
Methylation of Subtelomeric Chromatin Modifies the Expression of the lncRNA TERRA, Disturbing Telomere Homeostasis

 , , , , , , , and
, , , , , , , and 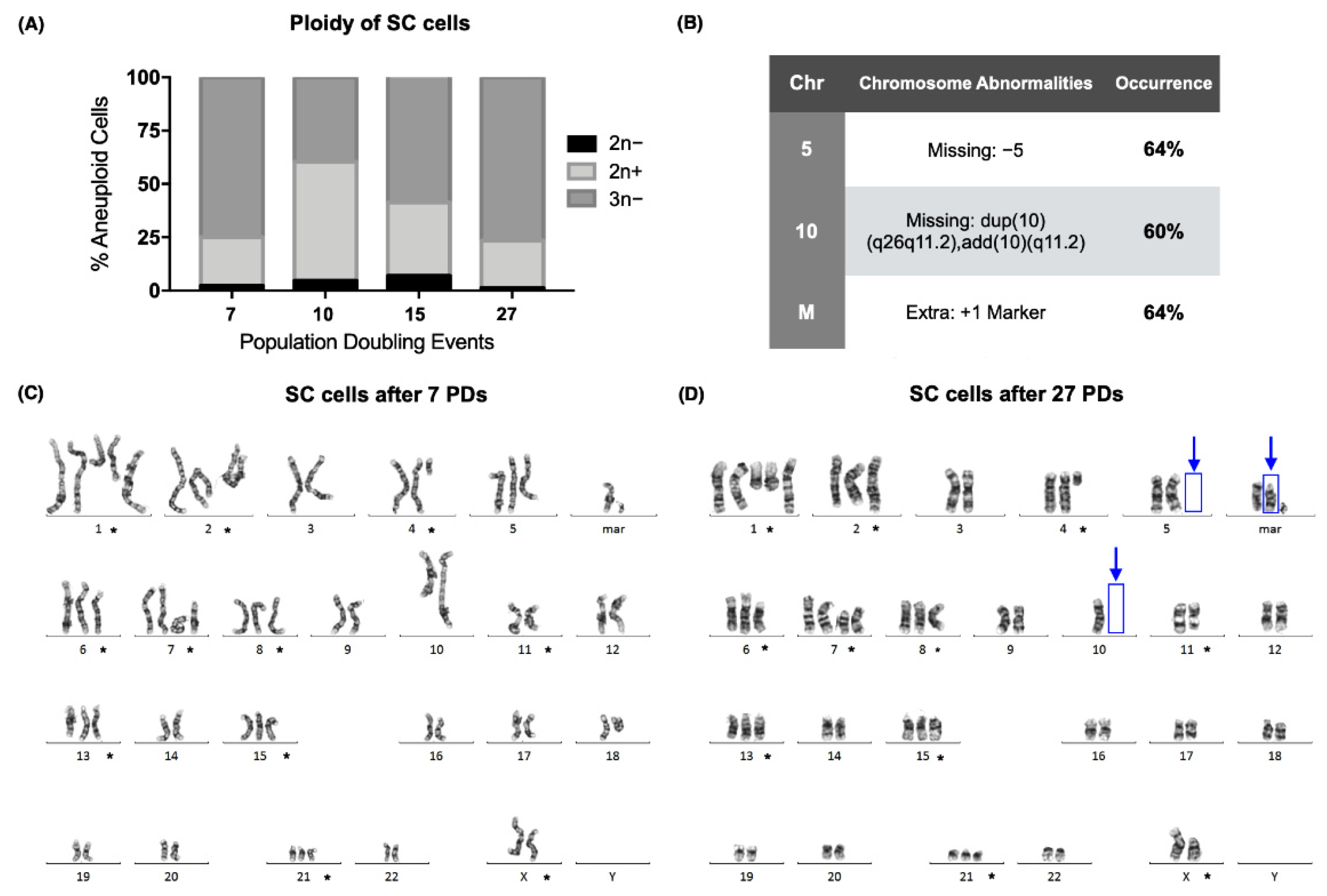{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
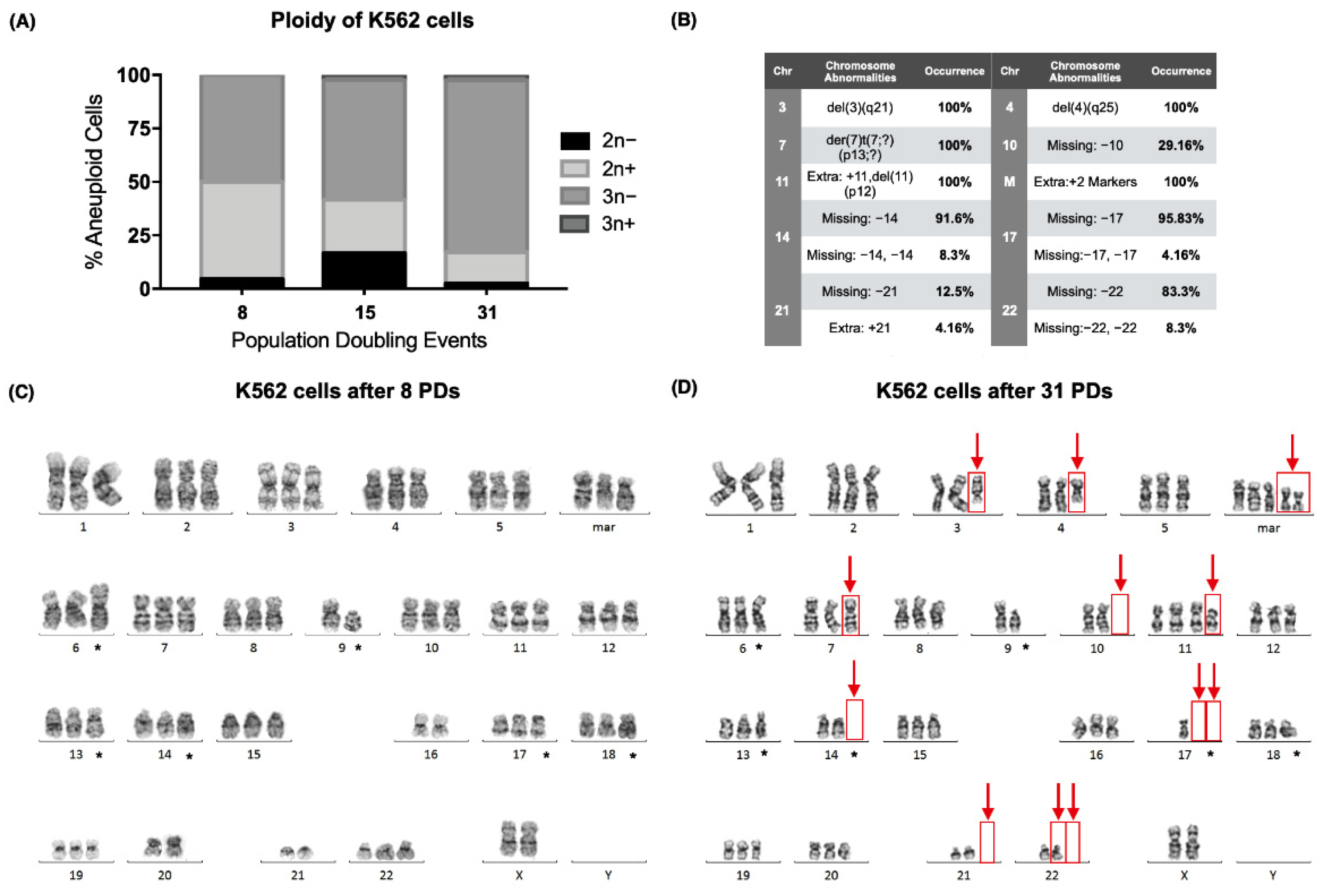
2.1. Karyotype Analysis
2.2. Telomerase Expression
2.3. Global TERRA Levels
2.4. Locus-Specific Expression of TERRA
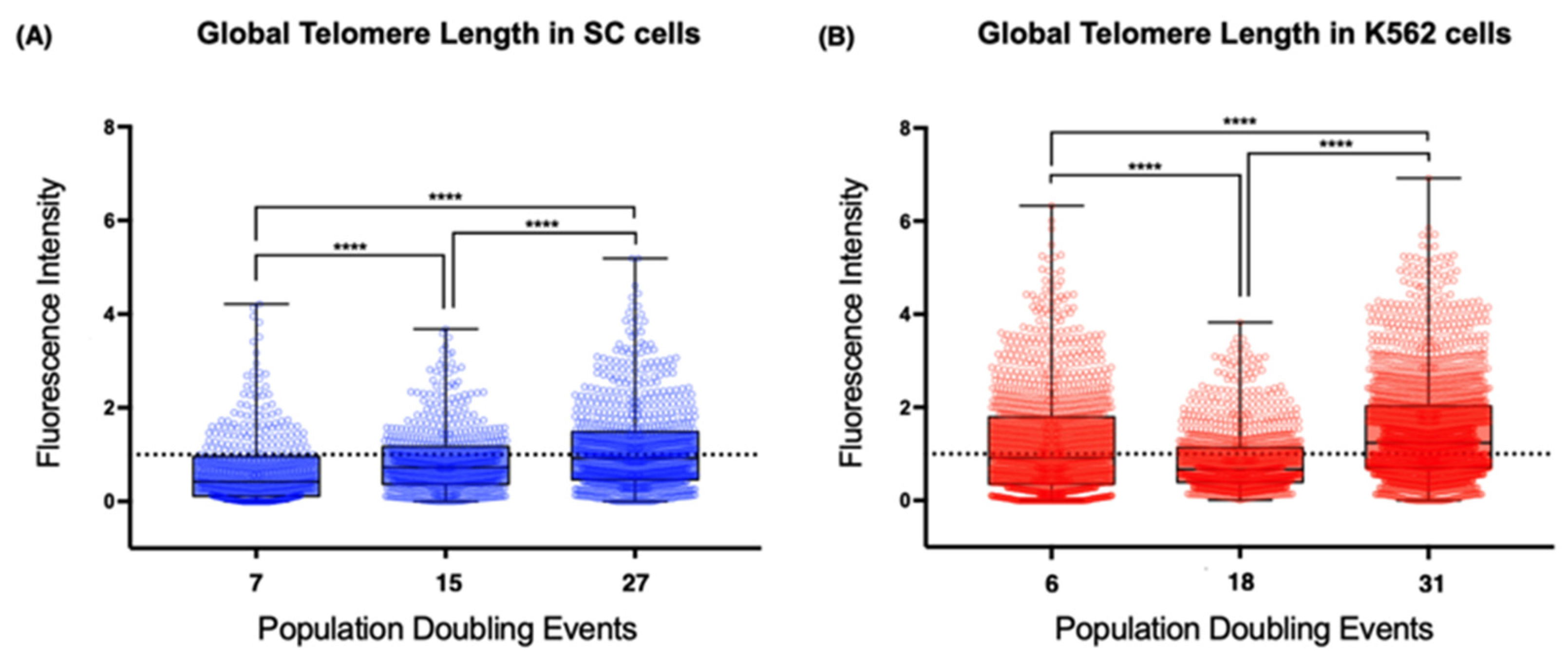
2.5. Global Telomeric Length
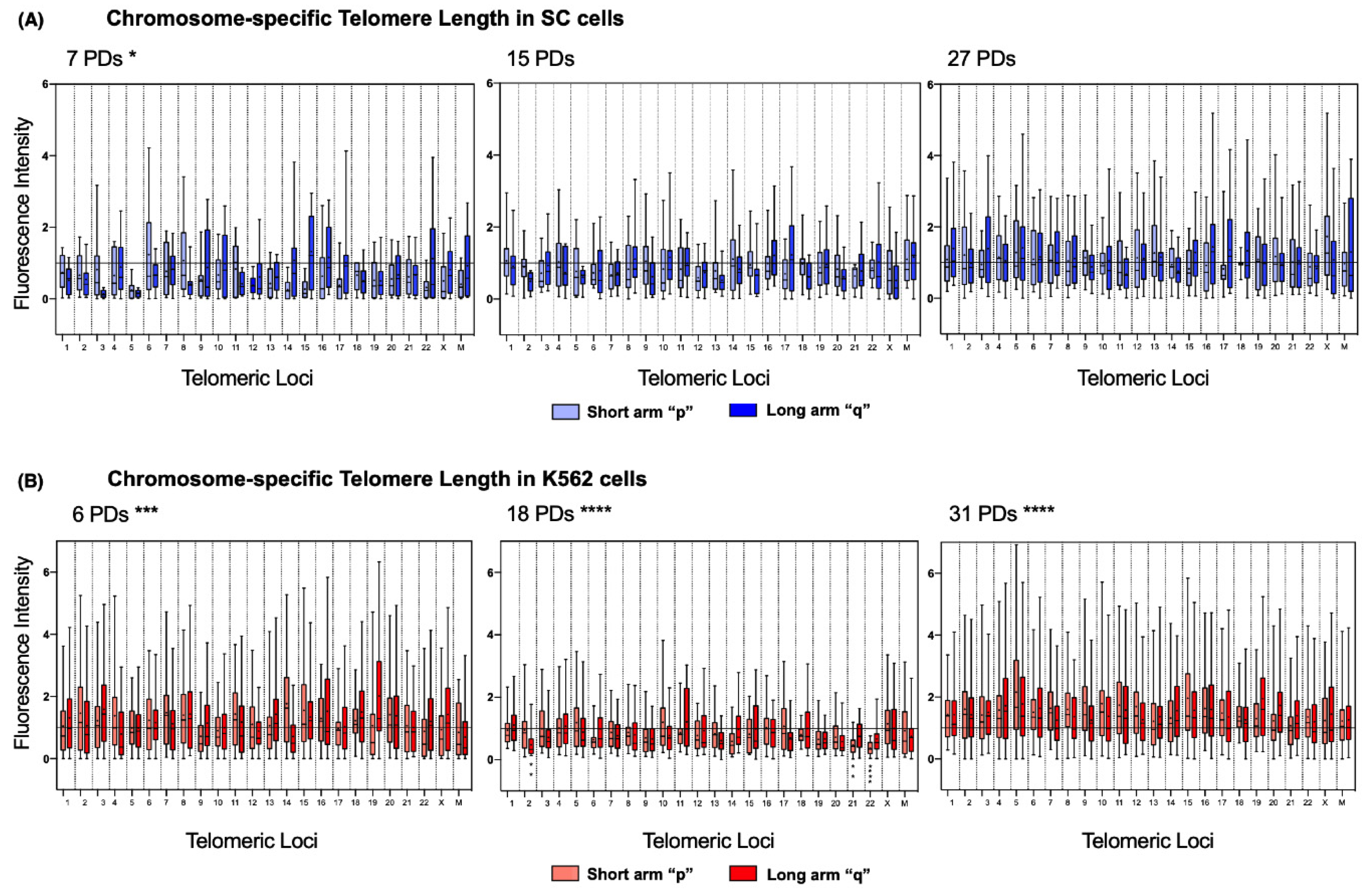
2.6. Chromosome-Specific Telomere Length Analysis
2.7. Locus 5p and 10q-Specific Telomeric Length
2.8. Subtelomeric Chromatin Analysis
2.8.1. SC Cell Line—Heterochromatin
