
Propagation and Dissemination Strategies of Transmissible Spongiform Encephalopathy Agents in Mammalian Cells


 ,
,
Abstract
:1. Introduction
2. Cell Biology of Cellular PrP
3. Cellular Models for Prion Propagation
4. The Infection Process—The Uptake of Prions
5. Detection of Productive Infections
6. The Site of PrPSc Formation in Persistently Infected Cells
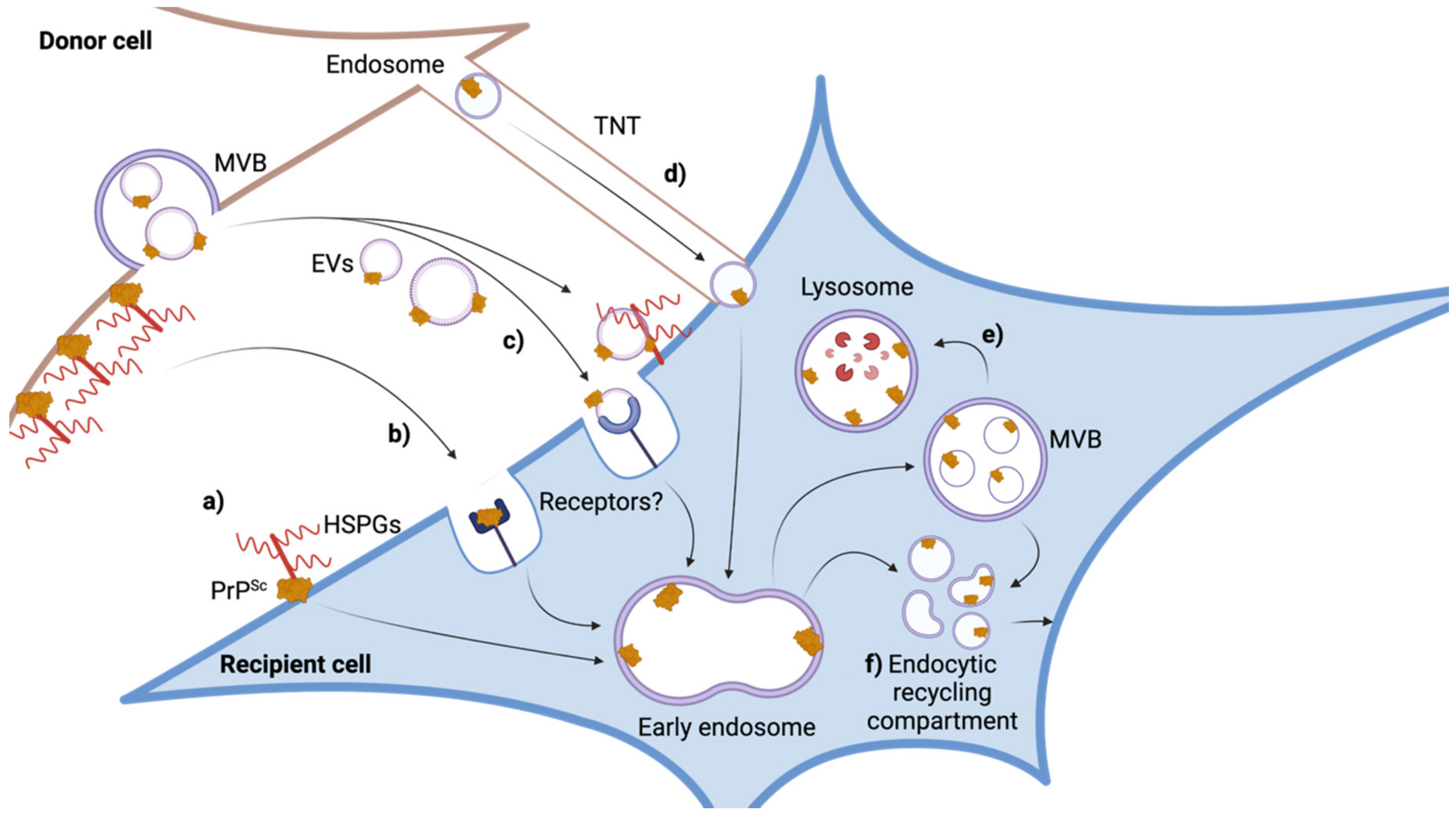
7. Intercellular Dissemination of Prions
8. Role of Viruses in Intercellular Prion Spreading
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Gallardo, M.J.; Delgado, F.O. Animal prion diseases: A review of intraspecies transmission. Open Vet. J. 2021, 11, 707–723. [Google Scholar] [CrossRef] [PubMed]
- Prusiner, S.B. Novel proteinaceous infectious particles cause scrapie. Science 1982, 216, 136–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oesch, B.; Westaway, D.; Walchli, M.; McKinley, M.P.; Kent, S.B.; Aebersold, R.; Barry, R.A.; Tempst, P.; Teplow, D.B.; Hood, L.E.; et al. A cellular gene encodes scrapie PrP 27-30 protein. Cell 1985, 40, 735–746. [Google Scholar] [CrossRef]
- Bessen, R.A.; Kocisko, D.A.; Raymond, G.J.; Nandan, S.; Lansbury, P.T.; Caughey, B. Non-genetic propagation of strain-specific properties of scrapie prion protein. Nature 1995, 375, 698–700. [Google Scholar] [CrossRef] [PubMed]
- Supattapone, S. Cofactor molecules: Essential partners for infectious prions. Prog. Mol. Biol. Transl. Sci. 2020, 175, 53–75. [Google Scholar] [CrossRef]
- Burke, C.M.; Walsh, D.J.; Mark, K.M.K.; Deleault, N.R.; Nishina, K.A.; Agrimi, U.; Di Bari, M.A.; Supattapone, S. Cofactor and glycosylation preferences for in vitro prion conversion are predominantly determined by strain conformation. PLoS Pathog. 2020, 16, e1008495. [Google Scholar] [CrossRef]
- Vella, L.J.; Greenwood, D.L.; Cappai, R.; Scheerlinck, J.P.; Hill, A.F. Enrichment of prion protein in exosomes derived from ovine cerebral spinal fluid. Vet. Immunol. Immunopathol. 2008, 124, 385–393. [Google Scholar] [CrossRef]
- Sunyach, C.; Jen, A.; Deng, J.; Fitzgerald, K.T.; Frobert, Y.; Grassi, J.; McCaffrey, M.W.; Morris, R. The mechanism of internalization of glycosylphosphatidylinositol-anchored prion protein. EMBO J. 2003, 22, 3591–3601. [Google Scholar] [CrossRef]
- Fehlinger, A.; Wolf, H.; Hossinger, A.; Duernberger, Y.; Pleschka, C.; Riemschoss, K.; Liu, S.; Bester, R.; Paulsen, L.; Priola, S.A.; et al. Prion strains depend on different endocytic routes for productive infection. Sci. Rep. 2017, 7, 6923. [Google Scholar] [CrossRef] [Green Version]
- Pineau, H.; Sim, V.L. From Cell Culture to Organoids-Model Systems for Investigating Prion Strain Characteristics. Biomolecules 2021, 11, 106. [Google Scholar] [CrossRef]
- Mouillet-Richard, S.; Nishida, N.; Pradines, E.; Laude, H.; Schneider, B.; Feraudet, C.; Grassi, J.; Launay, J.M.; Lehmann, S.; Kellermann, O. Prions impair bioaminergic functions through serotonin- or catecholamine-derived neurotoxins in neuronal cells. J. Biol. Chem. 2008, 283, 23782–23790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tahir, W.; Abdulrahman, B.; Abdelaziz, D.H.; Thapa, S.; Walia, R.; Schatzl, H.M. An astrocyte cell line that differentially propagates murine prions. J. Biol. Chem. 2020, 295, 11572–11583. [Google Scholar] [CrossRef] [PubMed]
- Marbiah, M.M.; Harvey, A.; West, B.T.; Louzolo, A.; Banerjee, P.; Alden, J.; Grigoriadis, A.; Hummerich, H.; Kan, H.M.; Cai, Y.; et al. Identification of a gene regulatory network associated with prion replication. EMBO J. 2014, 33, 1527–1547. [Google Scholar] [CrossRef] [PubMed]
- Vorberg, I.M. All the Same? The Secret Life of Prion Strains within Their Target Cells. Viruses 2019, 11, 334. [Google Scholar] [CrossRef] [Green Version]
- Marijanovic, Z.; Caputo, A.; Campana, V.; Zurzolo, C. Identification of an intracellular site of prion conversion. PLoS Pathog. 2009, 5, e1000426. [Google Scholar] [CrossRef] [Green Version]
- Borchelt, D.R.; Scott, M.; Taraboulos, A.; Stahl, N.; Prusiner, S.B. Scrapie and cellular prion proteins differ in their kinetics of synthesis and topology in cultured cells. J. Cell Biol. 1990, 110, 743–752. [Google Scholar] [CrossRef] [Green Version]
- Mattei, V.; Barenco, M.G.; Tasciotti, V.; Garofalo, T.; Longo, A.; Boller, K.; Lower, J.; Misasi, R.; Montrasio, F.; Sorice, M. Paracrine diffusion of PrP(C) and propagation of prion infectivity by plasma membrane-derived microvesicles. PLoS ONE 2009, 4, e5057. [Google Scholar] [CrossRef]
- Alais, S.; Simoes, S.; Baas, D.; Lehmann, S.; Raposo, G.; Darlix, J.L.; Leblanc, P. Mouse neuroblastoma cells release prion infectivity associated with exosomal vesicles. Biol. Cell 2008, 100, 603–615. [Google Scholar] [CrossRef] [Green Version]
