
Isolation and Functional Characterization of Two CONSTANS-like 16 (MiCOL16) Genes from Mango


Abstract
:1. Introduction
2. Results
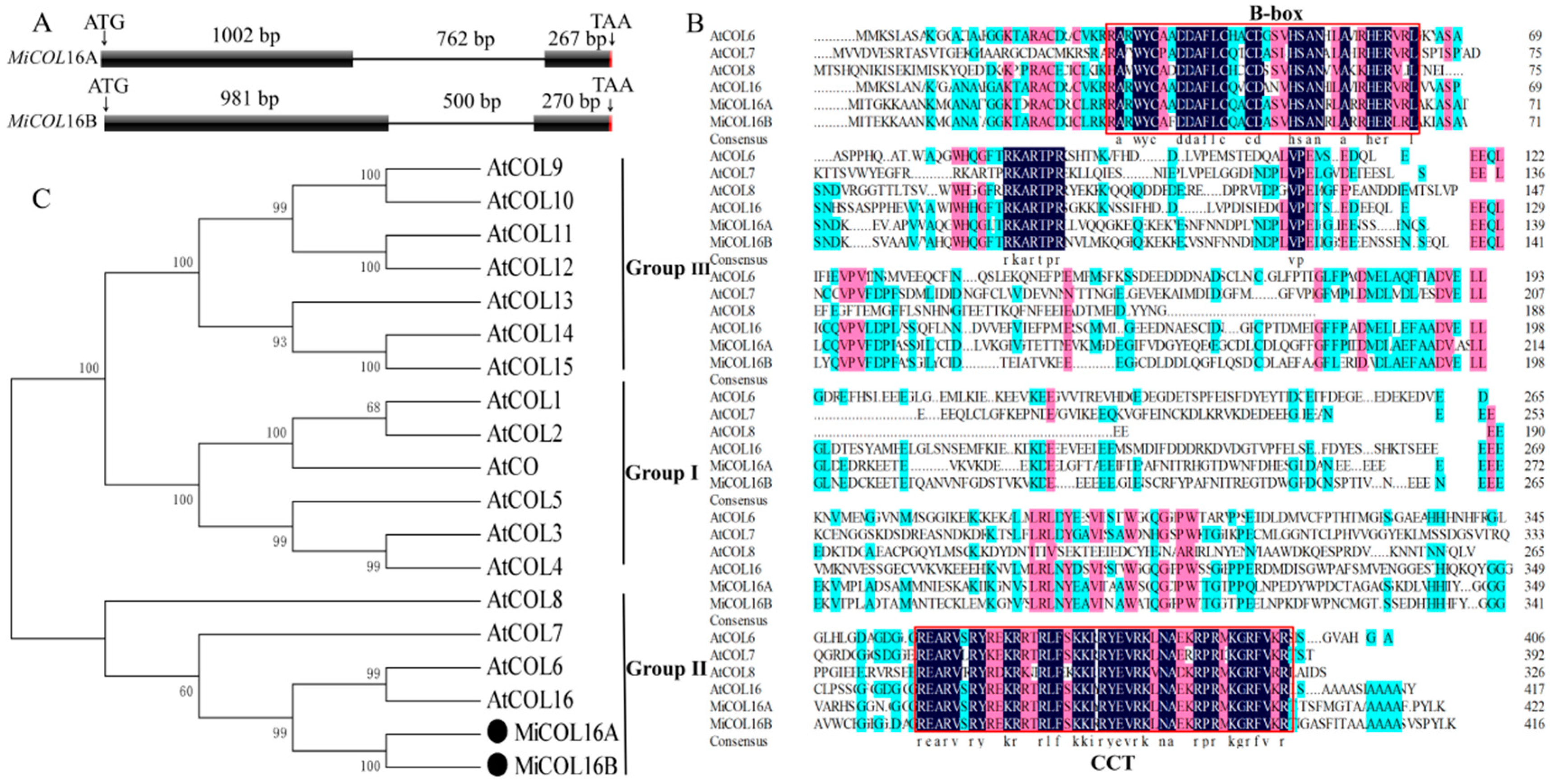
2.1. Isolation and Analysis of MiCOL16A and MiCOL16B
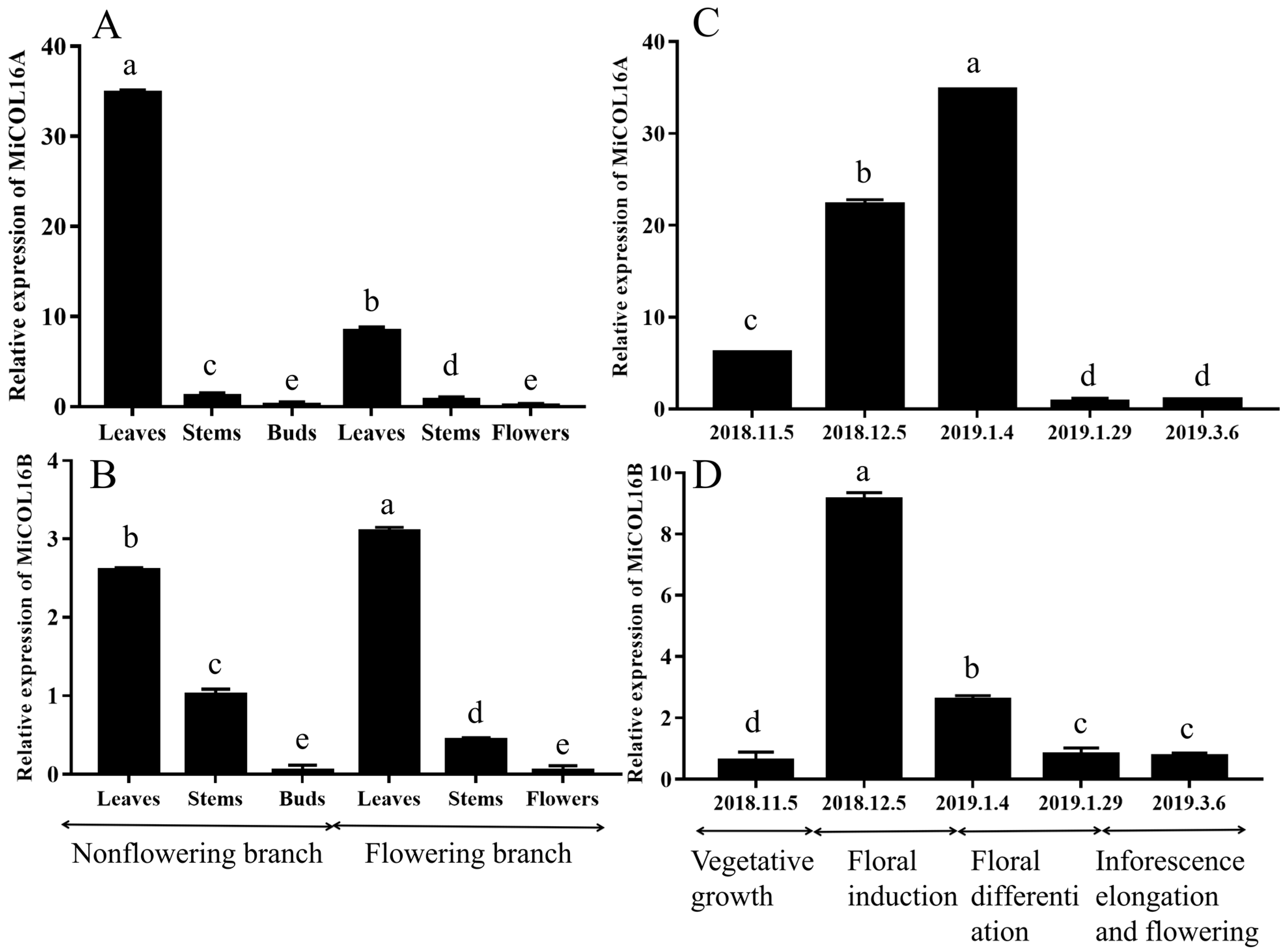
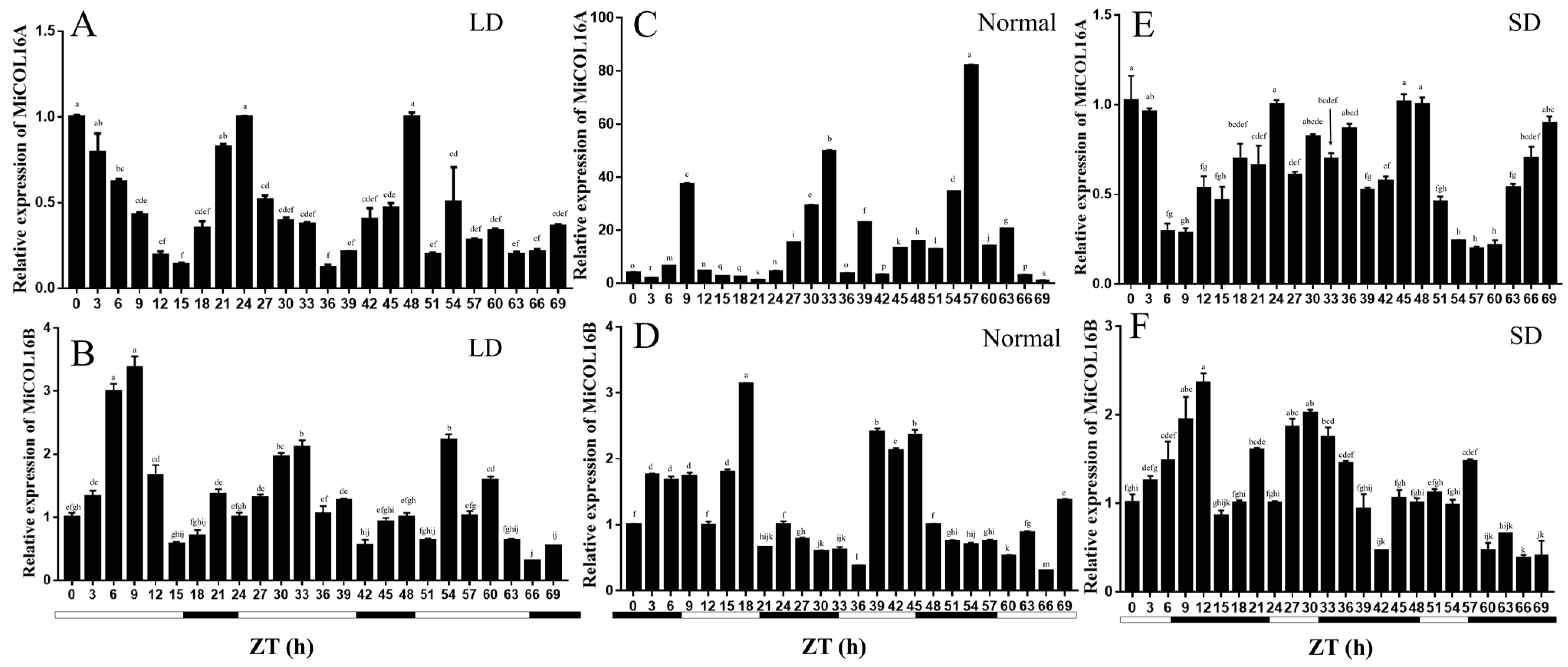
2.2. Expression of MiCOL16A and MiCOL16B in Mango
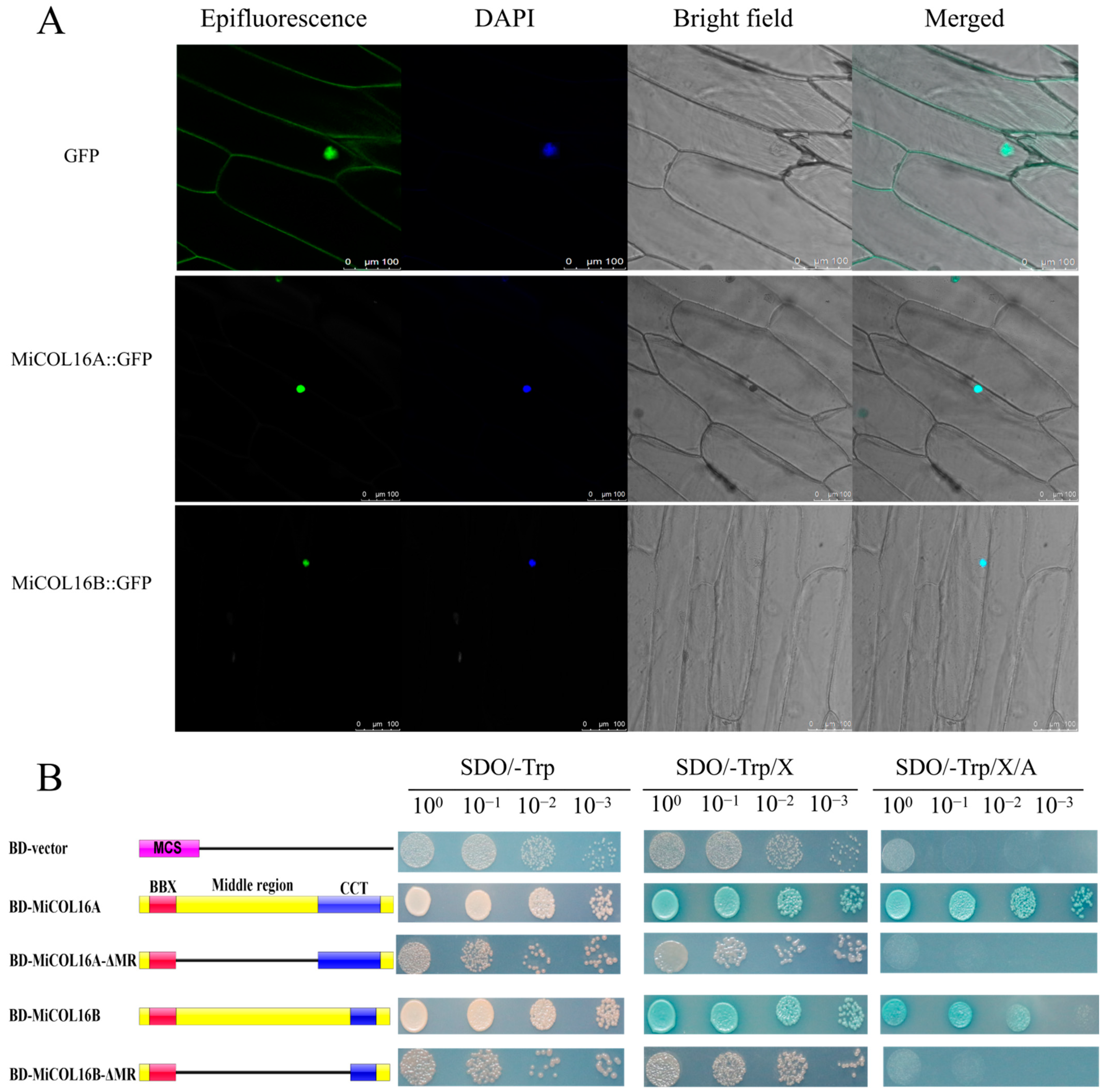
2.3. Both MiCOL16A and MiCOL16B Are Nuclear Proteins with Transcriptional Activation Activity
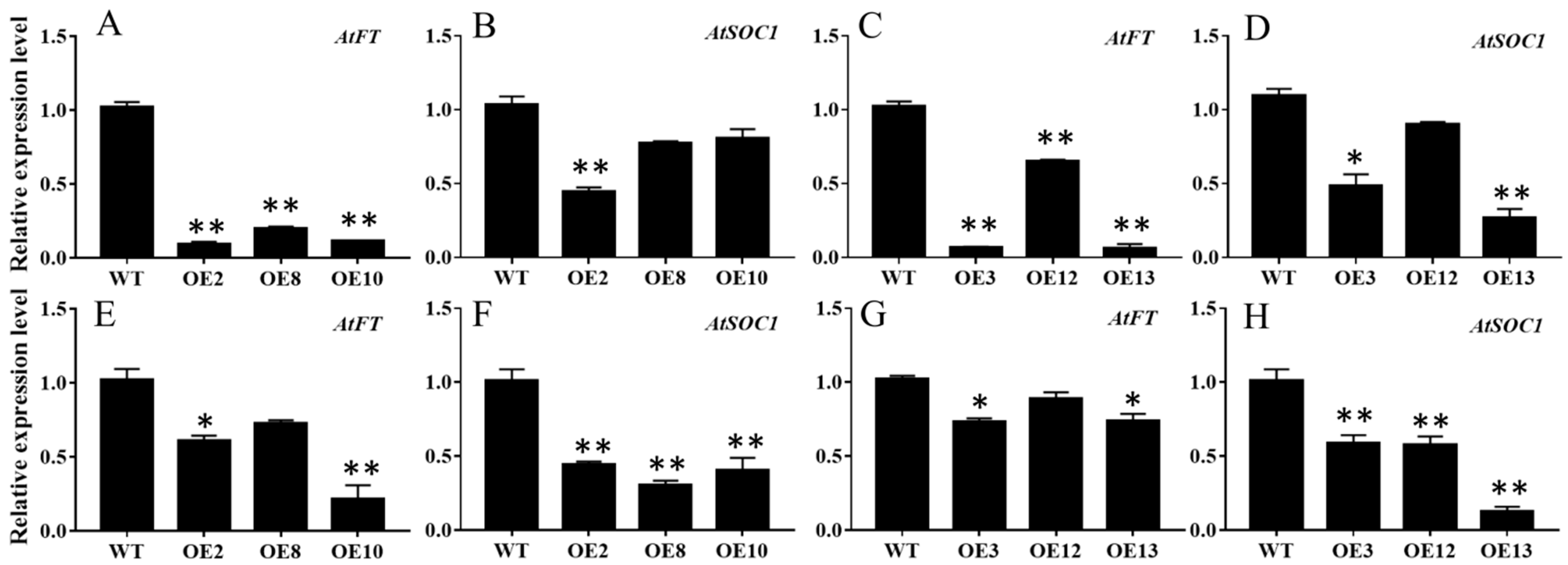
2.4. Overexpression of MiCOL16A and MiCOL16B Delayed Flowering in Arabidopsis
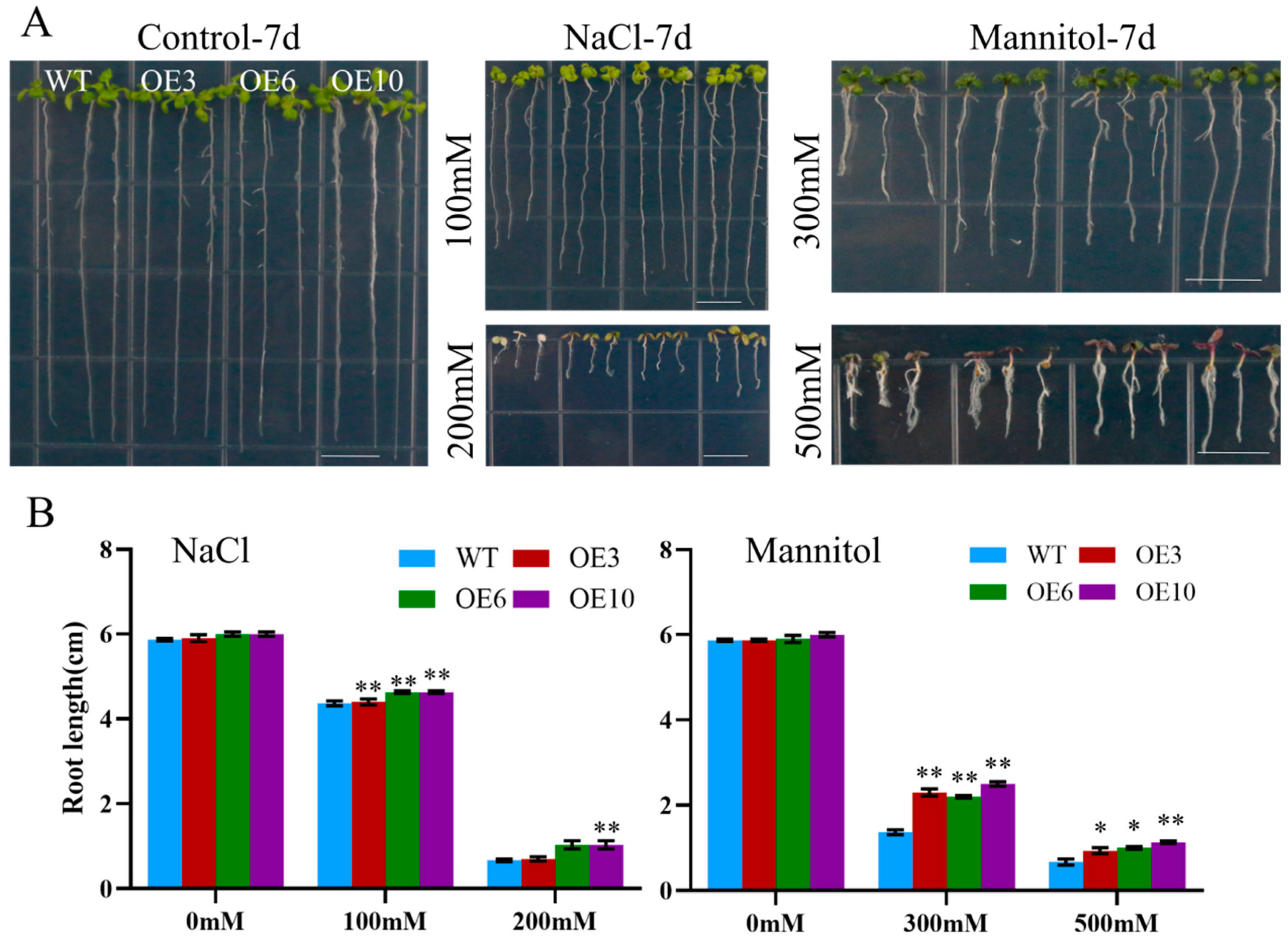
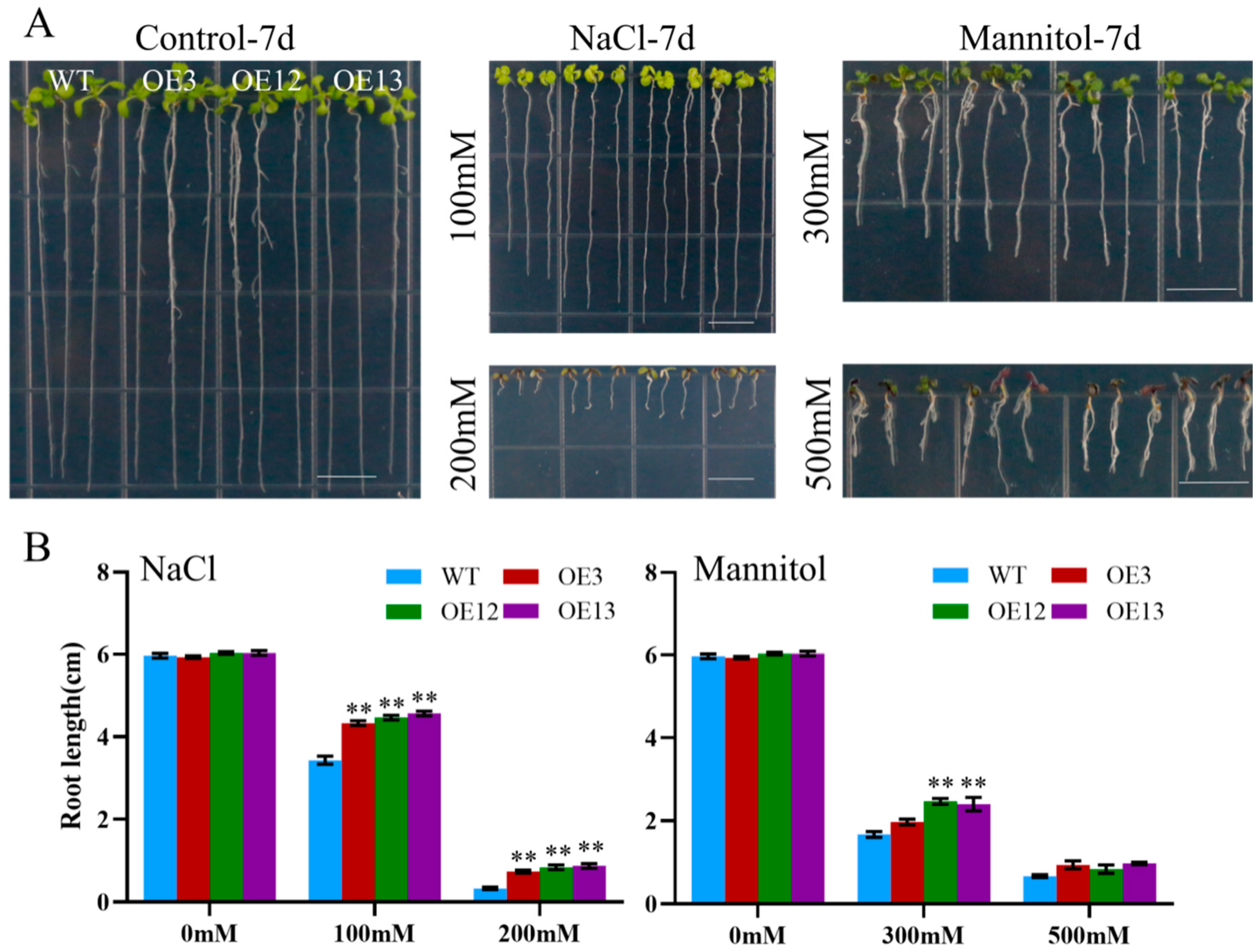
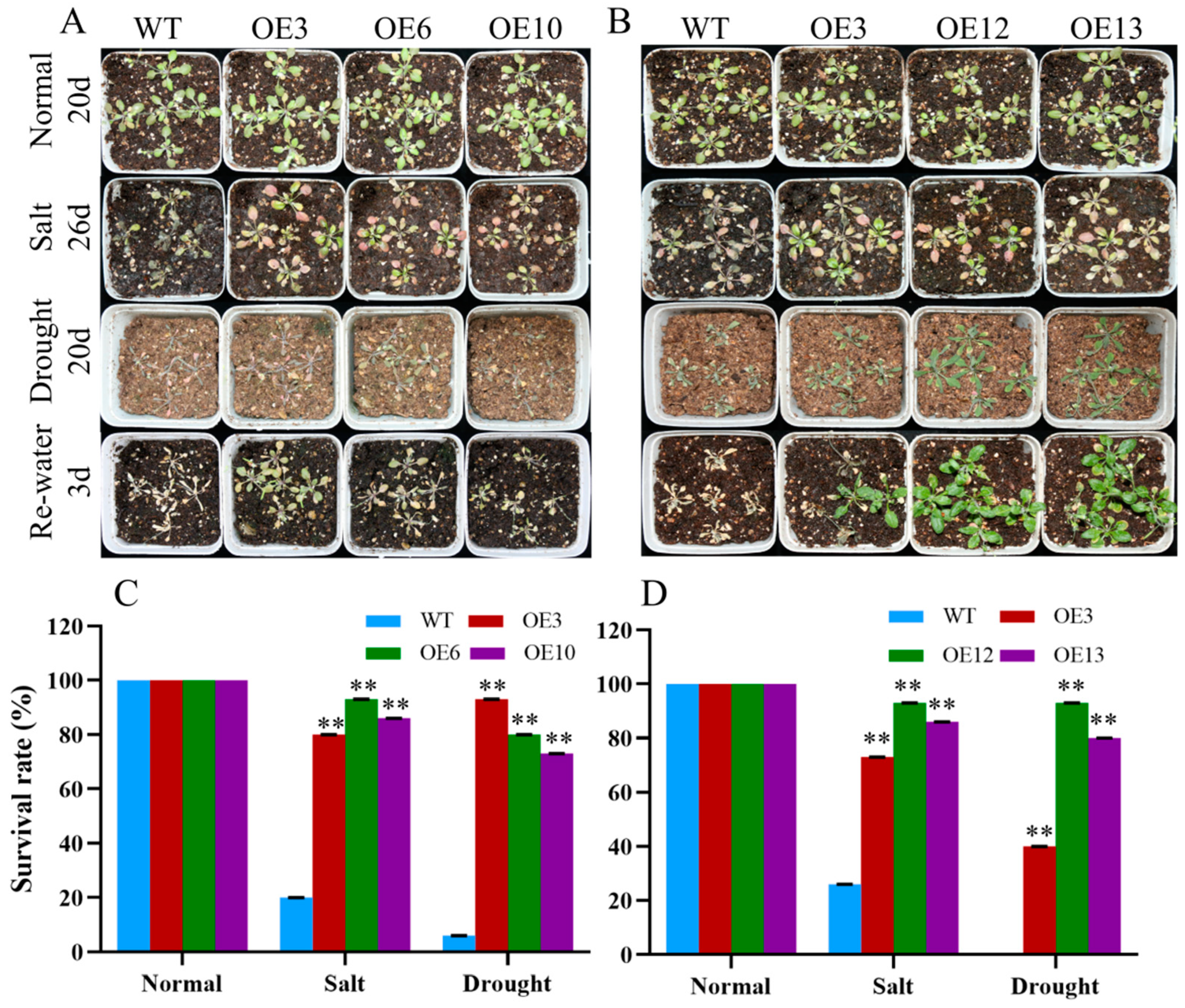
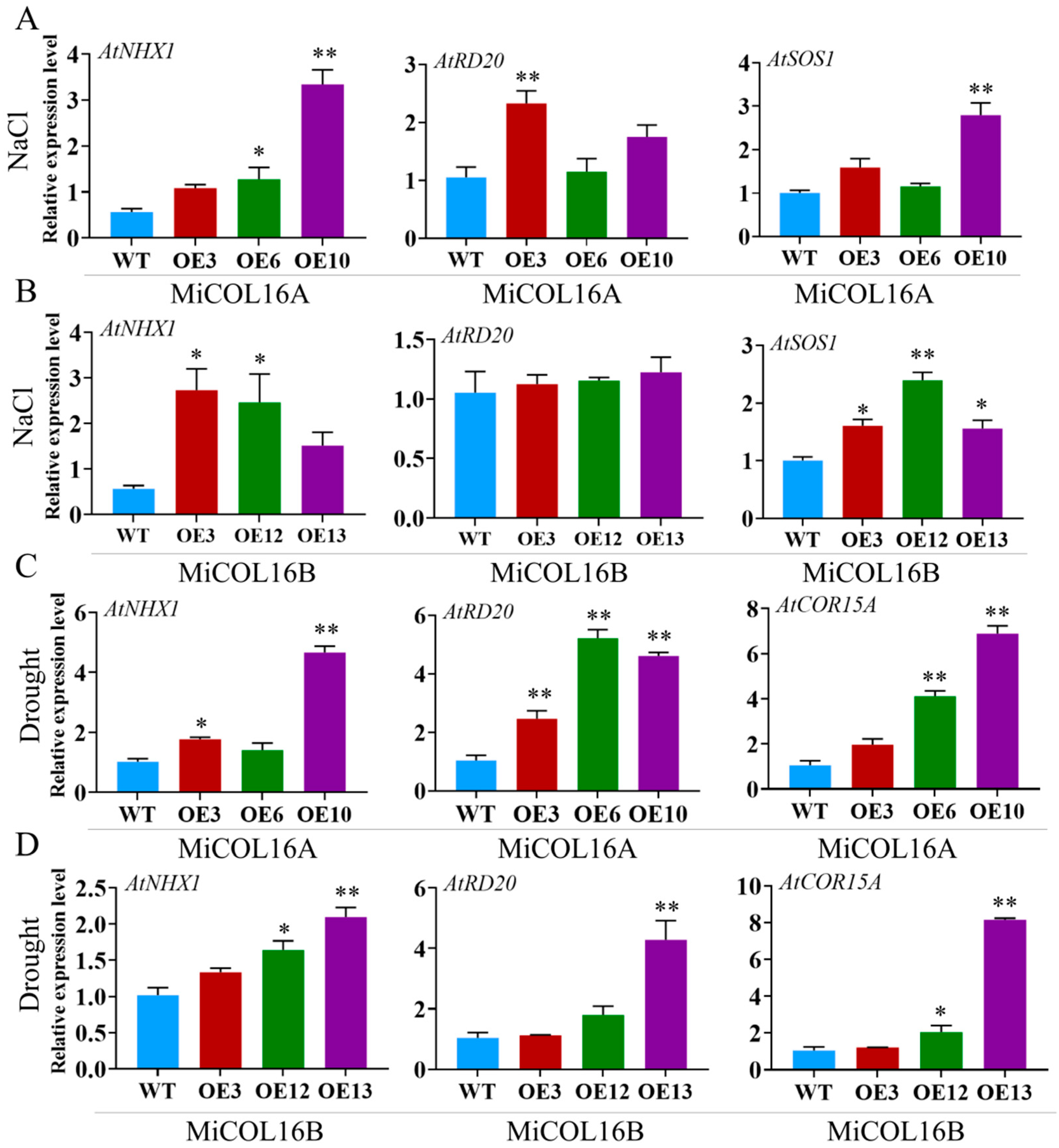
2.5. MiCOL16A and MiCOL16B Enhance Tolerance to Abiotic Stress
3. Discussion
4. Material and Methods
4.1. Materials
4.2. Identification and Sequence Analysis
4.3. qRT–PCR Analysis
4.4. Subcellular Localization
4.5. Analysis of Transcriptional Activity of MiCOL16A and MiCOL16B
4.6. Vector Construction and Arabidopsis Plant Transformation
4.7. Stress Treatments of Transgenic and WT Plants
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board statement
Informed Consent Statement
Conflicts of Interest
References
- Okamuro, J.K.; Szeto, W.; Lotys-Prass, C.; Jofuku, K.D. Photo and hormonal control of meristem identity in the Arabidopsis flower mutants apetala2 and apetala1. Plant Cell 1997, 9, 37–47. [Google Scholar] [PubMed] [Green Version]
- Wu, G.; Park, M.Y.; Conway, S.R.; Wang, J.W.; Weigel, D.; Poethig, R.S. The sequential action of miR156 and miR172 regulates developmental timing in Arabidopsis. Cell 2009, 138, 750–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blazquez, M.A.; Green, R.; Nilsson, O.; Sussman, M.R.; Weigel, D. Gibberellins promote flowering of Arabidopsis by activating the LEAFY promoter. Plant Cell 1998, 10, 791–800. [Google Scholar] [CrossRef] [Green Version]
- Huang, G.W.; Ma, J.H.; Han, Y.Z.; Chen, X.J.; Fu, Y.F. Cloning and expression analysis of the soybean CO-like gene GmCOL9. Plant Mol. Biol. Rep. 2010, 29, 352–359. [Google Scholar] [CrossRef]
- Komeda, Y. Genetic regulation of time to flower in Arabidopsis thaliana. Annu. Rev. Plant Biol. 2004, 55, 521–553. [Google Scholar] [CrossRef] [Green Version]
- Putterill, J.; Robson, F.; Lee, K.; Simon, R.; Coupland, G. The CONSTANS gene of Arabidopsis promotes flowering and encodes a protein showing similarities to zinc finger transcription factors. Cell 1995, 80, 847–857. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, S.; Dunford, R.P.; Coupland, G.; Laurie, D.A. The evolution of CONSTANS-like gene families in barley, rice, and Arabidopsis. Plant Physiol. 2003, 131, 1855–1867. [Google Scholar] [CrossRef] [Green Version]
