
Monitoring TRPC7 Conformational Changes by BRET Following GPCR Activation

 , ,
, ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
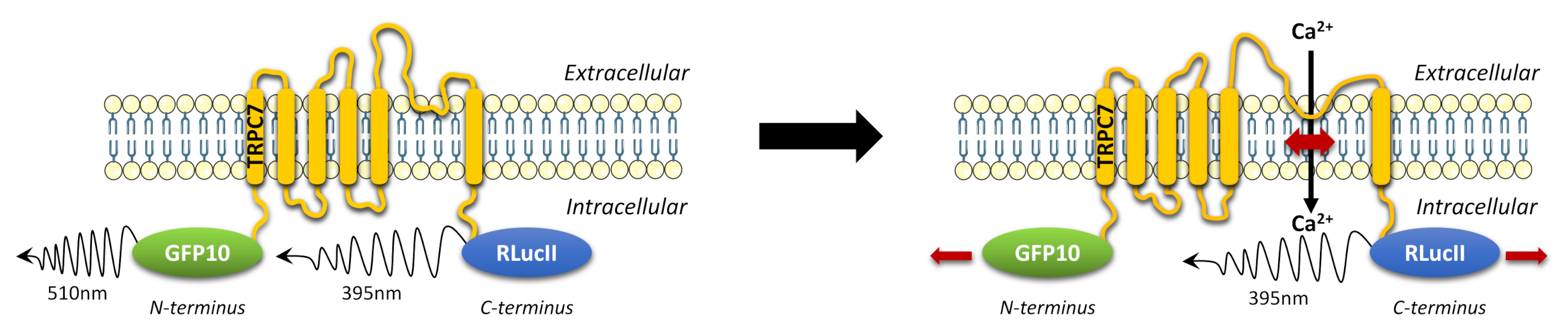
2.1. Design of TRPC7 BRET-Based Biosensors
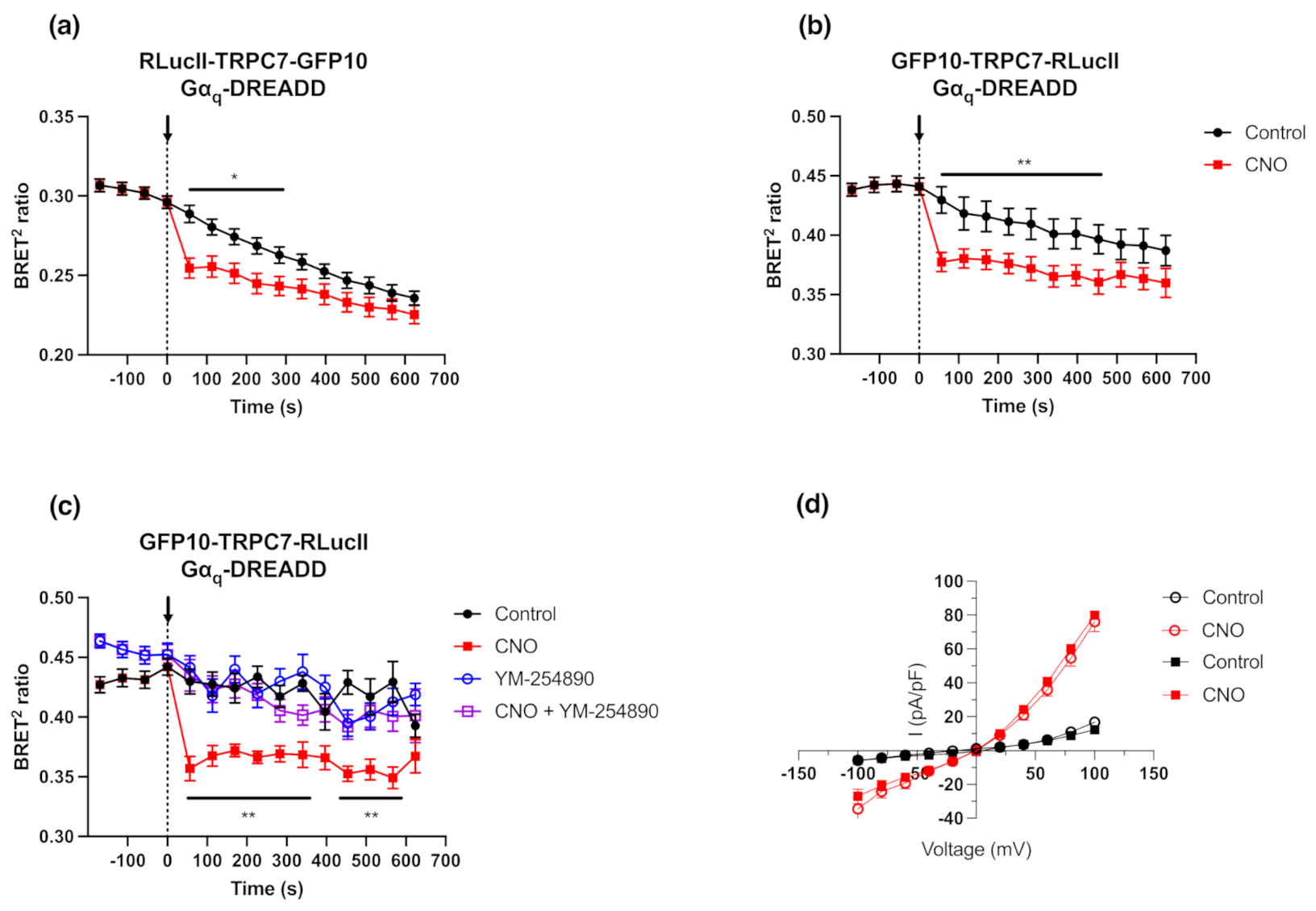
2.2. Activation of the Gαq Pathway Results in Conformational Changes of Functional Intramolecular TRPC7 Biosensors
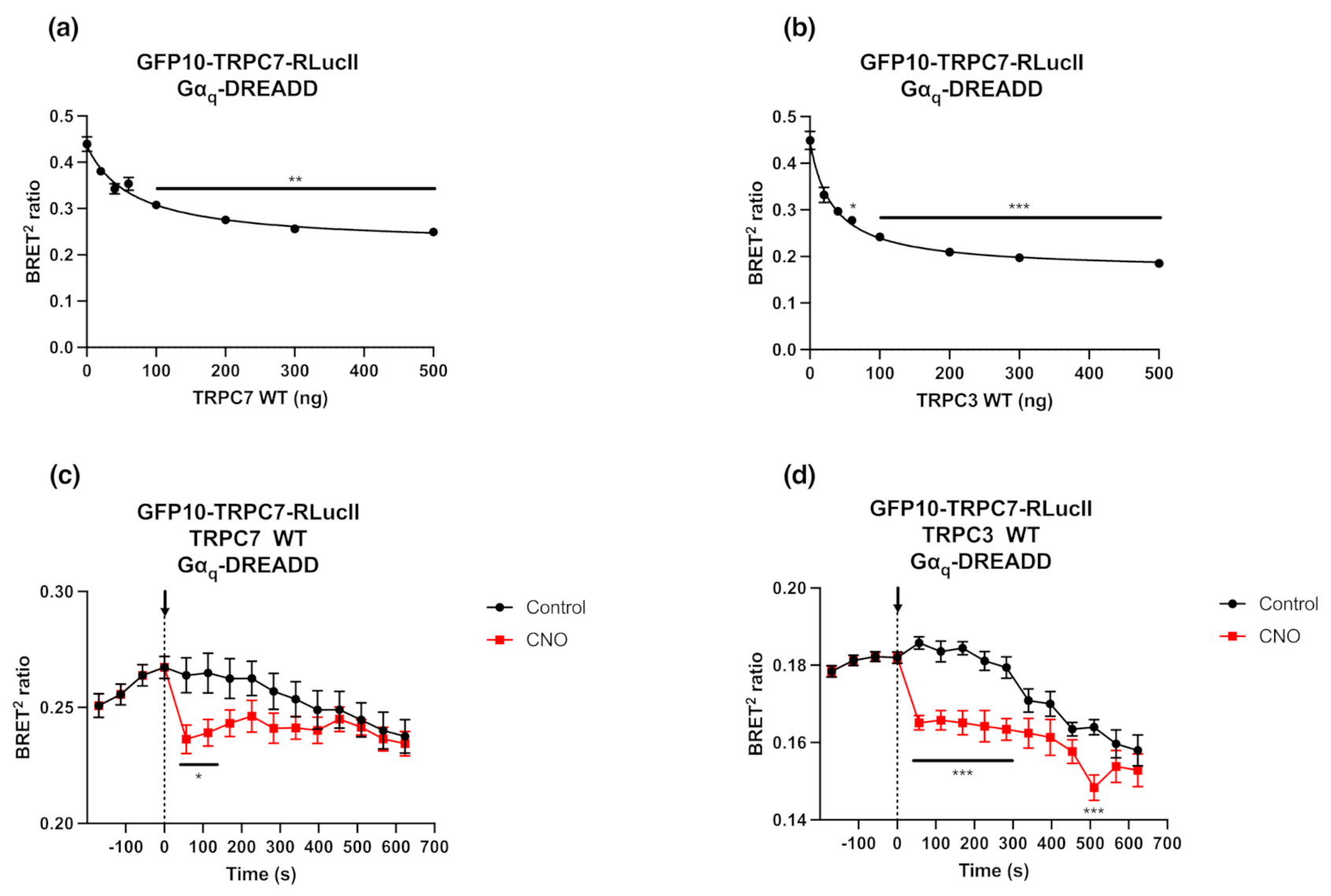
2.3. GFP10-TRPC7-RLucII Biosensor Competition with Untagged TRPC3 or TRPC7 Does Not Alter Its Conformational Changes in Response to Gαq-DREADD Stimulation
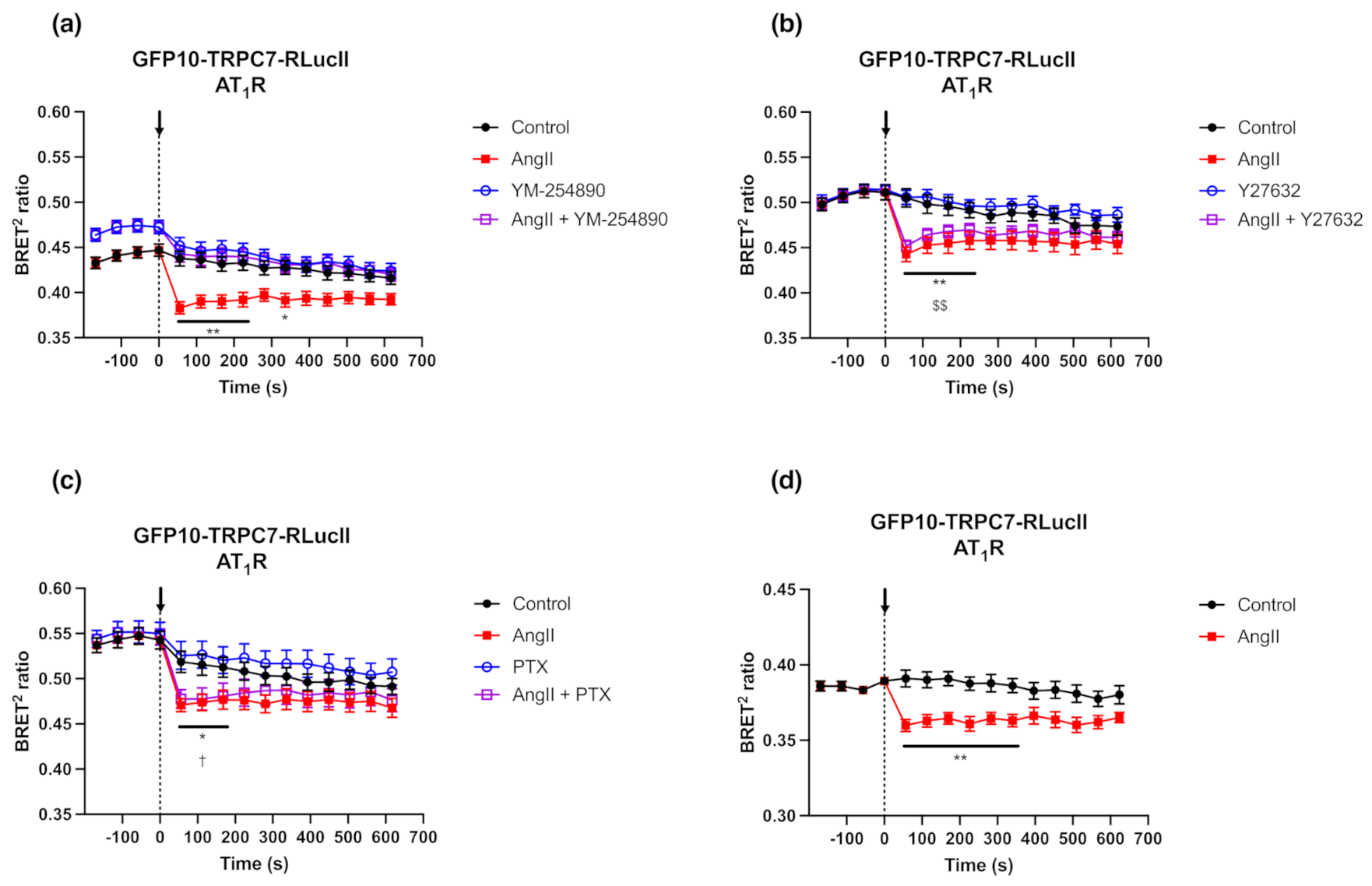
2.4. Gαq Signaling Is Required to Induce Conformational Changes in GFP10-TRPC7-RLucII BRET-Based Biosensor
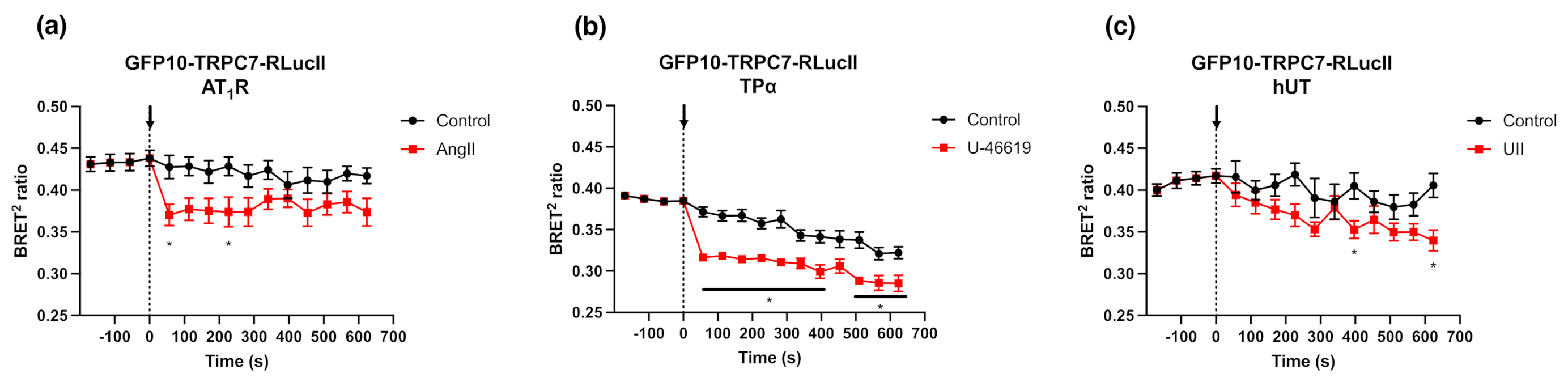
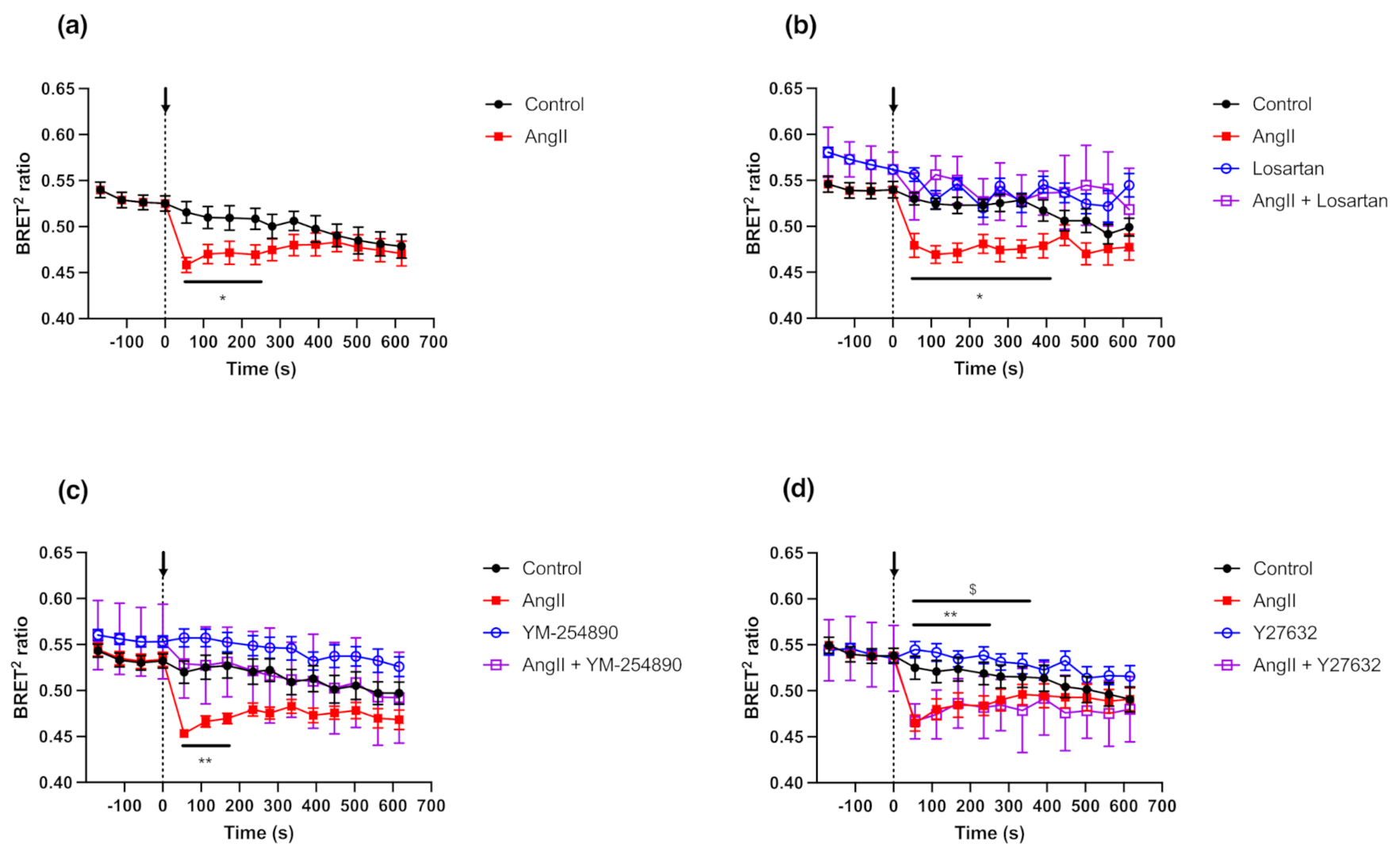
2.5. GFP10-TRPC7-RLucII Biosensor Shows Conformational Changes in Rat Cardiac Fibloblasts Expressing Endogenous AT1R
3. Materials and Methods
3.1. Materials
3.2. Plasmids
3.3. Cell Culture
3.4. BRET Assays
3.5. Electrophysiological Measurements
3.6. Cardiac Fibroblast Isolation and Infection
3.7. Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vannier, B.; Peyton, M.; Boulay, G.; Brown, D.; Qin, N.; Jiang, M.; Zhu, X.; Birnbaumer, L. Mouse trp2, the homologue of the human trpc2 pseudogene, encodes mTrp2, a store depletion-activated capacitative Ca2+ entry channel. Proc. Natl. Acad. Sci. USA 1999, 96, 2060–2064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vazquez, G.; Wedel, B.J.; Aziz, O.; Trebak, M.; Putney, J.W. The mammalian TRPC cation channels. Biochim. Biophys. Acta 2004, 1742, 21–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monteith, G.R.; Prevarskaya, N.; Roberts-Thomson, S.J. The calcium–cancer signalling nexus. Nat. Rev. Cancer 2017, 17, 373–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofmann, T.; Obukhov, A.G.; Schaefer, M.; Harteneck, C.; Gudermann, T.; Schultz, G. Direct activation of human TRPC6 and TRPC3 channels by diacylglycerol. Nature 1999, 397, 259–263. [Google Scholar] [CrossRef]
