
The Anti-Proliferative and Apoptotic Effects of Rutaecarpine on Human Esophageal Squamous Cell Carcinoma Cell Line CE81T/VGH In Vitro and In Vivo


Abstract
:1. Introduction
2. Results
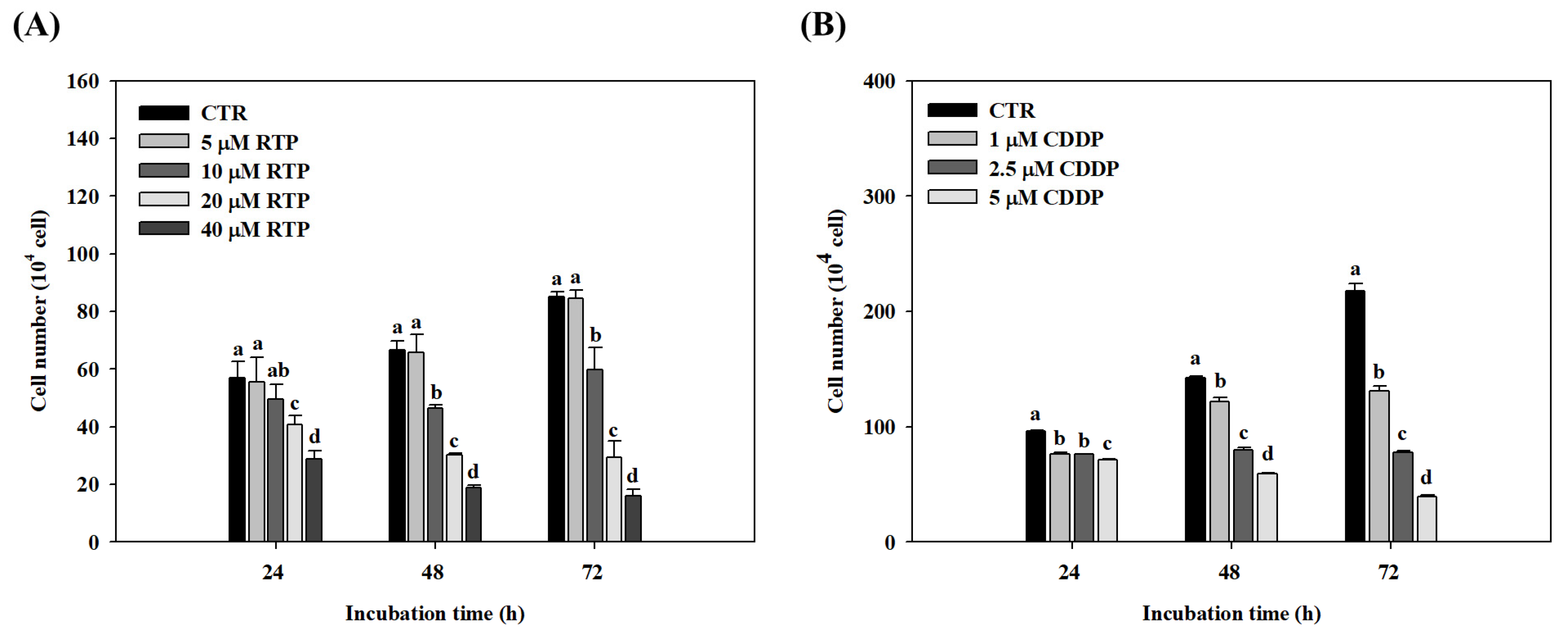
2.1. Effect of RTP and CDDP on the Growth of Human Esophageal Squamous Cell Carcinoma Cell Line CE81T/VGH
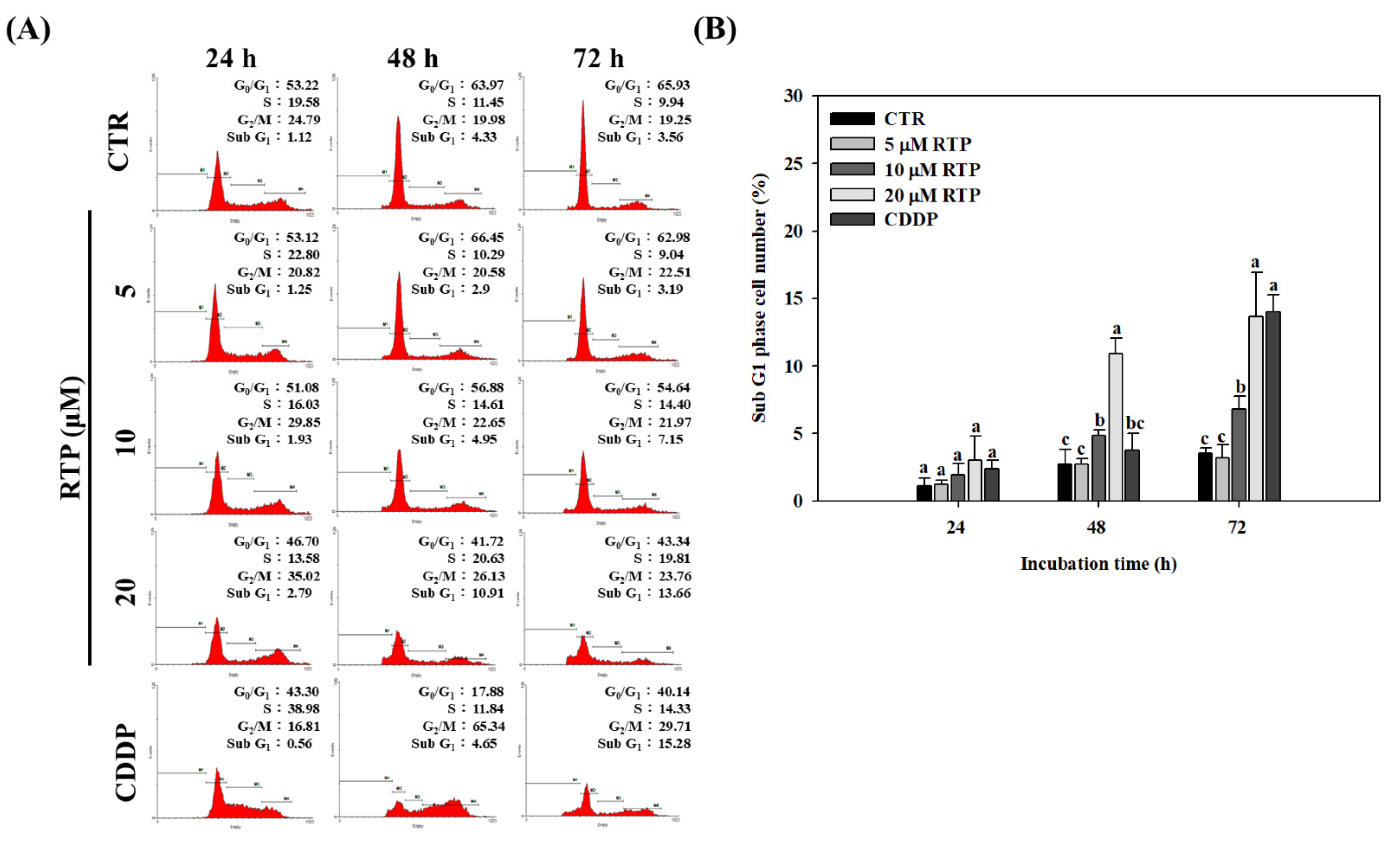
2.2. Effect of RTP on the Cell Cycle of Human Esophageal Squamous Cell Carcinoma Cell Line CE81T/VGH
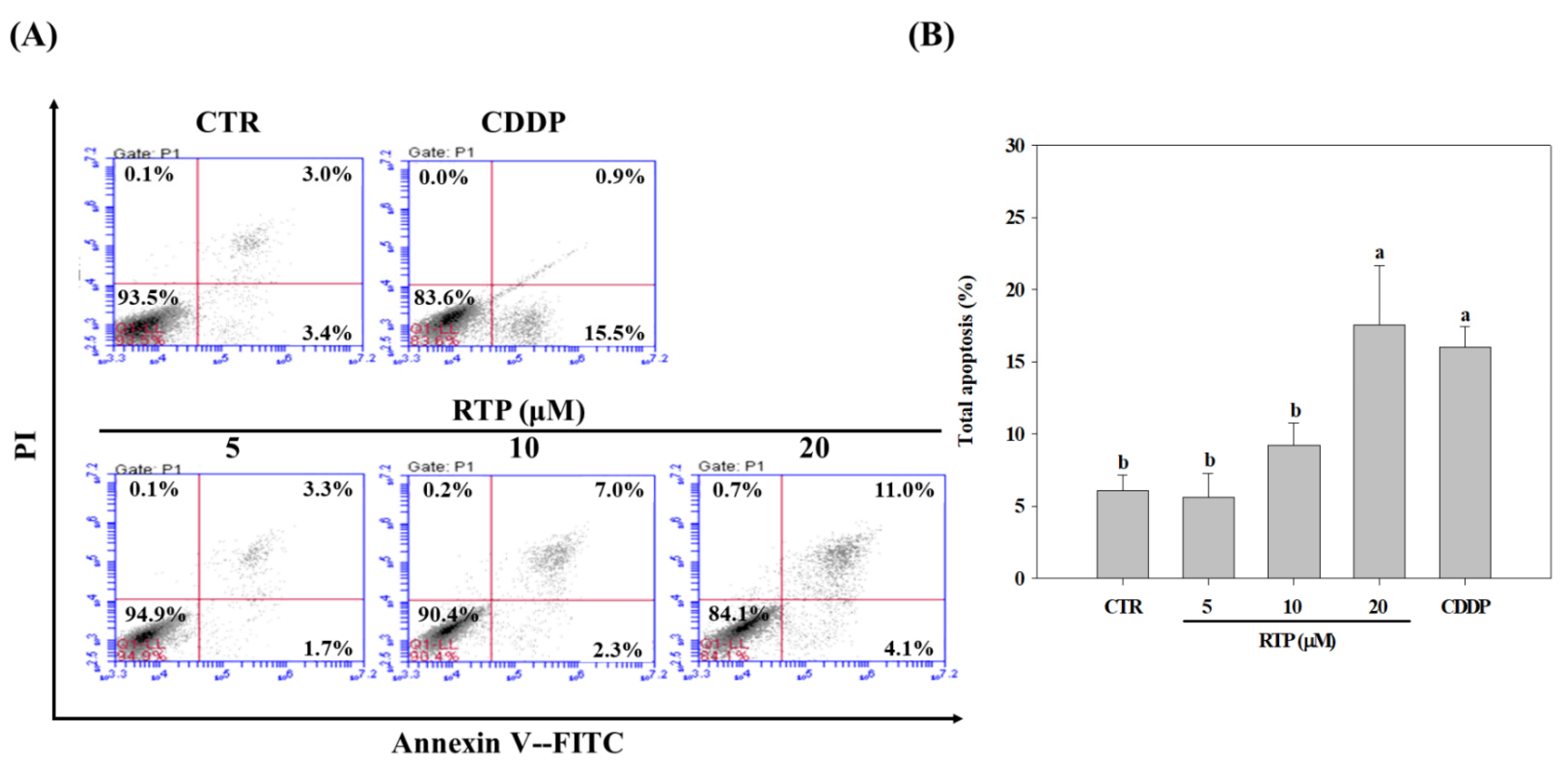
2.3. Effect of RTP on the Apoptosis of Human Esophageal Squamous Cell Carcinoma Cell Line CE81T/VGH
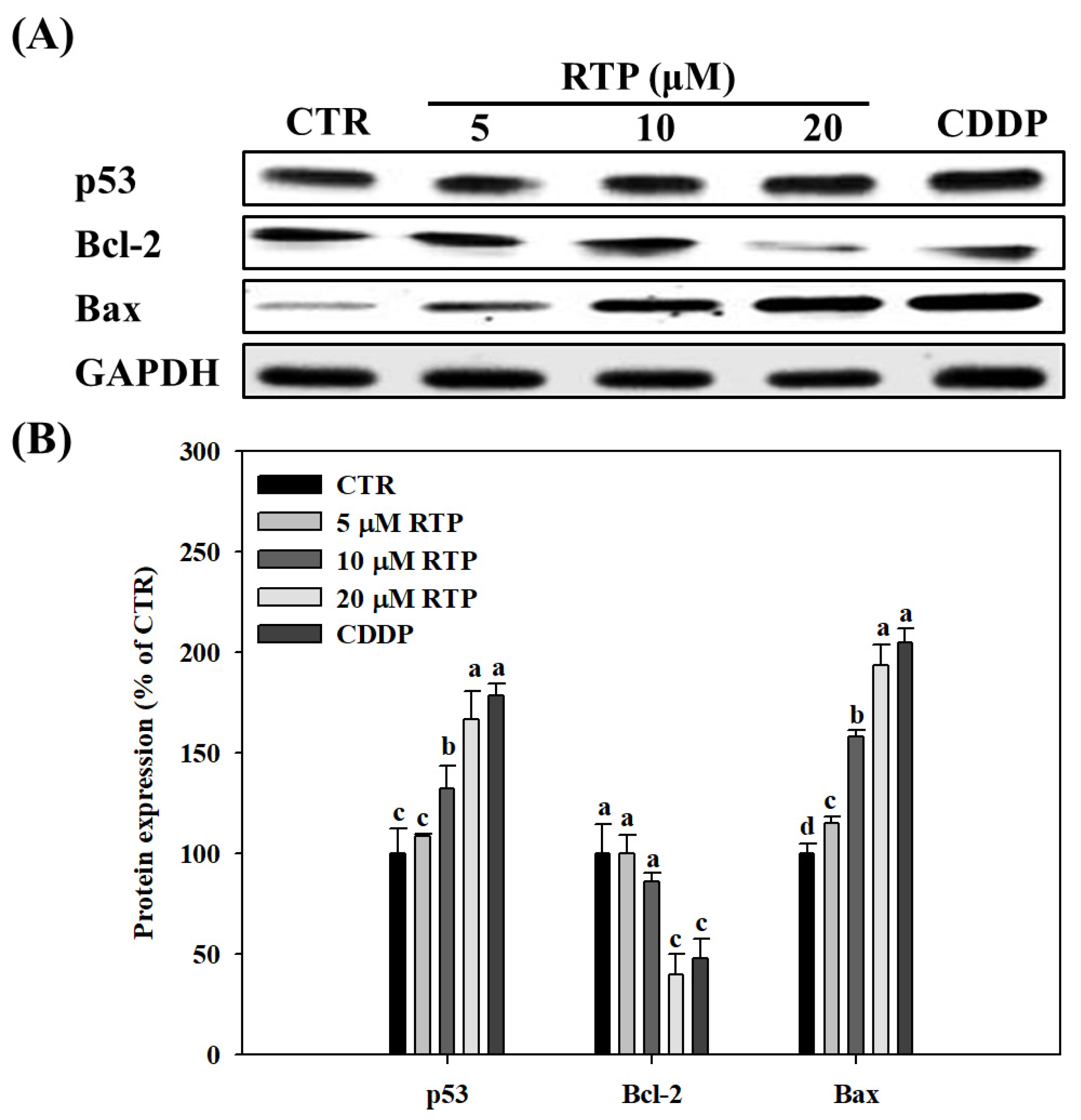
2.4. Effect of RTP on the Expressions of Human Esophageal Squamous Cell Carcinoma Cell Line CE81T/VGH, and p53, Bcl-2, and Bax Proteins
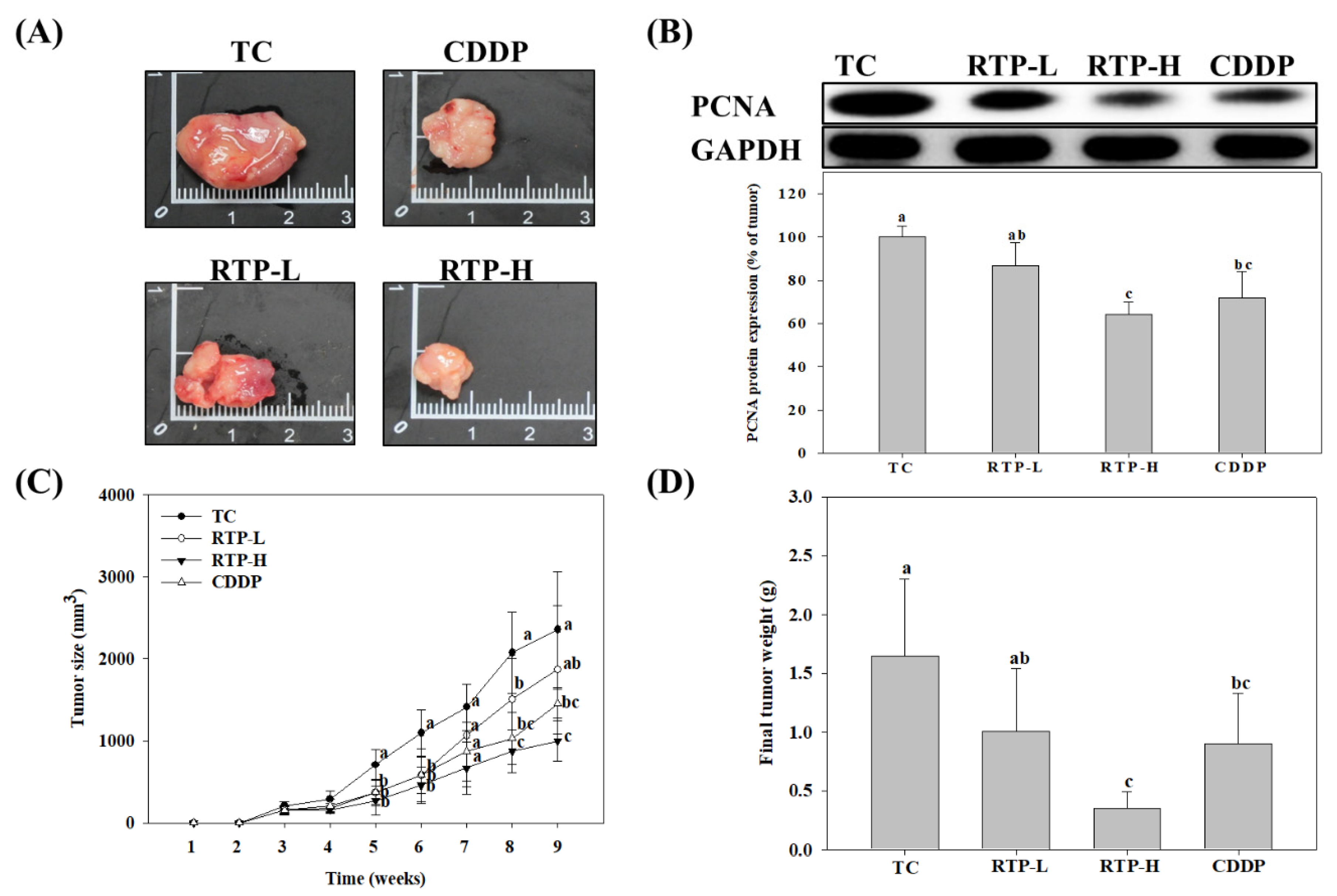
2.5. Effect of RTP on Tumor Growth in the Nude Mice Xenograft Model
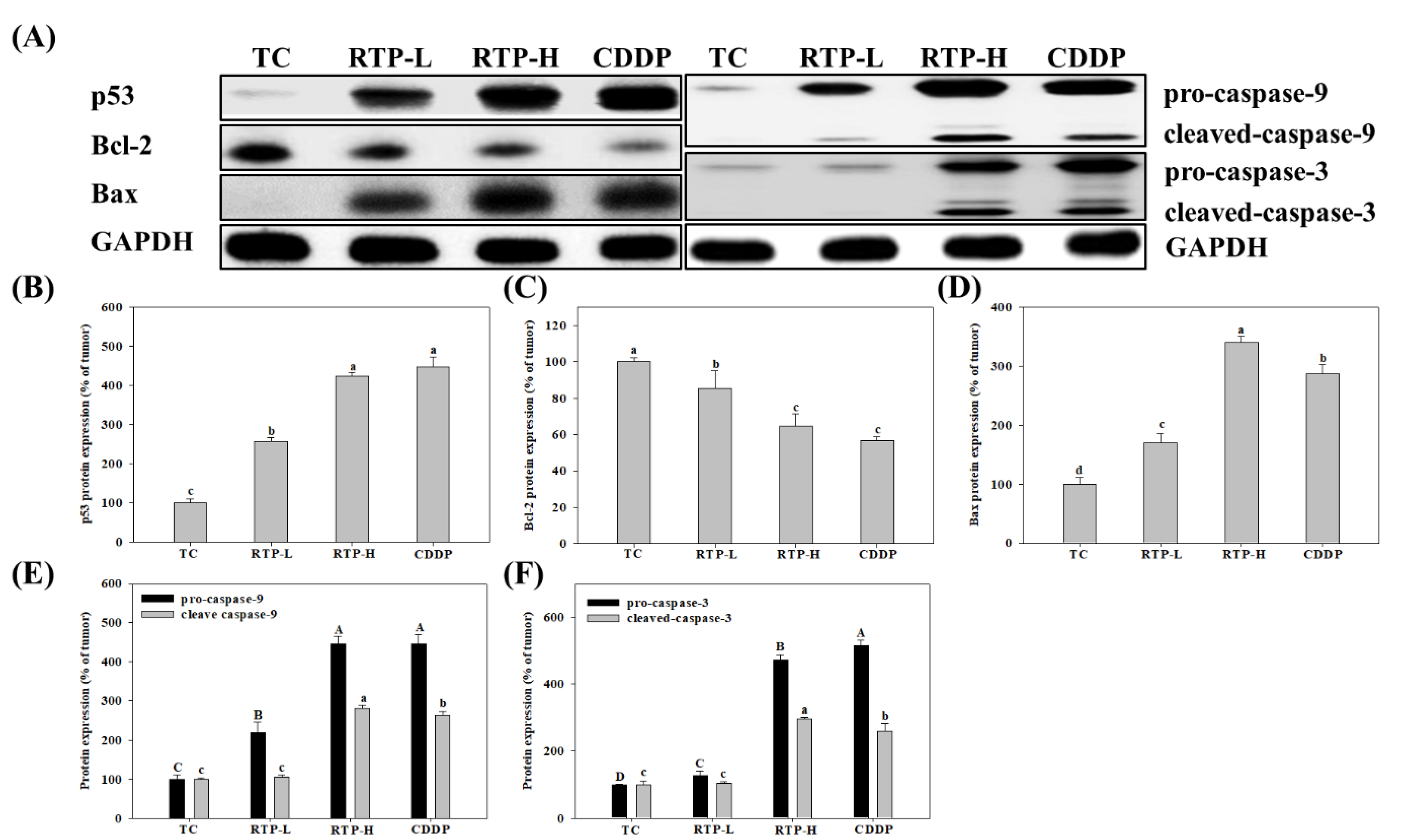
2.6. Effect of RTP on p53, Bcl-2, Bax, Caspase-9, and Caspase-3 Protein Expression in Tumor Tissue in the Nude Mice Xenograft Model
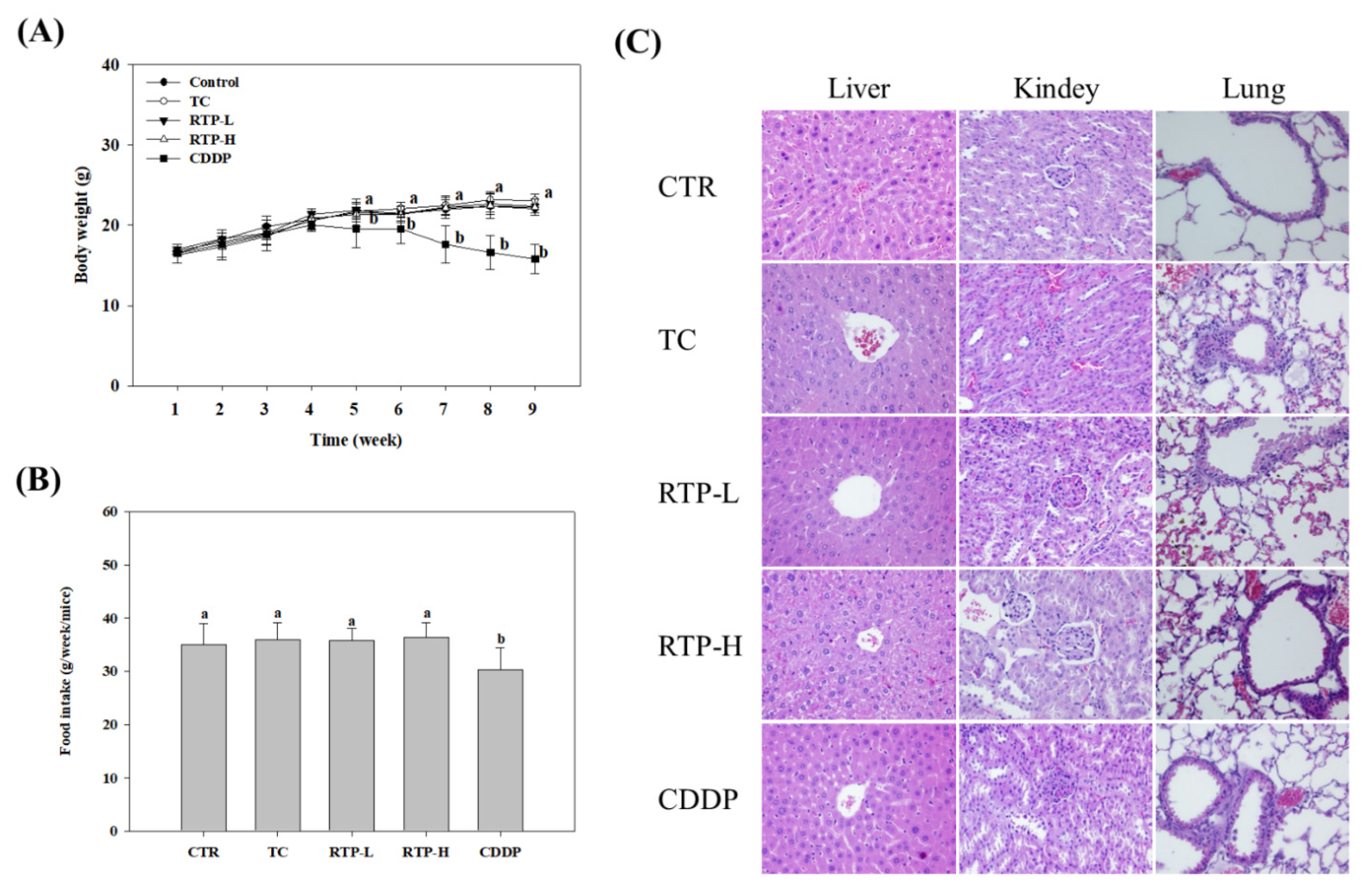
2.7. Effect of RTP Toxicity in the Nude Mice Xenograft Model
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Animal Study
4.3. Cell Culture
4.4. Cell Growth Analysis
4.5. Cell Cycle Analysis
4.6. Annexin V-FITC-Propidium Iodide Assay
4.7. Western Blotting
4.8. Hematological Assay
4.9. Biochemical Assay
4.10. Histopathological Examination
