
Differences in Transporters Rather than Drug Targets Are the Principal Determinants of the Different Innate Sensitivities of Trypanosoma congolense and Trypanozoon Subgenus Trypanosomes to Diamidines and Melaminophenyl Arsenicals

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
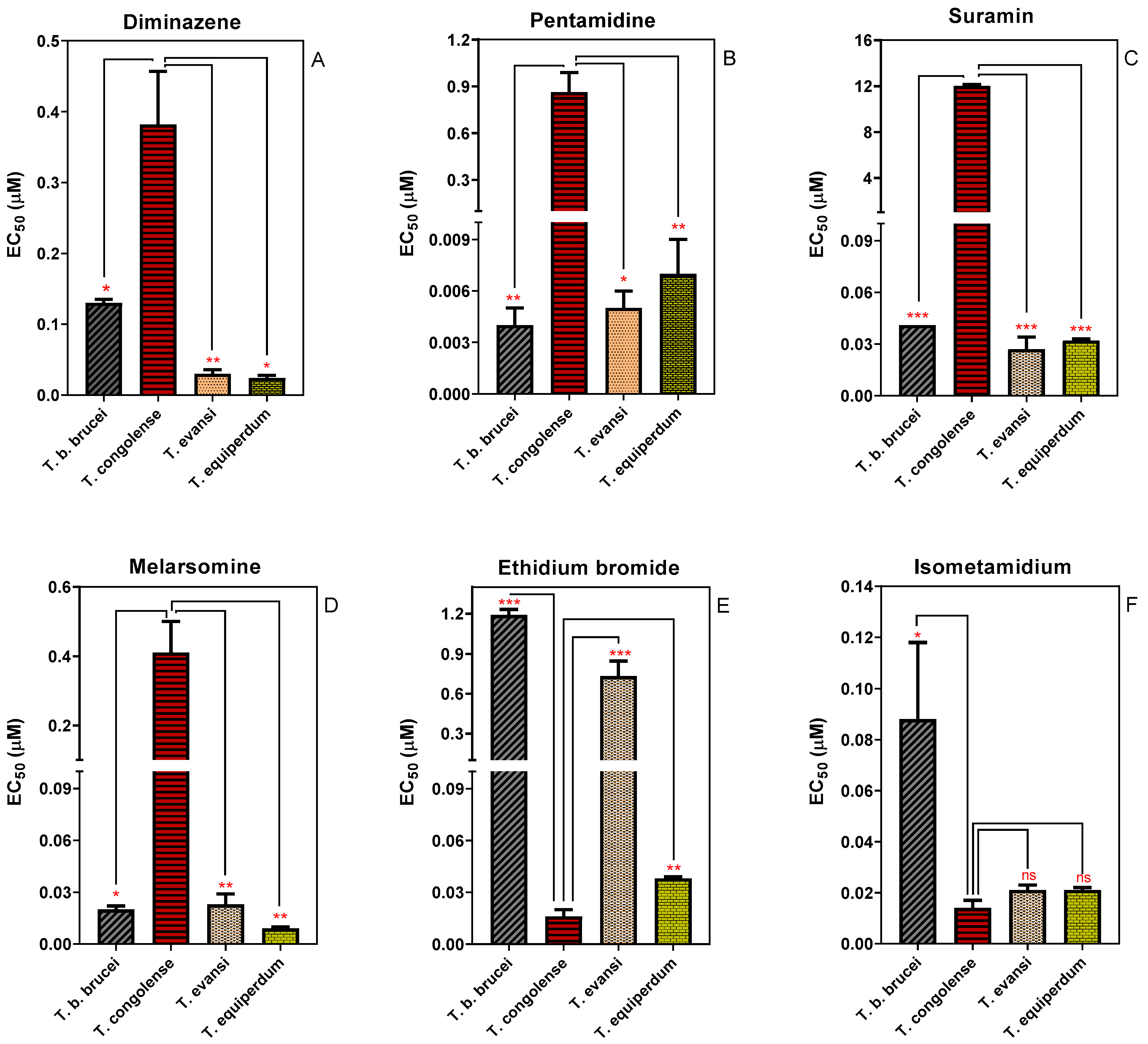
2.1. Sensitivity of Different Trypanosoma Species to Standard Trypanocides
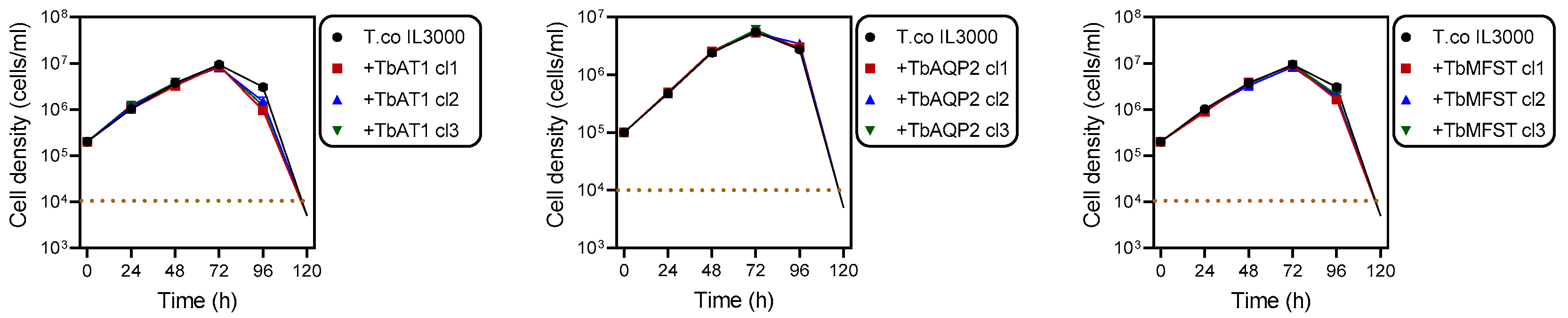
2.2. Cloning of T. brucei Drug Transporters and Their Expression in T. congolense
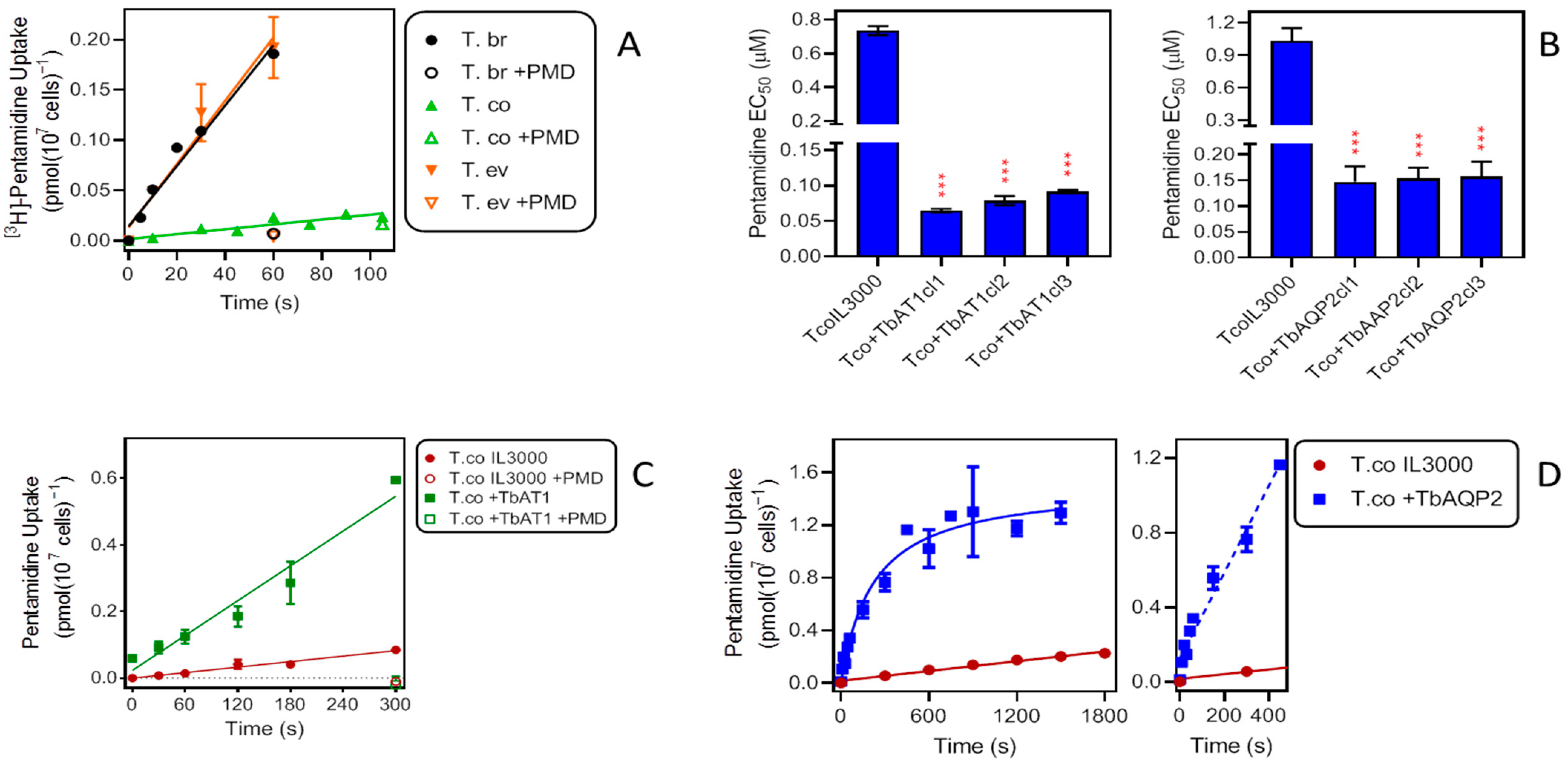
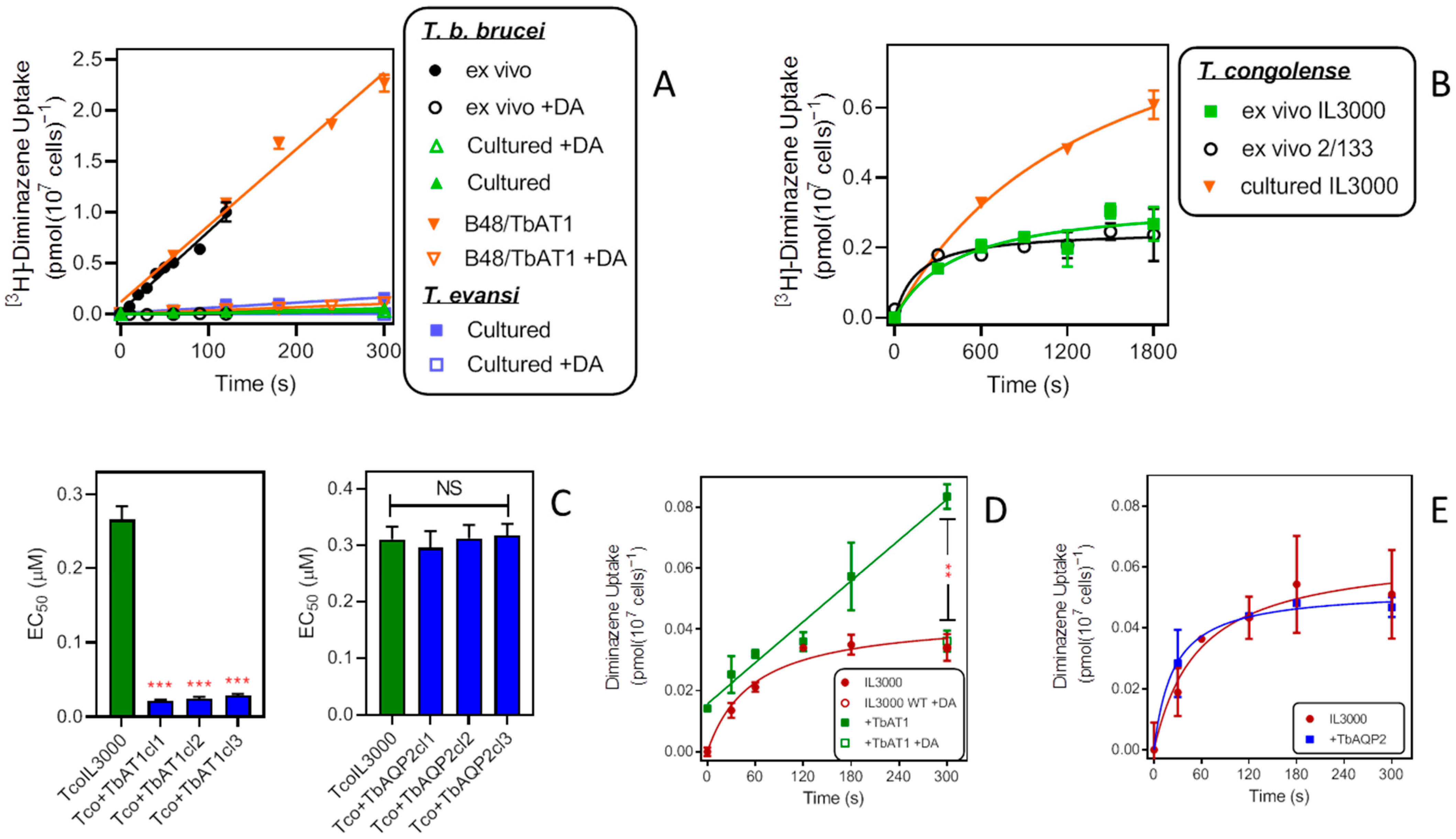
2.3. Uptake of Drugs in Different Trypanosome Species—Influence of TbAT1 and TbAQP2
2.3.1. Pentamidine
2.3.2. Diminazene Aceturate (DA)
2.3.3. Suramin
2.3.4. Melarsomine (Cymelarsan; MelCy)
3. Materials and Methods
3.1. Parasites and Cultures
3.2. Resazurin-Based Drug Sensitivity Assay
3.3. Growth Rate by Cell Count