- Yano, M.; Katayose, Y.; Ashikari, M.; Yamanouchi, U.; Monna, L.; Fuse, T.; Baba, T.; Yamamoto, K.; Umehara, Y.; Nagamura, Y.; et al. Hd1, a major photoperiod sensitivity quantitative trait locus in rice, is closely related to the Arabidopsis flowering time gene CONSTANS. Plant Cell 2000, 12, 2473–2484. [Google Scholar] [CrossRef] [Green Version]
- Serrano, G.; Herrera-Palau, R.; Romero, J.M.; Serrano, A.; Coupland, G.; Valverde, F. Chlamydomonas CONSTANS and the evolution of plant photoperiodic signaling. Curr. Biol. 2009, 19, 359–368. [Google Scholar] [CrossRef]
- Imaizumi, T.; Schultz, T.F.; Harmon, F.G.; Ho, L.A.; Kay, S.A. FKF1 F-box protein mediates cyclic degradation of a repressor of CONSTANS in Arabidopsis. Science 2005, 309, 293–297. [Google Scholar] [CrossRef]
- Sawa, M.; Nusinow, D.A.; Kay, S.A.; Imaizumi, T. FKF1 and GIGANTEA complex formation is required for day-length measurement in Arabidopsis. Science 2007, 318, 261–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imaizumi, T.; Kay, S.A. Photoperiodic control of flowering: Not only by coincidence. Trends Plant Sci. 2006, 11, 550–558. [Google Scholar] [CrossRef] [PubMed]
- Wigge, P.A.; Kim, M.C.; Jaeger, K.E.; Busch, W.; Schmid, M.; Lohmann, J.U.; Weigel, D. Integration of spatial and temporal information during floral induction in Arabidopsis. Science 2005, 309, 1056–1059. [Google Scholar] [CrossRef] [PubMed]
- Shogo, I.; Young, H.S.; Anna, R.J.; Ryan, J.M.; Ghislain, B.; Richard, G.O.; Takato, I. FLOWERING BHLH transcriptional activators control expression of the photoperiodic flowering regulator CONSTANS in Arabidopsis. Proc. Natl. Acad. Sci. USA 2012, 109, 3582–3587. [Google Scholar]
- Sun, J.J.; Lu, J.; Bai, M.J.; Chen, Y.Q.; Wang, W.N.; Fan, C.G.; Liu, J.Y.; Ning, G.G.; Wang, C.Q. Phytochrome-interacting factors interact with transcription factor CONSTANS to suppress flowering in rose. Plant Physiol. 2021, 186, 1186–1201. [Google Scholar] [CrossRef]
- Ana, L.; Federico, V.; Manuel, P.; Jose, A.J. The Arabidopsis E3 ubiquitin ligase HOS1 negatively regulates CONSTANS abundance in the photoperiodic control of flowering. Plant Cell 2012, 24, 982–999. [Google Scholar]
- Liu, L.J.; Zhang, Y.C.; Li, Q.H.; Sang, Y.; Mao, J.; Lian, H.L.; Wang, L.; Yang, H.Q. COP1-Mediated ubiquitination of CONSTANS is implicated in cryptochrome regulation of flowering in Arabidopsis. Plant Cell 2008, 20, 292–306. [Google Scholar] [CrossRef]