- Both loci 5p and 10q accumulated heterochromatin-associated histone marks as the cell population aged. In 5p, H3K9me3 increased 4.4-fold from its initial value after 15 PDs. The H3K27me3 mark increased 9.73-fold after 15 PDs and 7.29-fold after 25 PDs (Figure 8A,C). H4K20me3 was undetectable after 25 PDs (Figure 9A).
2.8.2. K562 Cell Line—Heterochromatin
- When we analyzed the same chromatin marks in locus 5p of the K562 cell populations, we identified a similar phenomenon to that in the SC cells. Both H3K9me3 and H3K27me3 displayed a 1.8- and 1.49-fold enrichment, respectively, after 24 PDs (Figure 8E,G). H4K20me3 showed a 1.12-fold increase after 14 PDs, but after 24 PDs, the levels of this mark returned to their initial value (Figure 9E).
- At locus 10q, neither H3K9me3 nor H3K27me3 showed differences. H3K9me3 increased 0.79-fold after 14 PDs and then returned to its initial level after 24 PDs. The levels of H3K27me3 diminished 0.61-fold after 14 PDs and 0.83-fold after 24 PDs (Figure 8F,H). H4K20me3 displayed a 0.79-fold enrichment after 14 PDs, but the levels of this mark also decreased up to 0.76-fold below its initial levels after 24 PDs (Figure 9F).
2.8.3. SC Cell Line—Euchromatin
2.8.4. K562 Cell Line—Euchromatin
- In the 5p locus of K562 cells, the levels of Pol II decreased 0.59-fold after 24 PDs. CTCF was enriched 1.5-fold after 14 PDs, but its levels returned to their initial value after 24 PDs (Figure 10E,G). H3K4me3 also increased 1.5-fold after 14 PDs, but after 24 PDs, the levels of this mark returned to their original value (Figure 9G).