3.4. Cell Absorbance Assay
3.5. Immunofluorescence Microscopy
3.6. Drug Uptake Assay
3.7. Genetic Manipulation of T. congolense
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Giordani, F.; Morrison, L.J.; Rowan, T.G.; De Koning, H.P.; Barrett, M.P. The animal trypanosomiases and their chemotherapy: A review. Parasitology 2016, 143, 1862–1889. [Google Scholar] [CrossRef] [PubMed]
- Büscher, P.; Gonzatti, M.I.; Hébert, L.; Inoue, N.; Pascucci, I.; Schnaufer, A.; Suganuma, K.; Touratier, L.; Van Reet, N. Equine trypanosomosis: Enigmas and diagnostic challenges. Parasit. Vectors 2019, 12, 234. [Google Scholar] [CrossRef] [PubMed]
- Osório, A.L.; Madruga, C.R.; Desquesnes, M.; Soares, C.O.; Ribeiro, L.R.; Costa, S.C. Trypanosoma (Duttonella) vivax: Its biology, epidemiology, pathogenesis, and introduction in the New World—A review. Mem. Inst. Oswaldo Cruz 2008, 103, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzatti, M.I.; Gonzalez-Baradat, B.; Aso, P.M.; Reyna-Bello, A. Trypanosoma (Duttonella) vivax and trypanosomosis in Latin America: Secadera/huequera/cacho hueco. In Trypanosomes and Trypanosomiasis; Magez, S., Radwanska, M., Eds.; Springer: Wien, Austria, 2014; pp. 261–285. [Google Scholar]
- Fetene, E.; Leta, S.; Regassa, F.; Büscher, P. Global distribution, host range and prevalence of Trypanosoma vivax: A systematic review and meta-analysis. Parasit. Vectors 2021, 14, 80–100. [Google Scholar] [CrossRef]
- Asgari, M.M.; Rassouli, M. First identification of Trypanosoma vivax among camels (Camelus dromedarius) in Yazd, central Iran, jointly with Trypanosoma evansi. Parasitol. Int. 2022, 86, 102450. [Google Scholar] [CrossRef]
- Desquesnes, M. Livestock Trypanosomoses and Their Vectors in Latin America; CIRAD-EMVT Publication, OIE: Paris, France, 2004; Available online: https://www.proquest.com/docview/214620829 (accessed on 2 March 2022).
