
MicroRNA 148a Suppresses Tuberculous Fibrosis by Targeting NOX4 and POLDIP2


Abstract
:1. Introduction
2. Results
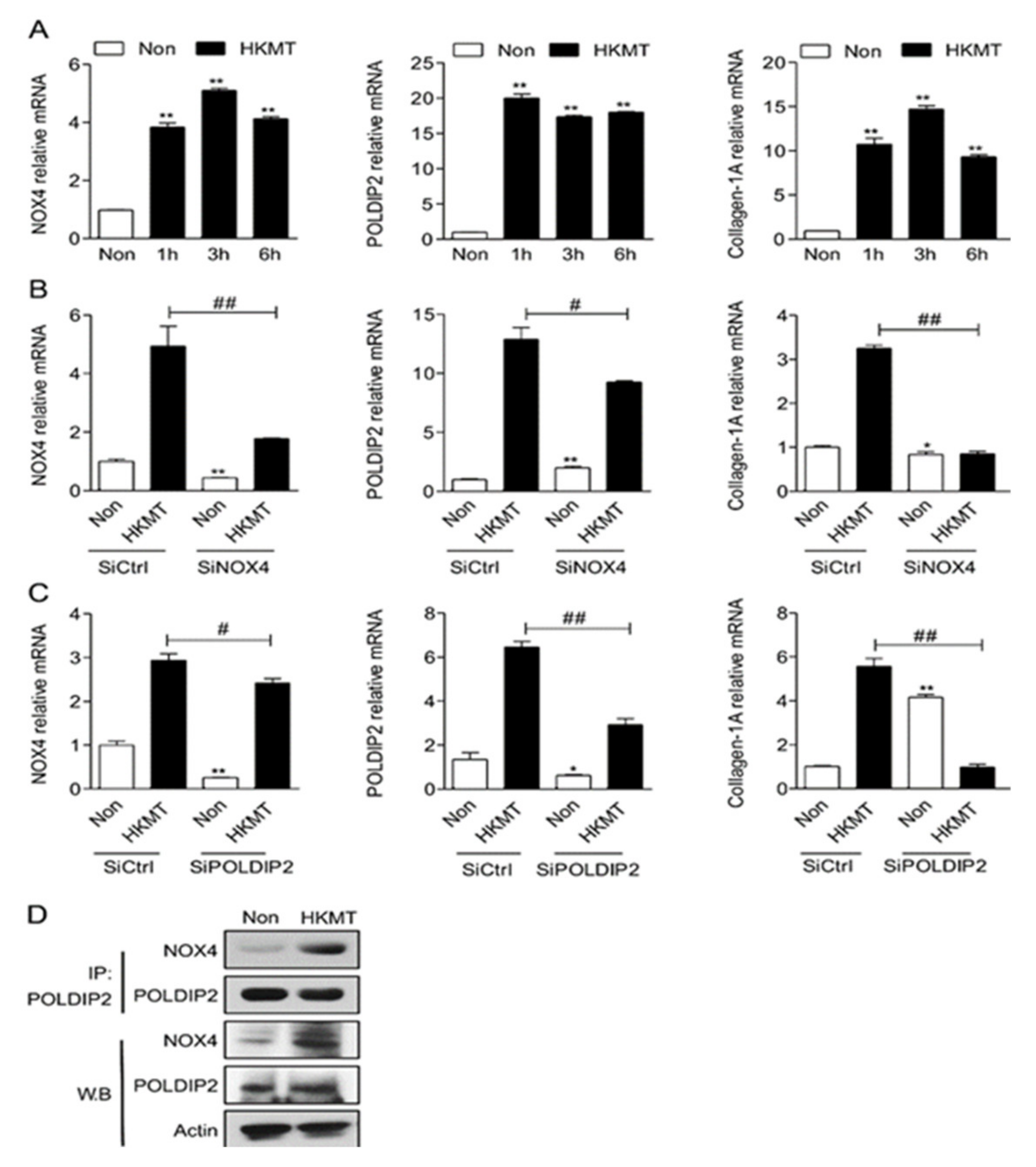
2.1. NOX4 and POLDIP2 Expression in Mesothelial Cells Is Upregulated after Treatment with HKMT
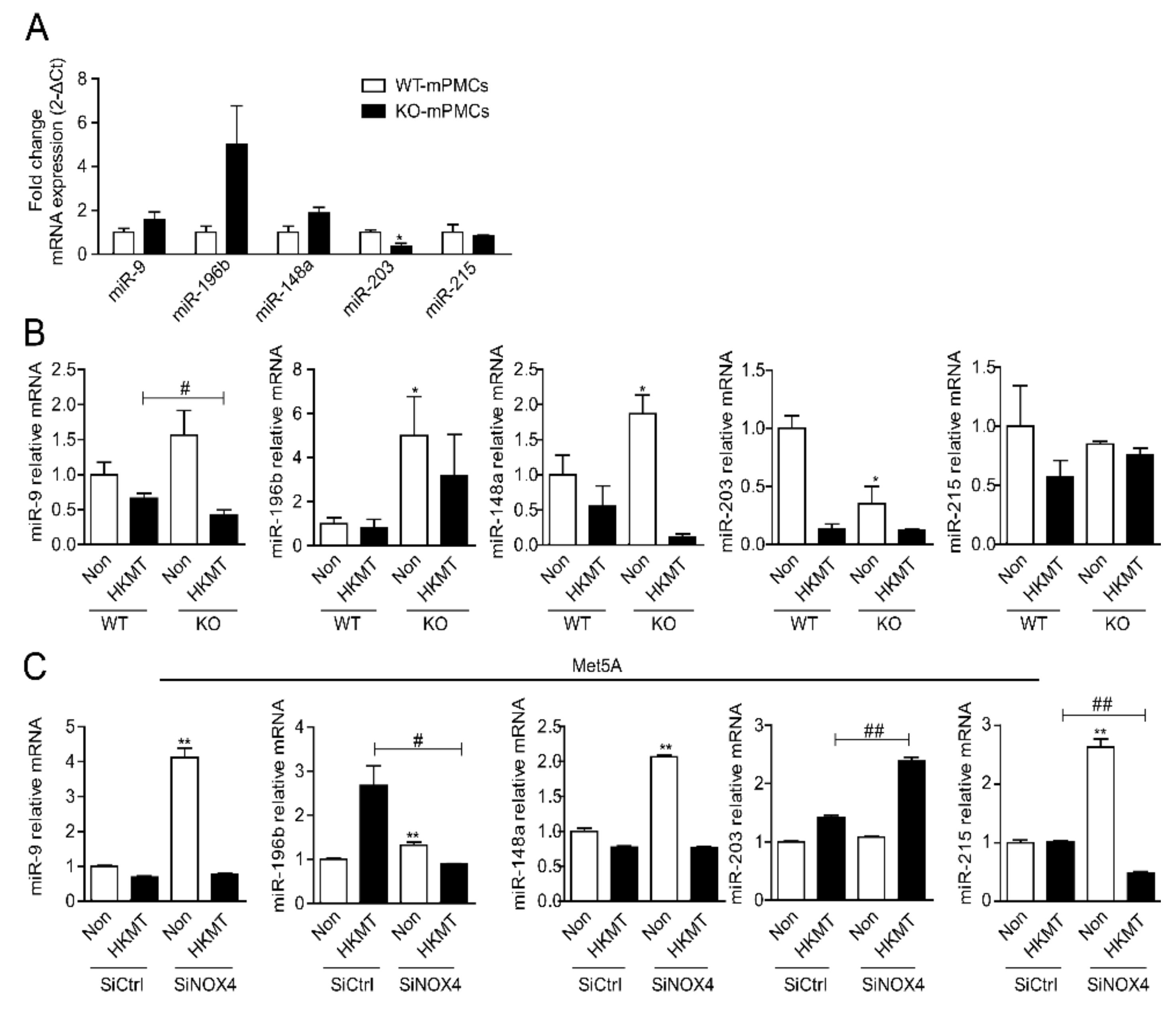