- At locus 10q, the levels of Pol II decreased 0.3-fold after 14 PDs and 0.675-fold after 24 PDs. However, the levels of CTCF increased 6.48-fold after 14 PDs but then returned to their initial values after 24 PDs (Figure 10F,H). Finally, H3K4me3 diminished steadily 0.36-fold after 14 PDs and 0.53-fold after 24 PDs (Figure 9H).
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Metaphase Preparation
4.3. Conventional Karyotype Analysis
4.4. Northern Blot
4.5. Primer Design
4.6. Analysis of hTERT and TERRA Expression
4.7. Q-FISH for Telomere Length Analysis
4.8. Image Processing
4.9. Chromatin Immunoprecipitation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Deng, Z.; Norseen, J.; Wiedmer, A.; Riethman, H.; Lieberman, P.M. TERRA RNA Binding to TRF2 Facilitates Heterochromatin Formation and ORC Recruitment at Telomeres. Mol. Cell 2009, 35, 403–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffith, J.D.; Comeau, L.; Rosenfield, S.; Stansel, R.M.; Bianchi, A.; Moss, H.; Lange, T. de Mammalian Telomeres End in a Large Duplex Loop. Cell 1999, 97, 503–514. [Google Scholar] [CrossRef] [Green Version]
- Arnoult, N.; Beneden, A.V.; Decottignies, A. Telomere Length Regulates TERRA Levels through Increased Trimethylation of Telomeric H3K9 and HP1α. Nat. Struct. Mol. Biol. 2012, 19, 948–956. [Google Scholar] [CrossRef] [PubMed]
- Oliva-Rico, D.; Herrera, L.A. Regulated Expression of the LncRNA TERRA and Its Impact on Telomere Biology. Mech. Ageing Dev. 2017, 167, 16–23. [Google Scholar] [CrossRef]
- Flynn, R.L.; Centore, R.C.; O’Sullivan, R.J.; Rai, R.; Tse, A.; Songyang, Z.; Chang, S.; Karlseder, J.; Zou, L. TERRA and HnRNPA1 Orchestrate an RPA-to-POT1 Switch on Telomeric Single-Stranded DNA. Nature 2012, 471, 532–536. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Komiyama, M. Structure, Function and Targeting of Human Telomere RNA. Methods 2012, 57, 100–105. [Google Scholar] [CrossRef]
- Deng, Z.; Wang, Z.; Stong, N.; Plasschaert, R.; Moczan, A.; Chen, H.-S.; Hu, S.; Wikramasinghe, P.; Davuluri, R.V.; Bartolomei, M.S.; et al. A Role for CTCF and Cohesin in Subtelomere Chromatin Organization, TERRA Transcription, and Telomere End Protection. EMBO J. 2012, 31, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Thijssen, P.E.; Tobi, E.W.; Balog, J.; Schouten, S.G.; Kremer, D.; Bouazzaoui, F.E.; Henneman, P.; Putter, H.; Slagboom, P.E.; Heijmans, B.T.; et al. Chromatin remodeling of human subtelomeres and TERRA promoters upon cellular senescence: Commonalities and differences between chromosomes. Epigenetics 2013, 8, 512–521. [Google Scholar] [CrossRef] [Green Version]
- Beishline, K.; Vladimirova, O.; Tutton, S.; Wang, Z.; Deng, Z.; Lieberman, P.M. CTCF Driven TERRA Transcription Facilitates Completion of Telomere DNA Replication. Nat. Commun. 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Mei, Y.; Deng, Z.; Vladimirova, O.; Gulve, N.; Johnson, F.B.; Drosopoulos, W.C.; Schildkraut, C.L.; Lieberman, P.M. TERRA G-Quadruplex RNA Interaction with TRF2 GAR Domain Is Required for Telomere Integrity. Sci. Rep. 2021, 11, 3509. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. The Hallmarks of Cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Farnung, B.O.; Brun, C.M.; Arora, R.; Lorenzi, L.E.; Azzalin, C.M. Telomerase Efficiently Elongates Highly Transcribing Telomeres in Human Cancer Cells. PLoS ONE 2012, 7, e35714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arora, R.; Lee, Y.; Wischnewski, H.; Brun, C.M.; Schwarz, T.; Azzalin, C.M. RNaseH1 Regulates TERRA-Telomeric DNA Hybrids and Telomere Maintenance in ALT Tumour Cells. Nat. Commun. 2014, 5, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutschner, T.; Diederichs, S. The Hallmarks of Cancer. RNA Biol. 2012, 9, 703–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryan, T.M.; Englezou, A.; Gupta, J.; Bacchetti, S.; Reddel, R.R. Telomere Elongation in Immortal Human Cells without Detectable Telomerase Activity. EMBO J. 1995, 14, 4240–4248. [Google Scholar] [CrossRef] [PubMed]
- Shay, J.W.; Bacchetti, S. A Survey of Telomerase Activity in Human Cancer. Eur. J. Cancer 1997, 33, 787–791. [Google Scholar] [CrossRef]
- Shay, J.W.; Wright, W.E. Hayflick, His Limit, and Cellular Ageing. Nat. Rev. Mol. Cell Biol. 2000, 1, 72–76. [Google Scholar] [CrossRef]
- Collado, M.; Blasco, M.A.; Serrano, M. Cellular Senescence in Cancer and Aging. Cell 2007, 130, 223–233. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Collins, G.W.; Largen, M.T. Continuous Mammalian Cell Lines Having Monocyte/Macrophage Characteristics and Their Establishment In Vitro. U.S. Patent US5447861A, 5 September 1995. [Google Scholar]
- Nogueira, B.M.D.; Machado, C.B.; Montenegro, R.C.; Moraes, M.E.A.D.; Moreira-Nunes, C.A. Telomere Length and Hematological Disorders: A Review. In Vivo 2020, 34, 3093–3101. [Google Scholar] [CrossRef]
- Zinn, R.L.; Pruitt, K.; Eguchi, S.; Baylin, S.B.; Herman, J.G. hTERT Is Expressed in Cancer Cell Lines Despite Promoter DNA Methylation by Preservation of Unmethylated DNA and Active Chromatin around the Transcription Start Site. Cancer Res. 2007, 67, 194–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGowan-Jordan, J.; Hastings, R.J.; Moore, S. ISCN 2020: An International System for Human Cytogenomic Nomenclature; Karger: Basel, Switzerland, 2020; ISBN 9783318067064. [Google Scholar]
- Scheel, C.; Schaefer, K.-L.; Jauch, A.; Keller, M.; Wai, D.; Brinkschmidt, C.; van Valen, F.; Boecker, W.; Dockhorn-Dworniczak, B.; Poremba, C. Alternative Lengthening of Telomeres Is Associated with Chromosomal Instability in Osteosarcomas. Oncogene 2001, 20, 3835–3844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coding Potential Calculator. Available online: http://cpc2.gao-lab.org/index.php (accessed on 25 January 2021).