- Chesebro, B. Introduction to the transmissible spongiform encephalopathies or prion diseases. Br. Med. Bull. 2003, 66, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Pan, K.M.; Baldwin, M.; Nguyen, J.; Gasset, M.; Serban, A.; Groth, D.; Mehlhorn, I.; Huang, Z.; Fletterick, R.J.; Cohen, F.E.; et al. Conversion of alpha-helices into beta-sheets features in the formation of the scrapie prion proteins. Proc. Natl. Acad. Sci. USA 1993, 90, 10962–10966. [Google Scholar] [CrossRef] [Green Version]
- Meisl, G.; Kurt, T.; Condado-Morales, I.; Bett, C.; Sorce, S.; Nuvolone, M.; Michaels, T.C.T.; Heinzer, D.; Avar, M.; Cohen, S.I.A.; et al. Scaling analysis reveals the mechanism and rates of prion replication in vivo. Nat. Struct. Mol. Biol. 2021, 28, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Bruce, M.E. Scrapie strain variation and mutation. Br. Med. Bull. 1993, 49, 822–838. [Google Scholar] [CrossRef] [PubMed]
- Bessen, R.A.; Marsh, R.F. Distinct PrP properties suggest the molecular basis of strain variation in transmissible mink encephalopathy. J. Virol. 1994, 68, 7859–7868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safar, J.; Wille, H.; Itri, V.; Groth, D.; Serban, H.; Torchia, M.; Cohen Gousset, K.; Schiff, E.; Langevin, C.; Marijanovic, Z.; et al. Prions hijack tunnelling nanotubes for intercellular spread. Nat. Cell Biol. 2009, 11, 328–336. [Google Scholar] [CrossRef]
- Safar, J.; Wille, H.; Itri, V.; Groth, D.; Serban, H.; Torchia, M.; Cohen, F.E.; Prusiner, S.B. Eight prion strains have PrP(Sc) molecules with different conformations. Nat. Med. 1998, 4, 1157–1165. [Google Scholar] [CrossRef]
- Arellano-Anaya, Z.E.; Huor, A.; Leblanc, P.; Lehmann, S.; Provansal, M.; Raposo, G.; Andreoletti, O.; Vilette, D. Prion strains are differentially released through the exosomal pathway. Cell Mol. Life Sci. 2015, 72, 1185–1196. [Google Scholar] [CrossRef]
- Kanu, N.; Imokawa, Y.; Drechsel, D.N.; Williamson, R.A.; Birkett, C.R.; Bostock, C.J.; Brockes, J.P. Transfer of scrapie prion infectivity by cell contact in culture. Curr. Biol. 2002, 12, 523–530. [Google Scholar] [CrossRef] [Green Version]
- Grassmann, A.; Wolf, H.; Hofmann, J.; Graham, J.; Vorberg, I. Cellular aspects of prion replication in vitro. Viruses 2013, 5, 374–405. [Google Scholar] [CrossRef] [Green Version]
- Castle, A.R.; Gill, A.C. Physiological Functions of the Cellular Prion Protein. Front. Mol Biosci. 2017, 4, 19. [Google Scholar] [CrossRef] [Green Version]
- Panes, J.D.; Saavedra, P.; Pineda, B.; Escobar, K.; Cuevas, M.E.; Moraga-Cid, G.; Fuentealba, J.; Rivas, C.I.; Rezaei, H.; Munoz-Montesino, C. PrP (C) as a Transducer of Physiological and Pathological Signals. Front. Mol. Neurosci. 2021, 14, 762918. [Google Scholar] [CrossRef]
- Go, G.; Yun, C.W.; Yoon, Y.M.; Lim, J.H.; Lee, J.H.; Lee, S.H. Role of PrP(C) in Cancer Stem Cell Characteristics and Drug Resistance in Colon Cancer Cells. Anticancer Res. 2020, 40, 5611–5620. [Google Scholar] [CrossRef] [PubMed]
- Schmitt-Ulms, G.; Mehrabian, M.; Williams, D.; Ehsani, S. The IDIP framework for assessing protein function and its application to the prion protein. Biol. Rev. Camb. Philos. Soc. 2021, 96, 1907–1932. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.R.; Hooper, N.M. The prion protein and lipid rafts. Mol. Membr. Biol. 2006, 23, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Leblanc, P.; Alais, S.; Porto-Carreiro, I.; Lehmann, S.; Grassi, J.; Raposo, G.; Darlix, J.L. Retrovirus infection strongly enhances scrapie infectivity release in cell culture. EMBO J. 2006, 25, 2674–2685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altmeppen, H.C.; Prox, J.; Puig, B.; Dohler, F.; Falker, C.; Krasemann, S.; Glatzel, M. Roles of endoproteolytic alpha-cleavage and shedding of the prion protein in neurodegeneration. FEBS J. 2013, 280, 4338–4347. [Google Scholar] [CrossRef] [PubMed]
- Shyng, S.L.; Moulder, K.L.; Lesko, A.; Harris, D.A. The N-terminal domain of a glycolipid-anchored prion protein is essential for its endocytosis via clathrin-coated pits. J. Biol. Chem. 1995, 270, 14793–14800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vey, M.; Pilkuhn, S.; Wille, H.; Nixon, R.; DeArmond, S.J.; Smart, E.J.; Anderson, R.G.; Taraboulos, A.; Prusiner, S.B. Subcellular colocalization of the cellular and scrapie prion proteins in caveolae-like membranous domains. Proc. Natl. Acad. Sci. USA 1996, 93, 14945–14949. [Google Scholar] [CrossRef] [Green Version]
- Uchiyama, K.; Tomita, M.; Yano, M.; Chida, J.; Hara, H.; Das, N.R.; Nykjaer, A.; Sakaguchi, S. Prions amplify through degradation of the VPS10P sorting receptor sortilin. PLoS Pathog. 2017, 13, e1006470. [Google Scholar] [CrossRef] [Green Version]