- Okada, T.; Inoue, R.; Yamazaki, K.; Maeda, A.; Kurosaki, T.; Yamakuni, T.; Tanaka, I.; Shimizu, S.; Ikenaka, K.; Imoto, K.; et al. Molecular and Functional Characterization of a Novel Mouse Transient Receptor Potential Protein Homologue TRP7. J. Biol. Chem. 1999, 274, 27359–27370. [Google Scholar] [CrossRef] [Green Version]
- Numaga, T.; Wakamori, M.; Mori, Y. TRPC7. In Transient Receptor Potential (TRP) Channels; Flockerzi, V., Nilius, B., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; Volume 179, pp. 143–151. [Google Scholar] [CrossRef]
- Hsu, W.; Tsai, M.; Wu, C.; Liang, J.; Lu, J.; Kahle, J.S.; Yu, H.; Yen, C.; Yen, C.; Hsieh, Y.; et al. Nociceptive transient receptor potential canonical 7 (TRPC7) mediates aging-associated tumorigenesis induced by ultraviolet B. Aging Cell 2019, 19, e13075. [Google Scholar] [CrossRef] [Green Version]
- Beck, B.; Zholos, A.; Sydorenko, V.; Roudbaraki, M.; Lehen’Kyi, V.; Bordat, P.; Prevarskaya, N.; Skryma, R. TRPC7 Is a Receptor-Operated DAG-Activated Channel in Human Keratinocytes. J. Investig. Dermatol. 2006, 126, 1982–1993. [Google Scholar] [CrossRef]
- Lievremont, J.-P.; Numaga, T.; Vazquez, G.; Lemonnier, L.; Hara, Y.; Mori, E.; Trebak, M.; Moss, S.E.; Bird, G.S.; Mori, Y.; et al. The Role of Canonical Transient Receptor Potential 7 in B-cell Receptor-activated Channels. J. Biol. Chem. 2005, 280, 35346–35351. [Google Scholar] [CrossRef] [Green Version]
- Itsuki, K.; Imai, Y.; Hase, H.; Okamura, Y.; Inoue, R.; Mori, M.X. PLC-mediated PI(4,5)P2 hydrolysis regulates activation and inactivation of TRPC6/7 channels. J. Gen. Physiol. 2014, 143, 183–201. [Google Scholar] [CrossRef] [Green Version]
- Vazquez, G.; Bird, G.S.J.; Mori, Y.; Putney, J. Native TRPC7 Channel Activation by an Inositol Trisphosphate Receptor-dependent Mechanism. J. Biol. Chem. 2006, 281, 25250–25258. [Google Scholar] [CrossRef] [Green Version]