- Kang, Y.-J.; Yang, D.-C.; Kong, L.; Hou, M.; Meng, Y.-Q.; Wei, L.; Gao, G. CPC2: A Fast and Accurate Coding Potential Calculator Based on Sequence Intrinsic Features. Nucleic Acids Res. 2017, 45, 12–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, L.; Zhang, Y.; Ye, Z.-Q.; Liu, X.-Q.; Zhao, S.-Q.; Wei, L.; Gao, G. CPC: Assess the Protein-Coding Potential of Transcripts Using Sequence Features and Support Vector Machine. Nucleic Acids Res. 2007, 35, 345–349. [Google Scholar] [CrossRef]
- Lange, T. de Shelterin: The Protein Complex That Shapes and Safeguards Human Telomeres. Gene Dev. 2005, 19, 2100–2110. [Google Scholar] [CrossRef] [Green Version]
- Jones, N. Human Cytogenetics: Malignancy and Acquired Abnormalities, Third Edition, A Practical Approach. Chromosome Res. 2002, 10, 176. [Google Scholar] [CrossRef]
- Halvorsen, T.L.; Leibowitz, G.; Levine, F. Telomerase Activity Is Sufficient To Allow Transformed Cells To Escape from Crisis. Mol. Cell Biol. 1999, 19, 1864–1870. [Google Scholar] [CrossRef] [Green Version]
- Kamal, S.; Junaid, M.; Ejaz, A.; Bibi, I.; Akash, M.S.H.; Rehman, K. The Secrets of Telomerase: Retrospective Analysis and Future Prospects. Life Sci. 2020, 257, 118115. [Google Scholar] [CrossRef]
- Lewis, K.A.; Tollefsbol, T.O. Regulation of the Telomerase Reverse Transcriptase Subunit through Epigenetic Mechanisms. Front. Genet. 2016, 7, 83. [Google Scholar] [CrossRef] [Green Version]
- Giardini, M.A.; Segatto, M.; da Silva, M.S.; Nunes, V.S.; Cano, M.I.N. Telomere and Telomerase Biology. In Telomeres in Health and Disease; Elsevier: Amsterdam, The Netherlands, 2014; Volume 125, pp. 1–40. ISBN 9780123978981. [Google Scholar]
- Ropio, J.; Merlio, J.-P.; Soares, P.; Chevret, E. Telomerase Activation in Hematological Malignancies. Genes 2016, 7, 61. [Google Scholar] [CrossRef] [Green Version]
- Vicente-Dueñas, C.; Barajas-Diego, M.; Romero-Camarero, I.; González-Herrero, I.; Flores, T.; Sánchez-García, I. Essential Role for Telomerase in Chronic Myeloid Leukemia Induced by BCR-ABL in Mice. Oncotarget 2012, 3, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Roake, C.M.; Artandi, S.E. Regulation of Human Telomerase in Homeostasis and Disease. Nat. Rev. Mol. Cell Biol. 2020, 21, 384–397. [Google Scholar] [CrossRef] [PubMed]
- Deville, L.; Hillion, J.; Ségal-Bendirdjian, E. Telomerase Regulation in Hematological Cancers: A Matter of Stemness? Biochim. Biophys. Acta Mol. Basis Dis. 2009, 1792, 229–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Wang, S.; Popova, E.Y.; Grigoryev, S.A.; Zhu, J. Rearrangement of Upstream Sequences of the hTERT Gene during Cellular Immortalization. Genes Chromosomes Cancer 2009, 48, 963–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, A.; Zheng, C.; Lindvall, C.; Hou, M.; Ekedahl, J.; Lewensohn, R.; Yan, Z.; Yang, X.; Henriksson, M.; Blennow, E.; et al. Frequent Amplification of the Telomerase Reverse Transcriptase Gene in Human Tumors. Cancer Res. 2000, 60, 6230–6235. [Google Scholar]
- Heidenreich, B.; Kumar, R. TERT Promoter Mutations in Telomere Biology. Mutat. Res. Rev. 2017, 771, 15–31. [Google Scholar] [CrossRef]
- Barthel, F.P.; Wei, W.; Tang, M.; Martinez-Ledesma, E.; Hu, X.; Amin, S.B.; Akdemir, K.C.; Seth, S.; Song, X.; Wang, Q.; et al. Systematic Analysis of Telomere Length and Somatic Alterations in 31 Cancer Types. Nat. Genet. 2017, 49, 349–357. [Google Scholar] [CrossRef]
- Greider, C.W. Telomerase RNA Levels Limit the Telomere Length Equilibrium. Cold Spring Harb. Sym. 2006, 71, 225–229. [Google Scholar] [CrossRef]
- Cristofari, G.; Lingner, J. Telomere Length Homeostasis Requires That Telomerase Levels Are Limiting. EMBO J. 2006, 25, 565–574. [Google Scholar] [CrossRef]
- Zhu, J.; Zhao, Y.; Wang, S. Chromatin and Epigenetic Regulation of the Telomerase Reverse Transcriptase Gene. Protein Cell 2010, 1, 22–32. [Google Scholar] [CrossRef] [Green Version]
- Episkopou, H.; Draskovic, I.; Beneden, A.V.; Tilman, G.; Mattiussi, M.; Gobin, M.; Arnoult, N.; Londoño-Vallejo, A.; Decottignies, A. Alternative Lengthening of Telomeres Is Characterized by Reduced Compaction of Telomeric Chromatin. Nucleic Acids Res. 2014, 42, 4391–4405. [Google Scholar] [CrossRef] [PubMed]
