
Sex Hormones and Their Effects on Ocular Disorders and Pathophysiology: Current Aspects and Our Experience



Abstract
:1. Introduction
2. Sex Hormones
2.1. Steroidogenesis
2.2. Mechanism of Action
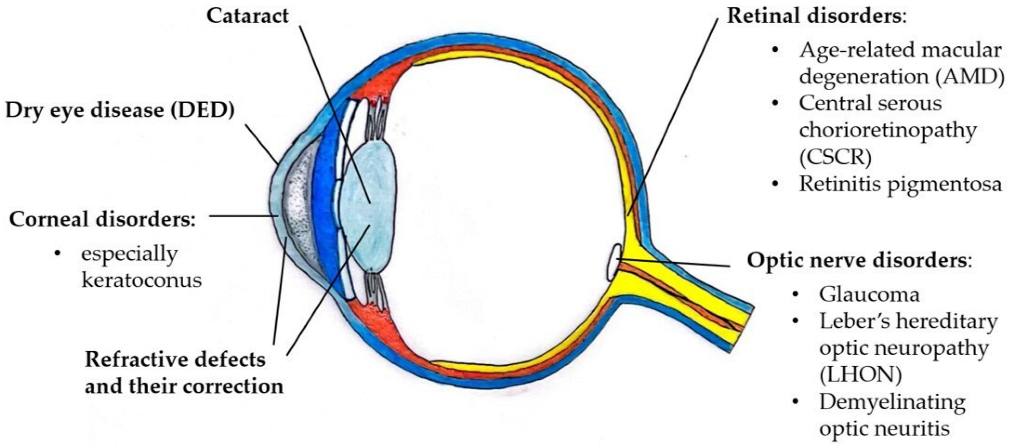
3. Effects of Sex Hormones on Ocular Disorders
3.1. Dry Eye Disease
3.2. Corneal Disorders
3.3. Cataract
3.4. Glaucoma and Optic Nerve Disorders
3.4.1. Glaucoma
3.4.2. Leber’s Hereditary Optic Neuropathy (LHON)
3.4.3. Optic Neuritis
3.5. Retinal Disorders
3.5.1. Age-Related Macular Degeneration (AMD)
3.5.2. Central Serous Chorioretinopathy (CSCR)
3.5.3. Retinitis Pigmentosa
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Azcoitia, I.; Arevalo, M.A.; de Nicola, A.F.; Garcia-Segura, L.M. Neuroprotective actions of estradiol revisited. Trends Endocrinol. Metab. TEM 2011, 22, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Clarke, B.L.; Khosla, S. Female reproductive system and bone. Arch. Biochem. Biophys. 2010, 503, 118–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosner, W.; Hankinson, S.E.; Sluss, P.M.; Vesper, H.W.; Wierman, M.E. Challenges to the measurement of estradiol: An endocrine society position statement. J. Clin. Endocrinol. Metab. 2013, 98, 1376–1387. [Google Scholar] [CrossRef] [Green Version]
- Melcangi, R.C.; Panzica, G.; Garcia-Segura, L.M. Neuroactive steroids: Focus on human brain. Neuroscience 2011, 191, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, K.-G.; Bergert, H.; Funk, R. Neurodegenerative diseases of the retina and potential for protection and recovery. Curr. Neuropharmacol. 2008, 6, 164–178. [Google Scholar] [CrossRef] [Green Version]
- Toker, E.; Yenice, Ö.; Akpinar, I.; Aribal, E.; Kazokoglu, H. The influence of sex hormones on ocular blood flow in women. Acta Ophthalmol. Scand. 2003, 81, 617–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faria AF, M.; de Souza, M.A.M.; Geber, S. Vascular resistance of central retinal artery is reduced in postmenopausal women after use of estrogen. Menopause 2011, 18, 869–872. [Google Scholar] [CrossRef] [PubMed]
- Toker, E.; Yenice, Ö.; Temel, A. Influence of serum levels of sex hormones on intraocular pressure in menopausal women. J. Glaucoma 2003, 12, 436–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tehrani, S. Gender difference in the pathophysiology and treatment of glaucoma. Curr. Eye Res. 2015, 40, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.D.; Johar, K.; Nagpal, K.; Vasavada, A.R. Sex hormone receptors in the human eye. Surv. Ophthalmol. 2005, 50, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Nuzzi, R.; Scalabrin, S.; Becco, A.; Panzica, G. Gonadal Hormones and Retinal Disorders: A Review. Front. Endocrinol. 2018, 9, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuzzi, R.; Scalabrin, S.; Becco, A.; Panzica, G. Sex Hormones and Optic Nerve Disorders: A Review. Front. Neurosci. 2019, 13, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, W.L.; Auchus, R.J. The molecular biology, biochemistry, and physiology of human steroidogenesis and its disorders. Endocr. Rev. 2011, 32, 81–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Black, S.M.; Harikrishna, J.A.; Szklarz, G.D.; Miller, W.L. The mitochondrial environment is required for activity of the cholesterol side-chain cleavage enzyme, cytochrome P450scc. Proc. Natl. Acad. Sci. USA 1994, 91, 7247–7251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauschenberger, K.; Schöler, K.; Sass, J.O.; Sauer, S.; Djuric, Z.; Rumig, C.; Wolf, N.I.; Okun, J.G.; Kölker, S.; Schwarz, H.; et al. A non-enzymatic function of 17beta-hydroxysteroid dehydrogenase type 10 is required for mitochondrial integrity and cell survival. EMBO Mol. Med. 2010, 2, 51–62. [Google Scholar] [CrossRef]