2.2. miR-148a Is Underexpressed during the Initial Stage of Exposure to Tuberculosis but Increases Later
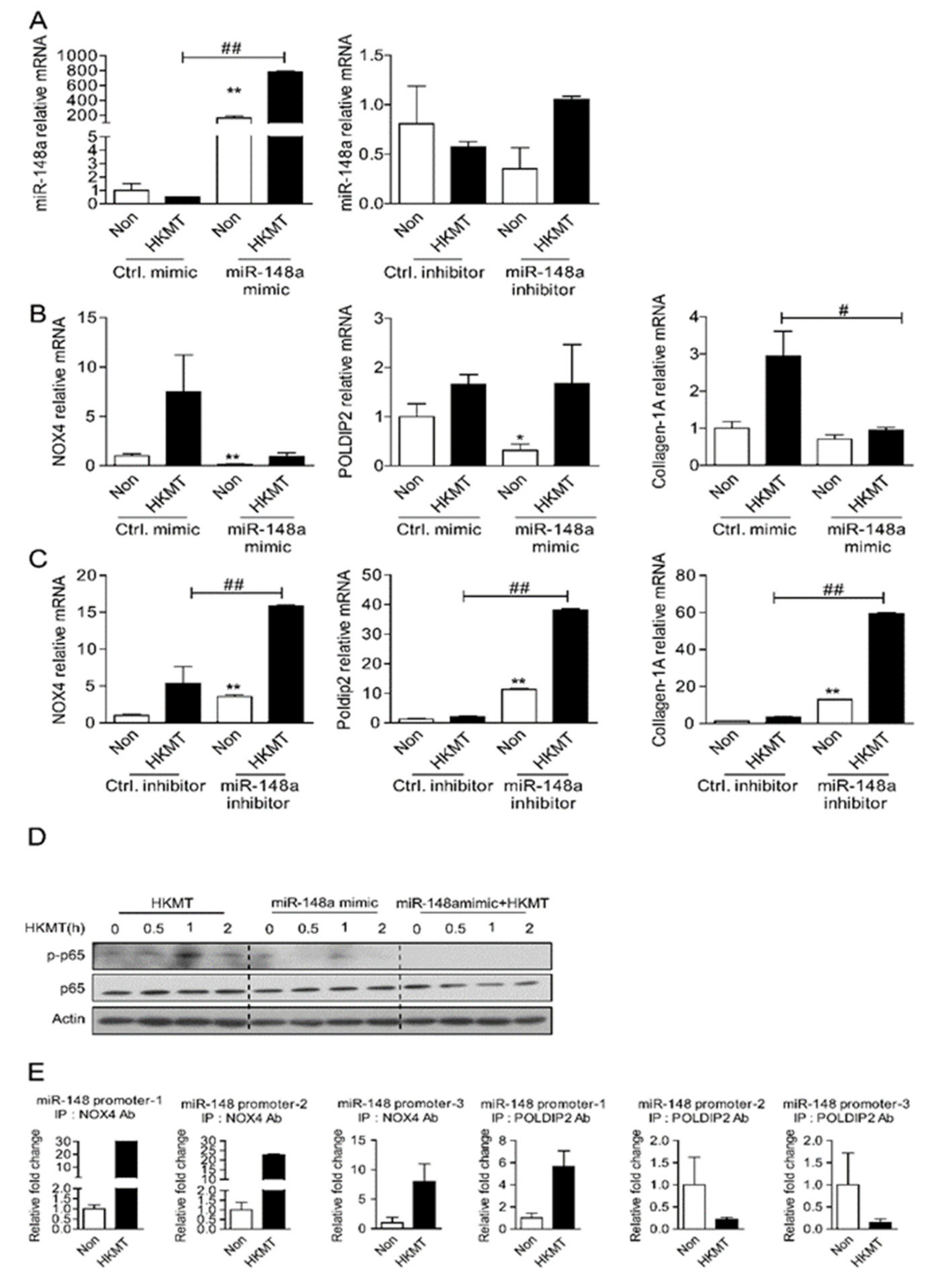
2.3. miR-148a Regulates Expression of NOX4/POLDIP2 in Met5A Cells
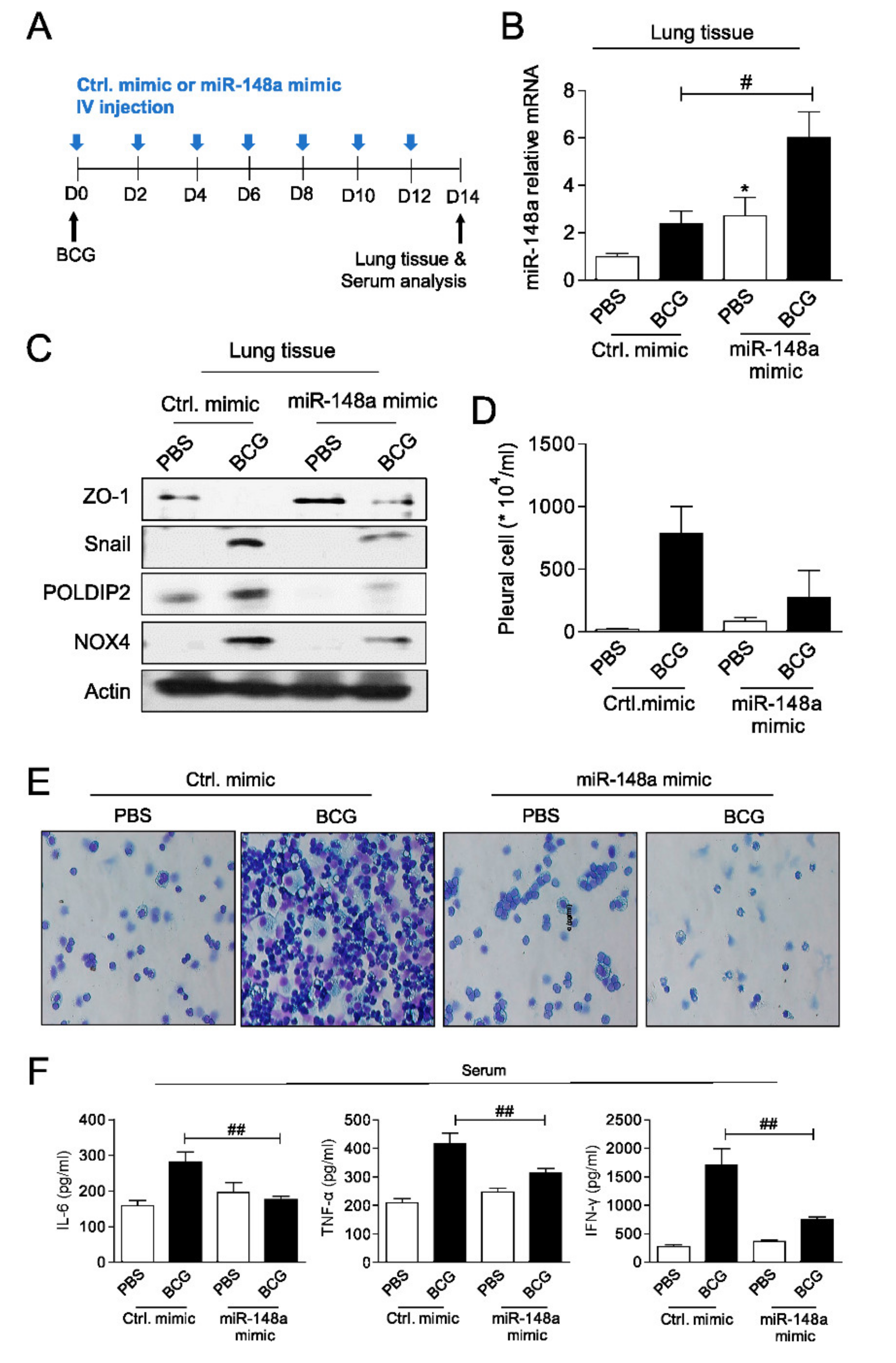
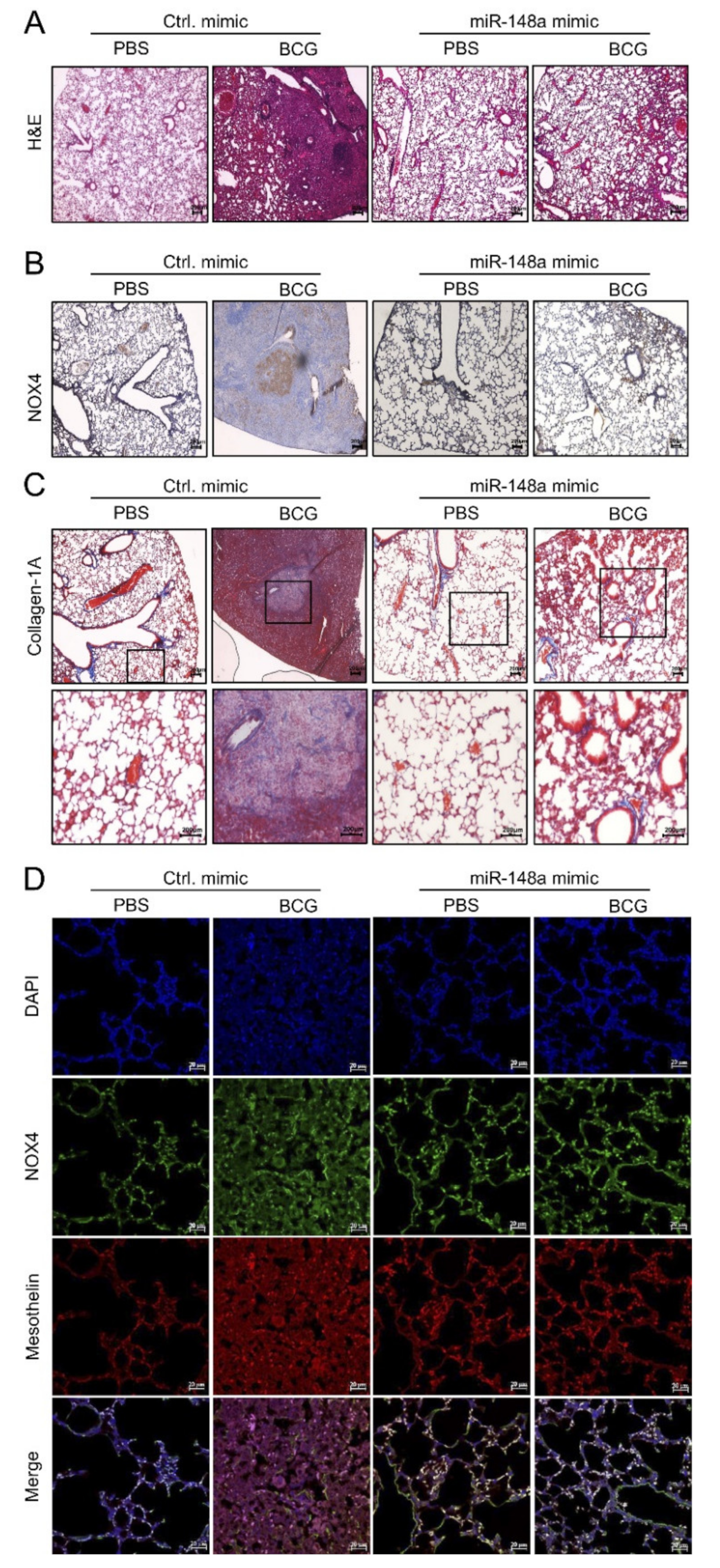
