–429. [Google Scholar] [CrossRef]
- Shi, Y.; Long, F. Hedgehog signaling via Gli2 prevents obesity induced by high-fat diet in adult mice. eLife 2017, 6, e31649. [Google Scholar] [CrossRef] [Green Version]
- Pospisilik, J.A.; Schramek, D.; Schnidar, H.; Cronin, S.J.; Nehme, N.T.; Zhang, X.; Knauf, C.; Cani, P.D.; Aumayr, K.; Todoric, J.; et al. Drosophila Genome-wide Obesity Screen Reveals Hedgehog as a Determinant of Brown versus White Adipose Cell Fate. Cell 2010, 140, 148–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nosavanh, L.; Yu, D.-H.; Jaehnig, E.J.; Tong, Q.; Shen, L.; Chen, M.-H. Cell-autonomous activation of Hedgehog signaling inhibits brown adipose tissue development. Proc. Natl. Acad. Sci. USA 2015, 112, 5069–5074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, W.-K.; Meliton, V.; Amantea, C.M.; Hahn, T.J.; Parhami, F. 20(S)-Hydroxycholesterol Inhibits PPARγ Expression and Adipogenic Differentiation of Bone Marrow Stromal Cells Through a Hedgehog-Dependent Mechanism. J. Bone Miner. Res. 2007, 22, 1711–1719. [Google Scholar] [CrossRef] [PubMed]
- Spinella-Jaegle, S.; Rawadi, G.; Kawai, S.; Gallea, S.; Faucheu, C.; Mollat, P.; Courtois, B.; Bergaud, B.; Ramez, V.; Blanchet, A.M.; et al. Sonic hedgehog increases the commitment of pluripotent mesenchymal cells into the osteoblastic lineage and abolishes adipocytic differentiation. J. Cell Sci. 2001, 114, 2085–2094. [Google Scholar] [CrossRef] [PubMed]
- Justesen, J.; Stenderup, K.; Eriksen, E.; Kassem, M. Maintenance of Osteoblastic and Adipocytic Differentiation Potential with Age and Osteoporosis in Human Marrow Stromal Cell Cultures. Calcif. Tissue Res. 2002, 71, 36–44. [Google Scholar] [CrossRef]
- Miki, H.; Yamauchi, T.; Suzuki, R.; Komeda, K.; Tsuchida, A.; Kubota, N.; Terauchi, Y.; Kamon, J.; Kaburagi, Y.; Matsui, J.; et al. Essential role of insulin receptor substrate 1 (IRS-1) and IRS-2 in adipocyte differentiation. Mol. Cell. Biol. 2001, 21, 2521–2532. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.H.; Huang, J.; Düvel, K.; Boback, B.; Wu, S.; Squillace, R.M.; Wu, C.-L.; Manning, B.D. Insulin Stimulates Adipogenesis through the Akt-TSC2-mTORC1 Pathway. PLoS ONE 2009, 4, e6189. [Google Scholar] [CrossRef]
- Yeh, W.C.; Bierer, B.; McKnight, S.L. Rapamycin inhibits clonal expansion and adipogenic differentiation of 3T3-L1 cells. Proc. Natl. Acad. Sci. USA 1995, 92, 11086–11090. [Google Scholar] [CrossRef] [Green Version]
- Boucher, J.; Mori, M.; Lee, K.; Smyth, G.; Liew, C.W.; Macotela, Y.; Rourk, M.; Bluher, M.; Russell, S.J.; Kahn, C.R. Impaired thermogenesis and adipose tissue development in mice with fat-specific disruption of insulin and IGF-1 signalling. Nat. Commun. 2012, 3, 902. [Google Scholar] [CrossRef]
- Sakaue, H.; Konishi, M.; Ogawa, W.; Asaki, T.; Mori, T.; Yamasaki, M.; Takata, M.; Ueno, H.; Kato, S.; Kasuga, M.; et al. Requirement of fibroblast growth factor 10 in development of white adipose tissue. Genes Dev. 2002, 16, 908–912. [Google Scholar] [CrossRef] [Green Version]
