
Platelets Purification Is a Crucial Step for Transcriptomic Analysis

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
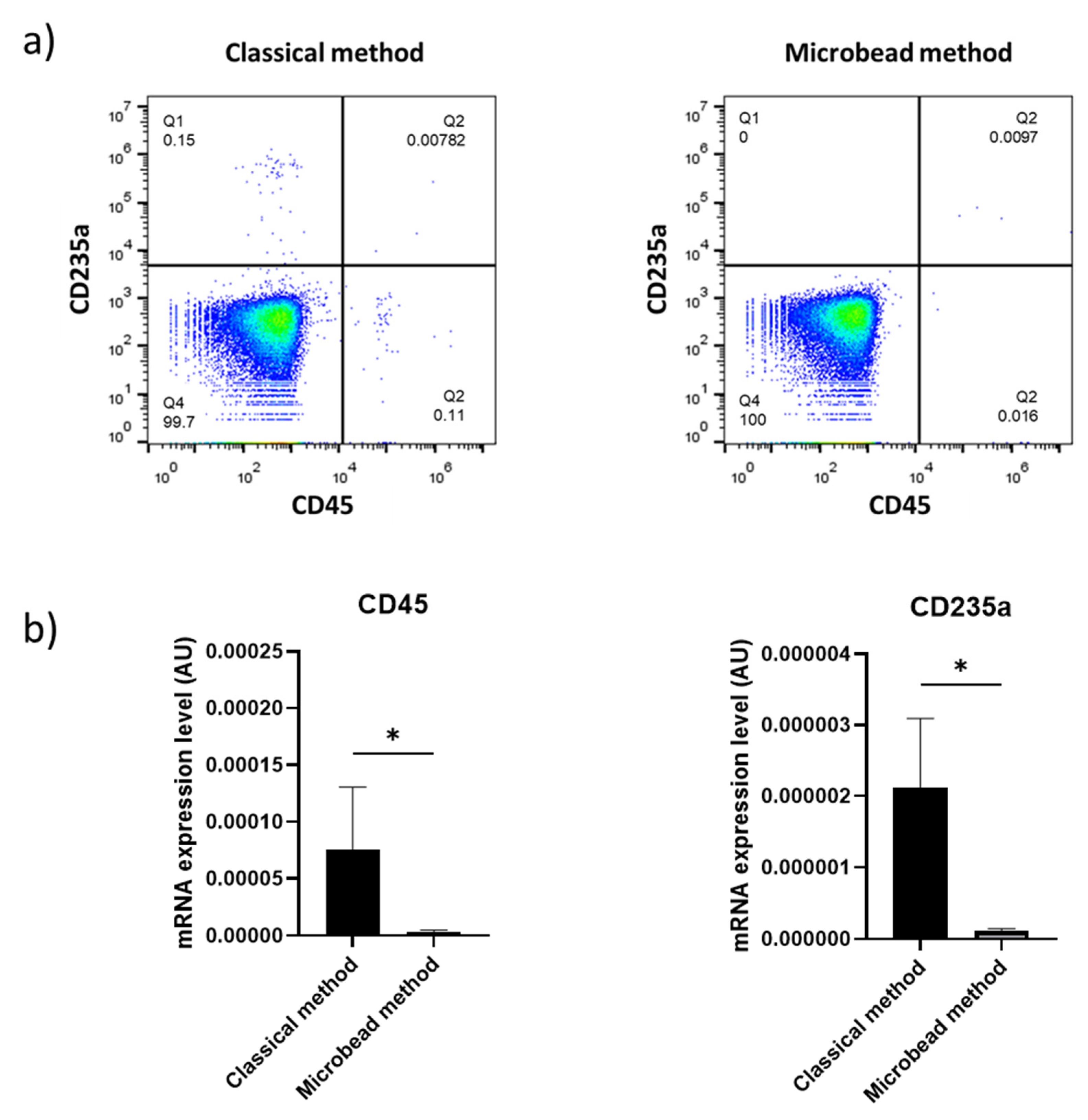
2.1. Magnetic Microbeads Greatly Reduce Leukocyte and Erythrocyte Contamination
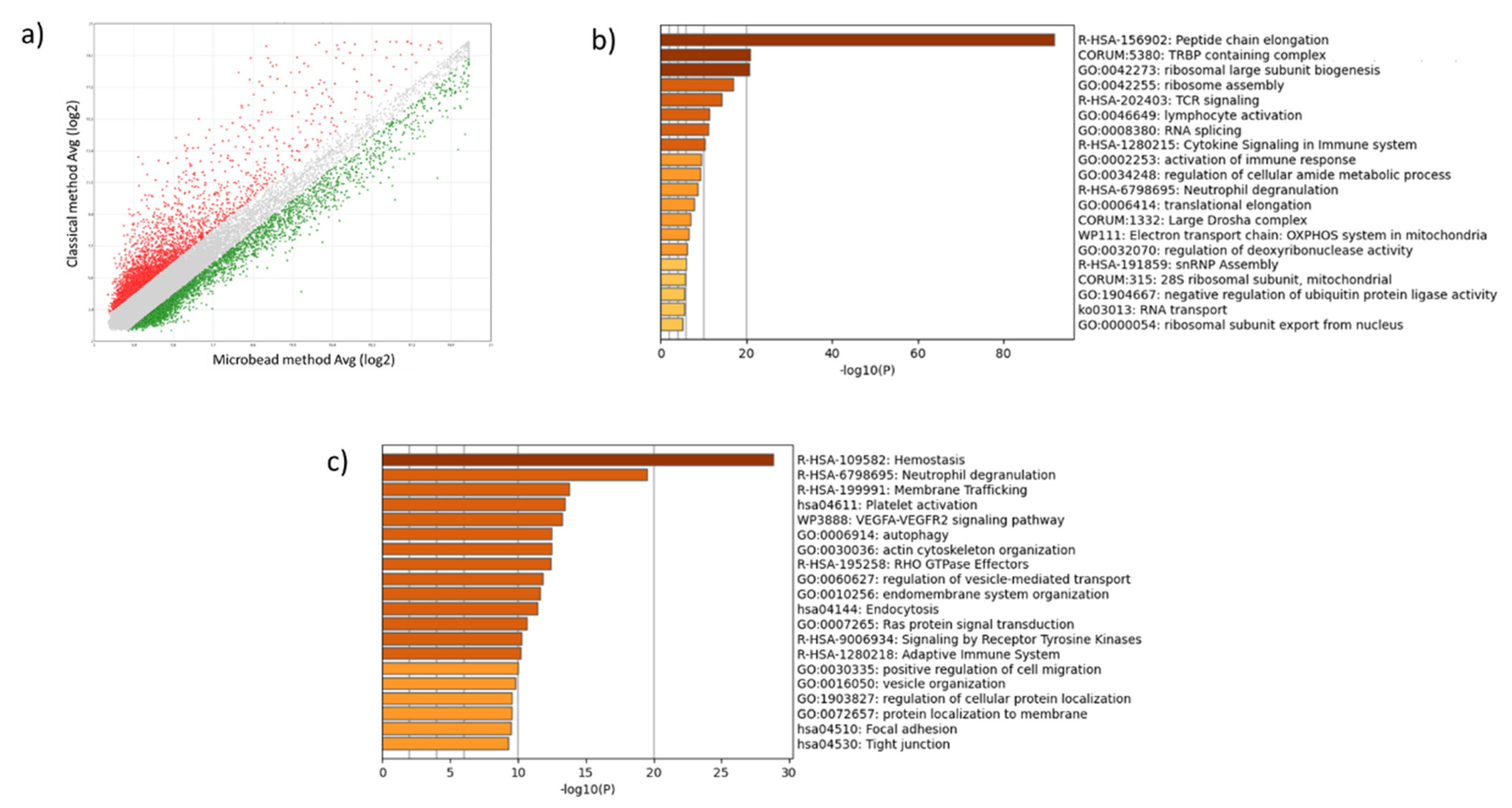
2.2. Leukocyte and Erythrocyte Contamination Strongly Alters Platelet Transcriptome Profile
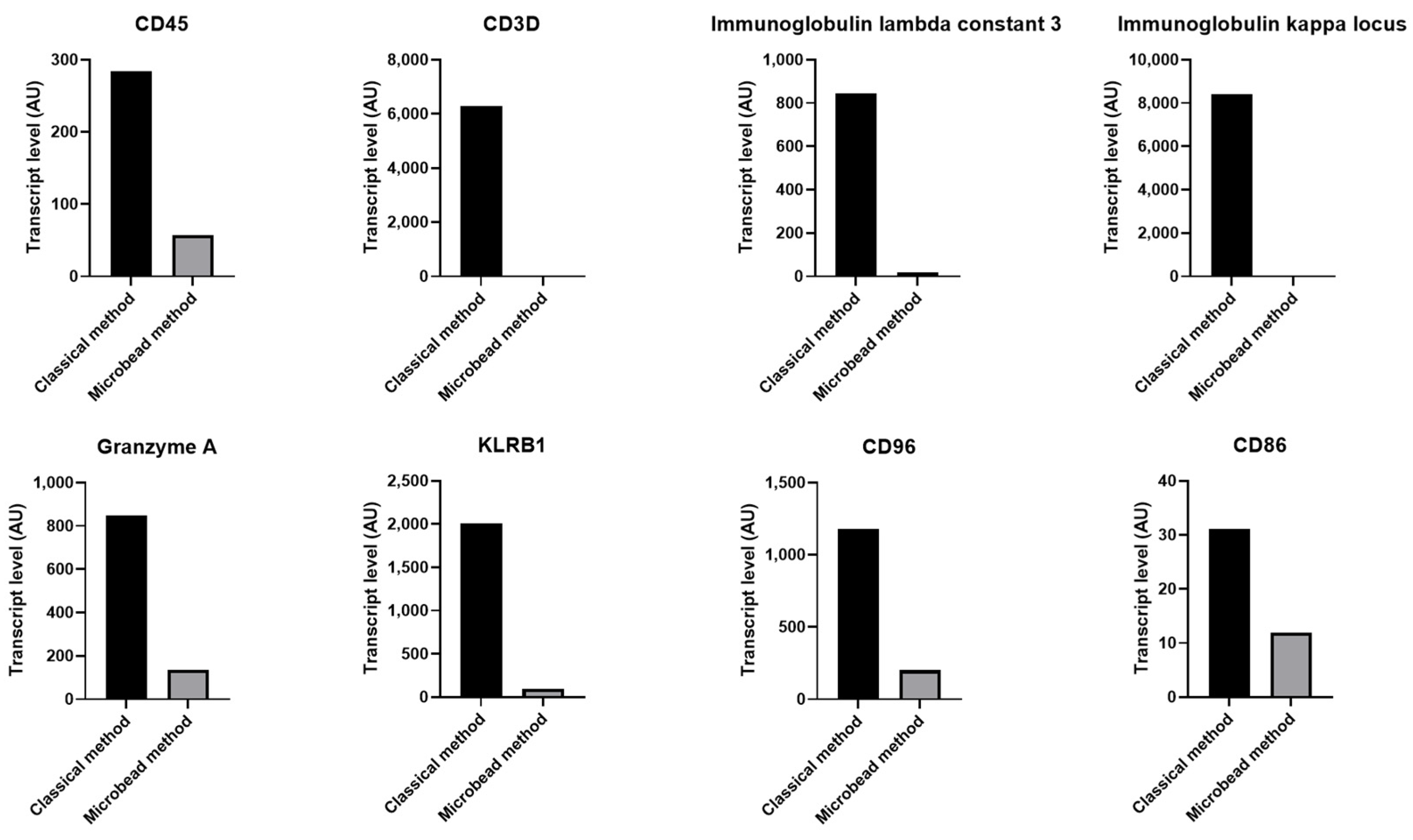
2.3. Increased Level of Specific Leukocyte and Erythrocyte Transcriptss in Contaminated Platelet Preparation
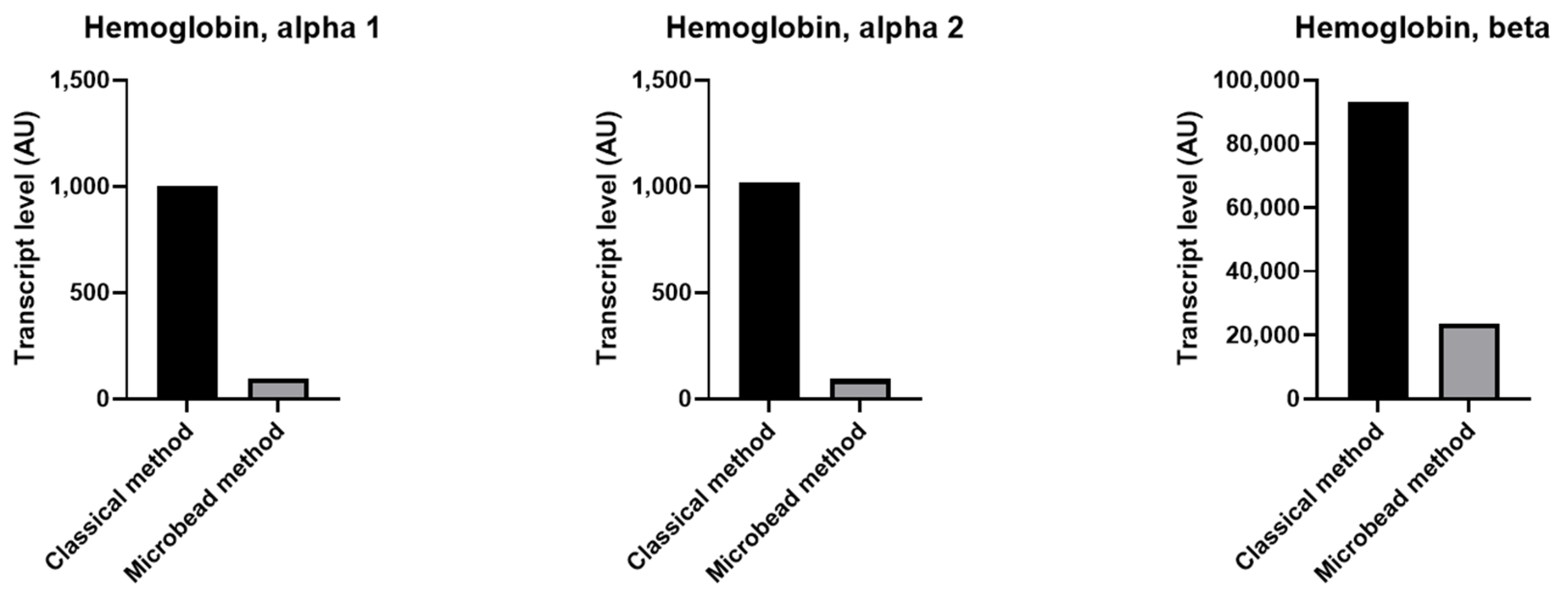
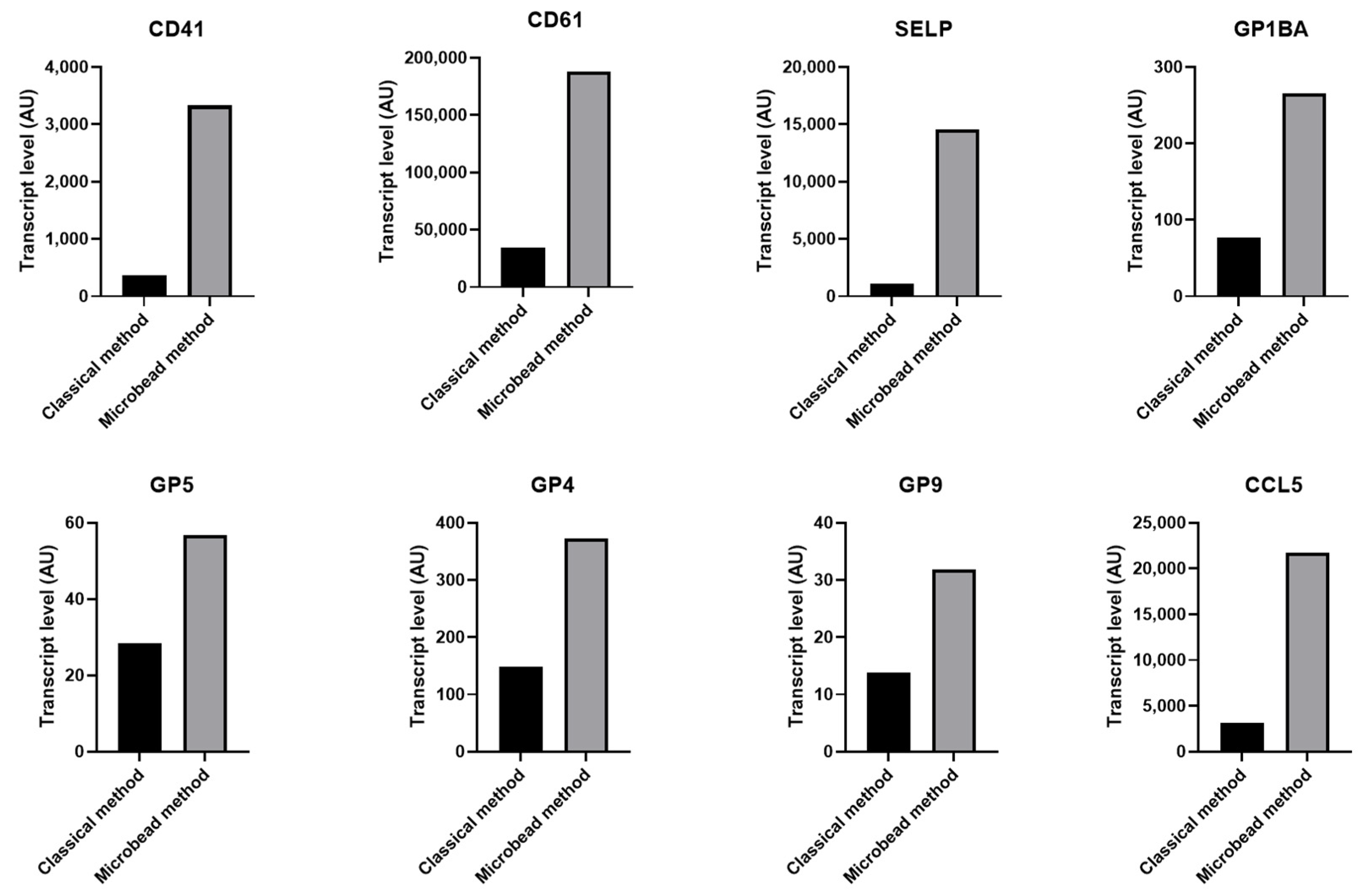
2.4. Decreased Level of Transcripts Related to Platelets in Contaminated Platelet Preparation
3. Discussion
4. Materials and Methods
4.1. Study Subjects
4.2. Platelet Isolation
4.3. RNA Purification
4.4. Real-Time Quantitative PCR Analysis
4.5. Flow Cytometry
4.6. Microarray Analysis
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chebbo, M.; Duez, C.; Alessi, M.C.; Chanez, P.; Gras, D. Platelets: A Potential Role in Chronic Respiratory Diseases? Eur. Respir. Rev. 2021, 30, 210062. [Google Scholar] [CrossRef] [PubMed]
- Rowley, J.W.; Schwertz, H.; Weyrich, A.S. Platelet MRNA: The Meaning behind the Message. Curr. Opin. Hematol. 