2.4. miR-148a Prevents Experimental Tuberculous Fibrosis
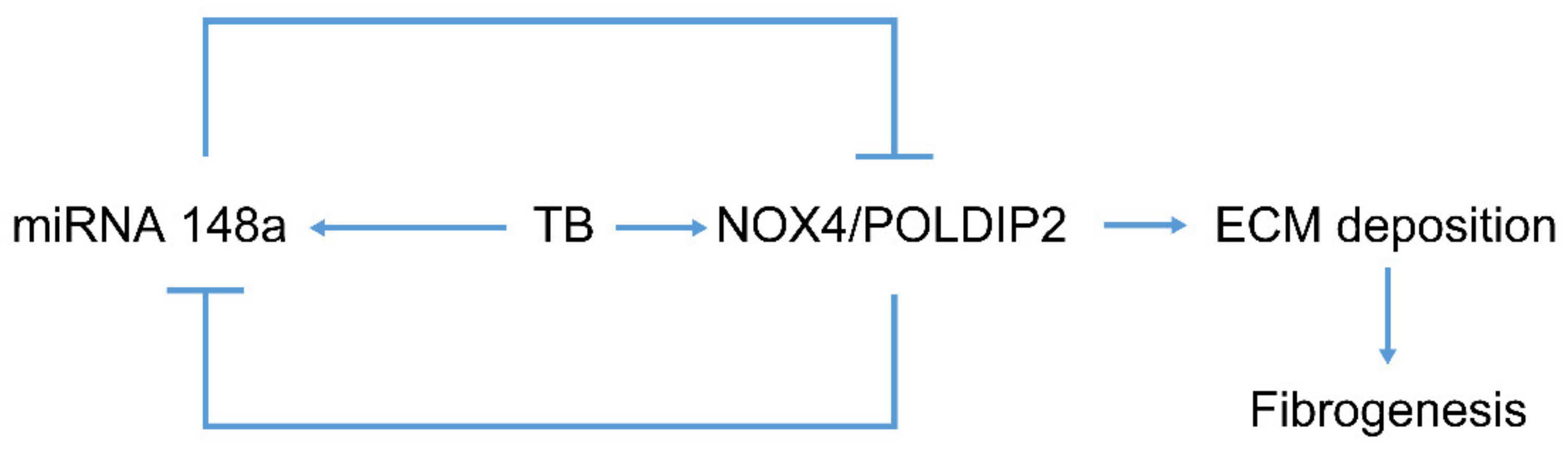
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Animals
4.2. Isolating Mouse Pleural Mesothelial Cells (mPMCs)
4.3. Treatment of Heat-Killed Mycobacterium Tuberculosis (HKMT)
4.4. The Identification of Potential miRNAs
4.5. Immunoblot and Immunoprecipitation (IP) Analysis
4.6. Quantitative Real Time Polymerase Chain Reaction (qRT-PCR)
4.7. Chromatin Immunoprecipitation Assay (ChIP Assay)
4.8. Enzyme-Linked Immunosorbent Assay (ELISA)
4.9. Establishment of BCG-Induced Pleurisy and In Vivo Treatment of miRNA Mimics
4.10. Histology, Immunohistochemistry, and Immunofluorescence Staining
4.11. Patients and Sample Collection
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Global Tuberculosis Report; World Health Organisation: Geneva, Switzerland, 2021.