- Taraboulos, A.; Rogers, M.; Borchelt, D.R.; McKinley, M.P.; Scott, M.; Serban, D.; Prusiner, S.B. Acquisition of protease resistance by prion proteins in scrapie-infected cells does not require asparagine-linked glycosylation. Proc. Natl. Acad. Sci. USA 1990, 87, 8262–8266. [Google Scholar] [CrossRef] [Green Version]
- Heisler, F.F.; Pechmann, Y.; Wieser, I.; Altmeppen, H.C.; Veenendaal, L.; Muhia, M.; Schweizer, M.; Glatzel, M.; Krasemann, S.; Kneussel, M. Muskelin Coordinates PrP(C) Lysosome versus Exosome Targeting and Impacts Prion Disease Progression. Neuron 2018, 99, 1155–1169.e1159. [Google Scholar] [CrossRef] [Green Version]
- Caughey, B.; Race, R.E.; Ernst, D.; Buchmeier, M.J.; Chesebro, B. Prion protein biosynthesis in scrapie-infected and uninfected neuroblastoma cells. J. Virol. 1989, 63, 175–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwamaru, Y.; Takenouchi, T.; Ogihara, K.; Hoshino, M.; Takata, M.; Imamura, M.; Tagawa, Y.; Hayashi-Kato, H.; Ushiki-Kaku, Y.; Shimizu, Y.; et al. Microglial cell line established from prion protein-overexpressing mice is susceptible to various murine prion strains. J. Virol. 2007, 81, 1524–1527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caughey, B.; Raymond, G.J. The scrapie-associated form of PrP is made from a cell surface precursor that is both protease- and phospholipase-sensitive. J. Biol. Chem. 1991, 266, 18217–18223. [Google Scholar] [CrossRef]
- Caughey, B.; Baron, G.S.; Chesebro, B.; Jeffrey, M. Getting a grip on prions: Oligomers, amyloids, and pathological membrane interactions. Annu. Rev. Biochem. 2009, 78, 177–204. [Google Scholar] [CrossRef] [Green Version]
- Hutti, C.R.; Welle, K.A.; Hryhorenko, J.R.; Ghaemmaghami, S. Publisher Correction: Global analysis of protein degradation in prion infected cells. Sci. Rep. 2020, 10, 12952. [Google Scholar] [CrossRef]
- Prusiner, S.B. The prion diseases. Brain Pathol. 1998, 8, 499–513. [Google Scholar] [CrossRef]
- Klohn, P.C.; Stoltze, L.; Flechsig, E.; Enari, M.; Weissmann, C. A quantitative, highly sensitive cell-based infectivity assay for mouse scrapie prions. Proc. Natl. Acad. Sci. USA 2003, 100, 11666–11671. [Google Scholar] [CrossRef] [Green Version]
- Nishida, N.; Harris, D.A.; Vilette, D.; Laude, H.; Frobert, Y.; Grassi, J.; Casanova, D.; Milhavet, O.; Lehmann, S. Successful transmission of three mouse-adapted scrapie strains to murine neuroblastoma cell lines overexpressing wild-type mouse prion protein. J. Virol. 2000, 74, 320–325. [Google Scholar] [CrossRef] [Green Version]
- Sarnataro, D.; Caputo, A.; Casanova, P.; Puri, C.; Paladino, S.; Tivodar, S.S.; Campana, V.; Tacchetti, C.; Zurzolo, C. Lipid rafts and clathrin cooperate in the internalization of PrP in epithelial FRT cells. PLoS ONE 2009, 4, e5829. [Google Scholar] [CrossRef] [Green Version]
- Schatzl, H.M.; Laszlo, L.; Holtzman, D.M.; Tatzelt, J.; DeArmond, S.J.; Weiner, R.I.; Mobley, W.C.; Prusiner, S.B. A hypothalamic neuronal cell line persistently infected with scrapie prions exhibits apoptosis. J. Virol. 1997, 71, 8821–8831. [Google Scholar] [CrossRef] [Green Version]
- Fevrier, B.; Vilette, D.; Archer, F.; Loew, D.; Faigle, W.; Vidal, M.; Laude, H.; Raposo, G. Cells release prions in association with exosomes. Proc. Natl. Acad. Sci. USA 2004, 101, 9683–9688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanez-Mo, M.; Siljander, P.R.; Andreu, Z.; Zavec, A.B.; Borras, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Race, R.E.; Fadness, L.H.; Chesebro, B. Characterization of scrapie infection in mouse neuroblastoma cells. J. Gen. Virol. 1987, 68, 1391–1399. [Google Scholar] [CrossRef] [PubMed]
- Cronier, S.; Laude, H.; Peyrin, J.M. Prions can infect primary cultured neurons and astrocytes and promote neuronal cell death. Proc. Natl. Acad. Sci. USA 2004, 101, 12271–12276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Archer, F.; Bachelin, C.; Andreoletti, O.; Besnard, N.; Perrot, G.; Langevin, C.; Le Dur, A.; Vilette, D.; Baron-Van Evercooren, A.; Vilotte, J.L.; et al. Cultured peripheral neuroglial cells are highly permissive to sheep prion infection. J. Virol. 2004, 78, 482–490. [Google Scholar] [CrossRef] [Green Version]
- Vilette, D.; Andreoletti, O.; Archer, F.; Madelaine, M.F.; Vilotte, J.L.; Lehmann, S.; Laude, H. Ex vivo propagation of infectious sheep scrapie agent in heterologous epithelial cells expressing ovine prion protein. Proc. Natl. Acad. Sci. USA 2001, 98, 4055–4059. [Google Scholar] [CrossRef] [Green Version]