- Lemonnier, L.; Trebak, M.; Putney, J.W. Complex regulation of the TRPC3, 6 and 7 channel subfamily by diacylglycerol and phosphatidylinositol-4,5-bisphosphate. Cell Calcium 2008, 43, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phelan, K.D.; Shwe, U.T.; Abramowitz, J.; Birnbaumer, L.; Zheng, F. Critical role of canonical transient receptor potential channel 7 in initiation of seizures. Proc. Natl. Acad. Sci. USA 2014, 111, 11533–11538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satoh, S.; Tanaka, H.; Ueda, Y.; Oyama, J.-I.; Sugano, M.; Sumimoto, H.; Mori, Y.; Makino, N. Transient receptor potential (TRP) protein 7 acts as a G protein-activated Ca2+ channel mediating angiotensin II-induced myocardial apoptosis. Mol. Cell. Biochem. 2006, 294, 205–215. [Google Scholar] [CrossRef]
- Gogebakan, B.; Bayraktar, R.; Suner, A.; Balakan, O.; Ulasli, M.; Izmirli, M.; Oztuzcu, S.; Camci, C. Do Fasudil and Y-27632 affect the level of transient receptor potential (TRP) gene expressions in breast cancer cell lines? Tumor Biol. 2014, 35, 8033–8041. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Iribe, G.; Nishida, M.; Naruse, K. Role of TRPC3 and TRPC6 channels in the myocardial response to stretch: Linking physiology and pathophysiology. Prog. Biophys. Mol. Biol. 2017, 130, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Onohara, N.; Nishida, M.; Inoue, R.; Kobayashi, H.; Sumimoto, H.; Sato, Y.; Mori, Y.; Nagao, T.; Kurose, H. TRPC3 and TRPC6 are essential for angiotensin II-induced cardiac hypertrophy. EMBO J. 2006, 25, 5305–5316. [Google Scholar] [CrossRef]
- Liu, C.-H.; Gong, Z.; Liang, Z.-L.; Liu, Z.-X.; Yang, F.; Sun, Y.-J.; Ma, M.-L.; Wang, Y.-J.; Ji, C.-R.; Wang, Y.-H.; et al. Arrestin-biased AT1R agonism induces acute catecholamine secretion through TRPC3 coupling. Nat. Commun. 2017, 8, 14335. [Google Scholar] [CrossRef] [PubMed]
- Ruigrok, H.J.; Shahid, G.; Goudeau, B.; de Gannes, F.P.; Poque-Haro, E.; Hurtier, A.; Lagroye, I.; Vacher, P.; Arbault, S.; Sojic, N.; et al. Full-Spectral Multiplexing of Bioluminescence Resonance Energy Transfer in Three TRPV Channels. Biophys. J. 2017, 112, 87–98. [Google Scholar] [CrossRef] [Green Version]
