
Mechanistic Insights into Stereospecific Antifungal Activity of Chiral Fungicide Prothioconazole against Fusarium oxysporum F. sp. cubense

 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
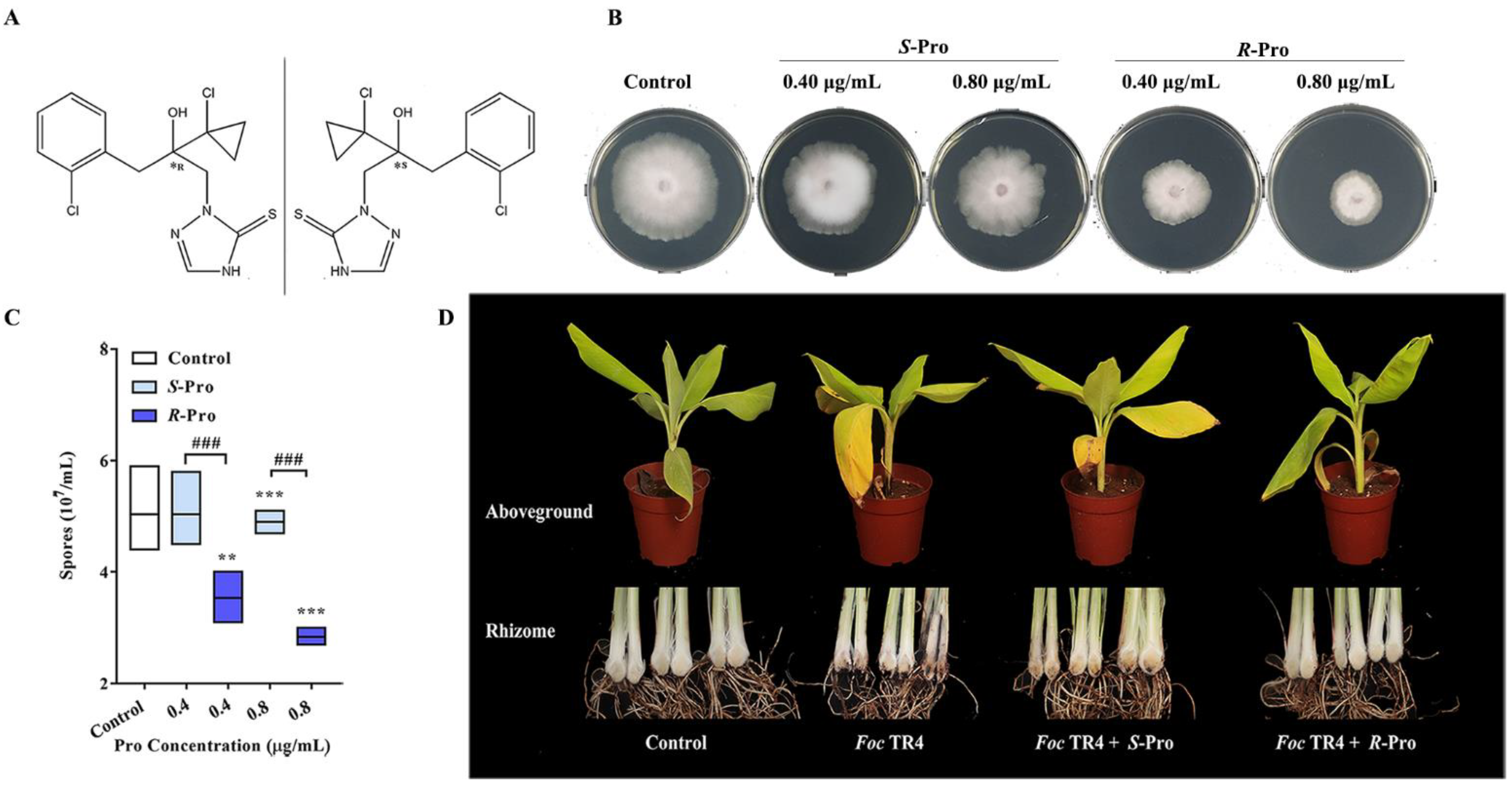
2.1. Pro Stereoselectively Inhibits TR4 Growth and Impairs Its Pathogenicity
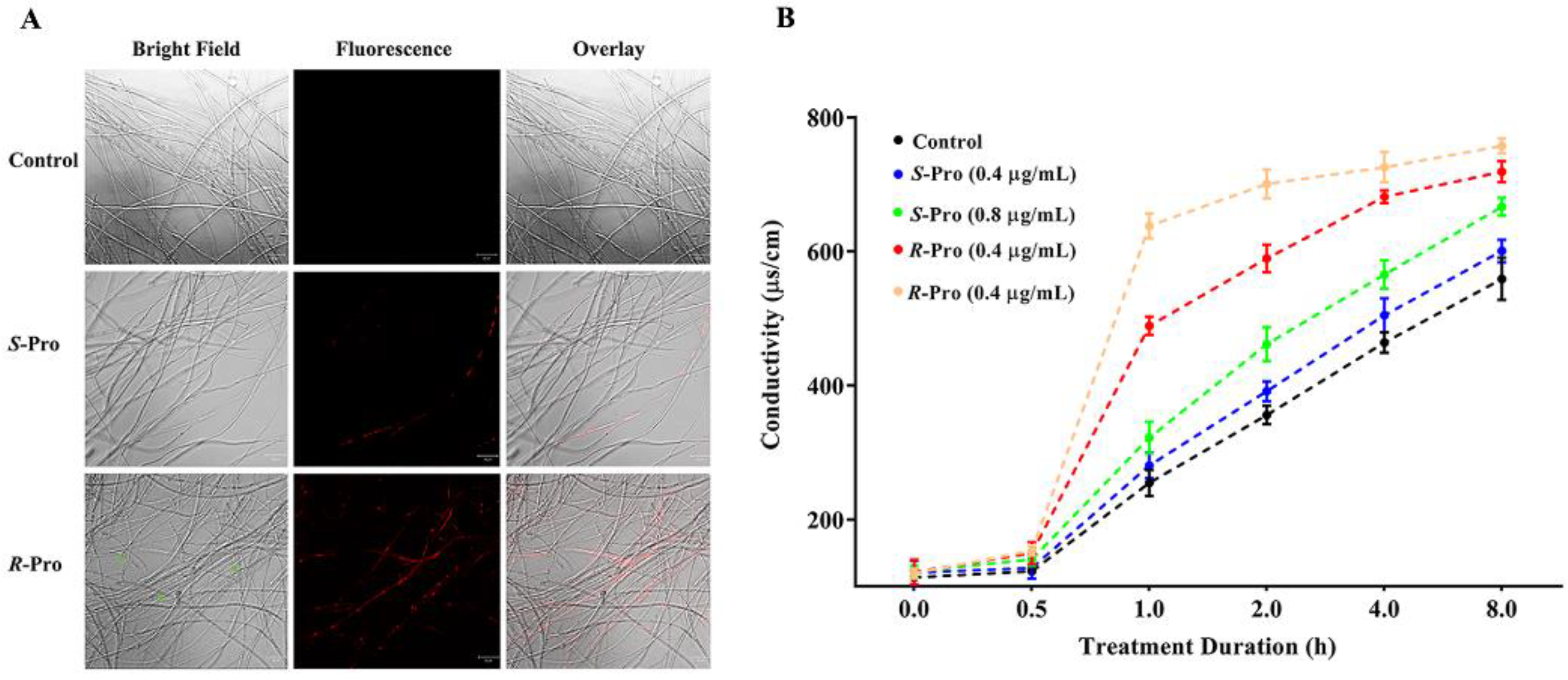
2.2. Pro Stereoselectively Leads to Morphological Changes and Impairment in Membrane Integrity in TR4
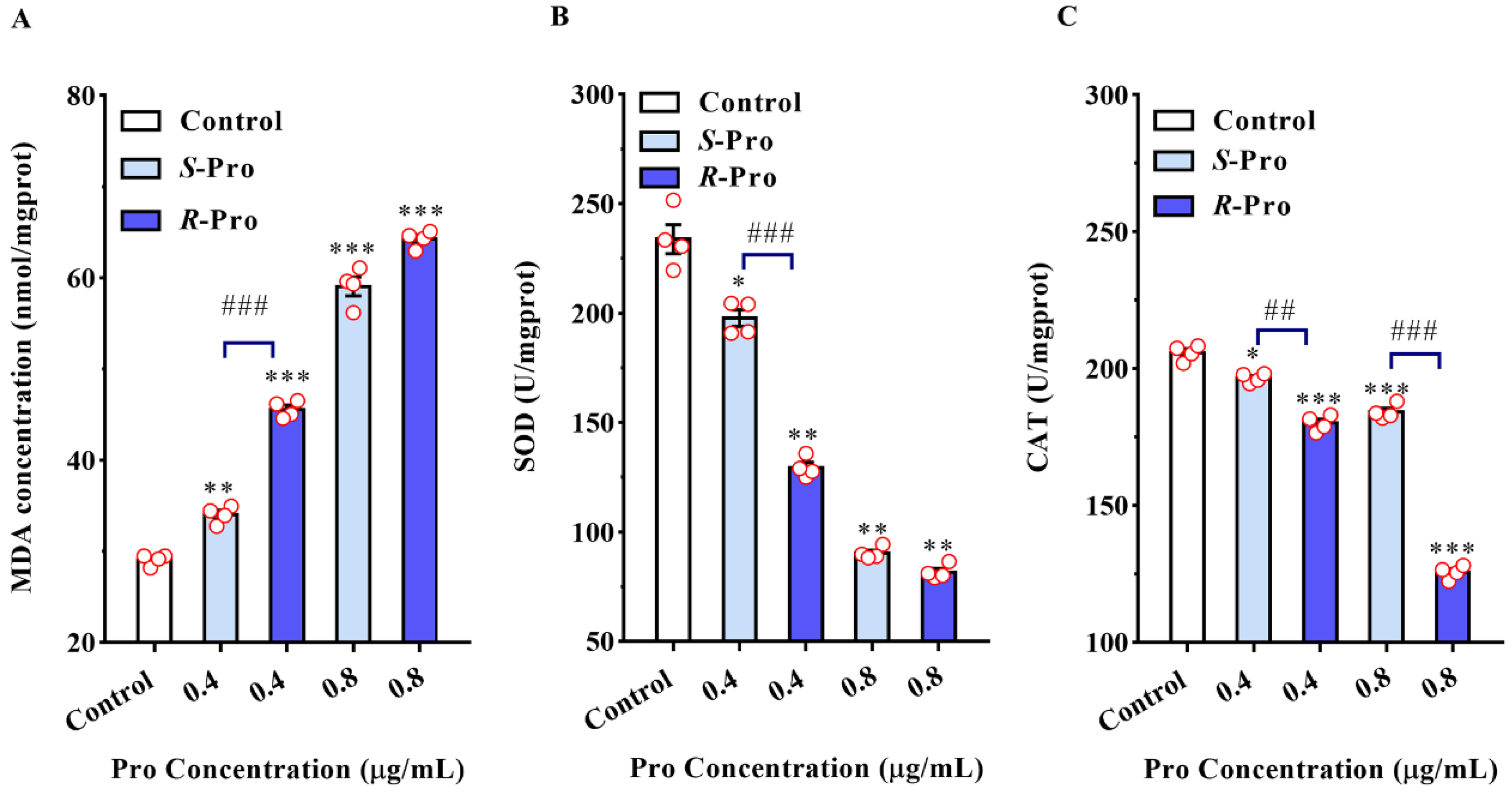
2.3. Pro Stereoselectively Triggers Oxidative Stress Reactions in TR4
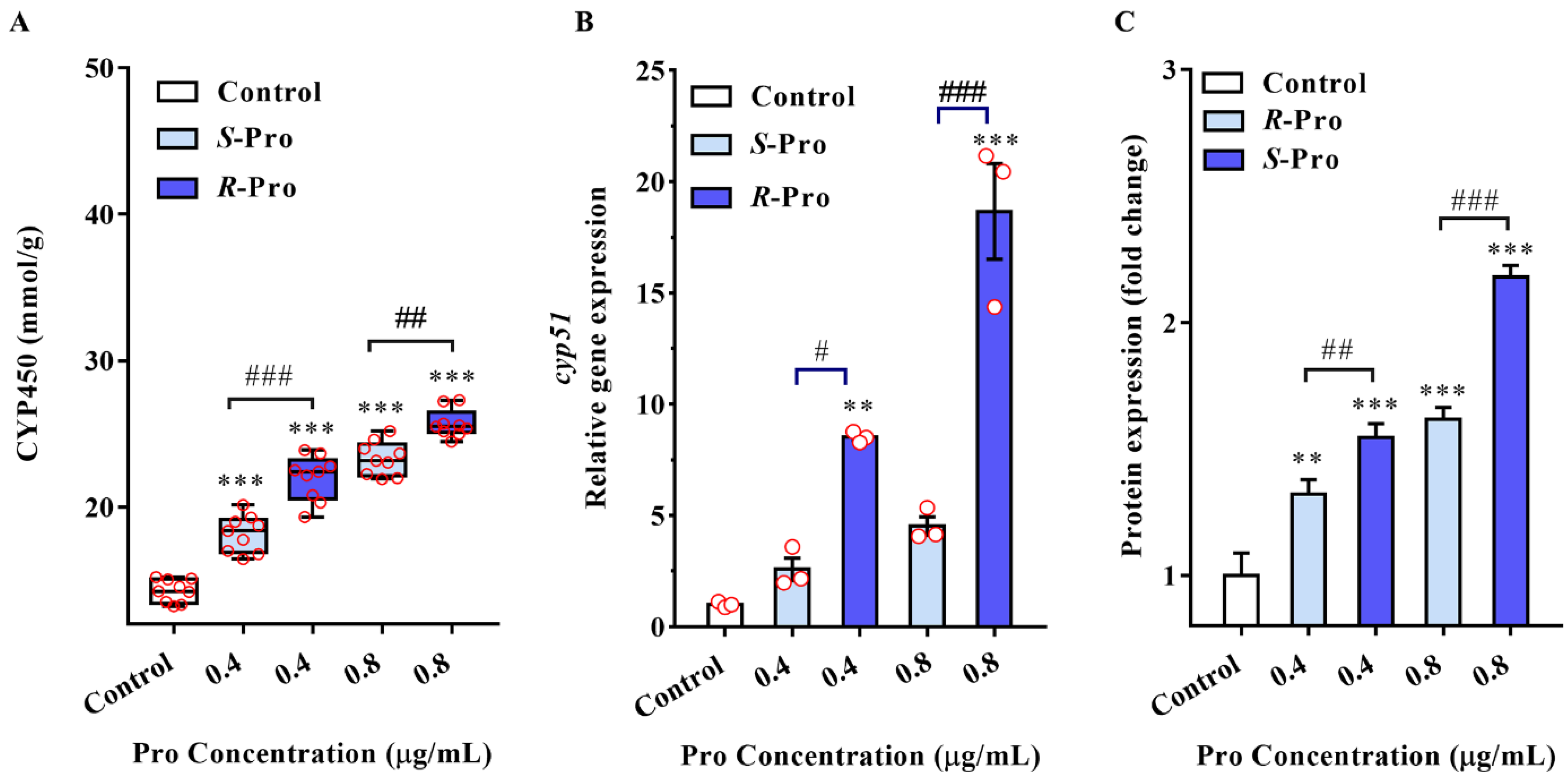
2.4. Pro Stereoselectively Upregulates the Expression of CYP51 in TR4
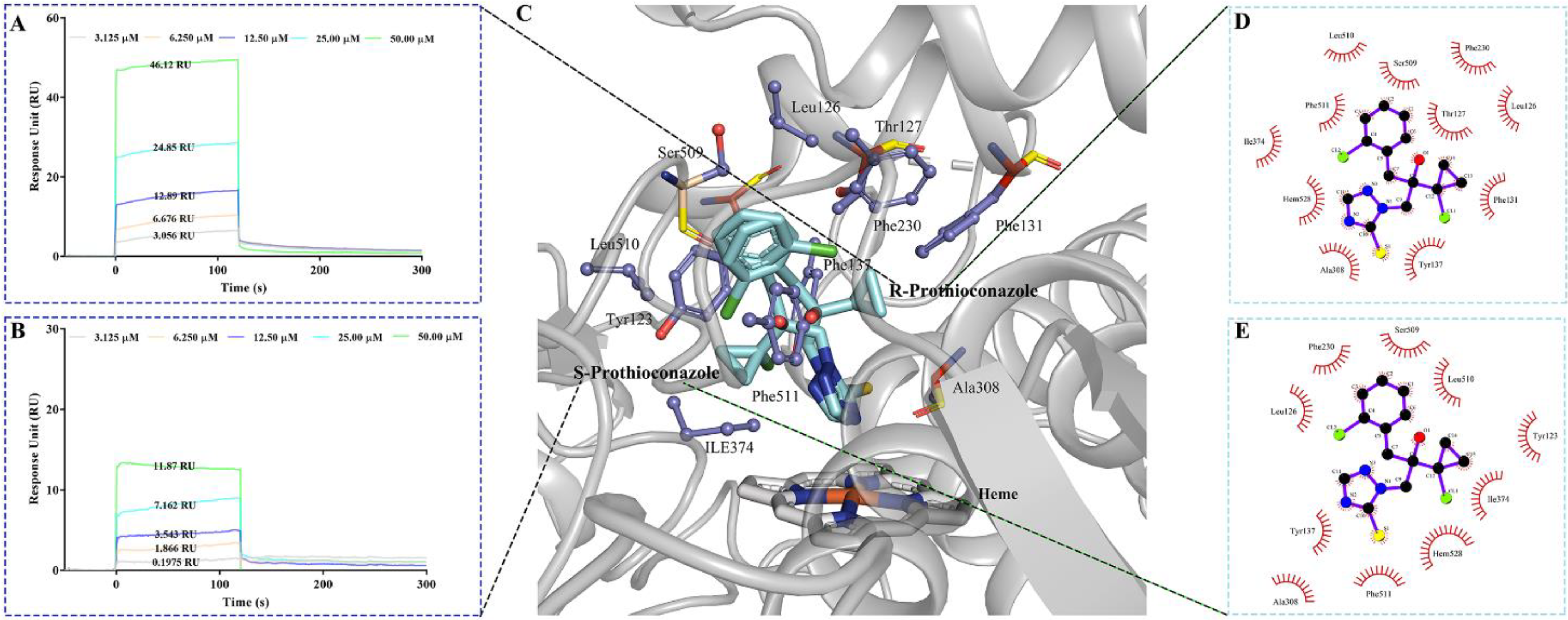
2.5. The Molecular Stereoselective Interactions between the Pro Enantiomers and CYP51 of TR4
3. Materials and Method
3.1. Chemicals and Reagents
3.2. Fungal Strains Culture Conditions and Experimental Design
3.3. Stereoselective Fungicidal Activity of Pro Enantiomers against TR4
3.4. Morphology and Propidium Iodide (PI) Influx of TR4
3.5. Cell Membrane Permeability of TR4
3.6. Determination of Cellular Enzymatic Activity
3.7. Real-Time PCR
3.8. Western Blot Analysis
3.9. Direct Binding Assays between CYP51 and Pro Enantiomers
3.10. Homology Modeling and Molecular Docking
3.11. Data Processing
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fernandez, D.; Assigbese, K.; Dubois, M.P.; Geiger, J.P. Molecular characterization of races and vegetative compatibility groups in Fusarium oxysporum f. sp. vasinfectum. Appl. Environ. Microbiol. 1994, 60, 4039–4046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nel, B.; Steinberg, C.; Labuschagne, N.; Viljoen, A. Isolation and characterization of nonpathogenic Fusarium oxysporum isolates from the rhizosphere of healthy banana plants. Plant Pathol. 2006, 55, 207–216. [Google Scholar] [CrossRef]