2012, 19, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, G.A.; Weyrich, A.S. Signal-Dependent Protein Synthesis by Activated Platelets: New Pathways to Altered Phenotype and Function. ATVB 2008, 28, s17–s24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laffont, B.; Corduan, A.; Plé, H.; Duchez, A.-C.; Cloutier, N.; Boilard, E.; Provost, P. Activated Platelets Can Deliver MRNA Regulatory Ago2•microRNA Complexes to Endothelial Cells via Microparticles. Blood 2013, 122, 253–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahr, W.H.; Hinckley, J.; Li, L.; Schwertz, H.; Christensen, H.; Rowley, J.W.; Pluthero, F.G.; Urban, D.; Fabbro, S.; Nixon, B.; et al. Mutations in NBEAL2, Encoding a BEACH Protein, Cause Gray Platelet Syndrome. Nat. Genet. 2011, 43, 738–740. [Google Scholar] [CrossRef] [PubMed]
- Davizon-Castillo, P.; Rowley, J.W.; Rondina, M.T. Megakaryocyte and Platelet Transcriptomics for Discoveries in Human Health and Disease. ATVB 2020, 40, 1432–1440. [Google Scholar] [CrossRef]
- Manne, B.K.; Denorme, F.; Middleton, E.A.; Portier, I.; Rowley, J.W.; Stubben, C.; Petrey, A.C.; Tolley, N.D.; Guo, L.; Cody, M.; et al. Platelet Gene Expression and Function in Patients with COVID-19. Blood 2020, 136, 1317–1329. [Google Scholar] [CrossRef]
- Sol, N.; Wurdinger, T. Platelet RNA Signatures for the Detection of Cancer. Cancer Metastasis Rev. 2017, 36, 263–272. [Google Scholar] [CrossRef]