- Luu-The, V.; Labrie, F. The intracrine sex steroid biosynthesis pathways. Prog. Brain Res. 2010, 181, 177–192. [Google Scholar] [PubMed]
- Lösel, R.; Wehling, M. Nongenomic actions of steroid hormones. Nat. Rev. Mol. Cell Biol. 2003, 4, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Tuohimaa, P.; Bläuer, M.; Pasanen, S.; Passinen, S.; Pekki, A.; Punnonen, R.; Syvälä, H.; Valkila, J.; Wallén, M.; Väliaho, J.; et al. Mechanisms of action of sex steroid hormones: Basic concepts and clinical correlations. Maturitas 1996, 23, S3–S12. [Google Scholar] [CrossRef]
- Singh, S.; Gupta, P.D. Induction of phosphoinositide-mediated signal transduction pathway by 17 beta-oestradiol in rat vaginal epithelial cells. J. Mol. Endocrinol. 1997, 19, 249–257. [Google Scholar] [CrossRef]
- Singh, S.; Shaul, P.W.; Gupta, P.D. Conventional estrogen receptors are found in the plasma membrane of vaginal epithelial cells of the rat. Steroids 2002, 67, 757–764. [Google Scholar] [CrossRef]
- Enmark, E.; Pelto-Huikko, M.; Grandien, K.; Lagercrantz, S.; Lagercrantz, J.; Fried, G.; Nordenskjöld, M.; Gustafsson, J.Å. Human estrogen receptor beta-gene structure, chromosomal localization, and expression pattern. J. Clin. Endocrinol. Metab. 1997, 82, 4258–4265. [Google Scholar] [PubMed] [Green Version]
- Lubahn, D.B.; Joseph, D.R.; Sullivan, P.M.; Willard, H.F.; French, F.S.; Wilson, E.M. Cloning of human androgen receptor complementary DNA and localization to the X chromosome. Science 1988, 240, 327–330. [Google Scholar] [CrossRef] [PubMed]
- Labrie, F. Adrenal androgens and intracrinology. Semin. Reprod. Med. 2004, 22, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Koos, R.D. Minireview: Putting physiology back into estrogens’ mechanism of action. Endocrinology 2011, 152, 4481–4488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Versura, P.; Giannaccare, G.; Campos, E.C. Sex-steroid imbalance in females and dry eye. Curr. Eye Res. 2015, 40, 162–175. [Google Scholar] [CrossRef] [PubMed]
- Stapleton, F.; Alves, M.; Bunya, V.Y.; Jalbert, I.; Lekhanont, K.; Malet, F.; Na, K.S.; Schaumberg, D.; Uchino, M.; Vehof, J.; et al. TFOS DEWS II Epidemiology Report. Ocul. Surf. 2017, 15, 334–365. [Google Scholar] [CrossRef]
- Lemp, M.A.; Baudouin, C.; Baum, J.; Dogru, M.; Foulks, G.N.; Kinoshita, S.; Laibson, P.; McCulley, J.; Murube, J.; Pflugfelder, S.C.; et al. The definition and classification of dry eye disease: Report of the Definition and Classification Subcommittee of the International Dry Eye WorkShop. Ocul. Surf. 2007, 5, 75–92. [Google Scholar]
- Schaumberg, D.A.; Sullivan, D.A.; Buring, J.E.; Dana, M.R. Prevalence of dry eye syndrome among US women. Am. J. Ophthalmol. 2003, 136, 318–326. [Google Scholar] [CrossRef]
- Mathers, W.D.; Stovall, D.; Lane, J.A.; Zimmerman, M.B.; Johnson, S. Menopause and tear function: The influence of prolactin and sex hormones on human tear production. Cornea 1998, 17, 353–358. [Google Scholar] [CrossRef]
- Sullivan, D.A.; Rocha, E.M.; Aragona, P.; Clayton, J.A.; Ding, J.; Golebiowski, B.; Hampel, U.; McDermott, A.M.; Schaumberg, D.A.; Srinivasan, S.; et al. TFOS DEWS II Sex, Gender, and Hormones Report. Ocul. Surf. 2017, 15, 284–333. [Google Scholar] [CrossRef]
- Rapoport, Y.; Singer, J.M.; Ling, J.D.; Gregory, A.; Kohanim, S. A Comprehensive Review of Sex Disparities in Symptoms, Pathophysiology, and Epidemiology of Dry Eye Syndrome. Semin. Ophthalmol. 2016, 31, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, D.A.; Yamagami, H.; Liu, M.; Steagall, R.J.; Schirra, F.; Suzuki, T.; Krenzer, K.L.; Cermak, J.M.; Sullivan, R.M.; Richards, S.M.; et al. Sex steroids, the meibomian gland and evaporative dry eye. Adv. Exp. Med. Biol. 2002, 506, 389–399. [Google Scholar] [PubMed]