- Van den Berg, N.; Berger, D.K.; Hein, I.; Birch, P.R.J.; Wingfield, M.J.; Viljoen, A. Tolerance in banana to Fusarium wilt is associated with early up-regulation of cell wall-strengthening genes in the roots. Mol. Plant Pathol. 2007, 8, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Zorrilla-Fontanesi, Y.; Pauwels, L.; Panis, B.; Signorelli, S.; Vanderschuren, H.; Swennen, R. Strategies to revise agrosystems and breeding to control Fusarium wilt of banana. Nat. Food 2020, 1, 599–604. [Google Scholar] [CrossRef]
- Ploetz, R.C. Fusarium wilt of banana is caused by several pathogens referred to as Fusarium oxysporum f. sp. cubense. Phytopathology 2006, 96, 648. [Google Scholar] [CrossRef] [Green Version]
- Galvis, S. Colombia confirms that dreaded fungus has hit its banana plantations. Science 2019, 1–5. [Google Scholar] [CrossRef]
- Ghag, S.B.; Shekhawat, U.K.S.; Ganapathi, T.R. Fusarium wilt of banana: Biology, epidemiology and management. Int. J. Pest Manag. 2015, 61, 250–263. [Google Scholar] [CrossRef]
- Thangavelu, R. Fusarium wilt: A threat to banana cultivation and its management. CAB Rev. Perspect. Agric. Veter.-Sci. Nutr. Nat. Resour. 2020, 15. [Google Scholar] [CrossRef]
- Meldrum, R.; Daly, A.; Tran-Nguyen, L.; Aitken, E. The effect of surface sterilants on spore germination of Fusarium oxysporum f. sp. cubense tropical race 4. Crop Prot. 2013, 54, 194–198. [Google Scholar] [CrossRef]
- Nel, B.; Steinberg, C.; Labuschagne, N.; Viljoen, A. Evaluation of fungicides and sterilants for potential application in the management of Fusarium wilt of banana. Crop Prot. 2007, 26, 697–705. [Google Scholar] [CrossRef] [Green Version]
- Ying, Z.; Ling, L.; Kunde, L.; Xinping, Z.; Weiping, L. Enantiomer separation of triazole fungicides by high-performance liquid chromatography. Chirality 2008, 21, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Dong, F.; Li, J.; Chankvetadze, B.; Cheng, Y.; Xu, J.; Liu, X.; Li, Y.; Chen, X.; Bertucci, C.; Tedesco, D.; et al. chiral triazole fungicide difenoconazole: Absolute stereochemistry, stereoselective bioactivity, aquatic toxicity, and environmental behavior in vegetables and soil. Environ. Sci. Technol. 2013, 47, 3386–3394. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Zhao, M.; Niu, L.; Liu, W. Enantioselective Environmental Toxicology of Chiral Pesticides. Chem. Res. Toxicol. 2015, 28, 325–338. [Google Scholar] [CrossRef] [PubMed]