- Charest, P.G.; Terrillon, S.; Bouvier, M. Monitoring agonist-promoted conformational changes of β-arrestin in living cells by intramolecular BRET. EMBO Rep. 2005, 6, 334–340. [Google Scholar] [CrossRef] [Green Version]
- Bacart, J.; Corbel, C.; Jockers, R.; Bach, S.; Couturier, C. The BRET technology and its application to screening assays. Biotechnol. J. 2008, 3, 311–324. [Google Scholar] [CrossRef]
- Hu, J.; Stern, M.; Gimenez, L.E.D.; Wanka, L.; Zhu, L.; Rossi, M.; Meister, J.; Inoue, A.; Beck-Sickinger, A.G.; Gurevich, V.V.; et al. A G Protein-biased Designer G Protein-coupled Receptor Useful for Studying the Physiological Relevance of Gq/11-dependent Signaling Pathways. J. Biol. Chem. 2016, 291, 7809–7820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takasaki, J.; Saito, T.; Taniguchi, M.; Kawasaki, T.; Moritani, Y.; Hayashi, K.; Kobori, M. A Novel Gαq/11-selective Inhibitor. J. Biol. Chem. 2004, 279, 47438–47445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.; Li, S.; Liu, B.; Susperreguy, S.; Formoso, K.; Yao, J.; Kang, J.; Shi, A.; Birnbaumer, L.; Liao, Y. Major contribution of the 3/6/7 class of TRPC channels to myocardial ischemia/reperfusion and cellular hypoxia/reoxygenation injuries. Proc. Natl. Acad. Sci. USA 2017, 114, E4582–E4591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chappe, Y.; Michel, P.; Joushomme, A.; Barbeau, S.; Pierredon, S.; Baron, L.; Garenne, A.; De Gannes, F.P.; Hurtier, A.; Mayer, S.; et al. High-Throughput Screening of TRPV1 Ligands in the Light of the Bioluminescence Resonance Energy Transfer Technique. Mol. Pharmacol. 2021, 100, 237–257. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, T.; Schaefer, M.; Schultz, G.; Gudermann, T. Subunit composition of mammalian transient receptor potential channels in living cells. Proc. Natl. Acad. Sci. USA 2002, 99, 7461–7466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkatachalam, K.; Montell, C. TRP Channels. Annu. Rev. Biochem. 2007, 76, 387–417. [Google Scholar] [CrossRef] [Green Version]