- Le Blanc, S.; Simann, M.; Jakob, F.; Schütze, N.; Schilling, T. Fibroblast growth factors 1 and 2 inhibit adipogenesis of human bone marrow stromal cells in 3D collagen gels. Exp. Cell Res. 2015, 338, 136–148. [Google Scholar] [CrossRef] [PubMed]
- Hutley, L.; Shurety, W.; Newell, F.; McGeary, R.; Pelton, N.; Grant, J.; Herington, A.; Cameron, D.; Whitehead, J.; Prins, J. Fibroblast Growth Factor 1: A key regulator of human adipogenesis. Diabetes 2004, 53, 3097–3106. [Google Scholar] [CrossRef] [PubMed]
- Widberg, C.H.; Newell, F.S.; Bachmann, A.W.; Ramnoruth, S.N.; Spelta, M.C.; Whitehead, J.P.; Hutley, L.J.; Prins, J.B. Fibroblast growth factor receptor 1 is a key regulator of early adipogenic events in human preadipocytes. Am. J. Physiol. Metab. 2009, 296, E121–E131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bannister, A.J.; Kouzarides, T. Regulation of chromatin by histone modifications. Cell Res. 2011, 21, 381–395. [Google Scholar] [CrossRef]
- Maunakea, A.K.; Chepelev, I.; Zhao, K. Epigenome Mapping in Normal and Disease States. Circ. Res. 2010, 107, 327–339. [Google Scholar] [CrossRef] [Green Version]
- Galdieri, L.; Vancura, A. Acetyl-CoA Carboxylase Regulates Global Histone Acetylation. J. Biol. Chem. 2012, 287, 23865–23876. [Google Scholar] [CrossRef] [Green Version]
- Boone, A.; Brownsey, R.; Elliott, J.; Kulpa, J.; Lee, W. Regulation of acetyl-CoA carboxylase. Biochem. Soc. Trans. 2006, 34, 223–227. [Google Scholar] [CrossRef]
- Takahashi, N.; Kawada, T.; Yamamoto, T.; Goto, T.; Taimatsu, A.; Aoki, N.; Kawasaki, H.; Taira, K.; Yokoyama, K.K.; Kamei, Y.; et al. Overexpression and Ribozyme-mediated Targeting of Transcriptional Coactivators CREB-binding Protein and p300 Revealed Their Indispensable Roles in Adipocyte Differentiation through the Regulation of Peroxisome Proliferator-activated Receptor γ. J. Biol. Chem. 2002, 277, 16906–16912. [Google Scholar] [CrossRef] [Green Version]
- Yamauchi, T.; Oike, Y.; Kamon, J.; Waki, H.; Komeda, K.; Tsuchida, A.; Date, Y.; Li, M.-X.; Miki, H.; Akanuma, Y.; et al. Increased insulin sensitivity despite lipodystrophy in Crebbp heterozygous mice. Nat. Genet. 2002, 30, 221–226. [Google Scholar] [CrossRef]
- Zhang, D.-W.; Fu, M.; Gao, S.-H.; Liu, J.-L. Curcumin and Diabetes: A Systematic Review. Evid.-Based Complement. Altern. Med. 2013, 2013, 636053. [Google Scholar] [CrossRef]
- Ahn, J.; Lee, H.; Kim, S.; Ha, T. Curcumin-induced suppression of adipogenic differentiation is accompanied by activation of Wnt/β-catenin signaling. Am. J. Physiol. Physiol. 2010, 298, C1510–C1516. [Google Scholar] [CrossRef] [PubMed]
- Ninomiya-Tsuji, J.; Torti, F.M.; Ringold, G.M. Tumor necrosis factor-induced c-myc expression in the absence of mitogenesis is associated with inhibition of adipocyte differentiation. Proc. Natl. Acad. Sci. USA 1993, 90, 9611–9615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, M.; Rao, M.; Bouras, T.; Wang, C.; Wu, K.; Zhang, X.; Li, Z.; Yao, T.-P.; Pestell, R.G. Cyclin D1 Inhibits Peroxisome Proliferator-activated Receptor γ-mediated Adipogenesis through Histone Deacetylase Recruitment. J. Biol. Chem. 2005, 280, 16934–16941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asai, A.; Miyazawa, T. Dietary Curcuminoids Prevent High-Fat Diet–Induced Lipid Accumulation in Rat Liver and Epididymal Adipose Tissue. J. Nutr. 2001, 131, 2932–2935. [Google Scholar] [CrossRef] [Green Version]