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Enzinger, P.C.; Mayer, R.J. Esophageal cancer. N. Engl. J. Med. 2003, 349, 2241–2252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, S.T.; Wang, P.J.; Liou, N.J.; Lin, P.S.; Chen, C.H.; Chang, W.C. ICAM1 Is a Potential Cancer Stem Cell Marker of Esophageal Squamous Cell Carcinoma. PLoS ONE 2015, 10, e0142834. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Otake, R.; Kozuki, R.; Toihata, T.; Takahashi, K.; Okamura, A.; Imamura, Y. Recent progress in multidisciplinary treatment for patients with esophageal cancer. Surg. Today. 2020, 50, 12–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grünberger, B.; Raderer, M.; Schmidinger, M.; Hejna, M. Palliative chemotherapy for recurrent and metastatic esophageal cancer. Anticancer Res. 2007, 27, 2705–2714. [Google Scholar] [PubMed]
- Watanabe, M.; Baba, Y.; Yoshida, N.; Ishimoto, T.; Nagai, Y.; Iwatsuki, M.; Iwagami, S.; Baba, H. Outcomes of preoperative chemotherapy with docetaxel, cisplatin, and 5-fluorouracil followed by esophagectomy in patients with resectable node-positive esophageal cancer. Ann. Surg. Oncol. 2014, 21, 2838–2844. [Google Scholar] [CrossRef] [PubMed]
- Pennathur, A.; Gibson, M.K.; Jobe, B.A.; Luketich, J.D. Oesophageal carcinoma. Lancet 2013, 381, 400–412. [Google Scholar] [CrossRef] [Green Version]
- Chiu, W.C.; Lee, Y.C.; Su, Y.H.; Wang, Y.Y.; Tsai, C.H.; Hou, Y.A.; Wang, C.H.; Huang, Y.F.; Huang, C.J.; Chou, S.H.; et al. The Synthetic β-Nitrostyrene Derivative CYT-Rx20 Inhibits Esophageal Tumor Growth and Metastasis via PI3K/AKT and STAT3 Pathways. PLoS ONE 2016, 11, e0166453. [Google Scholar] [CrossRef]