- Esmaeli, B.; Harvey, J.T.; Hewlett, B. Immunohistochemical evidence for estrogen receptors in meibomian glands. Ophthalmology 2000, 107, 180–184. [Google Scholar] [CrossRef]
- Golebiowski, B.; Badarudin, N.; Eden, J.; You, J.; Hampel, U.; Stapleton, F. Does endogenous serum oestrogen play a role in meibomian gland dysfunction in postmenopausal women with dry eye? Br. J. Ophthalmol. 2017, 101, 218–222. [Google Scholar] [CrossRef]
- Suzuki, T.; Schirra, F.; Richards, S.M.; Jensen R, v.; Sullivan, D.A. Estrogen and progesterone control of gene expression in the mouse meibomian gland. Investig. Ophthalmol. Vis. Sci. 2008, 49, 1797–1808. [Google Scholar] [CrossRef]
- Butler, L.; Santoro, N. The reproductive endocrinology of the menopausal transition. Steroids 2011, 76, 627–635. [Google Scholar] [CrossRef] [Green Version]
- Metka, M.; Enzelsberger, H.; Knogler, W.; Schurz, B.; Aichmair, H. Ophthalmic complaints as a climacteric symptom. Maturitas 1991, 14, 3–8. [Google Scholar] [CrossRef]
- Nichols, K.K.; Foulks, G.N.; Bron, A.J.; Glasgow, B.J.; Dogru, M.; Tsubota, K.; Lemp, M.A.; Sullivan, D.A. The international workshop on meibomian gland dysfunction: Executive summary. Investig. Ophthalmol. Vis. Sci. 2011, 52, 1922–1929. [Google Scholar] [CrossRef] [Green Version]
- Erdem, U.; Ozdegirmenci, O.; Sobaci, E.; Sobaci, G.; Göktolga, U.; Dagli, S. Dry eye in post-menopausal women using hormone replacement therapy. Maturitas 2007, 56, 257–262. [Google Scholar] [CrossRef]
- Ablamowicz, A.F.; Nichols, J.J.; Nichols, K.K. Association between Serum Levels of Testosterone and Estradiol with Meibomian Gland Assessments in Postmenopausal Women. Investig. Ophthalmol. Vis. Sci. 2016, 57, 295–300. [Google Scholar] [CrossRef]
- Sullivan, D.A.; Sullivan, B.D.; Evans, J.E.; Schirra, F.; Yamagami, H.; Liu, M.; Richards, S.M.; Suzuki, T.; Schaumberg, D.A.; Sullivan, R.M.; et al. Androgen deficiency, Meibomian gland dysfunction, and evaporative dry eye. Ann. N. Y. Acad. Sci. 2002, 966, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Schirra, F.; Richards, S.M.; Liu, M.; Suzuki, T.; Yamagami, H.; Sullivan, D.A. Androgen regulation of lipogenic pathways in the mouse meibomian gland. Exp. Eye Res. 2006, 83, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Beauregard, C.; Brandt, P.C. Down regulation of interleukin-1beta-induced nitric oxide production in lacrimal gland acinar cells by sex steroids. Curr. Eye Res. 2004, 29, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Cermak, J.M.; Krenzer, K.L.; Sullivan, R.M.; Dana, M.R.; Sullivan, D.A. Is complete androgen insensitivity syndrome associated with alterations in the meibomian gland and ocular surface? Cornea 2003, 22, 516–521. [Google Scholar] [CrossRef]
- Krenzer, K.L.; Reza Dana, M.; Ullman, M.D.; Cermak, J.M.; Tolls, D.B.; Evans, J.E.; Sullivan, D.A. Effect of androgen deficiency on the human meibomian gland and ocular surface. J. Clin. Endocrinol. Metab. 2000, 85, 4874–4882. [Google Scholar] [CrossRef]
- Grasso, A.; di Zazzo, A.; Giannaccare, G.; Sung, J.; Inomata, T.; Shih, K.C.; Micera, A.; Gaudenzi, D.; Spelta, S.; Romeo, M.A.; et al. Sex Hormones Related Ocular Dryness in Breast Cancer Women. J. Clin. Med. 2021, 10, 2620. [Google Scholar] [CrossRef]
- McKay, T.B.; Priyadarsini, S.; Karamichos, D. Sex Hormones, Growth Hormone, and the Cornea. Cells 2022, 11, 224. [Google Scholar] [CrossRef]
- Tiffany, J. The normal tear film. Dev. Ophthalmol. 2008, 41, 3–6. [Google Scholar]
- Suzuki, T.; Kinoshita, Y.; Tachibana, M.; Matsushima, Y.; Kobayashi, Y.; Adachi, W.; Sotozono, C.; Kinoshita, S. Expression of sex steroid hormone receptors in human cornea. Curr. Eye Res. 2001, 22, 28–33. [Google Scholar] [CrossRef]