- Myeong, J.; Ko, J.; Hong, C.; Yang, D.; Lee, K.P.; Jeon, J.-H.; So, I. The interaction domains of transient receptor potential canonical (TRPC)1/4 and TRPC1/5 heteromultimeric channels. Biochem. Biophys. Res. Commun. 2016, 474, 476–481. [Google Scholar] [CrossRef]
- St-Pierre, D.; Cabana, J.; Holleran, B.J.; Besserer-Offroy, A.; Escher, E.; Guillemette, G.; Lavigne, P.; Leduc, R. Angiotensin II cyclic analogs as tools to investigate AT1R biased signaling mechanisms. Biochem. Pharmacol. 2018, 154, 104–117. [Google Scholar] [CrossRef] [Green Version]
- Galandrin, S.; Denis, C.; Boularan, C.; Marie, J.; M’Kadmi, C.; Pilette, C.; Dubroca, C.; Nicaise, Y.; Seguelas, M.-H.; N’Guyen, D.; et al. Cardioprotective Angiotensin-(1–7) Peptide Acts as a Natural-Biased Ligand at the Angiotensin II Type 1 Receptor. Hypertension 2016, 68, 1365–1374. [Google Scholar] [CrossRef] [Green Version]
- Namkung, Y.; LeGouill, C.; Kumar, S.; Cao, Y.; Teixeira, L.B.; Lukasheva, V.; Giubilaro, J.; Simões, S.C.; Longpré, J.-M.; Devost, D.; et al. Functional selectivity profiling of the angiotensin II type 1 receptor using pathway-wide BRET signaling sensors. Sci. Signal. 2018, 11, eaat1631. [Google Scholar] [CrossRef] [Green Version]
- Jeon, J.-P.; Lee, K.P.; Park, E.J.; Sung, T.S.; Kim, B.J.; Jeon, J.-H.; So, I. The specific activation of TRPC4 by Gi protein subtype. Biochem. Biophys. Res. Commun. 2008, 377, 538–543. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.-P.; Hong, C.; Park, E.-J.; Jeon, J.-H.; Cho, N.-H.; Kim, I.-G.; Choe, H.; Muallem, S.; Kim, H.J.; So, I. Selective Gαi Subunits as Novel Direct Activators of Transient Receptor Potential Canonical (TRPC)4 and TRPC5 Channels. J. Biol. Chem. 2012, 287, 17029–17039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zholos, A.V. TRPC5. In Mammalian Transient Receptor Potential (TRP) Cation Channels; Springer: Berlin/Heidelberg, Germany, 2014; Volume 222, pp. 129–156. [Google Scholar]