- Wrzyszcz, A.; Urbaniak, J.; Sapa, A.; Woźniak, M. An Efficient Method for Isolation of Representative and Contamination-Free Population of Blood Platelets for Proteomic Studies. Platelets 2017, 28, 43–53. [Google Scholar] [CrossRef]
- Teruel-Montoya, R.; Kong, X.; Abraham, S.; Ma, L.; Kunapuli, S.P.; Holinstat, M.; Shaw, C.A.; McKenzie, S.E.; Edelstein, L.C.; Bray, P.F. MicroRNA Expression Differences in Human Hematopoietic Cell Lineages Enable Regulated Transgene Expression. PLoS ONE 2014, 9, e102259. [Google Scholar] [CrossRef]
- Macaulay, I.C.; Carr, P.; Farrugia, R.; Watkins, N.A. Analysing the Platelet Transcriptome. Vox Sang. 2004, 87, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Osman, A.; Hitzler, W.E.; Ameur, A.; Provost, P. Differential Expression Analysis by RNA-Seq Reveals Perturbations in the Platelet MRNA Transcriptome Triggered by Pathogen Reduction Systems. PLoS ONE 2015, 10, e0133070. [Google Scholar] [CrossRef]
- Palmer, C.; Diehn, M.; Alizadeh, A.A.; Brown, P.O. Cell-Type Specific Gene Expression Profiles of Leukocytes in Human Peripheral Blood. BMC Genom. 2006, 7, 115. [Google Scholar] [CrossRef] [PubMed]
- Rowley, J.W.; Oler, A.J.; Tolley, N.D.; Hunter, B.N.; Low, E.N.; Nix, D.A.; Yost, C.C.; Zimmerman, G.A.; Weyrich, A.S. Genome-Wide RNA-Seq Analysis of Human and Mouse Platelet Transcriptomes. Blood 2011, 118, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lood, C.; Amisten, S.; Gullstrand, B.; Jönsen, A.; Allhorn, M.; Truedsson, L.; Sturfelt, G.; Erlinge, D.; Bengtsson, A.A. Platelet