- Yang, N.; Cong, Q.; Cheng, L.J. BBX transcriptional factors family in plants-a review. Chin. J. Biotechnol. 2019, 36, 666–677. [Google Scholar]
- Ledger, S.; Strayer, C.; Ashton, F.; Kay, S.A.; Putterill, J. Analysis of the function of two circadian-regulated CONSTANS-LIKE genes. Plant J. 2001, 26, 15–22. [Google Scholar] [CrossRef]
- Graeff, M.; Straub, D.; Eguen, T.; Dolde, U.; Rodrigues, V.; Brandt, R.; Wenkel, S. Microprotein-mediated recruitment of CONSTANS into a topless trimeric complex represses flowering in Arabidopsis. PLoS Genet. 2016, 12, e1005959. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.G.; Zhang, Z.L.; Li, H.Y.; Zhao, X.Y.; Liu, X.M.; Ortiz, M.; Lin, C.T.; Liu, B. CONSTANS-LIKE 7 regulates branching and shade avoidance response in Arabidopsis. J. Exp. Bot. 2013, 64, 1017–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takase, T.; Kakikubo, Y.; Nakasone, A.; Nishiyama, Y.; Yasuhara, M.; Yoko, T.; Kiyosue, T. Characterization and transgenic study of CONSTANS-LIKE8 (COL8) gene in Arabidopsis thaliana: Expression of 35S:COL8 delays flowering under long-day conditions. Plant Biotechnol. 2011, 28, 439–446. [Google Scholar] [CrossRef] [Green Version]
- Tsuji, H.; Taoka, K.; Shimamoto, K. Regulation of flowering in rice: Two florigen genes, a complex gene network, and natural variation. Curr. Opin. Plant Biol. 2011, 14, 45–52. [Google Scholar] [CrossRef]
- Wu, W.X.; Zheng, X.M.; Chen, D.B.; Zhang, Y.X.; Ma, W.W.; Zhang, H.; Sun, L.P.; Yang, Z.F.; Zhao, C.D.; Zhan, X.D.; et al. OsCOL16, encoding a CONSTANS-like protein, represses flowering by upregulating Ghd7 expression in rice. Plant Sci. 2017, 260, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Schain, N.D.; Dıaz-Mendoza, M.; Zurczak, M.; Suárez-López, P. Potato CONSTANS is involved in photoperiodic tuberization in a graft-transmissible manner. Plant J. 2012, 70, 678–690. [Google Scholar] [CrossRef] [PubMed]
- Jeong, D.H.; Sung, S.K.; An, G. Molecular cloning and characterization of CONSTANS-like cDNA clones of the Fuji apple. J Plant Biol. 1999, 42, 23–31. [Google Scholar] [CrossRef]
- Luo, C.; Yu, H.X.; Fan, Y.; Zhang, X.J.; He, X.H. Research advance on the flowering mechanism of mango. Acta Hortic. 2019, 1244, 2. [Google Scholar] [CrossRef]
- Fan, Z.Y.; He, X.H.; Fan, Y.; Yu, H.X.; Wang, Y.H.; Xie, X.J.; Liu, Y.; Mo, X.; Wang, J.Y.; Luo, C. Isolation and functional characterization of three MiFTs genes from mango. Plant Physiol. Biochem. 2020, 155, 169–176. [Google Scholar] [CrossRef]