- Miyazawa, K.; Masujin, K.; Okada, H.; Ushiki-Kaku, Y.; Matsuura, Y.; Yokoyama, T. Selective propagation of mouse-passaged scrapie prions with long incubation period from a mixed prion population using GT1-7 cells. PLoS ONE 2017, 12, e0179317. [Google Scholar] [CrossRef] [Green Version]
- Walia, R.; Ho, C.C.; Lee, C.; Gilch, S.; Schatzl, H.M. Gene-edited murine cell lines for propagation of chronic wasting disease prions. Sci. Rep. 2019, 9, 11151. [Google Scholar] [CrossRef] [Green Version]
- Raymond, G.J.; Olsen, E.A.; Lee, K.S.; Raymond, L.D.; Bryant, P.K., 3rd; Baron, G.S.; Caughey, W.S.; Kocisko, D.A.; McHolland, L.E.; Favara, C.; et al. Inhibition of protease-resistant prion protein formation in a transformed deer cell line infected with chronic wasting disease. J. Virol. 2006, 80, 596–604. [Google Scholar] [CrossRef] [Green Version]
- Bian, J.; Napier, D.; Khaychuck, V.; Angers, R.; Graham, C.; Telling, G. Cell-based quantification of chronic wasting disease prions. J. Virol. 2010, 84, 8322–8326. [Google Scholar] [CrossRef] [Green Version]
- Hannaoui, S.; Gougerot, A.; Privat, N.; Levavasseur, E.; Bizat, N.; Hauw, J.J.; Brandel, J.P.; Haik, S. Cycline efficacy on the propagation of human prions in primary cultured neurons is strain-specific. J. Infect. Dis. 2014, 209, 1144–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahal, S.P.; Baker, C.A.; Demczyk, C.A.; Smith, E.W.; Julius, C.; Weissmann, C. Prion strain discrimination in cell culture: The cell panel assay. Proc. Natl. Acad. Sci. USA 2007, 104, 20908–20913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathieu, M.; Martin-Jaular, L.; Lavieu, G.; Thery, C. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat. Cell Biol. 2019, 21, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef] [PubMed]
- Marzo, L.; Marijanovic, Z.; Browman, D.; Chamoun, Z.; Caputo, A.; Zurzolo, C. 4-hydroxytamoxifen leads to PrPSc clearance by conveying both PrPC and PrPSc to lysosomes independently of autophagy. J. Cell Sci. 2013, 126, 1345–1354. [Google Scholar] [CrossRef] [Green Version]
- Fuentes, P.; Sese, M.; Guijarro, P.J.; Emperador, M.; Sanchez-Redondo, S.; Peinado, H.; Hummer, S.; Ramon, Y.C.S. ITGB3-mediated uptake of small extracellular vesicles facilitates intercellular communication in breast cancer cells. Nat. Commun. 2020, 11, 4261. [Google Scholar] [CrossRef]
- Christianson, H.C.; Svensson, K.J.; van Kuppevelt, T.H.; Li, J.P.; Belting, M. Cancer cell exosomes depend on cell-surface heparan sulfate proteoglycans for their internalization and functional activity. Proc. Natl. Acad. Sci. USA 2013, 110, 17380–17385. [Google Scholar] [CrossRef] [Green Version]
- Ligios, C.; Sigurdson, C.J.; Santucciu, C.; Carcassola, G.; Manco, G.; Basagni, M.; Maestrale, C.; Cancedda, M.G.; Madau, L.; Aguzzi, A. PrPSc in mammary glands of sheep affected by scrapie and mastitis. Nat. Med. 2005, 11, 1137–1138. [Google Scholar] [CrossRef]
- Vorberg, I.; Raines, A.; Story, B.; Priola, S.A. Susceptibility of common fibroblast cell lines to transmissible spongiform encephalopathy agents. J. Infect. Dis. 2004, 189, 431–439. [Google Scholar] [CrossRef] [Green Version]
- Hara, H.; Chida, J.; Uchiyama, K.; Pasiana, A.D.; Takahashi, E.; Kido, H.; Sakaguchi, S. Neurotropic influenza A virus infection causes prion protein misfolding into infectious prions in neuroblastoma cells. Sci. Rep. 2021, 11, 10109. [Google Scholar] [CrossRef]
- Birkett, C.R.; Hennion, R.M.; Bembridge, D.A.; Clarke, M.C.; Chree, A.; Bruce, M.E.; Bostock, C.J. Scrapie strains maintain biological phenotypes on propagation in a cell line in culture. EMBO J. 2001, 20, 3351–3358. [Google Scholar] [CrossRef]
- Casaccia-Bonnefil, P.; Kascsak, R.J.; Fersko, R.; Callahan, S.; Carp, R.I. Brain regional distribution of prion protein PrP27-30 in mice stereotaxically microinjected with different strains of scrapie. J. Infect. Dis. 1993, 167, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Gerdes, H.H.; Carvalho, R.N. Intercellular transfer mediated by tunneling nanotubes. Curr. Opin. Cell Biol. 2008, 20, 470–475. [Google Scholar] [CrossRef]
- Magalhaes, A.C.; Baron, G.S.; Lee, K.S.; Steele-Mortimer, O.; Dorward, D.; Prado, M.A.; Caughey, B. Uptake and neuritic transport of scrapie prion protein coincident with infection of neuronal cells. J. Neurosci. 2005, 25, 5207–5216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, K.E.; Hughson, A.; Vascellari, S.; Priola, S.A.; Sakudo, A.; Onodera, T.; Baron, G.S. PrP Knockout Cells Expressing Transmembrane PrP Resist Prion Infection. J. Virol. 2017, 91, e01686–e01716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chivet, M.; Javalet, C.; Laulagnier, K.; Blot, B.; Hemming, F.J.; Sadoul, R. Exosomes secreted by cortical neurons upon glutamatergic synapse activation specifically interact with neurons. J. Extracell Vesicles 2014, 3, 24722. [Google Scholar] [CrossRef] [Green Version]