- Beneden, A.V.; Arnoult, N.; Decottignies, A. Telomeric RNA Expression: Length Matters. Front. Oncol. 2013, 3, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arora, R.; Azzalin, C.M. Telomere Elongation Chooses TERRA ALTernatives. RNA Biol. 2015, 12, 938–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, B.; Arora, R.; Bione, S.; Azzalin, C.M. TERRA Transcription Destabilizes Telomere Integrity to Initiate Break-Induced Replication in Human ALT Cells. Nat. Commun. 2021, 12, 3760. [Google Scholar] [CrossRef]
- Véronèse, L.; Tournilhac, O.; Callanan, M.; Prie, N.; Kwiatkowski, F.; Combes, P.; Chauvet, M.; Davi, F.; Gouas, L.; Verrelle, P.; et al. Telomeres and Chromosomal Instability in Chronic Lymphocytic Leukemia. Leukemia 2013, 27, 490–493. [Google Scholar] [CrossRef]
- Shay, J.W.; Wright, W.E. Role of Telomeres and Telomerase in Cancer. Semin. Cancer Biol. 2011, 21, 349–353. [Google Scholar] [CrossRef] [Green Version]
- Counter, C.M.; Botelho, F.M.; Wang, P.; Harley, C.B.; Bacchetti, S. Stabilization of Short Telomeres and Telomerase Activity Accompany Immortalization of Epstein-Barr Virus-Transformed Human B Lymphocytes. J. Virol. 1994, 68, 3410–3414. [Google Scholar] [CrossRef] [Green Version]
- Counter, C.M.; Avilion, A.A.; LeFeuvre, C.E.; Stewart, N.G.; Greider, C.W.; Harley, C.B.; Bacchetti, S. Telomere Shortening Associated with Chromosome Instability Is Arrested in Immortal Cells Which Express Telomerase Activity. EMBO J. 1992, 11, 1921–1929. [Google Scholar] [CrossRef]
- Hemann, M.T.; Strong, M.A.; Hao, L.-Y.; Greider, C.W. The Shortest Telomere, Not Average Telomere Length, Is Critical for Cell Viability and Chromosome Stability. Cell 2001, 107, 67–77. [Google Scholar] [CrossRef] [Green Version]
- Kyo, S.; Takakura, M.; Fujiwara, T.; Inoue, M. Understanding and Exploiting hTERT Promoter Regulation for Diagnosis and Treatment of Human Cancers. Cancer Sci 2008, 99, 1528–1538. [Google Scholar] [CrossRef] [Green Version]
- McClintock, B. The Fusion of Broken Ends of Chromosomes Following Nuclear Fusion. Proc Natl. Acad. Sci. USA 1942, 28, 458–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barral, A.; Déjardin, J. Telomeric Chromatin and TERRA. J. Mol. Biol. 2020, 432, 4244–4256. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Ludlow, A.T.; Min, J.; Robin, J.D.; Stadler, G.; Mender, I.; Lai, T.-P.; Zhang, N.; Wright, W.E.; Shay, J.W. Regulation of the Human Telomerase Gene TERT by Telomere Position Effect-Over Long Distances (TPE-OLD): Implications for Aging and Cancer. PLoS Biol. 2016, 14, e2000016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, J.S.; Nicetto, D.; Zaret, K.S. H3K9me3-Dependent Heterochromatin: Barrier to Cell Fate Changes. Trends Genet. 2016, 32, 29–41. [Google Scholar] [CrossRef] [Green Version]
- Nicetto, D.; Zaret, K.S. Role of H3K9me3 Heterochromatin in Cell Identity Establishment and Maintenance. Curr. Opin. Genet. Dev. 2019, 55, 1–10. [Google Scholar] [CrossRef]
- Jamieson, K.; Wiles, E.T.; McNaught, K.J.; Sidoli, S.; Leggett, N.; Shao, Y.; Garcia, B.A.; Selker, E.U. Loss of HP1 Causes Depletion of H3K27me3 from Facultative Heterochromatin and Gain of H3K27me2 at Constitutive Heterochromatin. Genome Res. 2016, 26, 97–107. [Google Scholar] [CrossRef] [Green Version]
- Nelson, D.M.; Jaber-Hijazi, F.; Cole, J.J.; Robertson, N.A.; Pawlikowski, J.S.; Norris, K.T.; Criscione, S.W.; Pchelintsev, N.A.; Piscitello, D.; Stong, N.; et al. Mapping H4K20me3 onto the Chromatin Landscape of Senescent Cells Indicates a Function in Control of Cell Senescence and Tumor Suppression through Preservation of Genetic and Epigenetic Stability. Genome Biol. 2016, 17, 158. [Google Scholar] [CrossRef] [Green Version]
- Benetti, R.; Gonzalo, S.; Jaco, I.; Schotta, G.; Klatt, P.; Jenuwein, T.; Blasco, M.A. Suv4-20h Deficiency Results in Telomere Elongation and Derepression of Telomere Recombination. J. Cell Biol. 2007, 178, 925–936. [Google Scholar] [CrossRef] [Green Version]
- Wiles, E.T.; Selker, E.U. H3K27 Methylation: A Promiscuous Repressive Chromatin Mark. Curr. Opin. Genet. Dev. 2017, 43, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Guillemette, B.; Drogaris, P.; Lin, H.-H.S.; Armstrong, H.; Hiragami-Hamada, K.; Imhof, A.; Bonneil, É.; Thibault, P.; Verreault, A.; Festenstein, R.J. H3 Lysine 4 Is Acetylated at Active Gene Promoters and Is Regulated by H3 Lysine 4 Methylation. PLoS Genet. 2011, 7, e1001354. [Google Scholar] [CrossRef] [Green Version]