- Hnizdo, E.; Singh, T.; Churchyard, G. Chronic pulmonary function impairment caused by initial and recurrent pulmonary tuberculosis following treatment. Thorax 2000, 55, 32–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasipanodya, J.G.; Miller, T.L.; Vecino, M.; Munguia, G.; Garmon, R.; Bae, S.; Drewyer, G.; Weis, S.E. Pulmonary impairment after tuberculosis. Chest 2007, 131, 1817–1824. [Google Scholar] [CrossRef] [PubMed]
- Pasipanodya, J.G.; Miller, T.L.; Vecino, M.; Munguia, G.; Bae, S.; Drewyer, G.; Weis, S.E. Using the St. George respiratory questionnaire to ascertain health quality in persons with treated pulmonary tuberculosis. Chest 2007, 132, 1591–1598. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Majano, V. Ventilation and transfer of gases in pulmonary tuberculosis. Respiration 1973, 30, 48–63. [Google Scholar] [CrossRef]
- Skoogh, B.E. Lung mechanics in pulmonary tuberculosis. I. Static lung volumes. Scand. J. Respir. Dis. 1973, 54, 148–156. [Google Scholar]
- Dheda, K.; Booth, H.; Huggett, J.F.; Johnson, M.A.; Zumla, A.; Rook, G.A. Lung remodeling in pulmonary tuberculosis. J. Infect. Dis. 2005, 192, 1201–1209. [Google Scholar] [CrossRef]
- Bocchino, M.; Agnese, S.; Fagone, E.; Svegliati, S.; Grieco, D.; Vancheri, C.; Gabrielli, A.; Sanduzzi, A.; Avvedimento, E.V. Reactive oxygen species are required for maintenance and differentiation of primary lung fibroblasts in idiopathic pulmonary fibrosis. PLoS ONE 2010, 5, e14003. [Google Scholar] [CrossRef] [Green Version]
- Hecker, L.; Vittal, R.; Jones, T.; Jagirdar, R.; Luckhardt, T.R.; Horowitz, J.C.; Pennathur, S.; Martinez, F.J.; Thannickal, V.J. NADPH oxidase-4 mediates myofibroblast activation and fibrogenic responses to lung injury. Nat. Med. 2009, 15, 1077–1081. [Google Scholar] [CrossRef] [Green Version]
- Crestani, B.; Besnard, V.; Boczkowski, J. Signalling pathways from NADPH oxidase-4 to idiopathic pulmonary fibrosis. Int. J. Biochem. Cell Biol. 2011, 43, 1086–1089. [Google Scholar] [CrossRef]
- Amara, N.; Goven, D.; Prost, F.; Muloway, R.; Crestani, B.; Boczkowski, J. NOX4/NADPH oxidase expression is increased in pulmonary fibroblasts from patients with idiopathic pulmonary fibrosis and mediates TGFbeta1-induced fibroblast differentiation into myofibroblasts. Thorax 2010, 65, 733–738. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Lan, T.; Hou, J.; Li, J.; Fang, R.; Yang, Z.; Zhang, M.; Liu, J.; Liu, B. NOX4 promotes non-small cell lung cancer cell proliferation and metastasis through positive feedback regulation of PI3K/Akt signaling. Oncotarget 2014, 5, 4392–4405. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Park, S.Y.; Jung, H.; Noh, Y.S.; Lee, J.J.; Hong, J.Y. Inhibition of NADPH oxidase 4 (NOX4) signaling attenuates tuberculous pleural fibrosis. J. Clin. Med. 2019, 8, 116. [Google Scholar] [CrossRef] [Green Version]
- Reinhart, B.J.; Slack, F.J.; Basson, M.; Pasquinelli, A.E.; Bettinger, J.C.; Rougvie, A.E.; Horvitz, H.R.; Ruvkun, G. The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature 2000, 403, 901–906. [Google Scholar] [CrossRef]
- Garzon, R.; Fabbri, M.; Cimmino, A.; Calin, G.A.; Croce, C.M. MicroRNA expression and function in cancer. Trends Mol. Med. 2006, 12, 580–587. [Google Scholar] [CrossRef]