- Miller, N.F.; Standish, J.R.; Quesada-Ocampo, L. Sensitivity of Fusarium oxysporum f. sp. niveum to prothioconazole and pydiflumetofen in vitro and efficacy for Fusarium wilt management in watermelon. Plant Health Prog. 2020, 21, 13–18. [Google Scholar] [CrossRef]
- Limay-Rios, V.; Schaafsma, A.W. Effect of prothioconazole application timing on fusarium mycotoxin content in maize grain. J. Agric. Food Chem. 2018, 66, 4809–4819. [Google Scholar] [CrossRef]
- Zhang, J.; Li, L.; Lv, Q.; Yan, L.; Wang, Y.; Jiang, Y. The fungal CYP51s: Their functions, structures, related drug resistance, and inhibitors. Front. Microbiol. 2019, 10, 691. [Google Scholar] [CrossRef]
- Muellender, M.M.; Mahlein, A.-K.; Stammler, G.; Varrelmann, M. Evidence for the association of target-site resistance in cyp51 with reduced DMI sensitivity in European Cercospora beticola field isolates. Pest Manag. Sci. 2020, 77, 1765–1774. [Google Scholar] [CrossRef]
- Griffiths, K.M.; Bacic, A.; Howlett, B.J. Sterol composition of mycelia of the plant pathogenic ascomycete Leptosphaeria mac-ulans. Phytochemistry 2003, 62, 147–153. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, Z.; Tang, B.; Gao, B.; Tian, M.; Sanganyado, E.; Shi, H.; Wang, M. Mechanistic insights into stereospecific bioactivity and dissipation of chiral fungicide triticonazole in agricultural management. J. Agric. Food Chem. 2018, 66, 7286–7293. [Google Scholar] [CrossRef]
- Wang, X.; Liu, Y.; Xue, M.; Wang, Z.; Yu, J.; Guo, X. Enantioselective degradation of chiral fungicides triticonazole and prothioconazole in soils and their enantioselective accumulation in earthworms Eisenia fetida. Ecotoxicol. Environ. Saf. 2019, 183, 109491. [Google Scholar] [CrossRef]
- Zhang, Z.; Gao, B.; He, Z.; Li, L.; Zhang, Q.; Kaziem, A.E.; Wang, M. Stereoselective bioactivity of the chiral triazole fungicide prothioconazole and its metabolite. Pestic. Biochem. Physiol. 