- Shao, W.; Yu, Z.; Chiang, Y.; Yang, Y.; Chai, T.; Foltz, W.; Lu, H.; Fantus, I.G.; Jin, T. Curcumin Prevents High Fat Diet Induced Insulin Resistance and Obesity via Attenuating Lipogenesis in Liver and Inflammatory Pathway in Adipocytes. PLoS ONE 2012, 7, e28784. [Google Scholar] [CrossRef]
- Weisberg, S.P.; Leibel, R.; Tortoriello, D.V. Dietary Curcumin Significantly Improves Obesity-Associated Inflammation and Diabetes in Mouse Models of Diabesity. Endocrinology 2008, 149, 3549–3558. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, M.; Mimaki, Y.; Nishiyama, T.; Mae, T.; Kishida, H.; Tsukagawa, M.; Takahashi, K.; Kawada, T.; Nakagawa, K.; Kitahara, M. Hypoglycemic Effects of Turmeric (Curcuma longa L. Rhizomes) on Genetically Diabetic KK-Ay Mice. Biol. Pharm. Bull. 2005, 28, 937–939. [Google Scholar] [CrossRef] [Green Version]
- Nishiyama, T.; Mae, T.; Kishida, H.; Tsukagawa, M.; Mimaki, Y.; Kuroda, M.; Sashida, Y.; Takahashi, K.; Kawada, T.; Nakagawa, A.K.; et al. Curcuminoids and Sesquiterpenoids in Turmeric (Curcuma longa L.) Suppress an Increase in Blood Glucose Level in Type 2 Diabetic KK-Ay Mice. J. Agric. Food Chem. 2005, 53, 959–963. [Google Scholar] [CrossRef]
- Aggarwal, B.B. Targeting Inflammation-Induced Obesity and Metabolic Diseases by Curcumin and Other Nutraceuticals. Annu. Rev. Nutr. 2010, 30, 173–199. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi, A.; Sahebkar, A.; Iranshahi, M.; Amini, M.; Khojasteh, R.; Ghayour-Mobarhan, M.; Ferns, G.A. Effects of Supplementation with Curcuminoids on Dyslipidemia in Obese Patients: A Randomized Crossover Trial. Phytother. Res. 2012, 27, 374–379. [Google Scholar] [CrossRef]
- Bowers, E.M.; Yan, G.; Mukherjee, C.; Orry, A.; Wang, L.; Holbert, M.A.; Crump, N.T.; Hazzalin, C.A.; Liszczak, G.; Yuan, H.; et al. Virtual Ligand Screening of the p300/CBP Histone Acetyltransferase: Identification of a Selective Small Molecule Inhibitor. Chem. Biol. 2010, 17, 471–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimura, Y.; Sasagawa, S.; Ariyoshi, M.; Ichikawa, S.; Shimada, Y.; Kawaguchi, K.; Kawase, R.; Yamamoto, R.; Uehara, T.; Yanai, A.G.; et al. Systems pharmacology of adiposity reveals inhibition of EP300 as a common therapeutic mechanism of caloric restriction and resveratrol for obesity. Front. Pharmacol. 2015, 6, 199. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Wang, X.; Bi, M.; Hu, X.; Wang, Q.; Liang, H.; Liu, D. Effects of the histone acetylase inhibitor C646 on growth and differentiation of adipose-derived stem cells. Cell Cycle 2021, 20, 392–405. [Google Scholar] [CrossRef] [PubMed]