- Chen, C.Y.; Chen, S.T.; Fuh, C.S.; Juan, H.F.; Huang, H.C. Coregulation of transcription factors and microRNAs in human transcriptional regulatory network. BMC Bioinform. 2011, 12 (Suppl. 1), S41. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Marin-Muller, C.; Bharadwaj, U.; Chow, K.H.; Yao, Q.; Chen, C. MicroRNAs: Control and loss of control in human physiology and disease. World J. Surg. 2009, 33, 667–684. [Google Scholar] [CrossRef] [Green Version]
- Sinigaglia, A.; Peta, E.; Riccetti, S.; Venkateswaran, S.; Manganelli, R.; Barzon, L. Tuberculosis-associated microRNAs: From pathogenesis to disease biomarkers. Cells 2020, 9, 2160. [Google Scholar] [CrossRef]
- Manickam, N.; Patel, M.; Griendling, K.K.; Gorin, Y.; Barnes, J.L. RhoA/Rho kinase mediates TGF-beta1-induced kidney myofibroblast activation through Poldip2/Nox4-derived reactive oxygen species. Am. J. Physiol. Ren. Physiol. 2014, 307, F159–F171. [Google Scholar] [CrossRef] [Green Version]
- Cortez, M.A.; Bueso-Ramos, C.; Ferdin, J.; Lopez-Berestein, G.; Sood, A.K.; Calin, G.A. MicroRNAs in body fluids--the mix of hormones and biomarkers. Nat. Rev. Clin. Oncol. 2011, 8, 467–477. [Google Scholar] [CrossRef] [Green Version]
- Tijsen, A.J.; Pinto, Y.M.; Creemers, E.E. Circulating microRNAs as diagnostic biomarkers for cardiovascular diseases. Am. J. Physiol. Heart Circ. Physiol. 2012, 303, H1085–H1095. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Wang, Y.; Shu, S.; Cai, J.; Tang, C.; Dong, Z. Non-coding RNAs in kidney injury and repair. Am. J. Physiol. Cell Physiol. 2019, 317, C177–C188. [Google Scholar] [CrossRef]
- Usman, K.; Hsieh, A.; Hackett, T.L. The role of miRNAs in extracellular matrix repair and chronic fibrotic lung diseases. Cells 2021, 10, 1706. [Google Scholar] [CrossRef]
- Li, J.; Yu, T.; Cao, J.; Liu, L.; Liu, Y.; Kong, H.W.; Zhu, M.X.; Lin, H.C.; Chu, D.D.; Yao, M.; et al. MicroRNA-148a suppresses invasion and metastasis of human non-small-cell lung cancer. Cell. Physiol. Biochem. 2015, 37, 1847–1856. [Google Scholar] [CrossRef]
- Xiong, J.; Ni, J.; Chen, C.; Wang, K. miR148a-3p regulates alcoholic liver fibrosis through targeting ERBB3. Int. J. Mol. Med. 2020, 46, 1003–1012. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Min, L.; Ren, C.; Xu, X.; Yang, J.; Sun, X.; Wang, T.; Wang, F.; Sun, C.; Zhang, X. miRNA-148a serves as a prognostic factor and suppresses migration and invasion through Wnt1 in non-small cell lung cancer. PLoS ONE 2017, 12, e0171751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miotto, P.; Mwangoka, G.; Valente, I.C.; Norbis, L.; Sotgiu, G.; Bosu, R.; Ambrosi, A.; Codecasa, L.R.; Goletti, D.; Matteelli, A.; et al. miRNA signatures in sera of patients with active pulmonary tuberculosis. PLoS ONE 2013, 8, e80149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.; Haigh, S.; Barman, S.; Fulton, D.J. From form to function: The role of Nox4 in the cardiovascular system. Front. Physiol. 2012, 3, 412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anrather, J.; Racchumi, G.; Iadecola, C. NF-kappaB regulates phagocytic NADPH oxidase by inducing the expression of gp91phox. J. Biol. Chem. 2006, 281, 5657–5667. [Google Scholar] [CrossRef] [Green Version]
- Ryu, J.; Lee, C.W.; Shin, J.A.; Park, C.S.; Kim, J.J.; Park, S.J.; Han, K.H. FcgammaRIIa mediates C-reactive protein-induced inflammatory responses of human vascular smooth muscle cells by activating NADPH oxidase 4. Cardiovasc. Res. 2007, 75, 555–565. [Google Scholar] [CrossRef]
