
Control of Bacterial Diseases of Banana Using CRISPR/Cas-Based Gene Editing


Abstract
:1. Introduction
2. Bacterial Diseases of Banana
3. Banana Xanthomonas Wilt (BXW)
4. Moko and Bugtok Disease
5. Blood Disease
6. Recent Advances in Gene Editing of Banana
7. Strategies for Developing Bacterial Wilt Resistant Banana

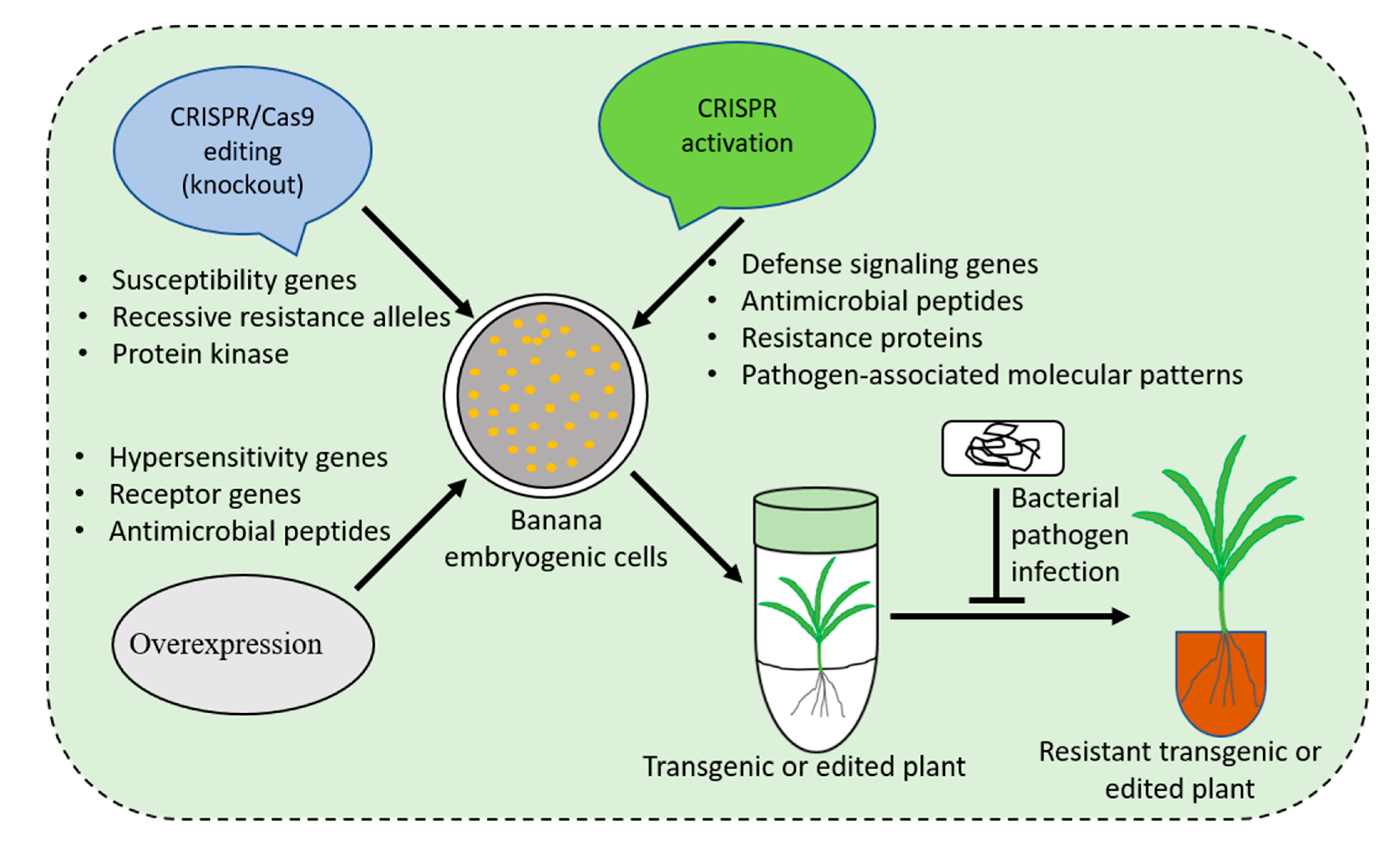{kind=link}
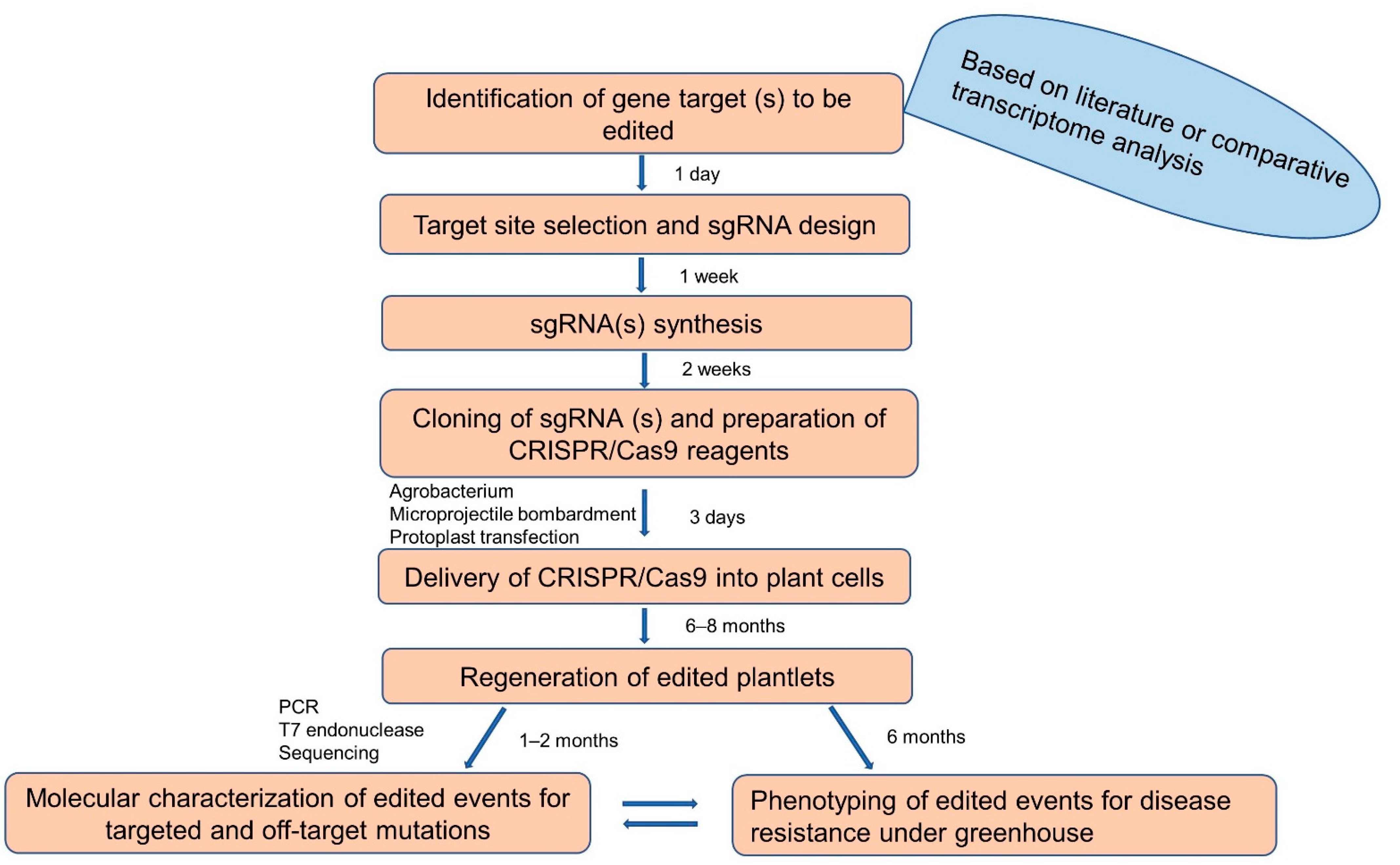{kind=link}
| Mode of Action | Potential Target Gene | Type of Manipulation | References |
|---|---|---|---|
| Hypersensitivity response | Hrap | Overexpression | [34] |
| Pflp | Overexpression | [34] | |
| Stacked Hrap and Pflp | Overexpression | [35] | |
| Pathogen recognition receptors induced immunity | Xa21 | Overexpression | [36] |
| Susceptibility genes | MusaDMR6 | Gene knockout | [11] |
| SlDMR6 | Gene knockout | [37] | |
| OsSWEET14 | Gene knockout | [38] | |
| OsSWEET11, OsSWEET13 OsSWEET14 promoter | Gene knockout | [39] | |
| DIPM1, DIPM2, DIPM4 | Gene knockout | [40] | |
| CsLOB1/CsLOB1promoter | Gene knockout | [41,42] | |
| MLO | Gene knockout | [43] | |
| AtAN9 | Gene knockout | [37] | |
| OsXa13 | Gene knockout | [44] | |
| OsCul3a | Gene knockout | [45] | |
| Protein kinases as a negative regulator of plant defense | OsMPK5 | Gene knockout | [46] |
| Nutrient transporter | ENOD | Gene knockout | [4] |
| E3 ubiquitin-protein ligase | PUB | Gene knockout | [4] |
| Pathogen-associated molecular patterns | LRR | Overexpression/CRISPR activation | [4] |
| Receptor kinases | WAK2, WAK5 | Overexpression/CRISPR activation | [4] |
| Antimicrobial peptides | Vicilin | Overexpression/CRISPR activation | [4] |
| Resistance proteins | RPM1 | Overexpression/CRISPR activation | [4] |
| Defense signaling | PR1 | Overexpression/CRISPR activation | [4] |
| NPR1 | Overexpression/CRISPR activation | [47] |
8. Knockout of Susceptibility Genes
9. Activation of Defense Genes through CRISPR Activation (CRISPRa)
10. Limitations in Gene Editing of Banana and Future Prospects
11. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAOSTAT. Banana Statistical Compendium 2020; Food and Agricultural Organization of the United Nations, FAO: Rome, Italy, 2021; Available online: www.fao.org (accessed on 31 January 2021).