2019, 160, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Roldán, M.C.; Köhli, M.; Roncero, M.I.G.; Philippsen, P.; Di Pietro, A.; Espeso, E.A. Nuclear dynamics during germination, conidiation, and hyphal fusion of Fusarium oxysporum. Eukaryot. Cell 2010, 9, 1216–1224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaziem, A.E.; Gao, B.; Li, L.; Zhang, Z.; He, Z.; Wen, Y.; Wang, M.-H. Enantioselective bioactivity, toxicity, and degradation in different environmental mediums of chiral fungicide epoxiconazole. J. Hazard. Mater. 2019, 386, 121951. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Hua, X.-D.; Shi, H.-Y.; Liu, J.-S.; Tian, M.-M.; Wang, M.-H. Enantioselective bioactivity, acute toxicity and dissipation in vegetables of the chiral triazole fungicide flutriafol. J. Hazard. Mater. 2015, 284, 65–72. [Google Scholar] [CrossRef]
- Kathiravan, M.K.; Salake, A.B.; Chothe, A.S.; Dudhe, P.B.; Watode, R.P.; Mukta, M.S.; Gadhwe, S. The biology and chemistry of antifungal agents: A review. Bioorg. Med. Chem. 2012, 20, 5678–5698. [Google Scholar] [CrossRef]
- Dichtl, K.; Samantaray, S.; Aimanianda, V.; Zhu, Z.; Prévost, M.-C.; Latgé, J.-P.; Ebel, F.; Wagener, J. Aspergillus fumigatus devoid of cell wall β-1,3-glucan is viable, massively sheds galactomannan and is killed by septum formation inhibitors. Mol. Microbiol. 2014, 95, 458–471. [Google Scholar] [CrossRef]
- Geißel, B.; Loiko, V.; Klugherz, I.; Zhu, Z.; Wagener, N.; Kurzai, O.; Hondel, C.A.M.J.J.V.D.; Wagener, J. Azole-induced cell wall carbohydrate patches kill Aspergillus fumigatus. Nat. Commun. 2018, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Campos, P.S.; Quartin, V.N.; Ramalho, J.C.; Nunes, M.A. Electrolyte leakage and lipid degradation account for cold sensi-tivity in leaves of Coffea sp. plants. J. Plant Physiol. 2003, 160, 283–292. [Google Scholar] [CrossRef] [Green Version]
- Stenzel, K.; Vors, J. Sterol biosynthesis inhibitors. In Modern Crop Protection Compounds; Jeschke, P., Witschel, M., Krämer, W., Schirmer, U., Eds.; Wiley-VCH: Weinheim, Germany, 2019; Volume 3, Chapter 19; pp. 797–844. [Google Scholar]
- Burckhardt, I.C.; Gozal, D.; Dayyat, E.; Cheng, Y.; Li, R.C.; Goldbart, A.D.; Row, B.W. Green tea catechin polyphenols at-tenuate behavioral and oxidative responses to intermittent hypoxia. Am. J. Resp. Crit. Care 2008, 177, 1135–1141. [Google Scholar] [CrossRef] [Green Version]