- Lu, X.; Murphy, T.C.; Nanes, M.S.; Hart, C.M. PPAR{gamma} regulates hypoxia-induced Nox4 expression in human pulmonary artery smooth muscle cells through NF-{kappa}B. Am. J. Physiol. Lung Cell. Mol. Physiol. 2010, 299, L559–L566. [Google Scholar] [CrossRef] [Green Version]
- Woo, S.J.; Kim, Y.; Jung, H.; Lee, J.J.; Hong, J.Y. Tuberculous fibrosis enhances tumorigenic potential via the NOX4-autophagy axis. Cancers 2021, 13, 687. [Google Scholar] [CrossRef]
- Liu, J.; Livingston, M.J.; Dong, G.; Tang, C.; Su, Y.; Wu, G.; Yin, X.M.; Dong, Z. Histone deacetylase inhibitors protect against cisplatin-induced acute kidney injury by activating autophagy in proximal tubular cells. Cell Death Dis. 2018, 9, 322. [Google Scholar] [CrossRef]
- Liu, Z.; Tang, C.; He, L.; Yang, D.; Cai, J.; Zhu, J.; Shu, S.; Liu, Y.; Yin, L.; Chen, G.; et al. The negative feedback loop of NF-κB/miR-376b/NFKBIZ in septic acute kidney injury. JCI Insight 2020, 5, e142272. [Google Scholar] [CrossRef]
- Light, R.W.; Macgregor, M.I.; Luchsinger, P.C.; Ball, W.C. Pleural effusions: The diagnostic separation of transudates and exudates. Ann. Intern. Med. 1972, 77, 507–513. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences (5′–3′) |
|---|---|
| Hu.NOX4 | FOR: CCGGCTGCATCAGTCTTAAC |
| REV: TCGGCACAGTACAGGCACAA | |
| Hu.Poldip2 | FOR:CAAAACAGAATGGAAAATATGAGACCGG |
| REV: TGATTGATGCTCGTGACTGCCCA | |
| Hu.Col1A1 | FOR: ACTGGTGAGACCTGCGTGTA |
| REV: AATCCATCGGTCATGCTCTC | |
| Hu.β-actin | FOR: GTGCTATCCCTGTACGCCTC |
| REV: GGCCATCTCTTGCTCGAAGT | |
| Mu.NOX4 | FOR: CATTCACCAAATGTTGGGC |
| REV: TGCACACCTGAGAAAATACA | |
| Mu.Poldip2 | FOR: TCCTCAGAGGCTGGACATCT |
| REV:ATGCTCAGAAGCCCACAGTT | |
| Mu.GAPDH | FOR: CGTCCCGTAGACAAAATGGT |
| REV: TTGATGGCAACAATCTCCAC |
| Gene | Primer Sequences (5′–3′) |
|---|---|
| has-miR-215 | ATGACCTATGAATTGACAGAC |
| has-miR-9 | TCTTTGGTTATCTAGCTGTATGA |
| has-miR-196b | TAGGTAGTTTCCTGTTGTTGGG |
| has-miR-203 | GTGAAATGTTTAGGACCACTAG |
| has-miR-148a | AAAGTTCTGAGACACTCCGACT |
| has-U6 | CTCGCTTCGGCAGCACA |
| has-miR-215 | ATGACCTATGATTTGACAGAC |
| has-miR-9 | TCTTTGGTTATCTAGCTGTATG |
| has-miR-196b | TAGGTAGTTTCCTGTTGTTGGG |
| has-miR-203 | GTGAAATGTTTAGGACCACTAG |
| has-miR-148a | AAAGTTCTGAGACACTCCGACT |
| has-U6 | CTCGCTTCGGCAGCACA |
| mmu-miR-215 | ATGACCTATGATTTGACAGAC |
| mmu-miR-9 | TCTTTGGTTATCTAGCTGTATGA |
| mmu-miR-196b | TAGGTAGTTTCCTGTTGTTGGG |
| mmu-miR-203 | GTGAAATGTTTAGGACCACTAG |
| mmu-miR148a | TCAGTGCACTACAGAACTTTGT |
| mmu-U6 | GCTTCGGCAGCACATATACTAAAAT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Woo, S.J.; Kim, Y.; Jung, H.; Lee, J.J.; Hong, J.Y. MicroRNA 148a Suppresses Tuberculous Fibrosis by Targeting NOX4 and POLDIP2. Int. J. Mol. Sci. 2022, 23, 2999. https://doi.org/10.3390/ijms23062999
Woo SJ, Kim Y, Jung H, Lee JJ, Hong JY. MicroRNA 148a Suppresses Tuberculous Fibrosis by Targeting NOX4 and POLDIP2. International Journal of Molecular Sciences. 2022; 23(6):2999. https://doi.org/10.3390/ijms23062999
Chicago/Turabian StyleWoo, Seong Ji, Youngmi Kim, Harry Jung, Jae Jun Lee, and Ji Young Hong. 2022. "MicroRNA 148a Suppresses Tuberculous Fibrosis by Targeting NOX4 and POLDIP2" International Journal of Molecular Sciences 23, no. 6: 2999. https://doi.org/10.3390/ijms23062999
APA StyleWoo, S. J., Kim, Y., Jung, H., Lee, J. J., & Hong, J. Y. (2022). MicroRNA 148a Suppresses Tuberculous Fibrosis by Targeting NOX4 and POLDIP2. International Journal of Molecular Sciences, 23(6), 2999. https://doi.org/10.3390/ijms23062999