- Leblanc, P.; Hasenkrug, K.; Ward, A.; Myers, L.; Messer, R.J.; Alais, S.; Timmes, A.; Priola, S.A. Co-infection with the friend retrovirus and mouse scrapie does not alter prion disease pathogenesis in susceptible mice. PLoS ONE 2012, 7, e30872. [Google Scholar] [CrossRef] [Green Version]
- Butler, D.A.; Scott, M.R.; Bockman, J.M.; Borchelt, D.R.; Taraboulos, A.; Hsiao, K.K.; Kingsbury, D.T.; Prusiner, S.B. Scrapie-infected murine neuroblastoma cells produce protease-resistant prion proteins. J. Virol. 1988, 62, 1558–1564. [Google Scholar] [CrossRef] [Green Version]
- Rubenstein, R.; Carp, R.I.; Callahan, S.M. In vitro replication of scrapie agent in a neuronal model: Infection of PC12 cells. J. Gen. Virol. 1984, 65, 2191–2198. [Google Scholar] [CrossRef]
- Hannaoui, S.; Maatouk, L.; Privat, N.; Levavasseur, E.; Faucheux, B.A.; Haik, S. Prion propagation and toxicity occur in vitro with two-phase kinetics specific to strain and neuronal type. J. Virol. 2013, 87, 2535–2548. [Google Scholar] [CrossRef] [Green Version]
- Clarke, M.C.; Haig, D.A. Multiplication of scrapie agent in cell culture. Res. Vet. Sci. 1970, 11, 500–501. [Google Scholar] [CrossRef]
- Roffe, M.; Beraldo, F.H.; Bester, R.; Nunziante, M.; Bach, C.; Mancini, G.; Gilch, S.; Vorberg, I.; Castilho, B.A.; Martins, V.R.; et al. Prion protein interaction with stress-inducible protein 1 enhances neuronal protein synthesis via mTOR. Proc. Natl. Acad. Sci. USA 2010, 107, 13147–13152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jen, A.; Parkyn, C.J.; Mootoosamy, R.C.; Ford, M.J.; Warley, A.; Liu, Q.; Bu, G.; Baskakov, I.V.; Moestrup, S.; McGuinness, L.; et al. Neuronal low-density lipoprotein receptor-related protein 1 binds and endocytoses prion fibrils via receptor cluster 4. J. Cell Sci. 2010, 123, 246–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coleman, B.M.; Hanssen, E.; Lawson, V.A.; Hill, A.F. Prion-infected cells regulate the release of exosomes with distinct ultrastructural features. FASEB J. 2012, 26, 4160–4173. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Hossinger, A.; Heumuller, S.E.; Hornberger, A.; Buravlova, O.; Konstantoulea, K.; Muller, S.A.; Paulsen, L.; Rousseau, F.; Schymkowitz, J.; et al. Highly efficient intercellular spreading of protein misfolding mediated by viral ligand-receptor interactions. Nat. Commun. 2021, 12, 5739. [Google Scholar] [CrossRef]
- Bourkas, M.E.C.; Arshad, H.; Al-Azzawi, Z.A.M.; Halgas, O.; Shikiya, R.A.; Mehrabian, M.; Schmitt-Ulms, G.; Bartz, J.C.; Watts, J.C. Engineering a murine cell line for the stable propagation of hamster prions. J. Biol. Chem. 2019, 294, 4911–4923. [Google Scholar] [CrossRef]
- Courageot, M.P.; Daude, N.; Nonno, R.; Paquet, S.; Di Bari, M.A.; Le Dur, A.; Chapuis, J.; Hill, A.F.; Agrimi, U.; Laude, H.; et al. A cell line infectible by prion strains from different species. J Gen Virol 2008, 89, 341–347. [Google Scholar] [CrossRef]
- Guo, B.B.; Bellingham, S.A.; Hill, A.F. Stimulating the Release of Exosomes Increases the Intercellular Transfer of Prions. J. Biol. Chem. 2016, 291, 5128–5137. [Google Scholar] [CrossRef] [Green Version]
- Hasebe, R.; Tanaka, M.; Suzuki, A.; Yamasaki, T.; Horiuchi, M. Complement factors alter the amount of PrP(Sc) in primary-cultured mouse cortical neurons associated with increased membrane permeability. Virology 2016, 496, 9–20. [Google Scholar] [CrossRef]
- Krejciova, Z.; Alibhai, J.; Zhao, C.; Krencik, R.; Rzechorzek, N.M.; Ullian, E.M.; Manson, J.; Ironside, J.W.; Head, M.W.; Chandran, S. Human stem cell-derived astrocytes replicate human prions in a PRNP genotype-dependent manner. J. Exp. Med. 2017, 214, 3481–3495. [Google Scholar] [CrossRef] [Green Version]
- Follet, J.; Lemaire-Vieille, C.; Blanquet-Grossard, F.; Podevin-Dimster, V.; Lehmann, S.; Chauvin, J.P.; Decavel, J.P.; Varea, R.; Grassi, J.; Fontes, M.; et al. PrP expression and replication by Schwann cells: Implications in prion spreading. J. Virol. 2002, 76, 2434–2439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cronier, S.; Beringue, V.; Bellon, A.; Peyrin, J.M.; Laude, H. Prion strain- and species-dependent effects of antiprion molecules in primary neuronal cultures. J. Virol. 2007, 81, 13794–13800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vella, L.J.; Sharples, R.A.; Lawson, V.A.; Masters, C.L.; Cappai, R.; Hill, A.F. Packaging of prions into exosomes is associated with a novel pathway of PrP processing. J. Pathol. 2007, 211, 582–590. [Google Scholar] [CrossRef] [PubMed]