- Gao, T.; Zhang, Y.; Shi, J.; Mohamed, S.R.; Xu, J.; Liu, X. The antioxidant guaiacol exerts fungicidal activity against fungal growth and deoxynivalenol production in Fusarium graminearum. Front. Microbiol. 2021, 12, 762844. [Google Scholar] [CrossRef]
- Wang, J.; Guo, J.; Zhang, Y.; Yan, X. Integrated transcriptomic and metabolomic analyses of yellow horn (Xanthoceras sorbifolia) in response to cold stress. PLoS ONE 2020, 15, e0236588. [Google Scholar] [CrossRef]
- Liu, Q.; Kong, W.; Hu, S.; Kang, Y.; Zhang, Y.; Ng, T.B. Effects of Oudemansiella radicata polysaccharide on postharvest quality of oyster mushroom (Pleurotus ostreatus) and its antifungal activity against Penicillium digitatum. Postharvest Biol. Technol. 2020, 166, 111207. [Google Scholar] [CrossRef]
- Vargas-Sánchez, R.D.; Torrescano-Urrutia, G.R.; Acedo-Félix, E.; Carvajal-Millán, E.; González-Córdova, A.F.; Valle-jo-Galland, B.; Torres-Llanez, M.J.; Sánchez-Escalante, A. Antioxidant and antimicrobial activity of commercial propolis ex-tract in beef patties. J. Food Sci. 2014, 79, 1499–1504. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Choi, A.G.J.; Cho, K.Y. Correlation of lipid peroxidation in Botrytis cinerea caused by dicarboximide fungicides with their fungicidal activity. J. Agric. Food Chem. 1998, 46, 737–741. [Google Scholar] [CrossRef]
- Heck, D.E.; Shakarjian, M.; Kim, H.D.; Laskin, J.D.; Vetrano, A.M. Mechanisms of oxidant generation by catalase. Ann. N. Acad. Sci. 2010, 1203, 120–125. [Google Scholar]
- Liu, T.; Fang, K.; Liu, Y.; Zhang, X.; Han, L.; Wang, X. Enantioselective residues and toxicity effects of the chiral triazole fun-gicide hexaconazole in earthworms (Eisenia fetida). Environ. Pollut. 2021, 270, 116269. [Google Scholar] [CrossRef]
- Lamb, D.C.; Waterman, M.R.; Kelly, S.L.; Guengerich, F.P. Cytochromes P450 and drug discovery. Curr. Opin. Biotechnol. 2007, 18, 504–512. [Google Scholar] [CrossRef]