- Vécsei, P.V.; Kircher, K.; Kaminski, S.; Nagel, G.; Breitenecker, G.; Kohlberger, P.D. Immunohistochemical detection of estrogen and progesterone receptor in human cornea. Maturitas 2000, 36, 169–172. [Google Scholar] [CrossRef]
- Goldich, Y.; Barkana, Y.; Pras, E.; Fish, A.; Mandel, Y.; Hirsh, A.; Tsur, N.; Morad, Y.; Avni, I.; Zadok, D. Variations in corneal biomechanical parameters and central corneal thickness during the menstrual cycle. J. Cataract Refract. Surg. 2011, 37, 1507–1511. [Google Scholar] [CrossRef] [PubMed]
- Giuffrè, G.; di Rosa, L.; Fiorino, F.; Bubella, D.M.; Lodato, G. Variations in central corneal thickness during the menstrual cycle in women. Cornea 2007, 26, 144–146. [Google Scholar] [CrossRef] [PubMed]
- Lopilly Park, H.Y.; Kim, J.H.; Lee, K.M.; Park, C.K. Effect of prostaglandin analogues on tear proteomics and expression of cytokines and matrix metalloproteinases in the conjunctiva and cornea. Exp. Eye Res. 2012, 94, 13–21. [Google Scholar] [CrossRef]
- Suzuki, T.; Sullivan, D.A. Estrogen stimulation of proinflammatory cytokine and matrix metalloproteinase gene expression in human corneal epithelial cells. Cornea 2005, 24, 1004–1009. [Google Scholar] [CrossRef]
- Naderan, M.; Jahanrad, A. Topographic, tomographic and biomechanical corneal changes during pregnancy in patients with keratoconus: A cohort study. Acta Ophthalmol. 2017, 95, e291–e296. [Google Scholar] [CrossRef] [PubMed]
- Bilgihan, K.; Hondur, A.; Sul, S.; Ozturk, S. Pregnancy-induced progression of keratoconus. Cornea 2011, 30, 991–994. [Google Scholar] [CrossRef] [PubMed]
- Hoogewoud, F.; Gatzioufas, Z.; Hafezi, F. Transitory topographical variations in keratoconus during pregnancy. J. Refract. Surg. 2013, 29, 144–146. [Google Scholar] [CrossRef] [Green Version]
- Soeters, N.; Tahzib, N.G.; Bakker, L.; van der Lelij, A. Two cases of keratoconus diagnosed after pregnancy. Optom. Vis. Sci. 2012, 89, 112–116. [Google Scholar] [CrossRef]
- Escandon, P.; Nicholas, S.E.; Cunningham, R.L.; Murphy, D.A.; Riaz, K.M.; Karamichos, D. The Role of Estriol and Estrone in Keratoconic Stromal Sex Hormone Receptors. Int. J. Mol. Sci. 2022, 23, 916. [Google Scholar] [CrossRef]
- Oh, T.H.; Chang, D.J.; Choi, J.S.; Joo, C.K. Effects of 17β-estradiol on human corneal wound healing in vitro. Cornea 2012, 31, 1158–1164. [Google Scholar] [CrossRef]
- Yan, L.; Wu, W.; Wang, Z.; Li, C.; Lu, X.; Duan, H.; Zhou, J.; Wang, X.; Wan, P.; Song, Y.; et al. Comparative study of the effects of recombinant human epidermal growth factor and basic fibroblast growth factor on corneal epithelial wound healing and neovascularization in vivo and in vitro. Ophthalmic Res. 2013, 49, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wu, X.; Shi, T.; Lu, L. Epidermal growth factor (EGF)-induced corneal epithelial wound healing through nuclear factor κB subtype-regulated CCCTC binding factor (CTCF) activation. J. Biol. Chem. 2013, 288, 24363–24371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leske, M.C.; Wu, S.Y.; Nemesure, B.; Yang, L.; Hennis, A. Nine-year incidence of lens opacities in the Barbados Eye Studies. Ophthalmology 2004, 111, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Livingston, P.M.; Guest, C.S.; Stanislavsky, Y.; Lee, S.; Bayley, S.; Walker, C.; McKean, C.; Taylor, H.R. A population-based estimate of cataract prevalence: The Melbourne Visual Impairment Project experience. Dev. Ophthalmol. 1994, 26, 1–6. [Google Scholar] [PubMed]