- Lasko, L.M.; Jakob, C.G.; Edalji, R.P.; Qiu, W.; Montgomery, D.; DiGiammarino, E.L.; Hansen, T.M.; Risi, R.M.; Frey, R.; Manaves, V.; et al. Discovery of a selective catalytic p300/CBP inhibitor that targets lineage-specific tumours. Nature 2017, 550, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Rajan, A.; Shi, H.; Xue, B. Class I and II Histone Deacetylase Inhibitors Differentially Regulate Thermogenic Gene Expression in Brown Adipocytes. Sci. Rep. 2018, 8, 13072. [Google Scholar] [CrossRef] [Green Version]
- Yoo, E.J.; Chung, J.-J.; Choe, S.S.; Kim, K.H.; Kim, J.B. Down-regulation of Histone Deacetylases Stimulates Adipocyte Differentiation. J. Biol. Chem. 2006, 281, 6608–6615. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Wu, R.; Cui, X.; Zha, L.; Yu, L.; Shi, H.; Xue, B. Histone Deacetylase 1 (HDAC1) Negatively Regulates Thermogenic Program in Brown Adipocytes via Coordinated Regulation of Histone H3 Lysine 27 (H3K27) Deacetylation and Methylation. J. Biol. Chem. 2016, 291, 4523–4536. [Google Scholar] [CrossRef] [Green Version]
- Dudakovic, A.; Camilleri, E.; Lewallen, E.; McGee-Lawrence, M.; Riester, S.M.; Kakar, S.; Montecino, M.; Stein, G.S.; Ryoo, H.-M.; Dietz, A.; et al. Histone Deacetylase Inhibition Destabilizes the Multi-Potent State of Uncommitted Adipose-Derived Mesenchymal Stromal Cells. J. Cell. Physiol. 2014, 230, 52–62. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Wang, Y.; Ding, J.; Zhang, S.; Yang, Y.; Sun, C. SAHA induces white fat browning and rectifies metabolic dysfunctions via activation of ZFPs. J. Endocrinol. 2021, 249, 177–193. [Google Scholar] [CrossRef]
- Bele, S.; Girada, S.B.; Ray, A.; Gupta, A.; Oruganti, S.; Babu, P.P.; Rayalla, R.S.; Kalivendi, S.V.; Ibrahim, A.; Puri, V.; et al. MS-275, a class 1 histone deacetylase inhibitor augments glucagon-like peptide-1 receptor agonism to improve glycemic control and reduce obesity in diet-induced obese mice. eLife 2020, 9, e52212. [Google Scholar] [CrossRef]
- Ferrari, A.; Fiorino, E.; Longo, R.; Barilla, S.; Mitro, N.; Cermenati, G.; Giudici, M.; Caruso, D.; Mai, A.; Guerrini, U.; et al. Attenuation of diet-induced obesity and induction of white fat browning with a chemical inhibitor of histone deacetylases. Int. J. Obes. 2016, 41, 289–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nebbioso, A.; Dell’Aversana, C.; Bugge, A.; Sarno, R.; Valente, S.; Rotili, D.; Manzo, F.; Teti, D.; Mandrup, S.; Ciana, P.; et al. HDACs class II-selective inhibition alters nuclear receptor-dependent differentiation. J. Mol. Endocrinol. 2010, 45, 219–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, J.; Ianni, A.; Smolka, C.; Vakhrusheva, O.; Nolte, H.; Krüger, M.; Wietelmann, A.; Simonet, N.; Adrian-Segarra, J.M.; Vaquero, A.; et al. Sirt7 promotes adipogenesis in the mouse by inhibiting autocatalytic activation of Sirt1. Proc. Natl. Acad. Sci. USA 2017, 114, E8352–E8361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanders, B.D.; Jackson, B.; Marmorstein, R. Structural basis for sirtuin function: What we know and what we don’t. Biochim. et Biophys. Acta (BBA)-Proteins Proteom. 