- Degradi, L.; Tava, V.; Kunova, A.; Cortesi, P.; Saracchi, M.; Pasquali, M. Telomere to telomere genome assembly of Fusarium musae F31, Causal agent of crown rot disease of banana. Mol. Plant-Microbe Interact. 2021, 34, 1455–1457. [Google Scholar] [CrossRef]
- Hargrove, T.Y.; Wawrzak, Z.; Lamb, D.C.; Guengerich, F.P.; Lepesheva, G.I. Structure-functional characterization of cy-tochrome P450 sterol 14α-demethylase (CYP51B) from Aspergillus fumigatus and molecular basis for the development of an-tifungal drugs. J. Biol. Chem. 2015, 290, 23916–23934. [Google Scholar] [CrossRef] [Green Version]
- Qian, H.; Duan, M.; Sun, X.; Chi, M.; Zhao, Y.; Liang, W.; Du, J.; Huang, J.; Li, B. The binding mechanism between azoles and FgCYP51B, sterol 14α-demethylase of Fusarium graminearum. Pest Manag. Sci. 2017, 74, 126–134. [Google Scholar] [CrossRef]
- Cools, H.J.; Hawkins, N.; Fraaije, B. Constraints on the evolution of azole resistance in plant pathogenic fungi. Plant Pathol. 2013, 62, 36–42. [Google Scholar] [CrossRef] [Green Version]
- Parker, J.E.; Warrilow, A.G.S.; Cools, H.J.; Martel, C.M.; Nes, W.D.; Fraaije, B.A.; Lucas, J.A.; Kelly, D.E.; Kelly, S.L. Mechanism of binding of prothioconazole to Mycosphaerella graminicola CYP51 differs from that of other azole antifungals. Appl. Environ. Microbiol. 2011, 77, 1460–1465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagatova, A.; Keniya, M.V.; Wilson, R.K.; Sabherwal, M.; Tyndall, J.D.A.; Monk, B.C. Triazole resistance mediated by mutations of a conserved active site tyrosine in fungal lanosterol 14α-demethylase. Sci. Rep. 2016, 6, 26213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beck, M.E.; Negroni, J.; Matthiesen, S.; Kohnen, M.; Riplinger, C. A binding mode hypothesis for prothioconazole binding to CYP51 derived from first principles quantum chemistry. J. Comput. Mol. Des. 2020, 35, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.; Du, J.; Chi, M.; Sun, X.; Liang, W.; Huang, J.; Li, B. The Y137H mutation in the cytochrome P450 FgCYP51B protein confers reduced sensitivity to tebuconazole in Fusarium graminearum. Pest Manag. Sci. 2018, 74, 1472–1477. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Peng, H.; Wang, X.; Shao, F.; Yuan, Z.; Han, H. Graphene oxide exhibits broad-spectrum antimicrobial activity against bacterial phytopathogens and fungal conidia by intertwining and membrane perturbation. Nanoscale 2013, 6, 1879–1889. [Google Scholar] [CrossRef]