Transcriptional Profile and Protein Expression in Patients with Systemic Lupus Erythematosus: Up-Regulation of the Type I Interferon System Is Strongly Associated with Vascular Disease. Blood 2010, 116, 1951–1957. [Google Scholar] [CrossRef] [Green Version]
- Middleton, E.A.; Rowley, J.W.; Campbell, R.A.; Grissom, C.K.; Brown, S.M.; Beesley, S.J.; Schwertz, H.; Kosaka, Y.; Manne, B.K.; Krauel, K.; et al. Sepsis Alters the Transcriptional and Translational Landscape of Human and Murine Platelets. Blood 2019, 134, 911–923. [Google Scholar] [CrossRef] [PubMed]
- Bray, P.F.; McKenzie, S.E.; Edelstein, L.C.; Nagalla, S.; Delgrosso, K.; Ertel, A.; Kupper, J.; Jing, Y.; Londin, E.; Loher, P.; et al. The Complex Transcriptional Landscape of the Anucleate Human Platelet. BMC Genom. 2013, 14, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Londin, E.R.; Hatzimichael, E.; Loher, P.; Edelstein, L.; Shaw, C.; Delgrosso, K.; Fortina, P.; Bray, P.F.; McKenzie, S.E.; Rigoutsos, I. The Human Platelet: Strong Transcriptome Correlations among Individuals Associate Weakly with the Platelet Proteome. Biol. Direct 2014, 9, 3. [Google Scholar] [CrossRef] [Green Version]
- Eicher, J.D.; Wakabayashi, Y.; Vitseva, O.; Esa, N.; Yang, Y.; Zhu, J.; Freedman, J.E.; McManus, D.D.; Johnson, A.D. Characterization of the Platelet Transcriptome by RNA Sequencing in Patients with Acute Myocardial Infarction. Platelets 2016, 27, 230–239. [Google Scholar] [CrossRef] [Green Version]