- Wei, J.Y.; Liu, D.B.; Liu, G.Y.; Tang, J.; Chen, Y.Y. Molecular cloning, characterization, and expression of MiSOC1: A homolog of the flowering gene SUPPRESSOR OF overexpression OF CONSTANS1 from mango (Mangifera indica L). Front. Plant Sci. 2016, 7, 1758. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.X.; Luo, C.; Fan, Y.; Zhang, X.J.; Huang, F.; Li, M.; He, X.H. Isolation and characterization of two APETALA1-Like genes from mango (Mangifera indica L.). Sci. Hortic. 2020, 259, 108814. [Google Scholar] [CrossRef]
- Liu, Y.; Luo, C.; Zhang, X.J.; Lu, X.X.; Yu, H.X.; Xie, X.J.; Fan, Z.Y.; Mo, X.; He, X.H. Overexpression of the mango MiCO gene delayed flowering time in transgenic Arabidopsis. Plant Cell Tissue Organ Cult. 2020, 143, 219–228. [Google Scholar] [CrossRef]
- Tan, J.J.; Jin, M.N.; Wang, J.C.; Wu, F.Q.; Sheng, P.K.; Cheng, Z.J.; Wang, J.L.; Zheng, X.M.; Chen, L.P.; Wang, M.; et al. OsCOL10, a CONSTANS-Like gene, functions as a flowering time repressor downstream of Ghd7 in rice. Plant Cell Physiol. 2016, 57, 798–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.X.; Zhang, Y.X.; Zhang, M.; Zhan, X.D.; Shen, X.H.; Yu, P.; Chen, D.B.; Liu, Q.E.; Sinumporn, S.; Hussain, K.; et al. The rice CONSTANS-like protein OsCOL15 suppresses flowering by promoting Ghd7 and repressing RID1. Biochem. Biophys. Res. Commun. 2018, 495, 1349–1355. [Google Scholar] [CrossRef]
- Valverde, F. CONSTANS and the evolutionary origin of photoperiodic timing of flowering. J. Exp. Bot. 2011, 62, 2453–2463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, J.; Mao, D.; Liu, X.M.; Wang, L.L.; Xu, F. Isolation and functional characterization of a circadian-regulated CONSTANS homolog (GbCO) from Ginkgo biloba. Plant Cell Rep. 2017, 36, 1387–1399. [Google Scholar] [CrossRef]
- Samach, A.; Onouchi, H.; Gold, S.E.; Ditta, G.S.; Schwarz-Sommer, Z.; Yanofsky, M.F.; Coupland, G. Distinct roles of CONSTANS target genes in reproductive development of Arabidopsis. Science 2000, 288, 1613–1616. [Google Scholar] [CrossRef] [Green Version]
- Sawa, M.; Kay, S.A. GIGANTEA directly activates FLOWERING LOCUS T in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2011, 108, 11698–11703. [Google Scholar] [CrossRef] [Green Version]
- Turck, F.; Fornara, F.; Coupland, G. Regulation and identity of florigen: FLOWERING LOCUS T moves center stage. Annu. Rev. Plant Biol. 2008, 59, 573–594. [Google Scholar] [CrossRef] [Green Version]