- Barski, A.; Cuddapah, S.; Cui, K.; Roh, T.-Y.; Schones, D.E.; Wang, Z.; Wei, G.; Chepelev, I.; Zhao, K. High-Resolution Profiling of Histone Methylations in the Human Genome. Cell 2007, 129, 823–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wendt, K.S.; Yoshida, K.; Itoh, T.; Bando, M.; Koch, B.; Schirghuber, E.; Tsutsumi, S.; Nagae, G.; Ishihara, K.; Mishiro, T.; et al. Cohesin Mediates Transcriptional Insulation by CCCTC-Binding Factor. Nature 2008, 451, 796–801. [Google Scholar] [CrossRef]
- Dávalos-Salas, M.; Furlan-Magaril, M.; González-Buendía, E.; Valdes-Quezada, C.; Ayala-Ortega, E.; Recillas-Targa, F. Gain of DNA Methylation Is Enhanced in the Absence of CTCF at the Human Retinoblastoma Gene Promoter. BMC Cancer 2011, 11, 232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luke, B.; Lingner, J. TERRA: Telomeric Repeat-Containing RNA. EMBO J. 2009, 28, 2503–2510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoeftner, S.; Blasco, M.A. Developmentally Regulated Transcription of Mammalian Telomeres by DNA-Dependent RNA Polymerase II. Nat. Cell Biol. 2008, 10, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Kidder, B.L. H4K20me3 Co-Localizes with Activating Histone Modifications at Transcriptionally Dynamic Regions in Embryonic Stem Cells. BMC Genom. 2018, 19, 514. [Google Scholar] [CrossRef] [Green Version]
- Kychygina, A.; Dall’Osto, M.; Allen, J.A.M.; Cadoret, J.-C.; Piras, V.; Pickett, H.A.; Crabbe, L. Progerin Impairs 3D Genome Organization and Induces Fragile Telomeres by Limiting the DNTP Pools. Sci Rep. 2021, 11, 13195. [Google Scholar] [CrossRef]
- Jost, D.; Vaillant, C. Epigenomics in 3D: Importance of Long-Range Spreading and Specific Interactions in Epigenomic Maintenance. Nucleic Acids Res. 2018, 46, 2252–2264. [Google Scholar] [CrossRef] [Green Version]
- Chuang, T.C.Y.; Moshir, S.; Garini, Y.; Chuang, A.Y.-C.; Young, I.T.; Vermolen, B.; van den Doel, R.; Mougey, V.; Perrin, M.; Braun, M.; et al. The Three-Dimensional Organization of Telomeres in the Nucleus of Mammalian Cells. BMC Biol. 2004, 2, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Fabian-Morales, E.; Vallejo-Escamilla, D.; Gudiño, A.; Rodríguez, A.; González-Barrios, R.; Torres, Y.L.R.; Hernández, C.C.; Torre-Luján, A.H.; Oliva-Rico, D.A.; Guzmán, E.C.O.; et al. Large-scale Topological Disruption of Chromosome Territories 9 and 22 Is Associated with Nonresponse to Treatment in CML. Int. J. Cancer 2021, 150, 1455–1470. [Google Scholar] [CrossRef]
- Zalensky, A.O.; Allen, M.J.; Kobayashi, A.; Zalenskaya, I.A.; Balhorn, R.; Bradbury, E.M. Well-Defined Genome Architecture in the Human Sperm Nucleus. Chromosoma 1995, 103, 577–590. [Google Scholar] [CrossRef] [PubMed]
- Lange, T. de Human Telomeres Are Attached to the Nuclear Matrix. EMBO J. 1992, 11, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Weierich, C.; Brero, A.; Stein, S.; von Hase, J.; Cremer, C.; Cremer, T.; Solovei, I. Three-Dimensional Arrangements of Centromeres and Telomeres in Nuclei of Human and Murine Lymphocytes. Chromosome Res. 2003, 11, 485–502. [Google Scholar] [CrossRef] [PubMed]
- Caria, P.; Dettori, T.; Frau, D.V.; Lichtenzstejn, D.; Pani, F.; Vanni, R.; Mai, S. Characterizing the Three-dimensional Organization of Telomeres in Papillary Thyroid Carcinoma Cells. J. Cell. Physiol. 2019, 234, 5175–5185. [Google Scholar] [CrossRef] [PubMed]
- Sunpaweravong, S.; Sunpaweravong, P.; Sathitruangsak, C.; Mai, S. Three-dimensional Telomere Architecture of Esophageal Squamous Cell Carcinoma: Comparison of Tumor and Normal Epithelial Cells. Dis. Esophagus 2016, 29, 307–313. [Google Scholar] [CrossRef]
- Klewes, L.; Vallente, R.; Dupas, E.; Brand, C.; Grün, D.; Guffei, A.; Sathitruangsak, C.; Awe, J.A.; Kuzyk, A.; Lichtensztejn, D.; et al. Three-Dimensional Nuclear Telomere Organization in Multiple Myeloma. Transl. Oncol. 2013, 6, 749-IN36. [Google Scholar] [CrossRef] [Green Version]
- Knecht, H.; Sawan, B.; Lichtensztejn, D.; Lemieux, B.; Wellinger, R.J.; Mai, S. The 3D Nuclear Organization of Telomeres Marks the Transition from Hodgkin to Reed–Sternberg Cells. Leukemia 2009, 23, 565–573. [Google Scholar] [CrossRef] [Green Version]
- Popova, L.V.; Nagarajan, P.; Lovejoy, C.M.; Sunkel, B.D.; Gardner, M.L.; Wang, M.; Freitas, M.A.; Stanton, B.Z.; Parthun, M.R. Epigenetic Regulation of Nuclear Lamina-Associated Heterochromatin by HAT1 and the Acetylation of Newly Synthesized Histones. Nucleic Acids Res. 2021, 49, 12136–12151. [Google Scholar] [CrossRef]
- The Human Genome Browser at UCSC. Available online: Genome.ucsc.edu (accessed on 5 January 2022).