- Raghavachari, N.; Xu, X.; Harris, A.; Villagra, J.; Logun, C.; Barb, J.; Solomon, M.A.; Suffredini, A.F.; Danner, R.L.; Kato, G.; et al. Amplified Expression Profiling of Platelet Transcriptome Reveals Changes in Arginine Metabolic Pathways in Patients With Sickle Cell Disease. Circulation 2007, 115, 1551–1562. [Google Scholar] [CrossRef] [Green Version]
- Healy, A.M.; Pickard, M.D.; Pradhan, A.D.; Wang, Y.; Chen, Z.; Croce, K.; Sakuma, M.; Shi, C.; Zago, A.C.; Garasic, J.; et al. Platelet Expression Profiling and Clinical Validation of Myeloid-Related Protein-14 as a Novel Determinant of Cardiovascular Events. Circulation 2006, 113, 2278–2284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Supernat, A.; Popęda, M.; Pastuszak, K.; Best, M.G.; Grešner, P.; Veld, S.I.; Siek, B.; Bednarz-Knoll, N.; Rondina, M.T.; Stokowy, T.; et al. Transcriptomic Landscape of Blood Platelets in Healthy Donors. Sci. Rep. 2021, 11, 15679. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Liu, Y.; Wang, X.; Yang, L.; Li, H.; Wang, Y.; Liu, M.; Zhao, X.; Xie, Y.; Yang, Y.; et al. SARS-CoV-2 Binds Platelet ACE2 to Enhance Thrombosis in COVID-19. J. Hematol. Oncol. 2020, 13, 120. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Yuan, L.; Cheng, B.; Dai, K. Identification of Seven Tumor-educated Platelets RNAs for Cancer Diagnosis. J. Clin. Lab. Anal. 2021, 35, e23791. [Google Scholar] [CrossRef]