- Bohlenius, H.; Huang, T.; Charbonnelcampaa, L.; Brunner, A.M.; Jansson, S.; Strauss, S.H.; Nilsson, O. CO/FT regulatory module controls timing of flowering and seasonal growth cessation in trees. Science 2006, 312, 1040–1043. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.L.; Yan, J.P.; Meng, X.X.; Ye, J.B.; Zhang, W.W.; Xu, F. Cloning and expression analysis of CONSTANS-like 16 (GbCOL16) gene from Ginkgo biloba. BMC Biotechnol. 2017, 16, 92–99. [Google Scholar] [CrossRef]
- Chia, T.Y.P.; Muller, A.; Jung, C.; Mutasa-Gottgens, E.S. Sugar beet contains a large CONSTANS-like gene family including a CO homologue that is independent of the early-bolting (B) gene locus. J. Exp. Bot. 2008, 59, 2735–2748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akemi, O.; Chihiro, O.Y.; Sanae, K. Overexpression of CONSTANS-like 16 enhances chlorophyll accumulation in petunia corollas. Plant Sci. 2019, 280, 90–96. [Google Scholar]
- Wang, L.; Xue, J.; Dai, W.; Tang, Y.; Gong, P.; Wang, Y.; Zhang, C. Genome-wide identification, phylogenetic analysis, and expression profiling of CONSTANS-like (COL) genes in vitis vinifera. J. Plant Growth Regul. 2019, 38, 631–643. [Google Scholar] [CrossRef]
- Suarez-Lopez, P.; Wheatley, K.; Robson, F. CONSTANS mediates between the circadian clock and the control of flowering in Arabidopsis. Nature 2001, 410, 1116–1120. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.X.; Zhao, X.L.; Tian, R.X.; Zeng, S.J.; Wu, K.L.; Silva, J.A.; Duan, J. Molecular cloning and functional analysis of three CONSTANS-Like genes from Chinese Cymbidium. J. Plant Growth Regul. 2019, 39, 1061–1074. [Google Scholar] [CrossRef]
- Zhang, Z.; Ji, R.; Li, H.; Tao, Z.; Liu, J.; Lin, C.; Liu, B. CONSTANS-LIKE 7 (COL7) is involved in phytochrome B (phyB)-mediated light-quality regulation of auxin homeostasis. Mol. Plant 2014, 7, 1674–2052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.H.; Shim, J.S.; Kinmonth-Schultz, H.A.; Imaizumi, T. Photoperiodic flowering: Time measurement mechanisms in leaves. Annu. Rev. Plant Biol. 2015, 66, 441–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinbach, Y. The Arabidopsis thaliana CONSTANS-LIKE 4 (COL4) a modulator of flowering time. Front. Plant Sci. 2019, 10, 651. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.K.; Yun, C.H.; Lee, J.H.; Jang, Y.H.; Park, H.Y.; Kim, J.K. OsCO3, a CONSTANS-LIKE gene, controls flowering by negatively regulating the expression of FT-like genes under SD conditions in rice. Planta 2008, 228, 355–365. [Google Scholar] [CrossRef]
- Kardailsky, I.; Shukla, V.K.; Ahn, J.H.; Dagenais, N.; Christensen, S.K.; Nguyen, J.T.; Chory, J.; Harrison, M.J.; Weigel, D. Activation tagging of the floral inducer FT. Science 1999, 286, 1962–1965. [Google Scholar] [CrossRef] [Green Version]