2010, 1804, 1604–1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picard, F.; Kurtev, M.; Chung, N.; Topark-Ngarm, A.; Senawong, T.; de Oliveira, R.M.; Leid, M.; McBurney, M.W.; Guarente, L. Sirt1 promotes fat mobilization in white adipocytes by repressing PPAR-γ. Nature 2004, 429, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Mayoral, R.; Osborn, O.; McNelis, J.; Johnson, A.M.; Oh, D.Y.; Izquierdo, C.L.; Chung, H.; Li, P.; Traves, P.G.; Bandyopadhyay, G.; et al. Adipocyte SIRT1 knockout promotes PPARγ activity, adipogenesis and insulin sensitivity in chronic-HFD and obesity. Mol. Metab. 2015, 4, 378–391. [Google Scholar] [CrossRef]
- Jimenez-Gomez, Y.; Mattison, J.A.; Pearson, K.J.; Martin-Montalvo, A.; Palacios, H.H.; Sossong, A.M.; Ward, T.M.; Younts, C.M.; Lewis, K.; Allard, J.S.; et al. Resveratrol Improves Adipose Insulin Signaling and Reduces the Inflammatory Response in Adipose Tissue of Rhesus Monkeys on High-Fat, High-Sugar. Diet. Cell Metab. 2013, 18, 533–545. [Google Scholar] [CrossRef] [Green Version]
- Konings, E.; Timmers, S.; Boekschoten, M.V.; Goossens, G.H.; Jocken, J.W.; Afman, L.A.; Muller, M.; Schrauwen, P.; Mariman, E.C.; Blaak, E.E. The effects of 30 days resveratrol supplementation on adipose tissue morphology and gene expression patterns in obese men. Int. J. Obes. 2014, 38, 470–473. [Google Scholar] [CrossRef]
- Guo, R.; Liu, B.; Wang, K.; Zhou, S.; Li, W.; Xu, Y. Resveratrol ameliorates diabetic vascular inflammation and macrophage infiltration in db/db mice by inhibiting the NF-κB pathway. Diabetes Vasc. Dis. Res. 2014, 11, 92–102. [Google Scholar] [CrossRef]
- Méndez-del Villar, M.; González-Ortiz, M.; Martínez-Abundis, E.; Pérez-Rubio, K.G.; Lizárraga-Valdez, R. Effect of Resveratrol Administration on Metabolic Syndrome, Insulin Sensitivity, and Insulin Secretion. Metab. Syndr. Relat. Disord. 2014, 12, 497–501. [Google Scholar] [CrossRef]
- Dash, S.; Xiao, C.; Morgantini, C.; Szeto, L.; Lewis, G.F. High-Dose Resveratrol Treatment for 2 Weeks Inhibits Intestinal and Hepatic Lipoprotein Production in Overweight/Obese Men. Arter. Thromb. Vasc. Biol. 2013, 33, 2895–2901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gualdoni, G.A.; Kovarik, J.J.; Hofer, J.; Dose, F.; Pignitter, M.; Doberer, D.; Steinberger, P.; Somoza, V.; Wolzt, M.; Zlabinger, G.J. Resveratrol enhances TNF-α production in human monocytes upon bacterial stimulation. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2013, 1840, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity: Implications for metabolic syndrome, diabetes, hypertension, dyslipidemia, atherosclerosis, and cancer. Obes. Res. Clin. Pract. 2013, 7, e330–e341. [Google Scholar] [CrossRef]