- Nishida, N.; Katamine, S.; Manuelidis, L. Reciprocal interference between specific CJD and scrapie agents in neural cell cultures. Science 2005, 310, 493–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arjona, A.; Simarro, L.; Islinger, F.; Nishida, N.; Manuelidis, L. Two Creutzfeldt-Jakob disease agents reproduce prion protein-independent identities in cell cultures. Proc. Natl. Acad. Sci. USA 2004, 101, 8768–8773. [Google Scholar] [CrossRef] [Green Version]
- Dinkel, K.D.; Schneider, D.A.; Munoz-Gutierrez, J.F.; McElliott, V.R.; Stanton, J.B. Correlation of cellular factors and differential scrapie prion permissiveness in ovine microglia. Virus Res. 2017, 240, 69–80. [Google Scholar] [CrossRef]
- Brown, C.A.; Schmidt, C.; Poulter, M.; Hummerich, H.; Klohn, P.C.; Jat, P.; Mead, S.; Collinge, J.; Lloyd, S.E. In vitro screen of prion disease susceptibility genes using the scrapie cell assay. Hum. Mol. Genet. 2014, 23, 5102–5108. [Google Scholar] [CrossRef] [Green Version]
- Bosque, P.J.; Prusiner, S.B. Cultured cell sublines highly susceptible to prion infection. J. Virol. 2000, 74, 4377–4386. [Google Scholar] [CrossRef] [Green Version]
- Philiastides, A.; Ribes, J.M.; Yip, D.C.; Schmidt, C.; Benilova, I.; Klohn, P.C. A New Cell Model for Investigating Prion Strain Selection and Adaptation. Viruses 2019, 11, 888. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Tark, D.S.; Lee, Y.H.; Kim, M.J.; Lee, W.Y.; Cho, I.S.; Sohn, H.J.; Yokoyama, T. Establishment of a cell line persistently infected with chronic wasting disease prions. J. Vet. Med. Sci. 2012, 74, 1377–1380. [Google Scholar] [CrossRef] [Green Version]
- Lawson, V.A.; Vella, L.J.; Stewart, J.D.; Sharples, R.A.; Klemm, H.; Machalek, D.M.; Masters, C.L.; Cappai, R.; Collins, S.J.; Hill, A.F. Mouse-adapted sporadic human Creutzfeldt-Jakob disease prions propagate in cell culture. Int. J. Biochem. Cell Biol. 2008, 40, 2793–2801. [Google Scholar] [CrossRef] [PubMed]
- Tark, D.; Kim, H.; Neale, M.H.; Kim, M.; Sohn, H.; Lee, Y.; Cho, I.; Joo, Y.; Windl, O. Generation of a persistently infected MDBK cell line with natural bovine spongiform encephalopathy (BSE). PLoS ONE 2015, 10, e0115939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baron, G.S.; Magalhaes, A.C.; Prado, M.A.; Caughey, B. Mouse-adapted scrapie infection of SN56 cells: Greater efficiency with microsome-associated versus purified PrP-res. J. Virol. 2006, 80, 2106–2117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greil, C.S.; Vorberg, I.M.; Ward, A.E.; Meade-White, K.D.; Harris, D.A.; Priola, S.A. Acute cellular uptake of abnormal prion protein is cell type and scrapie-strain independent. Virology 2008, 379, 284–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michiels, E.; Liu, S.; Gallardo, R.; Louros, N.; Mathelie-Guinlet, M.; Dufrene, Y.; Schymkowitz, J.; Vorberg, I.; Rousseau, F. Entropic Bristles Tune the Seeding Efficiency of Prion-Nucleating Fragments. Cell Rep. 2020, 30, 2834–2845 e2833. [Google Scholar] [CrossRef] [Green Version]
- Walzlein, J.H.; Schwenke, K.A.; Beekes, M. Propagation of CJD Prions in Primary Murine Glia Cells Expressing Human PrP(c). Pathogens 2021, 10, 1060. [Google Scholar] [CrossRef]
- Properzi, F.; Badhan, A.; Klier, S.; Schmidt, C.; Klohn, P.C.; Wadsworth, J.D.; Clarke, A.R.; Jackson, G.S.; Collinge, J. Physical, chemical and kinetic factors affecting prion infectivity. Prion 2016, 10, 251–261. [Google Scholar] [CrossRef] [Green Version]
- Horonchik, L.; Tzaban, S.; Ben-Zaken, O.; Yedidia, Y.; Rouvinski, A.; Papy-Garcia, D.; Barritault, D.; Vlodavsky, I.; Taraboulos, A. Heparan sulfate is a cellular receptor for purified infectious prions. J. Biol. Chem. 2005, 280, 17062–17067. [Google Scholar] [CrossRef] [Green Version]
- Wolf, H.; Grassmann, A.; Bester, R.; Hossinger, A.; Mohl, C.; Paulsen, L.; Groschup, M.H.; Schatzl, H.; Vorberg, I. Modulation of Glycosaminoglycans Affects PrPSc Metabolism but Does Not Block PrPSc Uptake. J. Virol. 2015, 89, 9853–9864. [Google Scholar] [CrossRef] [Green Version]
- Leucht, C.; Simoneau, S.; Rey, C.; Vana, K.; Rieger, R.; Lasmezas, C.I.; Weiss, S. The 37 kDa/67 kDa laminin receptor is required for PrP(Sc) propagation in scrapie-infected neuronal cells. EMBO Rep. 2003, 4, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Victoria, G.S.; Arkhipenko, A.; Zhu, S.; Syan, S.; Zurzolo, C. Astrocyte-to-neuron intercellular prion transfer is mediated by cell-cell contact. Sci. Rep. 2016, 6, 20762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamasaki, T.; Baron, G.S.; Suzuki, A.; Hasebe, R.; Horiuchi, M. Characterization of intracellular dynamics of inoculated PrP-res and newly generated PrP(Sc) during early stage prion infection in Neuro2a cells. Virology 2014, 450–451, 324–335. [Google Scholar] [CrossRef] [PubMed]