- Freeman, E.E.; Munoz, B.; Schein, O.D.; West, S.K. Hormone replacement therapy and lens opacities: The Salisbury Eye Evaluation project. Arch. Ophthalmol. 2001, 119, 1687–1692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, B.E.K. Lens opacities in women in Beaver Dam, Wisconsin: Is there evidence of an effect of sex hormones? Trans. Am. Ophthalmol. Soc. 1993, 91, 517–544. [Google Scholar]
- Beebe, D.C.; Holekamp, N.M.; Shui, Y.B. Oxidative damage and the prevention of age-related cataracts. Ophthalmic Res. 2010, 44, 155–165. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; John, M.; Subramanian, S.; Chen, H.; Carper, D. 17Beta-estradiol confers a protective effect against transforming growth factor-beta2-induced cataracts in female but not male lenses. Exp. Eye Res. 2004, 78, 67–74. [Google Scholar] [CrossRef]
- Hales, A.M.; Chamberlain, C.G.; Murphy, C.R.; McAvoy, J.W. Estrogen protects lenses against cataract induced by transforming growth factor-beta (TGFbeta). J. Exp. Med. 1997, 185, 273–280. [Google Scholar] [CrossRef] [Green Version]
- Younan, C.; Mitchell, P.; Cumming, R.G.; Panchapakesan, J.; Rochtchina, E.; Hales, A.M. Hormone replacement therapy, reproductive factors, and the incidence of cataract and cataract surgery: The Blue Mountains Eye Study. Am. J. Epidemiol. 2002, 155, 997–1006. [Google Scholar] [CrossRef]
- Calkins, D.J. Critical pathogenic events underlying progression of neurodegeneration in glaucoma. Prog. Retin. Eye Res. 2012, 31, 702–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tham, Y.C.; Li, X.; Wong, T.Y.; Quigley, H.A.; Aung, T.; Cheng, C.Y. Global prevalence of glaucoma and projections of glaucoma burden through 2040: A systematic review and meta-analysis. Ophthalmology 2014, 121, 2081–2090. [Google Scholar] [CrossRef] [PubMed]
- Quigley, H.; Broman, A.T. The number of people with glaucoma worldwide in 2010 and 2020. Br. J. Ophthalmol. 2006, 90, 262–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, J.D.; Khawaja, A.P.; Weizer, J.S. Glaucoma in Adults-Screening, Diagnosis, and Management: A Review. JAMA 2021, 325, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Allison, K.; Patel, D.; Alabi, O. Epidemiology of Glaucoma: The Past, Present, and Predictions for the Future. Cureus 2020, 12, e11686. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P.; Smith, W.; Attebo, K.; Healey, P.R. Prevalence of open-angle glaucoma in Australia. The Blue Mountains Eye Study. Ophthalmology 1996, 103, 1661–1669. [Google Scholar] [CrossRef]
- De Voogd, S.; Wolfs, R.C.W.; Jansonius, N.M.; Uitterlinden, A.G.; Pols, H.A.P.; Hofman, A.; De Jong, P.T.V.M. Estrogen receptors alpha and beta and the risk of open-angle glaucoma: The Rotterdam Study. Arch. Ophthalmol. 2008, 126, 110–114. [Google Scholar] [CrossRef] [Green Version]
- Mabuchi, F.; Sakurada, Y.; Kashiwagi, K.; Yamagata, Z.; Iijima, H.; Tsukahara, S. Estrogen receptor beta gene polymorphism and intraocular pressure elevation in female patients with primary open-angle glaucoma. Am. J. Ophthalmol. 2010, 149, 826–830. [Google Scholar] [CrossRef]
- Pasquale, L.R.; Loomis, S.J.; Weinreb, R.N.; Kang, J.H.; Yaspan, B.L.; Cooke Bailey, J.; Gaasterland, D.; Gaasterland, T.; Lee, R.K.; Scott, W.K.; et al. Estrogen pathway polymorphisms in relation to primary open angle glaucoma: An analysis accounting for gender from the United States. Mol. Vis. 2013, 19, 1471–1481. [Google Scholar]
- Feola, A.J.; Fu, J.; Allen, R.; Yang, V.; Campbell, I.C.; Ottensmeyer, A.; Ethier, C.R.; Pardue, M. Menopause exacerbates visual dysfunction in experimental glaucoma. Exp. Eye Res. 2019, 186, 107706. [Google Scholar] [CrossRef]