- Tomé-Carneiro, J.; Larrosa, M.; Yáñez-Gascón, M.J.; Davalos, A.; Gil Zamorano, J.; Gonzálvez, M.; García-Almagro, F.J.; Ros, J.A.R.; Tomás-Barberán, F.A.; Espín, J.C.; et al. One-year supplementation with a grape extract containing resveratrol modulates inflammatory-related microRNAs and cytokines expression in peripheral blood mononuclear cells of type 2 diabetes and hypertensive patients with coronary artery disease. Pharmacol. Res. 2013, 72, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Tomé-Carneiro, J.; Gonzálvez, M.; Larrosa, M.; Yáñez-Gascón, M.J.; García-Almagro, F.J.; Ruiz-Ros, J.A.; Conesa, M.T.G.; Tomás-Barberán, F.A.; Espín, J.C. One-Year Consumption of a Grape Nutraceutical Containing Resveratrol Improves the Inflammatory and Fibrinolytic Status of Patients in Primary Prevention of Cardiovascular Disease. Am. J. Cardiol. 2012, 110, 356–363. [Google Scholar] [CrossRef]
- Price, N.L.; Gomes, A.P.; Ling, A.J.; Duarte, F.V.; Martin-Montalvo, A.; North, B.J.; Agarwal, B.; Ye, L.; Ramadori, G.; Teodoro, J.S.; et al. SIRT1 Is Required for AMPK Activation and the Beneficial Effects of Resveratrol on Mitochondrial Function. Cell Metab. 2012, 15, 675–690. [Google Scholar] [CrossRef] [Green Version]
- Rayalam, S.; Yang, J.-Y.; Ambati, S.; Della-Fera, M.A.; Baile, C.A. Resveratrol induces apoptosis and inhibits adipogenesis in 3T3-L1 adipocytes. Phytother. Res. 2008, 22, 1367–1371. [Google Scholar] [CrossRef]
- Lagouge, M.; Argmann, C.; Gerhart-Hines, Z.; Meziane, H.; Lerin, C.; Daussin, F.; Messadeq, N.; Milne, J.; Lambert, P.; Elliott, P.; et al. Resveratrol improves mitochondrial function and protects against metabolic disease by activating SIRT1 and PGC-1α. Cell 2006, 127, 1109–1122. [Google Scholar] [CrossRef]
- Artsi, H.; Gurt, I.; El-Haj, M.; Müller, R.; Kuhn, G.A.; Ben Shalom, G.; Cohen-Kfir, E.; Abramowitz, E.; Kandel, L.; Safran, O.; et al. Sirt1 Promotes a Thermogenic Gene Program in Bone Marrow Adipocytes: From Mice to (Wo)Men. Front. Endocrinol. 2019, 10, 126. [Google Scholar] [CrossRef] [Green Version]
- Floyd, Z.E.; Wang, Z.Q.; Kilroy, G.; Cefalu, W.T. Modulation of peroxisome proliferator–activated receptor γ stability and transcriptional activity in adipocytes by resveratrol. Metabolism 2008, 57, S32–S38. [Google Scholar] [CrossRef] [Green Version]
- Imamura, H.; Nagayama, D.; Ishihara, N.; Tanaka, S.; Watanabe, R.; Watanabe, Y.; Sato, Y.; Yamaguchi, T.; Ban, N.; Kawana, H.; et al. Resveratrol attenuates triglyceride accumulation associated with upregulation of Sirt1 and lipoprotein lipase in 3T3-L1 adipocytes. Mol. Genet. Metab. Rep. 2017, 12, 44–50. [Google Scholar] [CrossRef]
- Qiang, L.; Wang, L.; Kon, N.; Zhao, W.; Lee, S.; Zhang, Y.; Rosenbaum, M.; Zhao, Y.; Gu, W.; Farmer, S.R.; et al. Brown Remodeling of White Adipose Tissue by SirT1-Dependent Deacetylation of Ppary. Cell 2012, 150, 620–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ku, C.R.; Cho, Y.H.; Hong, Z.-Y.; Lee, H.; Lee, S.J.; Hong, S.-S.; Lee, E.J. The Effects of High Fat Diet and Resveratrol on Mitochondrial Activity of Brown Adipocytes. Endocrinol. Metab. 