- Desquesnes, M.; Dargantes, A.; Lai, D.-H.; Lun, Z.-R.; Holzmuller, P.; Jittapalapong, S. Trypanosoma evansi and surra: A review and perspectives on transmission, epidemiology and control, impact, and zoonotic aspects. BioMed Res. Int. 2013, 2013, 321237. [Google Scholar] [CrossRef] [Green Version]
- Elata, A.; Galon, E.M.; Moumouni, P.F.A.; Ybanez, R.H.D.; Mossaad, E.; Salces, C.B.; Bajenting, G.P.; Ybanez, A.P.; Xuan, X.; Inoue, N.; et al. First molecular detection and identification of Trypanosoma evansi in goats from Cebu, Philippines using a PCR-based assay. Vet. Parasitol. Reg. Stud. Rep. 2020, 21, 100414. [Google Scholar] [CrossRef]
- Setiawan, A.; Nurcahyo, W.; Priyowidodo, D.; Budiati, R.T.; Susanti, D.S.R. Genetic and parasitological identification of Trypanosoma evansi infecting cattle in South Sulawesi, Indonesia. Vet. World 2021, 14, 113–119. [Google Scholar] [CrossRef]
- Angara, T.E.; Ismail, A.; Ibrahim, A. An overview on the economic impacts of animal trypanosomiasis. Glob. J. Res. Anal. 2012, 3, 275–276. [Google Scholar] [CrossRef] [Green Version]
- De Koning, H.P. The drugs of sleeping sickness: Their mechanisms of action and resistance, and a brief history. Trop. Med. Infect. Dis. 2020, 5, 14. [Google Scholar] [CrossRef] [Green Version]
- Kinabo, L.D.B. Pharmacology of existing drugs for animal trypanosomiasis. Acta Trop. 1993, 54, 169–183. [Google Scholar] [CrossRef]
- Williamson, J. Review of chemotherapeutic and chemoprophylactic agents. In The African Trypanosomiases; Mulligan, H.W., Ed.; George Allen and Unwin, Ltd.: London, UK, 1970; pp. 125–221. [Google Scholar]
- Delespaux, V.; De Koning, H.P. Transporters in antiparasitic drug development and resistance. In Trypanosomatid Diseases: Molecular Routes to Drug Discovery; Jäger, T., Koch, O., Flohe, L., Eds.; Wiley-Blackwell: Weinheim, Germany, 2013; pp. 335–349. [Google Scholar]
- Carter, N.S.; Berger, B.J.; Fairlamb, A.H. Uptake of diamidine drugs by the P2 nucleoside transporter in melarsen-sensitive and -resistant Trypanosoma brucei brucei. J. Biol. Chem. 1995, 270, 28153–28157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Koning, H.P.; Jarvis, S.M. Uptake of pentamidine in Trypanosoma brucei brucei is mediated by the P2 adenosine transporter and at least one novel, unrelated transporter. Acta Trop. 2001, 80, 245–250. [Google Scholar] [CrossRef]
- Bridges, D.; Gould, M.K.; Nerima, B.; Mäser, P.; Burchmore, R.J.S.; De Koning, H.P. Loss of the High Affinity Pentamidine Transporter is responsible for high levels of cross-resistance between arsenical and diamidine drugs in African Trypanosomes. Mol. Pharmacol. 2007, 71, 1098–1108. [Google Scholar] [CrossRef] [Green Version]
- Baker, N.; Glover, L.; Munday, J.C.; Aguinaga Andrés, D.; Barrett, M.P.; De Koning, H.P.; Horn, D. Aquaglyceroporin 2 controls susceptibility to melarsoprol and pentamidine in African Trypanosomes. Proc. Natl. Acad. Sci. USA 2012, 109, 10996–11001. [Google Scholar] [CrossRef] [Green Version]
- Alghamdi, A.H.; Munday, J.C.; Campagnaro, G.D.; Gurvic, D.; Svensson, F.; Okpara, C.E.; Kumar, A.; Quintana, J.; Martin Abril, M.E.; Milić, P.; et al. Positively selected modifications in the pore of TbAQP2 allow pentamidine to enter Trypanosoma brucei. eLife 2020, 9, e56416. [Google Scholar] [CrossRef]
- Munday, J.C.; Eze, A.A.; Baker, N.; Glover, L.; Clucas, C.; Aguinaga Andrés, D.; Natto, M.J.; Teka, I.A.; McDonald, J.; Lee, R.S.; et al. Trypanosoma brucei Aquaglyceroporin 2 is a high affinity transporter for pentamidine and melaminophenyl arsenic drugs and is the main genetic determinant of resistance to these drugs. J. Antimicrob. Chemother. 2014, 69, 651–663. [Google Scholar] [CrossRef] [PubMed]
- De Koning, H.P.; Anderson, L.F.; Stewart, M.; Burchmore, R.J.S.; Wallace, L.J.M.; Barrett, M.P. The trypanocide diminazene aceturate is accumulated predominantly through the TbAT1 purine transporter; additional insights in diamidine resistance in African Trypanosomes. Antimicrob. Agents Chemother. 2004, 48, 1515–1519. [Google Scholar] [CrossRef] [Green Version]
- Ward, C.P.; Wong, P.E.; Burchmore, R.J.; De Koning, H.P.; Barrett, M.P. Trypanocidal furamidine analogues: Influence of pyridine nitrogens on trypanocidal activity, transport kinetics and resistance patterns. Antimicrob. Agents Chemother. 2011, 55, 2352–2361. [Google Scholar] [CrossRef] [Green Version]
- Carter, N.S.; Fairlamb, A.H. Arsenical-resistant Trypanosomes lack an unusual adenosine transporter. Nature 1993, 361, 173–176. [Google Scholar] [CrossRef]
- Matovu, E.; Stewart, M.; Geiser, F.; Brun, R.; Mäser, P.; Wallace, L.J.M.; Burchmore, R.J.; Enyaru, J.C.K.; Barrett, M.P.; Kaminsky, R.; et al. The mechanisms of arsenical and diamidine uptake and resistance in Trypanosoma brucei. Eukaryot. Cell 2003, 2, 1003–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vincent, I.M.; Creek, D.; Watson, D.G.; Kamleh, M.A.; Woods, D.J.; Wong, P.E.; Burchmore, R.J.; Barrett, M.P. A molecular mechanism for eflornithine resistance in African Trypanosomes. PLoS Pathog. 2010, 6, e1001204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schumann Burkard, G.; Jutzi, P.; Roditi, I. Genome-wide RNAi screens in bloodstream form Trypanosomes identify drug transporters. Mol. Biochem. Parasitol. 2011, 175, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Alsford, S.; Eckert, S.; Baker, N.; Glover, L.; Sanchez-Flores, A.; Leung, K.F.; Turner, D.J.; Field, M.C.; Berriman, M.; Horn, D. High-throughput decoding of antitrypanosomal drug efficacy and resistance. Nature 2012, 482, 232–236. [Google Scholar] [CrossRef] [Green Version]