- Zhang, G.; Chen, H.; Guo, Y.; Zhang, W.; Jiang, Q.; Zhang, S.; Han, L.; Chen, S.; Xue, R. Activation of Platelet NLRP3 Inflammasome in Crohn’s Disease. Front. Pharmacol. 2021, 12, 705325. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, A.J. MicroRNA in Erythrocytes. Biochem. Soc. Trans. 2010, 38, 229–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanscombe, O.; Vidal, M.; Kaeda, J.; Luzzatto, L.; Greaves, D.R.; Grosveld, F. High-Level, Erythroid-Specific Expression of the Human a-Globin Gene in Transgenic Mice and the Production of Human Hemoglobin in Murine Erythrocytes. Genes Dev. 1989, 3, 1572–1581. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Cheng, L.; Zhang, Y.; Wu, R.; Wang, M.; Gu, Y.; Zhao, W.; Li, P.; Li, B.; Zhang, Y.; et al. Extensive Up-Regulation of Gene Expression in Cancer: The Normalised Use of Microarray Data. Mol. BioSyst. 2012, 8, 818. [Google Scholar] [CrossRef]
- Cheng, L.; Lo, L.-Y.; Tang, N.L.S.; Wang, D.; Leung, K.-S. CrossNorm: A Novel Normalization Strategy for Microarray Data in Cancers. Sci. Rep. 2016, 6, 18898. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape Provides a Biologist-Oriented Resource for the Analysis of Systems-Level Datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Direction | Sequences (5′ to 3′) |