2016, 31, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Arias, N.; Pico, C.; Macarulla, M.T.; Oliver, P.; Miranda, J.; Palou, A.; Portillo, M.P. A combination of resveratrol and quercetin induces browning in white adipose tissue of rats fed an obesogenic diet. Obesity 2016, 25, 111–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Liang, X.; Yang, Q.; Fu, X.; Rogers, C.J.; Zhu, M.; Rodgers, B.D.; Jiang, Q.; Dodson, M.V.; Du, M. Resveratrol induces brown-like adipocyte formation in white fat through activation of AMP-activated protein kinase (AMPK) α1. Int. J. Obes. 2015, 39, 967–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alberdi, G.; Rodríguez, V.M.; Miranda, J.; Macarulla, M.T.; Churruca, I.; Portillo, M.P. Thermogenesis is involved in the body-fat lowering effects of resveratrol in rats. Food Chem. 2013, 141, 1530–1535. [Google Scholar] [CrossRef]
- Wang, L.; Jin, Q.; Lee, J.-E.; Su, I.-H.; Ge, K. Histone H3K27 methyltransferase Ezh2 represses Wnt genes to facilitate adipogenesis. Proc. Natl. Acad. Sci. USA 2010, 107, 7317–7322. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Wang, Y.; Wang, Y.; Wang, X.; Li, J.; Chang, K.; Sun, C.; Jia, Z.; Gao, S.; Wei, J.; et al. GSK126 alleviates the obesity phenotype by promoting the differentiation of thermogenic beige adipocytes in diet-induced obese mice. Biochem. Biophys. Res. Commun. 2018, 501, 9–15. [Google Scholar] [CrossRef]
- Wu, X.; Li, J.; Chang, K.; Yang, F.; Jia, Z.; Sun, C.; Li, Q.; Xu, Y. Histone H3 methyltransferase Ezh2 promotes white adipocytes but inhibits brown and beige adipocyte differentiation in mice. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2021, 1866, 158901. [Google Scholar] [CrossRef]
- Yiew, N.K.H.; Greenway, C.; Zarzour, A.; Ahmadieh, S.; Goo, B.; Kim, D.; Benson, T.W.; Ogbi, M.; Tang, Y.; Chen, W.; et al. Enhancer of zeste homolog 2 (EZH2) regulates adipocyte lipid metabolism independent of adipogenic differentiation: Role of apolipoprotein E. J. Biol. Chem. 2019, 294, 8577–8591. [Google Scholar] [CrossRef]
- Duteil, D.; Tosic, M.; Lausecker, F.; Nenseth, H.Z.; Müller, J.M.; Urban, S.; Willmann, D.; Petroll, K.; Messaddeq, N.; Arrigoni, L.; et al. Lsd1 Ablation Triggers Metabolic Reprogramming of Brown Adipose Tissue. Cell Rep. 2016, 17, 1008–1021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sambeat, A.; Gulyaeva, O.; Dempersmier, J.; Tharp, K.M.; Stahl, A.; Paul, S.M.; Sul, H.S. LSD1 Interacts with Zfp516 to Promote UCP1 Transcription and Brown Fat Program. Cell Rep. 2016, 15, 2536–2549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Kim, J.; Zhang, R.; Yang, X.; Zhang, Y.; Fang, J.; Chen, Z.; Teng, L.; Chen, X.; Ge, H.; et al. Histone Demethylase LSD1 Promotes Adipocyte Differentiation through Repressing Wnt Signaling. Cell Chem. Biol. 2016, 23, 1228–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.; Kang, S. Functional Implications of DNA Methylation in Adipose Biology. Diabetes 2019, 68, 871–878. [Google Scholar] [CrossRef] [Green Version]
- Lei, H.; Oh, S.; Okano, M.; Juttermann, R.; Goss, K.; Jaenisch, R.; Li, E. De novo DNA cytosine methyltransferase activities in mouse embryonic stem cells. Development 1996, 122, 3195–3205. [Google Scholar] [CrossRef]