- Zoltner, M.; Horn, D.; De Koning, H.P.; Field, M.C. Exploiting the Achilles’ heel of membrane trafficking in Trypanosomes. Curr. Opin. Microbiol. 2016, 34, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Zoltner, M.; Campagnaro, G.D.; Taleva, G.; Burrell, A.; Cerone, M.; Leung, K.-F.; Achcar, F.; Horn, D.; Vaughan, S.; Gadelha, C.; et al. Suramin exposure alters cellular metabolism and mitochondrial energy production in African Trypanosomes. J. Biol. Chem. 2020, 295, 8331–8347. [Google Scholar] [CrossRef]
- Wilkes, J.M.; Mulugeta, W.; Wells, C.; Peregrine, A.S. Modulation of mitochondrial electrical potential: A candidate mechanism for drug resistance in African Trypanosomes. Biochem. J. 1997, 326, 755–761. [Google Scholar] [CrossRef] [Green Version]
- Eze, A.A.; Gould, M.K.; Munday, J.C.; Tagoe, D.N.A.; Stelmanis, V.; Schnaufer, A.; De Koning, H.P. Loss of mitochondrial membrane potential is a late adaptation of Trypanosoma brucei brucei to isometamidium preceded by mutations in the γ subunit of the F1F0-ATPase. PLoS Negl. Trop. Dis. 2016, 10, e0004791. [Google Scholar] [CrossRef] [Green Version]
- Kell, D.B. The transporter-mediated cellular uptake and efflux of pharmaceutical drugs and biotechnology products: How and why phospholipid bilayer transport Is negligible in real biomembranes. Molecules 2021, 26, 5629. [Google Scholar] [CrossRef]
- Jeganathan, S.; Sanderson, L.; Dogruel, M.; Rodgers, J.; Croft, S.; Thomas, S.A. The distribution of nifurtimox across the healthy and trypanosome-infected murine blood–brain and blood-CSF barriers. J. Pharmacol. Exp. Ther. 2011, 336, 506–515. [Google Scholar] [CrossRef] [Green Version]
- Watson, C.P.; Dogruel, M.; Mihoreanu, L.; Begley, D.J.; Weksler, B.B.; Couraud, P.O.; Romero, I.A.; Thomas, S.A. The transport of nifurtimox, an anti-trypanosomal drug, in an in vitro model of the human blood-brain barrier: Evidence for involvement of breast cancer resistance protein. Brain Res. 2012, 1436, 111–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyllie, S.; Foth, B.J.; Kelner, A.; Sokolova, A.Y.; Berriman, M.; Fairlamb, A.H. Nitroheterocyclic drug resistance mechanisms in Trypanosoma brucei. J. Antimicrob. Chemother. 2016, 71, 625–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Changtam, C.; De Koning, H.P.; Ibrahim, H.; Sajid, S.; Gould, M.K.; Suksamrarn, A. Curcuminoid analogues with potent activity against Trypanosoma and Leishmania species. Eur. J. Med. Chem. 2010, 45, 941–956. [Google Scholar] [CrossRef] [PubMed]
- Stewart, M.L.; Burchmore, R.J.S.; Clucas, C.; Hertz-Fowler, C.; Brook, K.; Tait, A.; McLeod, A.; Turner, C.M.R.; De Koning, H.P.; Wong, P.E.; et al. Multiple genetic mechanisms lead to the loss of functional TbAT1 expression in drug resistant Trypanosomes. Eukaryot. Cell 2010, 9, 336–343. [Google Scholar] [CrossRef] [Green Version]
- Graf, F.E.; Ludin, P.; Wenzler, T.; Kaiser, M.; Brun, R.; Pati Pyana, P.; Büscher, P.; De Koning, H.P.; Horn, D.; Mäser, P. Aquaporin 2 mutations in Trypanosoma b. gambiense field isolates correlate with decreased susceptibility to pentamidine and melarsoprol. PLoS Negl. Trop. Dis. 2013, 7, e2475. [Google Scholar] [CrossRef] [Green Version]
- Fueyo González, F.J.; Ebiloma, G.U.; Izquierdo García, C.; Bruggeman, V.; Sánchez Villamañán, J.M.; Donachie, A.; Balogun, E.O.; Inaoka, D.K.; Shiba, T.; Harada, S.; et al. Conjugates of 2,4-dihydroxybenzoate and salicylhydroxamate and lipocations display potent anti-parasite effects by efficiently targeting the Trypanosoma brucei and Trypanosoma congolense mitochondrion. J. Med. Chem. 2017, 60, 1509–1522. [Google Scholar] [CrossRef] [Green Version]
- Munday, J.C.; Settimo, L.; De Koning, H.P. Transport proteins determine drug sensitivity and resistance in a protozoan parasite, Trypanosoma brucei. Front. Pharmacol. 2015, 6, 32. [Google Scholar] [CrossRef]
- Carruthers, L.V.; Munday, J.C.; Ebiloma, G.U.; Steketee, P.; Jayaraman, S.; Campagnaro, G.D.; Ungogo, M.A.; Donnachie, A.; Lemgruber, L.; Rowan, T.G.; et al. Diminazene resistance in Trypanosoma congolense is not caused by reduced transport capacity but associated with reduced mitochondrial membrane potential. Mol. Microbiol. 2021, 116, 564–588. [Google Scholar] [CrossRef]
- Witola, W.H.; Inoue, N.; Ohashi, K.; Onuma, M. RNA-interference silencing of the adenosine transporter-1 gene in Trypanosoma evansi confers resistance to diminazene aceturate. Exp. Parasitol. 2004, 107, 47–57. [Google Scholar] [CrossRef]
- Barrett, M.P.; Zhang, Z.Q.; Denise, H.; Baltz, T. A diamidine-resistant Trypanosoma equiperdum clone contains a P2 purine transporter with reduced substrate affinity. Mol. Biochem. Parasitol. 1995, 73, 223–229. [Google Scholar] [CrossRef]
- Büscher, P.; Van Reet, N.; Institute of Tropical Medicine, Antwerp, Belgium. Personal communication, 2021.