- Duan, Y.; Xiu, Q.; Li, H.; Li, T.; Wang, J.; Zhou, M. Pharmacological characteristics and control efficacy of a novel SDHI fun-gicide pydiflumetofen against Sclerotinia sclerotiorum. Plant Dis. 2019, 103, 77–82. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−△△CT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Xiang, D.; Chu, T.; Li, M.; Wang, Q.; Zhu, G. Effects of pyrethroid pesticide cis-bifenthrin on lipogenesis in hepatic cell line. Chemosphere 2018, 201, 840–849. [Google Scholar] [CrossRef]
- Keniya, M.V.; Sabherwal, M.; Wilson, R.K.; Woods, M.A.; Sagatova, A.A.; Tyndall, J.D.A.; Monk, B.C. Crystal structures of full-length lanosterol 14α-demethylases of prominent fungal pathogens Candida albicans and Candida glabrata provide tools for antifungal discovery. Antimicrob. Agents Chemother. 2018, 62, e01134-18. [Google Scholar] [CrossRef] [Green Version]
- Sagatova, A.A.; Keniya, M.V.; Tyndall, J.D.A.; Monk, B.C. Impact of homologous resistance mutations from pathogenic yeast on Saccharomyces cerevisiae lanosterol 14α-demethylase. Antimicrob. Agents Chemother. 2018, 62, e02242-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martí-Renom, M.A.; Stuart, A.C.; Fiser, A.; Sánchez, R.; Sali, A. Comparative protein structure modeling of genes and ge-nomes. Annu. Rev. Biophys. Biomol. Struct. 2000, 29, 291–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rich, M.A.; Hoth, L.R.; Geoghegan, K.F.; Brown, T.A.; LeMotte, P.; Simons, S.P.; Hensley, P.; Myszka, D.G. Kinetic analysis of estrogen receptor/ligand interactions. Proc. Natl. Acad. Sci. USA 2002, 99, 8562–8567. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (h) | Control | Pro Enantiomers (μg/mL) | |||
|---|---|---|---|---|---|
| S-Pro | R-Pro | ||||
| 0.4 | 0.8 | 0.4 | 0.8 | ||
| 0 | 15.15 ± 1.42 | 16.04 ± 2.40 | 16.37 ± 1.17 | 16.16 ± 2.42 | 15.49 ± 0.94 |