- Ainembabazi, J.H.; Tripathi, L.; Rusike, J.; Abdoulaye, T.; Manyong, V. Ex-Ante Economic Impact Assessment of Genetically Modified Banana Resistant to Xanthomonas Wilt in the Great Lakes Region of Africa. PLoS ONE 2015, 10, e0138998. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, L.; Ntui, V.O.; Tripathi, J.N. CRISPR/Cas9 genome editing of banana for disease resistance. Curr. Opin. Plant Biol. 2020, 56, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, L.; Tripathi, J.N.; Shah, T.; Muiruri, S.K.; Katari, M. Molecular basis of disease resistance in banana progenitor Musa balbisiana against Xanthomonas campestris pv. musacearum. Sci. Rep. 2019, 9, 7007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, P.C.; Gerber, J.S.; Engstrom, P.M.; Mueller, N.D.; Brauman, K.A.; Carlson, K.M.; Cassidy, E.S.; Johnston, M.; MacDonald, G.K.; Ray, D.K.; et al. Leverage points for improving global food security and the environment. Science 2014, 345, 325–328. [Google Scholar] [CrossRef] [Green Version]
- Blomme, G.; Dita, M.; Jacobsen, K.S.; Pérez-Vicente, L.; Molina, A.; Ocimati, W.; Poussier, S.; Prior, P. Bacterial Diseases of Bananas and Enset: Current State of Knowledge and Integrated Approaches Toward Sustainable Management. Front. Plant Sci. 2017, 8, 1290. [Google Scholar] [CrossRef]
- Kagezi, G.H.; Kangire, A.; Tushmereirwe, W.; Bagamba, F.; Kikulwe, E.; Muhangi, J.; Gold, C.S.; Ragama, P.E. Banana bacterial wilt incidence in Uganda. Afr. Crop Sci. J. 2006, 14, 83–91. [Google Scholar] [CrossRef]
- Tripathi, L.; Mwangi, M.; Abele, S.; Aritua, V.; Tushemereirwe, W.K.; Bandyopadhyay, R. Xanthomonas Wilt: A Threat to Banana Production in East and Central Africa. Plant Dis. 2009, 93, 440–451. [Google Scholar] [CrossRef] [Green Version]
- Nakato, G.V.; Christelova, P.; Were, E.; Nyine, M.; Coutinho, T.A.; Dolezel, J.; Uwimana, B.; Swennen, R.; Mahuku, G. Sources of resistance in Musa to Xanthomonas campestris pv. Musacearum, the causal agent of banana xanthomonas wilt. Plant Pathol. 2019, 68, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Drenth, A.; Kema, G. The vulnerability of bananas to globally emerging disease threats. Phytopathology 2021, 111, 2146–2161. [Google Scholar] [CrossRef]
- Tripathi, J.N.; Ntui, V.O.; Shah, T.; Tripathi, L. CRISPR/Cas9-mediated editing of DMR6 orthologue in banana (Musa spp.) confers enhanced resistance to bacterial diseases. Plant Biotechnol. J. 2021, 19, 1291–1293. [Google Scholar] [CrossRef]
- Rorer, J.B. A bacterial disease of bananas and plantains. Phytopathology 1911, 1, 45–49. [Google Scholar]