- De Koning, H.P. Uptake of pentamidine in Trypanosoma brucei brucei is mediated by three distinct transporters. Implications for crossresistance with arsenicals. Mol. Pharmacol. 2001, 59, 586–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munday, J.C.; Rojas López, K.E.; Eze, A.A.; Delespaux, V.; Van Den Abbeele, J.; Rowan, T.; Barrett, M.P.; Morrison, L.J.; De Koning, H.P. Functional expression of TcoAT1 reveals it to be a P1-type nucleoside transporter with no capacity for diminazene uptake. Int. J. Parasitol. Drugs Drug Resist. 2013, 3, 69–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delespaux, V.; De Koning, H.P. Drugs and drug resistance in African trypanosomiasis. Drug Resist. Updat. 2007, 10, 30–50. [Google Scholar] [CrossRef] [PubMed]
- De Koning, H.P.; MacLeod, A.; Barrett, M.P.; Cover, B.; Jarvis, S.M. Further evidence for a link between melarsoprol resistance and P2 transporter function in African Trypanosomes. Mol. Biochem. Parasitol. 2000, 106, 181–185. [Google Scholar] [CrossRef]
- Fairlamb, A.H.; Carter, N.S.; Cunningham, M.; Smith, K. Characterisation of melarsen-resistant Trypanosoma brucei brucei with respect to cross-resistance to other drugs and trypanothione metabolism. Mol. Biochem. Parasitol. 1992, 53, 213–222. [Google Scholar] [CrossRef]
- Dean, S.; Gould, M.K.; Dewar, C.E.; Schnaufer, A.C. Single point mutations in ATP synthase compensate for mitochondrial genome loss in Trypanosomes. Proc. Natl. Acad. Sci. USA 2013, 110, 14741–14746. [Google Scholar] [CrossRef] [Green Version]
- Coustou, V.; Guegan, F.; Plazolles, N.; Baltz, T. Complete in vitro life cycle of Trypanosoma congolense: Development of genetic tools. PLoS Negl. Trop. Dis. 2010, 4, e618. [Google Scholar] [CrossRef]
- Giordani, F.; Khalaf, A.I.; Gillingwater, K.; Munday, J.C.; De Koning, H.P.; Suckling, C.J.; Barrett, M.P.; Scott, F.J. Novel minor groove binders cure animal African trypanosomiasis in an in vivo mouse model. J. Med. Chem. 2019, 62, 3021–3035. [Google Scholar] [CrossRef] [Green Version]
- Gould, M.K.; Vu, X.L.; Seebeck, T.; De Koning, H.P. Propidium iodide-based methods for monitoring drug action in the kinetoplastidae: Comparison with the Alamar Blue assay. Anal. Biochem. 2008, 382, 87–93. [Google Scholar] [CrossRef]