- Nishida, M.; Onohara, N.; Sato, Y.; Suda, R.; Ogushi, M.; Tanabe, S.; Inoue, R.; Mori, Y.; Kurose, H. Gα12/13-mediated Up-regulation of TRPC6 Negatively Regulates Endothelin-1-induced Cardiac Myofibroblast Formation and Collagen Synthesis through Nuclear Factor of Activated T Cells Activation. J. Biol. Chem. 2007, 282, 23117–23128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, R.A.; Hatano, N.; Ohya, S.; Imaizumi, Y.; Giles, W.R. C-type natriuretic peptide activates a non-selective cation current in acutely isolated rat cardiac fibroblasts via natriuretic peptide C receptor-mediated signalling. J. Physiol. 2007, 580, 255–274. [Google Scholar] [CrossRef]
- Villarreal, F.J.; Kim, N.N.; Ungab, G.D.; Printz, M.P.; Dillmann, W.H. Identification of functional angiotensin II receptors on rat cardiac fibroblasts. Circulation 1993, 88, 2849–2861. [Google Scholar] [CrossRef] [Green Version]
- Van Kesteren, C.; Van Heugten, H.; Lamers, J.; Saxena, P.; Schalekamp, M.; Danser, A. Angiotensin II-mediated Growth and Antigrowth Effects in Cultured Neonatal Rat Cardiac Myocytes and Fibroblasts. J. Mol. Cell. Cardiol. 1997, 29, 2147–2157. [Google Scholar] [CrossRef]
- Dostal, D.E.; Rothblum, K.N.; Conrad, K.M.; Cooper, G.R.; Baker, K.M. Detection of angiotensin I and II in cultured rat cardiac myocytes and fibroblasts. Am. J. Physiol. Physiol. 1992, 263, C851–C863. [Google Scholar] [CrossRef]
- Davis, J.; Burr, A.R.; Davis, G.F.; Birnbaumer, L.; Molkentin, J.D. A TRPC6-Dependent Pathway for Myofibroblast Transdifferentiation and Wound Healing In Vivo. Dev. Cell 2012, 23, 705–715. [Google Scholar] [CrossRef] [Green Version]
- Davis, J.; Molkentin, J.D. Myofibroblasts: Trust your heart and let fate decide. J. Mol. Cell. Cardiol. 2013, 70, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Londoño, J.E.C.; Marx, A.; Kraft, A.E.; Schürger, A.; Richter, C.; Dietrich, A.; Lipp, P.; Birnbaumer, L.; Freichel, M. Angiotensin-II-Evoked Ca2+ Entry in Murine Cardiac Fibroblasts Does Not Depend on TRPC Channels. Cells 2020, 9, 322. [Google Scholar] [CrossRef] [Green Version]