- Buddenhagen, I.W. Banana diseases caused by bacteria. In Compendium of Tropical Fruit Diseases; Ploetz, R.C., Zentmyer, G.A., Nishijima, W.T., Rohrbach, K.G., Ohr, H.D., Eds.; APS Press: St. Paul, MN, USA, 1994; pp. 15–17. [Google Scholar]
- Belalcazar, S.C.; Rosales, F.E.; Pocasangre, L.E. El Moko del banano y el plátano y el rol de las plantas hospederas en su epidemiología. In Proceedings of the XVI International ACORBAT Meeting, Oaxaca, Mexico, 26 September–1 October 2004; Orozco-Santos, M., Orozco-Romero, J., Robles-Gonzalez, M., Velazquez-Monreal, J., Medina-Urrutia, V., Hernandez-Bautista, J.A., Eds.; Artturi: Oaxaca, Mexico, 2004; pp. 16–35. [Google Scholar]
- Molina, A.B. Managing bacterial wilt/fruit rot disease of banana in Southeast Asia. In Proceedings of the Banana Xanthomonas wilt Regional Preparedness and Strategy Development Workshop Held in Kampala: Developping A Regional Strategy to Address the Outbreak of Banana Xanthomonas wilt in East and Central Africa, Kampala, Uganda, 14–18 February 2005; Karamura, E., Osiru, M., Blomme, G., Lusty, C., Picq, C., Eds.; INIBAP: Montpellier, France, 2006; pp. 26–31. [Google Scholar]
- Thwaites, R.; Eden-Green, S.J.; Black, R. Diseases caused by bacteria. In Diseases of Banana, Abacá and Enset; Jones, D.R., Ed.; CAB International: Wallingford, UK, 2000; pp. 213–239. [Google Scholar]
- Ray, J.D.; Subbandiya, S.; Rincon-Florez, V.A.; Prakoso, A.B.; Mudita, W.I.; Carvalhais, L.C.; Markus, J.E.R.; O’Dwyer, C.A.; Drenth, A. Geographic expansion of banana blood disease in South Asia. Plant Dis. 2021, 105, 2792–2800. [Google Scholar] [CrossRef]
- Roesmiyanto, L.H.; Hutagalung, L. Blood disease (P. celebesis) on banana in Jeneponto–Sulawesi Selatan. Hortikultura 1989, 27, 39–41. [Google Scholar]
- Subijanto, M. Status of banana disease in Indonesia. In Proceedings of the Technical Meeting on Diseases Affecting Banana and Plantain in Asia and the Pacific: Banana Diseases in Asia and the Pacific, Brisbane, Australia, 15–18 April 1991; Valmayor, R.V., Umali, B.E., Bejosano, C.P., Eds.; INIBAP: Los Baños, CA, USA, 1991; pp. 44–49. [Google Scholar]
- Nurhadi, M.; dan Harlion, R. Serangan bakteri dan cendawan pada tanaman pisang di propinsi dati I lampung. Info Hortik. 1994, 2, 37–40. (In Indonesian) [Google Scholar]
- Tripathi, L.; Ntui, V.O.; Tripathi, J.N.; Kumar, P.L. Application of CRISPR/Cas for Diagnosis and Management of Viral Diseases of Banana. Front. iMicrobiol. 2021, 11, 609784. [Google Scholar] [CrossRef]