- Lee, A.J.; Mitchell, P.; Rochtchina, E.; Healey, P.R. Female reproductive factors and open angle glaucoma: The Blue Mountains Eye Study. Br. J. Ophthalmol. 2003, 87, 1324–1328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasquale, L.R.; Rosner, B.A.; Hankinson, S.E.; Kang, J.H. Attributes of female reproductive aging and their relation to primary open-angle glaucoma: A prospective study. J. Glaucoma 2007, 16, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Newman-Casey, P.A.; Talwar, N.; Nan, B.; Musch, D.C.; Pasquale, L.R.; Stein, J.D. The potential association between postmenopausal hormone use and primary open-angle glaucoma. JAMA Ophthalmol. 2014, 132, 298–303. [Google Scholar] [CrossRef] [PubMed]
- Tint, N.L.; Alexander, P.; Tint, K.M.; Vasileiadis, G.T.; Yeung, A.M.; Azuara-Blanco, A. Hormone therapy and intraocular pressure in nonglaucomatous eyes. Menopause 2010, 17, 157–160. [Google Scholar] [CrossRef] [PubMed]
- Baker, A.E.; Brautigam, V.M.; Watters, J.J. Estrogen modulates microglial inflammatory mediator production via interactions with estrogen receptor beta. Endocrinology 2004, 145, 5021–5032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Członkowska, A.; Ciesielska, A.; Gromadzka, G.; Kurkowska-Jastrzebska, I. Gender differences in neurological disease: Role of estrogens and cytokines. Endocrine 2006, 29, 243–256. [Google Scholar] [CrossRef]
- Panchami, S.R.P.; Shenoy, J.P.; Shivakumar, J.; Kole, S.B. Postmenopausal intraocular pressure changes in South Indian females. J. Clin. Diagn. Res. JCDR 2013, 7, 1322–1324. [Google Scholar] [PubMed]
- Wu, A.; Khawaja, A.P.; Pasquale, L.R.; Stein, J.D. A review of systemic medications that may modulate the risk of glaucoma. Eye 2020, 34, 12–28. [Google Scholar] [CrossRef] [PubMed]
- Giorgio, A.; Zhang, J.; Costantino, F.; de Stefano, N.; Frezzotti, P. Diffuse brain damage in normal tension glaucoma. Hum. Brain Mapp. 2018, 39, 532–541. [Google Scholar] [CrossRef] [Green Version]
- Prokai-Tatrai, K.; Xin, H.; Nguyen, V.; Szarka, S.; Blazics, B.; Prokai, L.; Koulen, P. 17β-estradiol eye drops protect the retinal ganglion cell layer and preserve visual function in an in vivo model of glaucoma. Mol. Pharm. 2013, 10, 3253–3261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Man, P.Y.W.; Griffiths, P.G.; Brown, D.T.; Howell, N.; Turnbull, D.M.; Chinnery, P.F. The epidemiology of Leber hereditary optic neuropathy in the North East of England. Am. J. Hum. Genet. 2003, 72, 333–339. [Google Scholar] [CrossRef] [Green Version]
- Carelli, V.; Ross-Cisneros, F.N.; Sadun, A.A. Mitochondrial dysfunction as a cause of optic neuropathies. Prog. Retin. Eye Res. 2004, 23, 53–89. [Google Scholar] [CrossRef] [PubMed]
- Giordano, C.; Montopoli, M.; Perli, E.; Orlandi, M.; Fantin, M.; Ross-Cisneros, F.N.; Caparrotta, L.; Martinuzzi, A.; Ragazzi, E.; Ghelli, A.; et al. Oestrogens ameliorate mitochondrial dysfunction in Leber’s hereditary optic neuropathy. Brain J. Neurol. 2011, 134, 220–234. [Google Scholar] [CrossRef]
- Pisano, A.; Preziuso, C.; Iommarini, L.; Perli, E.; Grazioli, P.; Campese, A.F.; Maresca, A.; Montopoli, M.; Masuelli, L.; Sadun, A.A.; et al. Targeting estrogen receptor β as preventive therapeutic strategy for Leber’s hereditary optic neuropathy. Hum. Mol. Genet. 2015, 24, 6921–6931. [Google Scholar] [CrossRef] [Green Version]
- Miller, D.; Barkhof, F.; Montalban, X.; Thompson, A.; Filippi, M. Clinically isolated syndromes suggestive of multiple sclerosis, part I: Natural history, pathogenesis, diagnosis, and prognosis. Lancet. Neurol. 2005, 4, 281–288. [Google Scholar] [CrossRef]
- Soldan, S.S.; Retuerto, A.I.A.; Sicotte, N.L.; Voskuhl, R.R. Immune modulation in multiple sclerosis patients treated with the pregnancy hormone estriol. J. Immunol. 2003, 171, 6267–6274. [Google Scholar] [CrossRef] [PubMed]