- Luccioni, L.; Krzymuski, M.; Sanchez-Lamas, M.; Karayekov, E.; Cerdan, P.D.; Casal, J.J. CONSTANS delays Arabidopsis flowering under short days. Plant J. 2019, 97, 923–932. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.F.; Wang, Z.Y. Overexpression of COL9, a CONSTANS-LIKE gene, delays flowering by reducing expression of CO and FT in Arabidopsis thaliana. Plant J. 2005, 43, 758–768. [Google Scholar] [CrossRef] [PubMed]
- Xiao, G.H.; Li, B.J.; Chen, H.J.; Wang, Z.Y.; Mao, B.Z.; Gui, R.Y.; Guo, X.Q. Overexpression of PvCO1, a bamboo CONSTANS-LIKE gene, delays flowering by reducing expression of the FT gene in transgenic Arabidopsis. BMC Plant Biol. 2018, 18, 232. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Gu, F.W.; Dong, S.Y.; Liu, W.; Wang, H.; Chen, Z.Q.; Wang, J.F. CONSTANS-like 9 (COL9) delays the flowering time in Oryza sativa by repressing the Ehd1 pathway. Biochem. Biophys. Res. Commun. 2016, 479, 173–178. [Google Scholar] [CrossRef] [Green Version]
- Cramer, G.R. Abiotic stress & plant responses from the whole vine to the genes. Aust. J. Grape Wine Res. 2010, 16, 86–93. [Google Scholar]
- Skirycz, A.; Inze, D. More from less: Plant growth under limited water. Curr. Opin. Biotechnol. 2010, 21, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Seki, M.; Ishida, J.; Satou, M.; Sakurai, T.; Narusaka, M.; Kamiya, A.; Nakajima, M.; Enju, A.; Akiyama, K.; et al. Monitoring the expression profiles of genes induced by hyperosmotic, high salinity, and oxidative stress and abscisic acid treatment in Arabidopsis cell culture using a full-length cDNA microarray. Plant Mol. Biol. 2004, 56, 29–55. [Google Scholar] [CrossRef] [PubMed]
- Cramer, G.R.; Urano, K.; Delrot, S.; Pezzotti, M.; Shinozaki, K. Effects of abiotic stress on plants: A systems biology perspective. BMC Plant Biol. 2011, 11, 163. [Google Scholar] [CrossRef] [Green Version]
- Min, J.H.; Chung, J.S.; Lee, K.H.; Kim, C.S. The CONSTANS-like 4 transcription factor, AtCOL4, positively regulates abiotic stress tolerance through an abscisic acid-dependent manner in Arabidopsis. J. Integr. Plant Biol. 2015, 57, 313–324. [Google Scholar] [CrossRef]
- Qin, W.Q.; Yu, Y.; Jin, Y.Y.; Wang, X.D.; Liu, J.; Xi, J.P.; Li, Z.; Li, H.Q.; Zhao, G.; Hu, W.; et al. Genome-wide analysis elucidates the role of CONSTANS-like genes in stress responses of cotton. Int. J. Mol. Sci. 2018, 19, 2658. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.D.; Ding, Q.Y.; Liu, J.; Yang, C.L.; Chen, Y.; Zhang, S.F.; Zhu, J.C.; Wang, D.J. Brassica napus COL transcription factor BnCOL2 negatively affects the tolerance of transgenic Arabidopsis to drought stress Environ. Exp. Bot. 2020, 178, 104171. [Google Scholar] [CrossRef]