- Aman, R.; Ali, Z.; Butt, H.; Mahas, A.; Aljedaani, F.; Khan, M.Z.; Ding, S.; Mahfouz, M. RNA virus interference via CRISPR/Cas13a system in plants. Genome Biol. 2018, 19, 1. [Google Scholar] [CrossRef]
- Modrzejewski, D.; Hartung, F.; Sprink, T.; Krause, D.; Kohl, C.; Schiemann, J.; Wilhelm, R. What is the available evidence for the application of genome editing as a new tool for plant trait modification and the potential occurrence of associated of-target effects: A systematic map protocol. Environ. Evid. 2019, 8, 27. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.J.; Hussmann, J.A.; Yan, J.; Knipping, F.; Ravisankar, P.; Chen, P.-F.; Chen, C.; Nelson, J.W.; Newby, G.A.; Sahin, M.; et al. Enhanced prime editing systems by manipulating cellular determinants of editing outcomes. Cell 2021, 181, 5635–5652.e29. [Google Scholar] [CrossRef]
- Kaur, N.; Alok, A.; Shivani; Kaur, N.; Pandey, P.; Awasthi, P.; Tewari, S. CRISPR/Cas9- mediated efficient editing in phytoene desaturase (PDS) demonstrates precise manipulation in banana cv. Rasthali genome. Funct. Integr. Genom. 2018, 18, 89–99. [Google Scholar] [CrossRef]
- Naim, F.; Dugdale, B.; Kleidon, J.; Brinin, A.; Shand, K.; Waterhouse, P.; Dale, J. Gene editing the phytoene desaturase alleles of Cavendish banana using CRISPR/Cas9. Transgenic Res. 2018, 27, 451–460. [Google Scholar] [CrossRef] [Green Version]
- Ntui, V.O.; Tripathi, J.N.; Tripathi, L. Robust CRISPR/Cas9 mediated genome editing tool for banana and plantain (Musa spp.). Cur. Plant Biol. 2020, 21, 100128. [Google Scholar] [CrossRef]
- Zorrilla-Fontanesi, Y.; Pauwels, L.; Panis, B.; Signorelli, S.; Vanderschuren, H.; Swennen, R. Strategies to revise agrosystems and breeding to control fusarium wilt of banana. Nat. Food 2020, 1, 599–604. [Google Scholar] [CrossRef]
- Tripathi, J.N.; Ntui, V.O.; Ron, M.; Muiruri, S.K.; Britt, A.; Tripathi, L. CRISPR/Cas9 editing of endogenous banana streak virus in the B genome of Musa spp. overcomes a major challenge in banana breeding. Commun. Biol. 2019, 2, 46. [Google Scholar] [CrossRef] [Green Version]
- Kaur, N.; Alok, A.; Shivani; Kumar, P.; Kaur, N.; Awasthi, P.; Chaturvedi, S.; Pandey, P.; Ashutosh Pandey, A.; Pandey, A.K.; et al. CRISPR/Cas9 directed editing of lycopene epsilon-cyclase modulates metabolic flux for β-carotene biosynthesis in banana fruit. Metab. Eng. 2020, 59, 76–86. [Google Scholar] [CrossRef]