| 0.5 | 16.35 ± 0.86 | 16.98 ± 2.08 | 18.82 ± 1.32 | 19.88 ± 2.19 | 19.90 ± 0.95 |
| 1.0 | 33.94 ± 2.59 | 37.34 ± 2.53 | 43.13 ± 3.16 ** | 64.97 ± 1.80 | 82.65 ± 2.40 |
| 2.0 | 47.45 ± 1.87 | 52.06 ± 1.96 | 61.78 ± 3.42 *** | 78.34 ± 2.74 | 90.76 ± 2.79 |
| 4.0 | 61.84 ± 2.01 | 67.14 ± 3.37 | 75.69 ± 2.83 *** | 90.57 ± 1.28 | 93.96 ± 2.94 |
| 8.0 | 74.50 ± 4.17 | 79.91 ± 2.23 | 89.21 ± 1.78 *** | 95.53 ± 2.06 | 98.10 ± 1.44 |
| Item | Unit | Control | Pro Enantiomers (μg/mL) | |||
|---|---|---|---|---|---|---|
| S-Pro | R-Pro | |||||
| 0.4 | 0.8 | 0.4 | 0.8 | |||
| MDA | nmol/mgprot | 29.07 ± 0.61 | 34.02 ± 0.94 ** | 59.05 ± 2.06 *** | 45.57 ± 0.92 | 64.24 ± 0.92 *** |
| SOD | U/mgprot | 233.82 ± 13.33 | 197.71 ± 7.56 * | 90.31 ± 2.77 ** | 129.39 ± 4.66 | 81.69 ± 3.33 |
| CAT | U/mgprot | 205.80 ± 2.82 | 196.60 ± 1.69 * | 184.18 ± 2.73 *** | 180.02 ± 2.92 | 125.64 ± 2.46 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Gong, R.; Chu, Y.; Liu, S.; Xiang, D.; Li, C. Mechanistic Insights into Stereospecific Antifungal Activity of Chiral Fungicide Prothioconazole against Fusarium oxysporum F. sp. cubense. Int. J. Mol. Sci. 2022, 23, 2352. https://doi.org/10.3390/ijms23042352
Yang X, Gong R, Chu Y, Liu S, Xiang D, Li C. Mechanistic Insights into Stereospecific Antifungal Activity of Chiral Fungicide Prothioconazole against Fusarium oxysporum F. sp. cubense. International Journal of Molecular Sciences. 2022; 23(4):2352. https://doi.org/10.3390/ijms23042352
Chicago/Turabian StyleYang, Xiaofang, Ronggao Gong, Yuanqi Chu, Siwen Liu, Dandan Xiang, and Chunyu Li. 2022. "Mechanistic Insights into Stereospecific Antifungal Activity of Chiral Fungicide Prothioconazole against Fusarium oxysporum F. sp. cubense" International Journal of Molecular Sciences 23, no. 4: 2352. https://doi.org/10.3390/ijms23042352
APA StyleYang, X., Gong, R., Chu, Y., Liu, S., Xiang, D., & Li, C. (2022). Mechanistic Insights into Stereospecific Antifungal Activity of Chiral Fungicide Prothioconazole against Fusarium oxysporum F. sp. cubense. International Journal of Molecular Sciences, 23(4), 2352. https://doi.org/10.3390/ijms23042352