- Xiong, L.; Schumaker, K.S.; Zhu, J.K. Cell signaling during cold, drought, and salt stress. Plant Cell 2002, 14, S165–S183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aubert, Y.; Vile, D.; Pervent, M.; Aldon, D.; Ranty, B.; Simonneau, T.; Vavasseur, A.; Galau, J.P. RD20, a stress-inducible caleosin, participates in stomatal control, transpiration and drought tolerance in Arabidopsis thaliana. Plant Cell Physiol. 2010, 51, 1975–1987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asif, M.A.; Zafar, Y.; Iqbal, J.; Iqbal, M.M.; Rashid, U.; Ali, G.M.; Arif, A.; Nazir, F. Enhanced expression of AtNHX1, in transgenic groundnut (Arachis hypogaea L.) improves salt and drought tolerence. Mol. Biotechnol. 2011, 49, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Wright, J. RD20/CLO3, a Stress-Induced Calcium-Binding Protein, Acts as a Negative Regulator of GPA1 in Arabidopsis through GAP Activity. Ph.D. Thesis, Concordia University, Montreal, QC, Canada, 2014. [Google Scholar]
- Shi, H.; Lee, B.H.; Wu, S.J.; Zhu, J.K. Overexpression of a plasma membrane Na+/H+ antiporter gene improves salt tolerance in Arabidopsis thaliana. Nat. Biotechnol. 2003, 21, 81–85. [Google Scholar] [CrossRef]
- Meng, L.S.; Wang, Z.B.; Yao, S.Q.; Liu, A. The ARF2-ANT-COR15A gene cascade regulates ABA-signaling-mediated resistance of large seeds to drought in Arabidopsis. J. Cell Sci. 2015, 128, 3922–3932. [Google Scholar] [PubMed] [Green Version]
- Yu, H.X.; Luo, C.; Xu, C.; He, X.H. A simple and efcient method for high quality DNA extraction from transgenic Arabidopsis and Tobacco. Mol. Plant Breed. 2016, 14, 1436–1440. [Google Scholar]
- Luo, C.; He, X.H.; Chen, H.; Hu, Y.; Ou, S.J. Molecular cloning and expression analysis of four action genes (MiACT) from mango. Biol. Plantarum. 2013, 57, 238–244. [Google Scholar] [CrossRef]
- Libak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2–△△Ct method. Methods 2001, 25, 402–408. [Google Scholar]
- Clough, S.J.; Bent, A.F. Floral dip: A simplifed method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Zhang, X.F.; Duan, H.; Lian, C.L.; Liu, C.; Yin, W.L.; Xia, X.L. Three stress-responsive NAC transcription factors from Populus euphratica differentially regulate salt and drought tolerance in transgenic plants. Physiol. Plantarum. 2017, 162, 73–97. [Google Scholar] [CrossRef] [Green Version]