- Stortz, J.A.; Serafim, T.D.; Alsford, S.; Wilkes, J.; Fernandez-Cortes, F.; Hamilton, G.; Briggs, E.; Lemgruber, L.; Horn, D.; Mottram, J.C.; et al. Genome-wide and protein kinase-focused RNAi screens reveal conserved and novel damage response pathways in Trypanosoma brucei. PLoS Pathog. 2017, 13, e1006477. [Google Scholar] [CrossRef] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Campagnaro, G.D.; de Freitas Nascimento, J.; Girard, R.B.M.; Silber, A.M.; De Koning, H.P. Cloning and characterisation of the Equilibrative Nucleoside Transporter family of Trypanosoma cruzi: Ultra-high affinity and selectivity to survive in the intracellular niche. Biochim. Biophys. Acta Gen. Subj. 2018, 12, 2750–2763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proudfoot, C.; McCulloch, R. Distinct roles for two RAD51-related genes in Trypanosoma brucei antigenic variation. Nucleic Acids Res. 2005, 33, 6906–6919. [Google Scholar] [CrossRef] [PubMed]
- Giordani, F.; Paape, D.; Vincent, I.M.; Pountain, A.W.; Fernández-Cortés, F.; Rico, E.; Zhang, N.; Morrison, L.J.; Freund, Y.; Witty, M.J.; et al. Veterinary trypanocidal benzoxaboroles are peptidase-activated prodrugs. PLoS Pathog. 2020, 16, e1008932. [Google Scholar] [CrossRef] [PubMed]
- Awuah-Mensah, G.; McDonald, J.; Steketee, P.C.; Autheman, D.; Whipple, S.; D’Archivio, S.; Brandt, C.; Clare, S.; Harcourt, K.; Wright, G.J.; et al. Reliable, scalable functional genetics in bloodstream-form Trypanosoma congolense in vitro and in vivo. PLoS Pathog. 2021, 17, e1009224. [Google Scholar] [CrossRef]
- Campagnaro, G.D.; De Koning, H.P. Purine and pyrimidine transporters of pathogenic protozoa—Conduits for therapeutic agents. Med. Res. Rev. 2020, 40, 1679–1714. [Google Scholar] [CrossRef]
- Munday, J.C.; Tagoe, D.N.A.; Eze, A.A.; Krezdorn, J.A.; Rojas López, K.E.; Alkhaldi, A.A.M.; McDonald, F.; Still, J.; Alzahrani, K.J.; Settimo, L.; et al. Functional analysis of drug resistance-associated mutations in the Trypanosoma brucei adenosine transporter 1 (TbAT1) and the proposal of a structural model for the protein. Mol. Microbiol. 2015, 96, 887–900. [Google Scholar] [CrossRef]
- Jackson, A.P.; Allison, H.C.; Barry, J.D.; Field, M.C.; Hertz-Fowler, C.; Berriman, M. A Cell-surface phylome for African Trypanosomes. PLOS Negl. Trop. Dis. 2013, 7, e2121. [Google Scholar] [CrossRef]
- Jeacock, L.; Baker, N.; Wiedemar, N.; Mäser, P.; Horn, D. Aquaglyceroporin-null Trypanosomes display glycerol transport defects and respiratory-inhibitor sensitivity. PLoS Pathog. 2017, 13, e1006307. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
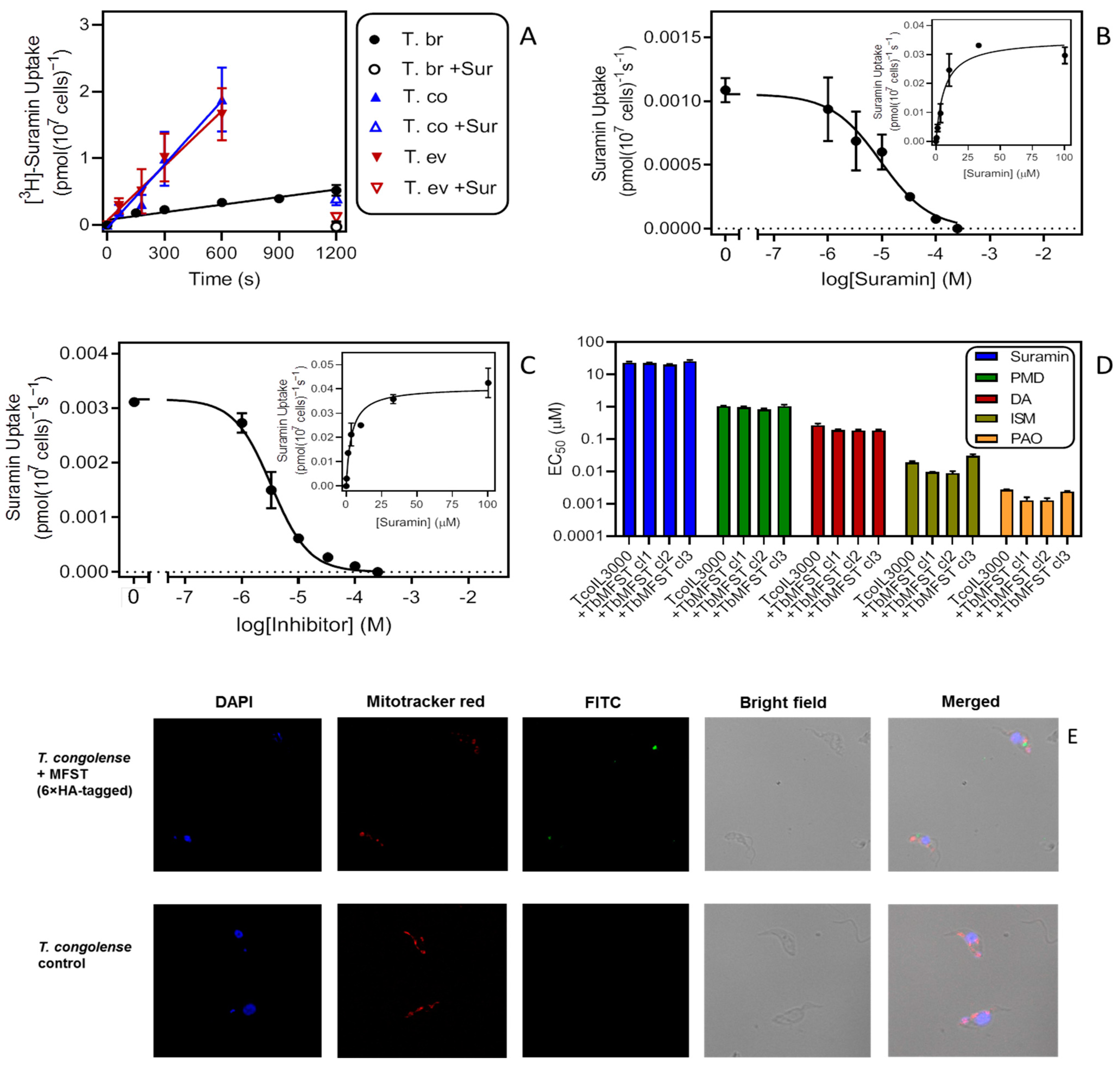{kind=link}
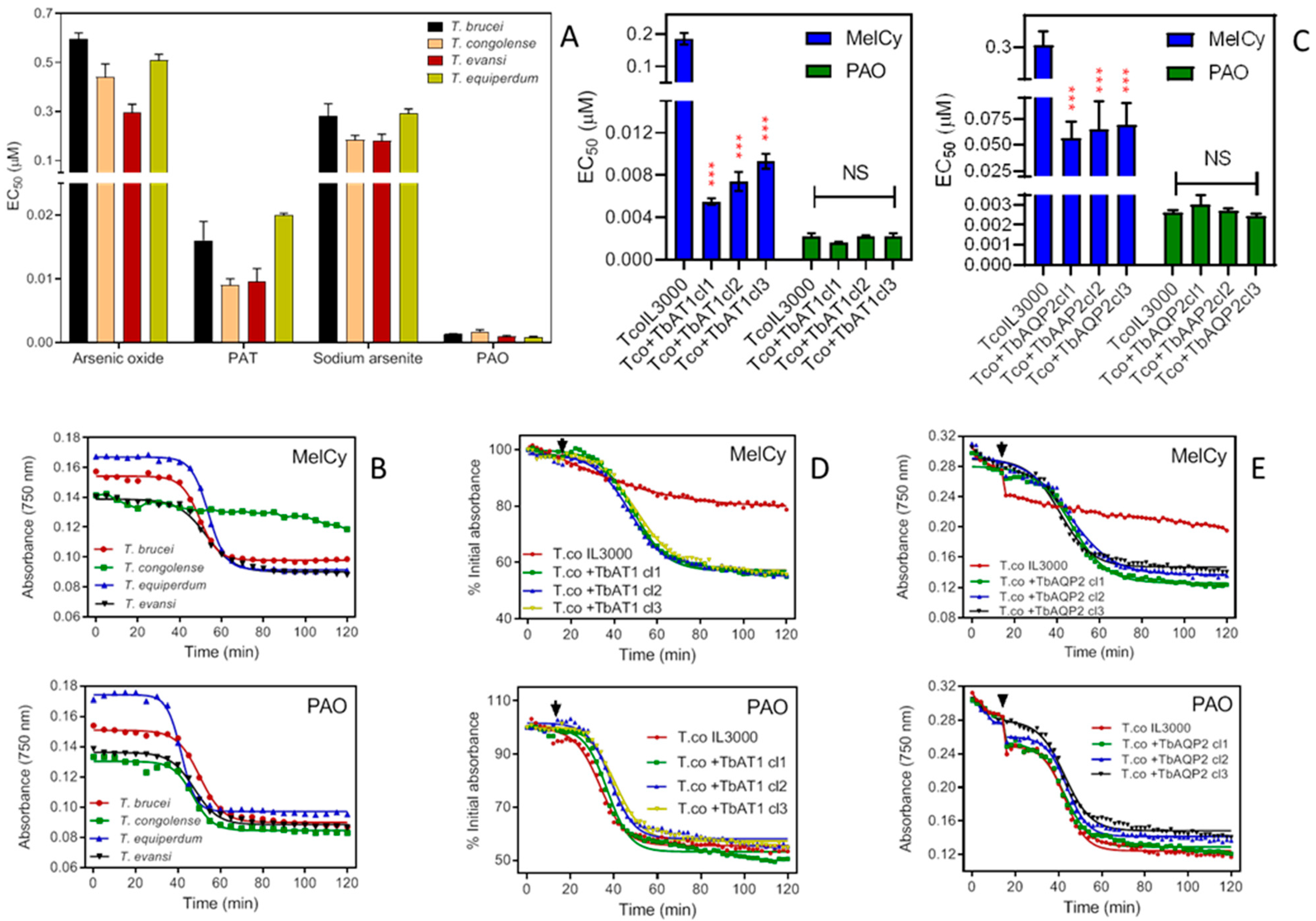{kind=link}