- Kim, R.Y.; Mangu, D.; Hoffman, A.S.; Kovash, R.; Jung, E.; Itoh, N.; Voskuhl, R. Oestrogen receptor β ligand acts on CD11c+ cells to mediate protection in experimental autoimmune encephalomyelitis. Brain J. Neurol. 2018, 141, 132–147. [Google Scholar]
- El-Etr, M.; Rame, M.; Boucher, C.; Ghoumari, A.M.; Kumar, N.; Liere, P.; Pianos, A.; Schumacher, M.; Sitruk-Ware, R. Progesterone and nestorone promote myelin regeneration in chronic demyelinating lesions of corpus callosum and cerebral cortex. Glia 2015, 63, 104–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, R.; Ghoumari, A.M.; Bielecki, B.; Steibel, J.; Boehm, N.; Liere, P.; MacKlin, W.B.; Kumar, N.; Habert, R.; Mhaouty-Kodja, S.; et al. The neural androgen receptor: A therapeutic target for myelin repair in chronic demyelination. Brain J. Neurol. 2013, 136, 132–146. [Google Scholar] [CrossRef] [PubMed]
- Cascio, C.; Russo, D.; Drago, G.; Galizzi, G.; Passantino, R.; Guarneri, R.; Guarneri, P. 17beta-estradiol synthesis in the adult male rat retina. Exp. Eye Res. 2007, 85, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Mellon, S.H.; Griffin, L.D.; Compagnone, N.A. Biosynthesis and action of neurosteroids. Brain Res. Brain Res. Rev. 2001, 37, 3–12. [Google Scholar] [CrossRef]
- Cascio, C.; Deidda, I.; Russo, D.; Guarneri, P. The estrogenic retina: The potential contribution to healthy aging and age-related neurodegenerative diseases of the retina. Steroids 2015, 103, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Gotovac, M.; Kaštelan, S.; Lukenda, A. Eye and pregnancy. Coll. Antropol. 2013, 37, 189–193. [Google Scholar] [PubMed]
- Mangiamele, L.A.; Gomez, J.R.; Curtis, N.J.; Thompson, R.R. GPER/GPR30, a membrane estrogen receptor, is expressed in the brain and retina of a social fish (Carassius auratus) and colocalizes with isotocin. J. Comp. Neurol. 2017, 525, 252–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakravarthy, U.; Wong, T.Y.; Fletcher, A.; Piault, E.; Evans, C.; Zlateva, G.; Buggage, R.; Pleil, A.; Mitchell, P. Clinical risk factors for age-related macular degeneration: A systematic review and meta-analysis. BMC Ophthalmol. 2010, 10, 31. [Google Scholar] [CrossRef] [PubMed]
- Ardourel, J.E. Risk factors associated with age-related macular degeneration. A case-control study in the age-related eye disease study: Age-Related Eye Disease Study Report Number 3. Ophthalmology 2000, 107, 2224–2232. [Google Scholar]
- Kaarniranta, K.; Machalińska, A.; Veréb, Z.; Salminen, A.; Petrovski, G.; Kauppinen, A. Estrogen signalling in the pathogenesis of age-related macular degeneration. Curr. Eye Res. 2015, 40, 226–233. [Google Scholar] [CrossRef]
- Snow, K.K.; Cote, J.; Yang, W.; Davis, N.J.; Seddon, J.M. Association between reproductive and hormonal factors and age-related maculopathy in postmenopausal women. Am. J. Ophthalmol. 2002, 134, 842–848. [Google Scholar] [CrossRef]
- Chakravarthy, U.; Williams, M.; Amoaku, W.; Bailey, C.; Bishop, P.; Brand, C.; Chong, V.; Downes, S.; Evans, J.; Lotery, A.; et al. The Royal College of Ophthalmologists Guidelines on AMD: Executive Summary. Eye 2013, 27, 1429–1431. [Google Scholar] [CrossRef]
- Evans, J.R. Risk factors for age-related macular degeneration. Prog. Retin. Eye Res. 2001, 20, 227–253. [Google Scholar] [CrossRef]
- Liew, G.; Quin, G.; Gillies, M.; Fraser-Bell, S. Central serous chorioretinopathy: A review of epidemiology and pathophysiology. Clin. Exp. Ophthalmol. 2013, 41, 201–214. [Google Scholar] [CrossRef] [PubMed]
- Ross, A.; Ross, A.H.; Mohamed, Q. Review and update of central serous chorioretinopathy. Curr. Opin. Ophthalmol. 2011, 22, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Iida, T.; Kishi, S.; Hagimura, N.; Shimizu, K. Persistent and bilateral choroidal vascular abnormalities in central serous chorioretinopathy. Retina 1999, 19, 508. [Google Scholar] [CrossRef] [PubMed]