- Vorberg, I.; Raines, A.; Priola, S.A. Acute formation of protease-resistant prion protein does not always lead to persistent scrapie infection in vitro. J. Biol. Chem. 2004, 279, 29218–29225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghaemmaghami, S.; Phuan, P.W.; Perkins, B.; Ullman, J.; May, B.C.; Cohen, F.E.; Prusiner, S.B. Cell division modulates prion accumulation in cultured cells. Proc. Natl. Acad. Sci. USA 2007, 104, 17971–17976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krauss, S.; Vorberg, I. Prions Ex Vivo: What Cell Culture Models Tell Us about Infectious Proteins. Int. J. Cell. Biol. 2013, 2013, 704546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maas, E.; Geissen, M.; Groschup, M.H.; Rost, R.; Onodera, T.; Schatzl, H.; Vorberg, I.M. Scrapie infection of prion protein-deficient cell line upon ectopic expression of mutant prion proteins. J. Biol. Chem. 2007, 282, 18702–18710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bach, C.; Gilch, S.; Rost, R.; Greenwood, A.D.; Horsch, M.; Hajj, G.N.; Brodesser, S.; Facius, A.; Schadler, S.; Sandhoff, K.; et al. Prion-induced activation of cholesterogenic gene expression by Srebp2 in neuronal cells. J. Biol. Chem. 2009, 284, 31260–31269. [Google Scholar] [CrossRef] [Green Version]
- Borchelt, D.R.; Taraboulos, A.; Prusiner, S.B. Evidence for synthesis of scrapie prion proteins in the endocytic pathway. J. Biol. Chem. 1992, 267, 16188–16199. [Google Scholar] [CrossRef]
- McNally, K.L.; Ward, A.E.; Priola, S.A. Cells expressing anchorless prion protein are resistant to scrapie infection. J. Virol. 2009, 83, 4469–4475. [Google Scholar] [CrossRef] [Green Version]
- Rouvinski, A.; Karniely, S.; Kounin, M.; Moussa, S.; Goldberg, M.D.; Warburg, G.; Lyakhovetsky, R.; Papy-Garcia, D.; Kutzsche, J.; Korth, C.; et al. Live imaging of prions reveals nascent PrPSc in cell-surface, raft-associated amyloid strings and webs. J. Cell Biol. 2014, 204, 423–441. [Google Scholar] [CrossRef] [Green Version]
- Yim, Y.I.; Park, B.C.; Yadavalli, R.; Zhao, X.; Eisenberg, E.; Greene, L.E. The multivesicular body is the major internal site of prion conversion. J. Cell Sci. 2015, 128, 1434–1443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chernoff, Y.O.; Lindquist, S.L.; Ono, B.; Inge-Vechtomov, S.G.; Liebman, S.W. Role of the chaperone protein Hsp104 in propagation of the yeast prion-like factor [psi+]. Science 1995, 268, 880–884. [Google Scholar] [CrossRef]
- Zhu, S.; Victoria, G.S.; Marzo, L.; Ghosh, R.; Zurzolo, C. Prion aggregates transfer through tunneling nanotubes in endocytic vesicles. Prion 2015, 9, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Vilette, D.; Laulagnier, K.; Huor, A.; Alais, S.; Simoes, S.; Maryse, R.; Provansal, M.; Lehmann, S.; Andreoletti, O.; Schaeffer, L.; et al. Efficient inhibition of infectious prions multiplication and release by targeting the exosomal pathway. Cell Mol. Life Sci. 2015, 72, 4409–4427. [Google Scholar] [CrossRef] [PubMed]
- Krasemann, S.; Neumann, M.; Luepke, J.P.; Grashorn, J.; Wurr, S.; Stocking, C.; Glatzel, M. Persistent retroviral infection with MoMuLV influences neuropathological signature and phenotype of prion disease. Acta Neuropathol. 2012, 124, 111–126. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Species Inoculum | TSE | Strain | Prion Susceptible Cell Line | Expressed PrP | References |
|---|---|---|---|---|---|
| Sheep | Scrapie | Natural Scrapie | RK13, hTERT microglia, MovS6/ MovS2 cells | Ovine | [42,43,44] |
| Primary neurons: cerebellar granular, iPSC-derived astrocytes | Ovine | [45] | |||
| 127S | RK13, MovS6/ MovS2 cells | Ovine | [13,41] | ||
| PG127 | Rov cells | Ovine | [46] | ||
| LA404 | Rov cells | Ovine | [46] | ||
| Kanagawa Scrapie | GT1 | Murine | [47] | ||
| Obihiro Scrapie | MG20 | Murine | [48] | ||
| Elk/Deer | Cervid chronic wasting disease (CWD) | Mule Deer CWD (MD-CWD) | CAD5 | Cervid | [49] |
| MEF | bank Vole/cervid | [49] | |||
| MDB | Mule Deer | [50] | |||
| White-Tailed Deer CWD (WT-CWD) | CAD5 | Cervid | [49] | ||
| MEF | Bank Vole/cervid | ||||
| Elk CWD | RK13 | Elk | [51] | ||
| Cattle | Bovine spongiform encephalopathy (BSE) | BSE | MG20 | Murine | [48] |
| Human | Creutzfeldt–Jakob disease (CJD) | Sporadic CJD | Primary neurons: cerebellar granular, iPSC-derived astrocytes | Human | [52,53] |
| Variant CJD | Primary neurons: cerebellar granular, iPSC-derived astrocytes | Human | [52,53] | ||
| Iatrogenic (iCJD) | Primary neurons: cerebellar granular | Human | [52] | ||
| Mouse-adapted | Scrapie | Ch./RML | N2a (and subclones), SMB, GT1, CAD5, SN56, 1C11, MG20, C8D1A, MSC-80, L929, RK13 | Murine | [38,42,48,54,55,56,57,58,59,60,61,62] |