- Zhao, Z.; He, X.; Han, W.; Chen, X.; Liu, P.; Zhao, X.; Wang, X.; Zhang, L.; Wu, S.; Zheng, X. Genus Tetradium L.: A comprehensive review on traditional uses, phytochemistry, and pharmacological activities. J. Ethnopharmacol. 2019, 231, 337–354. [Google Scholar] [CrossRef]
- Li, X.; Ge, J.; Zheng, Q.; Zhang, J.; Sun, R.; Liu, R. Evodiamine and rutaecarpine from Tetradium ruticarpum in the treatment of liver diseases. Phytomedicine 2020, 68, 153180. [Google Scholar] [CrossRef]
- Xue, Y.; Guo, T.; Zou, L.; Gong, Y.; Wu, B.; Yi, Z.; Jia, T.; Zhao, S.; Shi, L.; Li, L.; et al. Evodiamine attenuates P2X7-Mediated inflammatory injury of human umbilical vein endothelial cells exposed to high free fatty acids. Oxid. Med. Cell Longev. 2018, 2018, 5082817. [Google Scholar] [CrossRef] [Green Version]
- Ivanova, B.; Spiteller, M. Evodiamine and rutaecarpine alkaloids as highly selective transient receptor potential vanilloid 1 agonists. Int. J. Biol. Macromol. 2014, 65, 314–324. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.P.; Xiao, L.; Deng, H.W.; Li, Y.J. The depressor and vasodilator effects of rutaecarpine are mediated by calcitonin gene-related peptide. Planta Med. 2003, 69, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Sheu, J.R.; Kan, Y.C.; Hung, W.C.; Su, C.H.; Lin, C.H.; Lee, Y.M.; Yen, M.H. The antiplatelet activity of rutaecarpine, an alkaloid isolated from Evodia rutaecarpa, is mediated through inhibition of phospholipase C. Thromb. Res. 1998, 92, 53–64. [Google Scholar] [CrossRef]
- Lee, C.; Liao, J.; Chen, S.; Yen, C.; Lee, Y.; Huang, S.; Huang, S.; Lin, C.; Chang, V.H. Fluorine-modified rutaecarpine exerts cyclooxygenase-2 inhibition and anti-inflammatory effects in lungs. Front. Pharmacol. 2019, 10, 91. [Google Scholar] [CrossRef]