| Species | Gene ID | T. brucei | ||
|---|---|---|---|---|
| AT1 | ||||
| Tb927.5.286b | ||||
| T. evansi | AGT37292.1 | 99.35 | ||
| T. equiperdum | SCU70586.1 | 99.78 | ||
| T. congolense | - | <75 | ||
| AQP2 | ||||
| Tb927.10.14170 | ||||
| T. evansi | TevSTIB805.10.14910 | 100 | ||
| T. equiperdum | not annotated | 100 1 | ||
| T. congolense | - | <75 | ||
| MFST | ||||
| Tb927.9.6360 2 | Tb927.9.6370 | Tb927.9.6380 | ||
| T. evansi | TevSTIB805.9.4540 | 99.58 | 89.15 | 96.87 |
| TevSTIB805.9.4550 | 89.79 | 98.30 | 90.43 | |
| TevSTIB805.9.4560 | 96.24 | 88.72 | 98.96 | |
| T. equiperdum | RHW70022.1 | 96.24 | 88.72 | 98.96 |
| RHW70658.1 | 88.30 | 97.23 | 90.21 | |
| RHW70215.1 | 96.45 | 88.94 | 99.16 | |
| T. congolense | - | <75 | <75 | <75 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ungogo, M.A.; Campagnaro, G.D.; Alghamdi, A.H.; Natto, M.J.; de Koning, H.P. Differences in Transporters Rather than Drug Targets Are the Principal Determinants of the Different Innate Sensitivities of Trypanosoma congolense and Trypanozoon Subgenus Trypanosomes to Diamidines and Melaminophenyl Arsenicals. Int. J. Mol. Sci. 2022, 23, 2844. https://doi.org/10.3390/ijms23052844
Ungogo MA, Campagnaro GD, Alghamdi AH, Natto MJ, de Koning HP. Differences in Transporters Rather than Drug Targets Are the Principal Determinants of the Different Innate Sensitivities of Trypanosoma congolense and Trypanozoon Subgenus Trypanosomes to Diamidines and Melaminophenyl Arsenicals. International Journal of Molecular Sciences. 2022; 23(5):2844. https://doi.org/10.3390/ijms23052844
Chicago/Turabian StyleUngogo, Marzuq A., Gustavo D. Campagnaro, Ali H. Alghamdi, Manal J. Natto, and Harry P. de Koning. 2022. "Differences in Transporters Rather than Drug Targets Are the Principal Determinants of the Different Innate Sensitivities of Trypanosoma congolense and Trypanozoon Subgenus Trypanosomes to Diamidines and Melaminophenyl Arsenicals" International Journal of Molecular Sciences 23, no. 5: 2844. https://doi.org/10.3390/ijms23052844
APA StyleUngogo, M. A., Campagnaro, G. D., Alghamdi, A. H., Natto, M. J., & de Koning, H. P. (2022). Differences in Transporters Rather than Drug Targets Are the Principal Determinants of the Different Innate Sensitivities of Trypanosoma congolense and Trypanozoon Subgenus Trypanosomes to Diamidines and Melaminophenyl Arsenicals. International Journal of Molecular Sciences, 23(5), 2844. https://doi.org/10.3390/ijms23052844