- Kent, W.J.; Sugnet, C.W.; Furey, T.S.; Roskin, K.M.; Pringle, T.H.; Zahler, A.M.; Haussler, D. The Human Genome Browser at UCSC. Genome Res. 2002, 12, 996–1006. [Google Scholar] [CrossRef] [Green Version]
- Galiveti, C.R.; Rozhdestvensky, T.S.; Brosius, J.; Lehrach, H.; Konthur, Z. Application of Housekeeping NpcRNAs for Quantitative Expression Analysis of Human Transcriptome by Real-Time PCR. RNA 2010, 16, 450–461. [Google Scholar] [CrossRef] [Green Version]
- Whitaker, N.J.; Bryan, T.M.; Bonnefin, P.; Chang, A.C.; Musgrove, E.A.; Braithwaite, A.W.; Reddel, R.R. Involvement of RB-1, P53, P16INK4 and Telomerase in Immortalisation of Human Cells. Oncogene 1995, 11, 971–976. [Google Scholar] [PubMed]
- Hahn, W.C.; Counter, C.M.; Lundberg, A.S.; Beijersbergen, R.L.; Brooks, M.W.; Weinberg, R.A. Creation of Human Tumour Cells with Defined Genetic Elements. Nature 1999, 400, 464–468. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.-T.; Li, F.; Luan, R.-H.; Xing, J.-L.; Wang, R.-A.; Guo, W.-Y.; Wang, H. Telomerase Activity Is Not Enough for Tumor Initiation in Human Cells. Afr. J. Biotechnol. 2009, 8, 4734–4737. [Google Scholar]
- Galigniana, N.M.; Charó, N.L.; Uranga, R.; Cabanillas, A.M.; Piwien-Pilipuk, G. Oxidative Stress Induces Transcription of Telomeric Repeat-Containing RNA (TERRA) by Engaging PKA Signaling and Cytoskeleton Dynamics. Biochim. Biophys. Acta Bba. Mol. Cell. Res. 2020, 1867, 118643. [Google Scholar] [CrossRef] [PubMed]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliva-Rico, D.; Fabian-Morales, E.; Cáceres-Gutiérrez, R.E.; Gudiño, A.; Cisneros-Soberanis, F.; Dominguez, J.; Almaraz-Rojas, O.; Arriaga-Canon, C.; Castro-Hernández, C.; De la Rosa, C.; et al. Methylation of Subtelomeric Chromatin Modifies the Expression of the lncRNA TERRA, Disturbing Telomere Homeostasis. Int. J. Mol. Sci. 2022, 23, 3271. https://doi.org/10.3390/ijms23063271
Oliva-Rico D, Fabian-Morales E, Cáceres-Gutiérrez RE, Gudiño A, Cisneros-Soberanis F, Dominguez J, Almaraz-Rojas O, Arriaga-Canon C, Castro-Hernández C, De la Rosa C, et al. Methylation of Subtelomeric Chromatin Modifies the Expression of the lncRNA TERRA, Disturbing Telomere Homeostasis. International Journal of Molecular Sciences. 2022; 23(6):3271. https://doi.org/10.3390/ijms23063271
Chicago/Turabian StyleOliva-Rico, Diego, Eunice Fabian-Morales, Rodrigo E. Cáceres-Gutiérrez, Adriana Gudiño, Fernanda Cisneros-Soberanis, Julieta Dominguez, Oscar Almaraz-Rojas, Cristian Arriaga-Canon, Clementina Castro-Hernández, Carlos De la Rosa, and et al. 2022. "Methylation of Subtelomeric Chromatin Modifies the Expression of the lncRNA TERRA, Disturbing Telomere Homeostasis" International Journal of Molecular Sciences 23, no. 6: 3271. https://doi.org/10.3390/ijms23063271
APA StyleOliva-Rico, D., Fabian-Morales, E., Cáceres-Gutiérrez, R. E., Gudiño, A., Cisneros-Soberanis, F., Dominguez, J., Almaraz-Rojas, O., Arriaga-Canon, C., Castro-Hernández, C., De la Rosa, C., Reyes, J. L., & Herrera, L. A. (2022). Methylation of Subtelomeric Chromatin Modifies the Expression of the lncRNA TERRA, Disturbing Telomere Homeostasis. International Journal of Molecular Sciences, 23(6), 3271. https://doi.org/10.3390/ijms23063271