- Li, Z.; Yang, M.; Peng, Y.; Gao, M.; Yang, B. Rutaecarpine ameliorated sepsisinduced peritoneal resident macrophages apoptosis and inflammation responses. Life Sci. 2019, 228, 11–20. [Google Scholar] [CrossRef]
- Liao, Y.; Liu, Y.; Xia, X.; Shao, Z.; Huang, C.; He, J.; Jiang, L.; Tang, D.; Liu, J.; Huang, H. Targeting GRP78-dependent AR-V7 protein degradation overcomes castration-resistance in prostate cancer therapy. Theranostics 2020, 10, 3366–3381. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.Y.; Yeh, T.H. Rutaecarpine administration inhibits cancer cell growth in allogenic TRAMP-C1 prostate cancer mice correlating with immune balance in vivo. Biomed. Pharmacother. 2021, 139, 111648. [Google Scholar] [CrossRef] [PubMed]
- Cokluk, E.; Ozman, Z.; Eskiler, G.G.; Ozkan, A.D.; Sekeroglu, M.R. Comparison of the effects of rutaecarpine on molecular subtypes of breast cancer. J. Cancer Res. Ther. 2021, 17, 988–993. [Google Scholar]
- Ji, L.; Wu, M.; Li, Z. Rutacecarpine Inhibits Angiogenesis by Targeting the VEGFR2 and VEGFR2-Mediated Akt/mTOR/p70s6k Signaling Pathway. Molecules 2018, 23, 2047. [Google Scholar] [CrossRef] [Green Version]
- Stewart, D.J. Mechanisms of resistance to cisplatin and carboplatin. Crit. Rev. Oncol. Hematol. 2007, 63, 12–31. [Google Scholar] [CrossRef] [PubMed]
- Launay-Vacher, V.; Rey, J.B.; Isnard-Bagnis, C.; Deray, G.; Daouphars, M. Prevention of cisplatin nephrotoxicity: State of the art and recommendations from the European Society of Clinical Pharmacy Special Interest Group on Cancer Care. Cancer Chemother. Pharmacol. 2008, 61, 903–909. [Google Scholar] [CrossRef]