- Wen, H.; Gwathmey, J.K.; Xie, L.-H. Role of Transient Receptor Potential Canonical Channels in Heart Physiology and Pathophysiology. Front. Cardiovasc. Med. 2020, 7, 24. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Armillei, M.K.; Yu, A.S.; Liang, B.T.; Runnels, L.W.; Yue, L. Ca2+ Signaling in Cardiac Fibroblasts and Fibrosis-Associated Heart Diseases. J. Cardiovasc. Dev. Dis. 2019, 6, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Besser, L.; Chorin, E.; Sekler, I.; Silverman, W.F.; Atkin, S.; Russell, J.T.; Hershfinkel, M. Synaptically Released Zinc Triggers Metabotropic Signaling via a Zinc-Sensing Receptor in the Hippocampus. J. Neurosci. 2009, 29, 2890–2901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Sakoda, H.; Nakazato, Y.; Islam, N.; Pattou, F.; Kerr-Conte, J.; Nakazato, M. Neuromedin U uses Gαi2 and Gαo to suppress glucose-stimulated Ca2+ signaling and insulin secretion in pancreatic β cells. PLoS ONE 2021, 16, e0250232. [Google Scholar] [CrossRef] [PubMed]
- Lavenus, S.; Simard, É.; Besserer-Offroy, É.; Froehlich, U.; Leduc, R.; Grandbois, M. Label-free cell signaling pathway deconvolution of angiotensin type 1 receptor reveals time-resolved G-protein activity and distinct AngII and AngIIIIV responses. Pharmacol. Res. 2018, 136, 108–120. [Google Scholar] [CrossRef] [Green Version]
- Namkung, Y.; Le Gouill, C.; Lukashova, V.; Kobayashi, H.; Hogue, M.; Khoury, E.; Song, M.; Bouvier, M.; Laporte, S.A. Monitoring G protein-coupled receptor and β-arrestin trafficking in live cells using enhanced bystander BRET. Nat. Commun. 2016, 7, 12178. [Google Scholar] [CrossRef] [Green Version]
- Ehrhardt, C.; Schmolke, M.; Matzke, A.; Knoblauch, A.; Will, C.; Wixler, V.; Ludwig, S. Polyethylenimine, a cost-effective transfection reagent. Signal Transduct. 2006, 6, 179–184. [Google Scholar] [CrossRef]
- Flynn, R.; Chapman, K.; Iftinca, M.; Aboushousha, R.; Varela, D.; Altier, C. Targeting the Transient Receptor Potential Vanilloid Type 1 (TRPV1) Assembly Domain Attenuates Inflammation-Induced Hypersensitivity. J. Biol. Chem. 2014, 289, 16675–16687. [Google Scholar] [CrossRef] [Green Version]