- Shao, X.; Wu, S.; Dou, T.; Zhu, C.; Hu, C.; Huo, H.; He, W.; Sheng, D.O.; Bi, F.; Gao, H.; et al. Using CRISPR/Cas9 genome editing system to create MaGA20ox2 gene-modified semi-dwarf banana. Plant Biotechnol. J. 2020, 18, 17019. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.; Sheng, O.; Deng, G.; He, W.; Dong, T.; Yang, Q.; Dou, T.; Li, C.; Gao, H.; Liu, S.; et al. CRISPR/Cas9-mediated Genome Editing of MaACO1 (aminocyclopropane-1-carboxylate oxidase1) Promotes the Shelf Life of Banana Fruit. Plant Biotechnol. J. 2021, 19, 654–656. [Google Scholar] [CrossRef]
- Kourelis, J.; van der Hoorn, R. Defended to the nines: 25 years of resistance gene cloning identifies nine mechanisms for R protein function. Plant Cell 2018, 30, 285–299. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, L.; Tripathi, J.; Kiggundu, A.; Korie, S.; Shotkoski, F.; Tushemereirwe, W.K. Field trial of Xanthomonas wilt disease-resistant bananas in East Africa. Nat. Biotechnol. 2014, 32, 868–870. [Google Scholar] [CrossRef]
- Muwonge, A.; Tripathi, J.N.; Kunert, K.; Tripathi, L. Expressing stacked HRAP and PFLP genes in transgenic banana has no synergistic effect on resistance to Xanthomonas wilt disease. S. Afr. J. Bot. 2016, 104, 125–133. [Google Scholar] [CrossRef]
- Tripathi, J.N.; Lorenzen, J.; Bahar, O.; Ronald, P.; Tripathi, L. Transgenic expression of the rice Xa21 pattern-recognition receptor in banana (Musa sp.) confers resistance to Xanthomonas campestris pv. Musacearum. Plant Biotehnol. J. 2014, 12, 663–673. [Google Scholar] [CrossRef] [Green Version]
- De Toledo Thomazella, D.P.; Seong, K.; Mackelprang, R.; Dahlbeck, D.; Geng, Y.; Gill, U.S.; Qi, T.; Pham, J.; Giuseppe, P.; Lee, C.Y.; et al. Loss of function of a DMR6 ortholog in tomato confers broad-spectrum disease resistance. Proc. Natl. Acad. Sci. USA 2021, 118, e2026152118. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Liu, B.; Spalding, M.H.; Weeks, D.P.; Yang, B. High-efficient TALEN-based gene editing produces disease-resistant rice. Nat. Biotechnol. 2012, 30, 390–392. [Google Scholar] [CrossRef] [PubMed]
- Oliva, R.; Ji, C.; Atienza-Grande, G.; Huguet-Tapia, J.C.; Perez-Quintero, S.; Li, T.; Eom, J.; Li, C.; Nguyen, H.; Liu, B.; et al. Broad-spectrum resistance to bacterial blight in rice using genome editing. Nat. Biotechnol. 2019, 37, 1344–1350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malnoy, M.; Viola, R.; Jung, M.-H.; Koo, O.-J.; Kim, S.; Kim, J.-S.; Velasco, R.; Kanchiswamy, C.N. DNA-Free Genetically Edited Grapevine and Apple Protoplast Using CRISPR/Cas9 Ribonucleoproteins. Front. Plant Sci. 2016, 7, 1904. [Google Scholar] [CrossRef]