- Ojha, S.; Venkataraman, B.; Kurdi, A.; Mahgoub, E.; Sadek, B.; Rajesh, M. Plant-Derived Agents for Counteracting Cisplatin-Induced Nephrotoxicity. Oxid. Med. Cell Longev. 2016, 2016, 4320374. [Google Scholar] [CrossRef] [Green Version]
- Fridman, Y.; Palgi, N.; Dovrat, D.; Ben-Aroya, S.; Hieter, P.; Aharoni, A. Subtle alterations in PCNA-partner interactions severely impair DNA replication and repair. PLoS Biol. 2010, 8, e1000507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, Q.; Zhang, J.; Yi, Y.; Huang, Y.; Wang, Y.; Wang, Y.; Zhang, W. Proliferating Cell Nuclear Antigen Has an Association with Prognosis and Risks Factors of Cancer Patients: A Systematic Review. Mol. Neurobiol. 2016, 53, 6209–6217. [Google Scholar] [CrossRef] [PubMed]
- Mattern, J.; Volm, M. Imbalance of cell proliferation and apoptosis during progression of lung carcinomas. Anticancer Res. 2004, 24, 4243–4276. [Google Scholar]
- Lee, S.H.; Son, J.K.; Jeong, B.S.; Jeong, T.C.; Chang, H.W.; Lee, E.S.; Jahng, Y. Progress in the studies on Rutaecarpine. Molecules 2008, 13, 272–300. [Google Scholar] [CrossRef]
- Fuchs, Y. Steller, H. Live to die another way: Modes of programmed cell death and the signals emanating from dying cells. Nat. Rev. Mol. Cell Biol. 2015, 16, 329–344. [Google Scholar] [CrossRef]
- Lukin, D.J.; Carvajal, L.A.; Liu, W.J.; Resnick-Silverman, L.; Manfredi, J.J. p53 Promotes cell survival due to the reversibility of its cell-cycle checkpoints. Mol. Cancer Res. 2015, 13, 16–28. [Google Scholar] [CrossRef] [Green Version]