|---|---|---|
| CD45 | Forward | ACCAGGAATGGATGTCGCTA |
| Reverse | TGGGGCCTGTAAAAGTGTCC | |
| CD235a | Forward | CAAACGGGACACATATGCAG |
| Reverse | GTCGGCGAATACCGTAAGAA | |
| 18S | Forward | TCAAGAACGAAAGTCGGAGG |
| Reverse | CAGCTTTGCAACCATACTCC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chebbo, M.; Assou, S.; Pantesco, V.; Duez, C.; Alessi, M.C.; Chanez, P.; Gras, D. Platelets Purification Is a Crucial Step for Transcriptomic Analysis. Int. J. Mol. Sci. 2022, 23, 3100. https://doi.org/10.3390/ijms23063100
Chebbo M, Assou S, Pantesco V, Duez C, Alessi MC, Chanez P, Gras D. Platelets Purification Is a Crucial Step for Transcriptomic Analysis. International Journal of Molecular Sciences. 2022; 23(6):3100. https://doi.org/10.3390/ijms23063100
Chicago/Turabian StyleChebbo, Mohamad, Said Assou, Veronique Pantesco, Catherine Duez, Marie C. Alessi, Pascal Chanez, and Delphine Gras. 2022. "Platelets Purification Is a Crucial Step for Transcriptomic Analysis" International Journal of Molecular Sciences 23, no. 6: 3100. https://doi.org/10.3390/ijms23063100
APA StyleChebbo, M., Assou, S., Pantesco, V., Duez, C., Alessi, M. C., Chanez, P., & Gras, D. (2022). Platelets Purification Is a Crucial Step for Transcriptomic Analysis. International Journal of Molecular Sciences, 23(6), 3100. https://doi.org/10.3390/ijms23063100