- Dubois-Chevalier, J.; Oger, F.; Dehondt, H.; Firmin, F.F.; Gheeraert, C.; Staels, B.; Lefebvre, P.; Eeckhoute, J. A dynamic CTCF chromatin binding landscape promotes DNA hydroxymethylation and transcriptional induction of adipocyte differentiation. Nucleic Acids Res. 2014, 42, 10943–10959. [Google Scholar] [CrossRef]
- Qian, H.; Zhao, J.; Yang, X.; Wu, S.; An, Y.; Qu, Y.; Li, Z.; Ge, H.; Li, E.; Qi, W. TET1 promotes RXRα expression and adipogenesis through DNA demethylation. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2021, 1866, 158919. [Google Scholar] [CrossRef]
- Singh, N.; Dueñas-González, A.; Lyko, F.; Medina-Franco, J.L. Molecular Modeling and Molecular Dynamics Studies of Hydralazine with Human DNA Methyltransferase 1. ChemMedChem 2009, 4, 792–799. [Google Scholar] [CrossRef]
- Boon, N.; Goossens, G.; Blaak, E.E.; Saris, W.H. The effects of hydralazine on lipolysis in subcutaneous adipose tissue in humans. Metabolism 2007, 56, 1742–1748. [Google Scholar] [CrossRef]
- Lee, B.H.; Yegnasubramanian, S.; Lin, X.; Nelson, W.G. Procainamide Is a Specific Inhibitor of DNA Methyltransferase 1. J. Biol. Chem. 2005, 280, 40749–40756. [Google Scholar] [CrossRef] [Green Version]
- Halby, L.; Champion, C.; Sénamaud-Beaufort, C.; Ajjan, S.; Drujon, T.; Rajavelu, A.; Ceccaldi, A.; Jurkowska, R.; Lequin, O.; Nelson, W.G.; et al. Rapid Synthesis of New DNMT Inhibitors Derivatives of Procainamide. ChemBioChem 2011, 13, 157–165. [Google Scholar] [CrossRef] [PubMed]
- You, D.; Nilsson, E.; Tenen, D.E.; Lyubetskaya, A.; Lo, J.C.; Jiang, R.; Deng, J.; Dawes, B.A.; Vaag, A.; Ling, C.; et al. Dnmt3a is an epigenetic mediator of adipose insulin resistance. eLife 2017, 6, e30766. [Google Scholar] [CrossRef] [PubMed]
- Villivalam, S.D.; You, D.; Kim, J.; Lim, H.W.; Xiao, H.; Zushin, P.-J.H.; Oguri, Y.; Amin, P.; Kang, S. TET1 is a beige adipocyte-selective epigenetic suppressor of thermogenesis. Nat. Commun. 2020, 11, 4313. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Jing, J.; Movahed, M.; Cui, X.; Cao, Q.; Wu, R.; Chen, Z.; Yu, L.; Pan, Y.; Shi, H.; et al. Epigenetic interaction between UTX and DNMT1 regulates diet-induced myogenic remodeling in brown fat. Nat. Commun. 2021, 12, 6838. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, J.; Zhou, A.; Qi, W. The Potential to Fight Obesity with Adipogenesis Modulating Compounds. Int. J. Mol. Sci. 2022, 23, 2299. https://doi.org/10.3390/ijms23042299
Zhao J, Zhou A, Qi W. The Potential to Fight Obesity with Adipogenesis Modulating Compounds. International Journal of Molecular Sciences. 2022; 23(4):2299. https://doi.org/10.3390/ijms23042299
Chicago/Turabian StyleZhao, Jiaqi, Ailin Zhou, and Wei Qi. 2022. "The Potential to Fight Obesity with Adipogenesis Modulating Compounds" International Journal of Molecular Sciences 23, no. 4: 2299. https://doi.org/10.3390/ijms23042299
APA StyleZhao, J., Zhou, A., & Qi, W. (2022). The Potential to Fight Obesity with Adipogenesis Modulating Compounds. International Journal of Molecular Sciences, 23(4), 2299. https://doi.org/10.3390/ijms23042299