- Vousden, K.H.; Prives, C. Blinded by the Light: The Growing Complexity of p53. Cell 2009, 137, 413–431. [Google Scholar] [CrossRef] [Green Version]
- Visconti, R.; Della Monica, R.; Grieco, D. Cell cycle checkpoint in cancer: A therapeutically targetable double-edged sword. J. Exp. Clin. Cancer Res. 2016, 35, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Du, H.; Geng, G.; Zhou, H.; Xu, M.; Cao, H.; Zhang, B.; Song, G.; Hu, T. Matrine inhibits proliferation and induces apoptosis via BID-mediated mitochondrial pathway in esophageal cancer cells. Mol. Biol Rep. 2014, 41, 3009–3020. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Li, S.; Chen, Z.; Lu, Z.; Gao, J.; Zou, J.; Lin, X.; Li, Y.; Zhang, C.; Shen, L. A novel oral camptothecin analog, gimatecan, exhibits superior antitumor efficacy than irinotecan toward esophageal squamous cell carcinoma in vitro and in vivo. Cell Death Dis. 2018, 9, 661. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Zhang, R.; Feng, C.; Zhang, J.; Liu, D.; Xu, K.; Wang, X.; Zhang, S.; Li, Z.; Liu, X.; et al. Diallyl disulfide induces G2/M arrest and promotes apoptosis through the p53/p21 and MEK-ERK pathways in human esophageal squamous cell carcinoma. Oncol. Rep. 2014, 32, 1748–1756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Fiandalo, M.V.; Kyprianou, N. Caspase control: Protagonists of cancer cell apoptosis. Exp. Oncol. 2012, 4, 165–175. [Google Scholar]
- Yang, X.W.; Zhang, H.; Li, M.; Du, L.J.; Yang, Z.; Xiao, S.Y. Studies on the alkaloid constituents of Evodia rutaecarpa (Juss) Benth var. bodinaieri (Dode) Huang and their acute toxicity in mice. J. Asian Nat. Prod. Res. 2006, 8, 697–703. [Google Scholar] [CrossRef]