- Iftinca, M.; Flynn, R.; Basso, L.; Melo, H.; Aboushousha, R.; Taylor, L.; Altier, C. The stress protein heat shock cognate 70 (Hsc70) inhibits the Transient Receptor Potential Vanilloid type 1 (TRPV1) channel. Mol. Pain 2016, 12, 1744806916663945. [Google Scholar] [CrossRef]
- Malette, J.; Degrandmaison, J.; Giguère, H.; Berthiaume, J.; Frappier, M.; Parent, J.-L.; Auger-Messier, M.; Boulay, G. MURC/CAVIN-4 facilitates store-operated calcium entry in neonatal cardiomyocytes. Biochim. Biophys. Acta 2019, 1866, 1249–1259. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pétigny, C.; Dumont, A.-A.; Giguère, H.; Collette, A.; Holleran, B.J.; Iftinca, M.; Altier, C.; Besserer-Offroy, É.; Auger-Messier, M.; Leduc, R. Monitoring TRPC7 Conformational Changes by BRET Following GPCR Activation. Int. J. Mol. Sci. 2022, 23, 2502. https://doi.org/10.3390/ijms23052502
Pétigny C, Dumont A-A, Giguère H, Collette A, Holleran BJ, Iftinca M, Altier C, Besserer-Offroy É, Auger-Messier M, Leduc R. Monitoring TRPC7 Conformational Changes by BRET Following GPCR Activation. International Journal of Molecular Sciences. 2022; 23(5):2502. https://doi.org/10.3390/ijms23052502
Chicago/Turabian StylePétigny, Cécile, Audrey-Ann Dumont, Hugo Giguère, Audrey Collette, Brian J. Holleran, Mircea Iftinca, Christophe Altier, Élie Besserer-Offroy, Mannix Auger-Messier, and Richard Leduc. 2022. "Monitoring TRPC7 Conformational Changes by BRET Following GPCR Activation" International Journal of Molecular Sciences 23, no. 5: 2502. https://doi.org/10.3390/ijms23052502
APA StylePétigny, C., Dumont, A. -A., Giguère, H., Collette, A., Holleran, B. J., Iftinca, M., Altier, C., Besserer-Offroy, É., Auger-Messier, M., & Leduc, R. (2022). Monitoring TRPC7 Conformational Changes by BRET Following GPCR Activation. International Journal of Molecular Sciences, 23(5), 2502. https://doi.org/10.3390/ijms23052502