- Grieshaber, M.C.; Staub, J.J.; Flammer, J. The potential role of testosterone in central serous chorioretinopathy. Br. J. Ophthalmol. 2007, 91, 118–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahad, M.A.; Chua, C.N.; Evans, N.M. Central serous chorioretinopathy associated with testosterone therapy. Eye 2006, 20, 503–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nudleman, E.; Witmer, M.T.; Kiss, S.; Williams, G.A.; Wolfe, J.D. Central serous chorioretinopathy in patients receiving exogenous testosterone therapy. Retina 2014, 34, 2128–2132. [Google Scholar] [CrossRef]
- Campochiaro, P.A.; Mir, T.A. The mechanism of cone cell death in Retinitis Pigmentosa. Prog. Retin. Eye Res. 2018, 62, 24–37. [Google Scholar] [CrossRef]
- Kennan, A.; Aherne, A.; Humphries, P. Light in retinitis pigmentosa. Trends Genet. TIG 2005, 21, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Roche, S.L.; Ruiz-Lopez, A.M.; Moloney, J.N.; Byrne, A.M.; Cotter, T.G. Microglial-induced Müller cell gliosis is attenuated by progesterone in a mouse model of retinitis pigmentosa. Glia 2018, 66, 295–310. [Google Scholar] [CrossRef]
- Sánchez-Vallejo, V.; Benlloch-Navarro, S.; López-Pedrajas, R.; Romero, F.J.; Miranda, M. Neuroprotective actions of progesterone in an in vivo model of retinitis pigmentosa. Pharmacol. Res. 2015, 99, 276–288. [Google Scholar] [CrossRef]
- Roche, S.L.; Kutsyr, O.; Cuenca, N.; Cotter, T.G. Norgestrel, a Progesterone Analogue, Promotes Significant Long-Term Neuroprotection of Cone Photoreceptors in a Mouse Model of Retinal Disease. Investig. Ophthalmol. Vis. Sci. 2019, 60, 3221–3235. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Ocular Disorder | Evidence of a Role of Sex Hormones on Pathophysiology | Sex Hormones Involved |
|---|---|---|
| Dry eye disease (DED) | ++ | Androgens, Estrogens |
| Corneal disorders | ++ | Estrogens |
| Cataract | + | Estrogens |
| Glaucoma | + | Estrogens |
| Leber’s hereditary optic neuropathy (LHON) | ++ | Estrogens |
| Demyelinating optic neuritis | ++ | Estrogens, Progesterone, Androgens |
| Age-related macular degeneration (AMD) | ± | Estrogens |
| Central serous chorioretinopathy (CSCR) | ± | Estrogens |
| Retinitis pigmentosa | ++ | Progesterone |
| Ocular Disorder | Effects of Sex Hormones |
|---|---|
| Dry eye disease (DED) |
|
| Corneal disorders |
|
| Cataract |
|
| Glaucoma |
|
| Leber’s hereditary optic neuropathy (LHON) |
|
| Demyelinating optic neuritis |
|
| Age-related macular degeneration (AMD) |
|
| Central serous chorioretinopathy (CSCR) |
|
| Retinitis pigmentosa |
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nuzzi, R.; Caselgrandi, P. Sex Hormones and Their Effects on Ocular Disorders and Pathophysiology: Current Aspects and Our Experience. Int. J. Mol. Sci. 2022, 23, 3269. https://doi.org/10.3390/ijms23063269
Nuzzi R, Caselgrandi P. Sex Hormones and Their Effects on Ocular Disorders and Pathophysiology: Current Aspects and Our Experience. International Journal of Molecular Sciences. 2022; 23(6):3269. https://doi.org/10.3390/ijms23063269
Chicago/Turabian StyleNuzzi, Raffaele, and Paolo Caselgrandi. 2022. "Sex Hormones and Their Effects on Ocular Disorders and Pathophysiology: Current Aspects and Our Experience" International Journal of Molecular Sciences 23, no. 6: 3269. https://doi.org/10.3390/ijms23063269
APA StyleNuzzi, R., & Caselgrandi, P. (2022). Sex Hormones and Their Effects on Ocular Disorders and Pathophysiology: Current Aspects and Our Experience. International Journal of Molecular Sciences, 23(6), 3269. https://doi.org/10.3390/ijms23063269