- Cantor, J.P.; Iliopoulos, D.; Rao, A.S.; Druck, T.; Semba, S.; Han, S.Y.; McCorkell, K.A.; Lakshman, T.V.; Collins, J.E.; Wachsberger, P.; et al. Epigenetic modulation of endogenous tumor suppressor expression in lung cancer xenografts suppresses tumorigenicity. Int. J. Cancer 2007, 120, 24–31. [Google Scholar] [CrossRef]
- Jeon, T.W.; Jin, C.H.; Lee, S.K.; Jun, I.H.; Kim, G.H.; Lee, D.J.; Jeong, H.G.; Lee, K.B.; Jahng, Y.; Jeong, T.C. Immunosuppressive effects of rutaecarpine in female BALB/c mice. Toxicol. Lett. 2006, 164, 155–166. [Google Scholar] [CrossRef]
- Yeh, S.L.; Yeh, C.L.; Chan, S.T.; Chuang, C.H. Plasma rich in quercetin metabolites induces G2/M arrest by upregulating PPAR-γ expression in human A549 lung cancer cells. Planta Med. 2011, 77, 992–998. [Google Scholar] [CrossRef]
- Chuang, C.H.; Chan, S.T.; Chen, C.H.; Yeh, S.L. Quercetin enhances the antitumor activity of trichostatin A through up-regulation of p300 protein expression in p53 null cancer cells. Chem. Biol. Interact. 2019, 306, 54–61. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | RBC (×106/µL) | Hb (g/dL) | HCT (%) | MCH (Pg) | MCHC (%) | BUN (mg/dL) | Cre (mg/dL) |
|---|---|---|---|---|---|---|---|
| CTR 2 | 6.68 ± 0.6 a,3 | 11.5 ± 0.7 a | 32.6 ± 2.6 a | 17.2 ± 0.4 a | 35.2 ± 0.8 a3 | 32.9 ± 2.7 c | 0.337 ± 0.012 b |
| TC | 7.26 ± 0.2 a | 12.1 ± 0.4 a | 34.7 ± 0.7 a | 16.6 ± 0.4 a | 34.8 ± 1.2 a | 36.3 ± 0.5 b | 0.345 ± 0.005 b |
| RTP-L | 6.34 ± 0.9 a | 12.0 ± 0.8 a | 32.4 ± 0.6 a | 17.9 ± 1.4 a | 35.6 ± 0.5 a | 34.2 ± 1.3b c | 0.333 ± 0.006 b |