- Pla, M.; Vilardell, J.; Guiltinan, M.J.; Marcotte, W.R.; Niogret, M.F.; Quatrano, R.S.; Pages, M. The cis-regulatory element CCACGTGG is involved in ABA and water-stress responses of the maize gene rab28. Plant Mol. Biol. 1993, 21, 259–266. [Google Scholar] [CrossRef]
- Li, S.X.; Liu, J.L.; An, Y.R.; Cao, Y.M.; Liu, Y.S.; Zhang, J.; Geng, J.C.; Hu, T.M.; Yang, P.Z. MsPIP2;2, a novel aquaporin gene from Medicago sativa, confers salt tolerance in transgenic Arabidopsis. Environ. Exp. Bot. 2019, 165, 39–52. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Days to Flowering | No. Rosette Leaves | Plant Height a (cm) | |||
|---|---|---|---|---|---|---|
| LD | SD | LD | SD | LD | SD | |
| WT | 24.8 ± 0.6 | 50.1 ± 0.8 | 5.5 ± 0.5 | 7.2 ± 0.3 | 6.4 ± 0.8 | 11.4 ± 0.6 |
| pBI121 | 25.6 ± 0.4 | 49.6 ± 0.7 | 7.6 ±0.6 | 7.0 ± 0.5 | 6.7 ± 0.4 | 11.7 ± 0.4 |
| MiCOL16A | ||||||
| OE2 | 28.0 ± 0.3 * | 51.3 ± 0.8 | 6.3 ± 0.4 | 7.6 ± 0.3 | 4.6 ± 1.1 * | 14.2 ± 0.9 * |
| OE8 | 28.0 ± 0.5 * | 52.8 ± 0.2 * | 6.8 ± 0.6 * | 7.3 ± 0.7 | 4.5 ± 0.6 * | 14.2 ± 0.8 * |
| OE10 | 28.1 ± 0.8 * | 52.8 ± 0.3 * | 7.5 ± 1.0 * | 7.3 ± 0.5 | 6.2 ± 1.1 | 13.0 ± 0.3 * |
| MiCOL16B | ||||||
| OE3 | 27.6 ± 0.5 * | 53.0 ± 0.4 * | 6.6 ± 0.7 * | 7.4 ± 0.6 | 4.7 ± 1.0 * | 17.7 ± 1.1 * |
| OE12 | 26.9 ± 0.8 * | 51.2 ± 0.9 | 7.1 ± 0.5 * | 7.3 ± 0.4 | 5.2 ± 1.1 * | 17.1 ± 1.0 * |
| OE13 | 28.4 ± 0.5 * | 51.5 ± 0.4 * | 6.7 ± 0.7 * | 6.6 ± 0.6 | 4.3 ± 0.8 * | 13.4 ± 0.2 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Luo, C.; Guo, Y.; Liang, R.; Yu, H.; Chen, S.; Mo, X.; Yang, X.; He, X. Isolation and Functional Characterization of Two CONSTANS-like 16 (MiCOL16) Genes from Mango. Int. J. Mol. Sci. 2022, 23, 3075. https://doi.org/10.3390/ijms23063075
Liu Y, Luo C, Guo Y, Liang R, Yu H, Chen S, Mo X, Yang X, He X. Isolation and Functional Characterization of Two CONSTANS-like 16 (MiCOL16) Genes from Mango. International Journal of Molecular Sciences. 2022; 23(6):3075. https://doi.org/10.3390/ijms23063075
Chicago/Turabian StyleLiu, Yuan, Cong Luo, Yihang Guo, Rongzhen Liang, Haixia Yu, Shuquan Chen, Xiao Mo, Xiaozhou Yang, and Xinhua He. 2022. "Isolation and Functional Characterization of Two CONSTANS-like 16 (MiCOL16) Genes from Mango" International Journal of Molecular Sciences 23, no. 6: 3075. https://doi.org/10.3390/ijms23063075
APA StyleLiu, Y., Luo, C., Guo, Y., Liang, R., Yu, H., Chen, S., Mo, X., Yang, X., & He, X. (2022). Isolation and Functional Characterization of Two CONSTANS-like 16 (MiCOL16) Genes from Mango. International Journal of Molecular Sciences, 23(6), 3075. https://doi.org/10.3390/ijms23063075