| Primary neurons: cortical, hippocampal | Murine | [56,63,64] | |||
| 79 A | N2a PK1 subclone, SMB, CAD5, L929 | Murine | [39,40,48,65] | ||
| 139 A | N2a (and subclone), SMB, GT1, CAD5, CRBL, L929 | Murine | [39,40,48,65,66] | ||
| PC12 | Rat | [67] | |||
| Primary neurons: cortical, striatal | Murine | [68] | |||
| Primary neurons: cerebellar granular, astrocytes | Murine/ovine | [41,69] | |||
| 22L | N2a (and subclones), GT1, CAD5, SN56, 1C11, HpL3-4, CF10, C8D1A, L929, NIH/3T3, RK13 | Murine | [39,40,57,62,70,71,72,73] | ||
| Primary neurons: cerebellar granular, cortical, striatal, hippocampal | Murine | [56,63,74] | |||
| ME 7 | N2a subclones, CAD5, SN56, MG20, L929 | Murine | [39,48,61,75] | ||
| Primary neurons: cerebellar granular | Murine | [76] | |||
| PC12 | Rat | [67] | |||
| 22F | SMB | Mouse | [54] | ||
| BSE | 301 C | CAD5 | Murine | [55] | |
| CJD | M1000 | RK13 | Murine | [60,77] | |
| SY | GT1 | Murine | [78] | ||
| FU | N2a, GT1 | Murine | [79] | ||
| GSS | Fukuoka 1 (Fu-1) | GT1, 1C11, RK13 | Murine | [57,72] | |
| Primary neurons: cerebellar granular | Murine | [68,76] | |||
| Bank vole-adapted | BSE | Bank vole-adapted BSE | RK13 | Bank vole | [60] |
| Hamster-adapted | Transm. mink encephalopathy (TME) | Hyper (HY) | CAD5 | Hamster | [80] |
| Scrapie | 263K | CAD5 | Hamster | [80] | |
| 139H | CAD5 | Hamster | [80] |
| Cell Line | Origin | Prion Strain | EV Isolation Method | PrPSc Association with EV | EM Confirmation EV | Recipient Cell | Detection of PrPSc p.i. | Reference |
|---|---|---|---|---|---|---|---|---|
| GT1 | Mouse hypothalamic neurons | RML | Not isolated, conditioned medium used | n.d. | No | N2a GT1 | 6–8 weeks | [57] |
| GT1-7 | Mouse hypothalamic neurons | M1000 | Differential centrifugation | WB | No | RK13 expressing murine PrP | 6 passages | [122] |
| M1000 | Differential centrifugation | WB | Yes | GT1-7; RK13 expressing murine PrP | One month | [77] | ||
| NIH/3T3 | Mouse fibroblast+/− infectionMuLV | 22L | Differential centrifugation | WB | Yes | NIH/3T3 | 16 passages Infection only when donors were MuLV infected | [22] |
| RK13: Rov | Rabbit kidney epithelial cells ectopically expressing ovine PrP | Sheep scrapie PG127 | Differential centrifugation | WB | Yes | Rov | Several weeks | [118] |
| Sheep scrapie PG127 | Differential centrifugation | WB | No | Rov | 4 weeks | [13] | ||
| RK13: moRK13 | Rabbit kidney epithelial cells ectopically expressing murine PrP | 22L | Differential centrifugation | WB | No | moRK13 | 4 weeks | [13] |
| M1000 | Differential centrifugation | WB | Yes | GT1-7; RK13 expressing murine PrP | One month | [77] | ||
| N2a | Mouse neuroblastoma expressing VSV-G | 22L | Differential centrifugation | n.d. | No | L929 CAD | 7–8 passages | [72] |
| Mouse neuroblastoma overexpressing murine PrP | 22L | Differential centrifugation | WB | Yes | N2a | 3 weeks | [75] | |
| Mov | Immortalized Schwann cell-like cells from transgenic mouse expressing ovine PrP | Sheep scrapie PG127 | Differential centrifugation | WB | Yes | Mov | Several weeks | [118] |
| Hpl3-4 moPrP-3F4 | Mouse hippocampus- derived, ectopically expressing epitope-tagged mouse PrP | 22L | Not isolated, conditioned medium used | n.d. | No | Hpl3-4 moPrP-3F4 | 14–28 passages | [101] |
| L929 | Mouse fibroblasts | 22L | Differential centrifugation | WB | No | L929 | Several weeks | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heumüller, S.-E.; Hornberger, A.C.; Hebestreit, A.S.; Hossinger, A.; Vorberg, I.M. Propagation and Dissemination Strategies of Transmissible Spongiform Encephalopathy Agents in Mammalian Cells. Int. J. Mol. Sci. 2022, 23, 2909. https://doi.org/10.3390/ijms23062909
Heumüller S-E, Hornberger AC, Hebestreit AS, Hossinger A, Vorberg IM. Propagation and Dissemination Strategies of Transmissible Spongiform Encephalopathy Agents in Mammalian Cells. International Journal of Molecular Sciences. 2022; 23(6):2909. https://doi.org/10.3390/ijms23062909
Chicago/Turabian StyleHeumüller, Stefanie-Elisabeth, Annika C. Hornberger, Alina S. Hebestreit, André Hossinger, and Ina M. Vorberg. 2022. "Propagation and Dissemination Strategies of Transmissible Spongiform Encephalopathy Agents in Mammalian Cells" International Journal of Molecular Sciences 23, no. 6: 2909. https://doi.org/10.3390/ijms23062909
APA StyleHeumüller, S. -E., Hornberger, A. C., Hebestreit, A. S., Hossinger, A., & Vorberg, I. M. (2022). Propagation and Dissemination Strategies of Transmissible Spongiform Encephalopathy Agents in Mammalian Cells. International Journal of Molecular Sciences, 23(6), 2909. https://doi.org/10.3390/ijms23062909