| RTP-H | 7.46 ± 0.6 a | 12.1 ± 0.4 a | 33.3 ± 2.0 a | 18.1 ± 0.5 a | 36.0 ± 1.0 a | 34.9 ± 0.9b c | 0.333 ± 0.012 b |
| CDDP | 6.65 ± 0.4 a | 11.2 ± 0.4 a | 33.9 ± 2.9 a | 16.9 ± 0.4 a | 34.1 ± 1.3 b | 40.7 ± 2.0 a | 0.447 ± 0.006 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.-Y.; Yeh, S.-L.; Hsu, S.-T.; Chen, C.-H.; Chen, C.-C.; Chuang, C.-H. The Anti-Proliferative and Apoptotic Effects of Rutaecarpine on Human Esophageal Squamous Cell Carcinoma Cell Line CE81T/VGH In Vitro and In Vivo. Int. J. Mol. Sci. 2022, 23, 2843. https://doi.org/10.3390/ijms23052843
Wang L-Y, Yeh S-L, Hsu S-T, Chen C-H, Chen C-C, Chuang C-H. The Anti-Proliferative and Apoptotic Effects of Rutaecarpine on Human Esophageal Squamous Cell Carcinoma Cell Line CE81T/VGH In Vitro and In Vivo. International Journal of Molecular Sciences. 2022; 23(5):2843. https://doi.org/10.3390/ijms23052843
Chicago/Turabian StyleWang, Li-Yu, Shu-Lan Yeh, Shih-Tien Hsu, Chao-Hsiang Chen, Chien-Chih Chen, and Cheng-Hung Chuang. 2022. "The Anti-Proliferative and Apoptotic Effects of Rutaecarpine on Human Esophageal Squamous Cell Carcinoma Cell Line CE81T/VGH In Vitro and In Vivo" International Journal of Molecular Sciences 23, no. 5: 2843. https://doi.org/10.3390/ijms23052843
APA StyleWang, L. -Y., Yeh, S. -L., Hsu, S. -T., Chen, C. -H., Chen, C. -C., & Chuang, C. -H. (2022). The Anti-Proliferative and Apoptotic Effects of Rutaecarpine on Human Esophageal Squamous Cell Carcinoma Cell Line CE81T/VGH In Vitro and In Vivo. International Journal of Molecular Sciences, 23(5), 2843. https://doi.org/10.3390/ijms23052843