- Peng, A.; Chen, S.; Lei, T.; Xu, L.; He, Y.; Wu, L.; Yao, L.; Zou, X. Engineering canker resistant plants through CRISPR/Cas9- targeted editing of the susceptibility gene CsLOB1 promoter in citrus. Plant Biotechnol. J. 2017, 10, 1011–1101. [Google Scholar] [CrossRef] [Green Version]
- Jia, H.; Orbovic, V.; Jones, J.F.; Wang, N. Modification of the PthA4 effector binding elements in Type I CsLOB1 promoter using Cas9/sgRNA to produce transgenic Duncan grapefruit alleviating XccDpthA4:dCsLOB1.3 infection. Plant Biotechnol. J. 2016, 14, 1291–1301. [Google Scholar] [CrossRef]
- Jiang, W.; Zhou, H.; Bi, H.; Fromm, M.; Yang, B.; Weeks, D.P. Demonstration of CRISPR/Cas9/sgRNA-mediated targeted gene modification in Arabidopsis, tobacco, sorghum and rice. Nucleic Acids Res. 2013, 41, e188. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Y.; Rosas-Diaz, T.; Caceres-Moreno, C.; Lozano-Dura’n, R.; Macho, A.P. The IMMUNE-ASSOCIATED NUCLEOTIDE-BINDING 9 protein is a regulator of basal immunity in Arabidopsis thaliana. Mol. Plant Microbe Interact. 2018, 32, 65–75. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.; Liu, Q.; Zhang, Y.; Chen, D.; Zhan, X.; Deng, C.; Cheng, S.; Cao, L. OsCUL3a-associated molecular switches have functions in cell metabolism, cell death, and disease resistance. J. Agric. Food Chem. 2020, 68, 5471–5482. [Google Scholar] [CrossRef]
- Xie, K.; Yang, Y. RNA-Guided Genome Editing in Plants Using a CRISPR–Cas System. Mol. Plant. 2013, 6, 1975–1983. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.; Yuan, M.; Ai, C.; Liu, L.; Zhuang, E.; Karapetyan, S. uORF-mediated translation allows engineered plant disease resistance without fitness costs. Nature 2017, 545, 491–494. [Google Scholar] [CrossRef]
- Ntui, V.O.; Uyoh, E.A.; Ita, E.E.; Markson, A.A.; Tripathi, J.N.; Okon, N.I.; Akpan, M.O.; Phillip, J.O.; Brisibe, E.A.; Ene-Obong, E.E.; et al. Strategies to combat the problems of yam anthracnose disease: Status and prospects. Mol. Plant Pathol. 2021, 22, 1302–1314. [Google Scholar] [CrossRef]
- Moniruzzaman, M.; Zhong, Y.; Yan, H.; Yuanda, L.; Jiang, B.; Zhong, G. Exploration of Susceptible Genes with Clustered Regularly Interspaced Short Palindromic Repeats–Tissue-Specific Knockout (CRISPR-TSKO) to Enhance Host Resistance. Crit. Rev. Plant Sci. 2020, 39, 387–417. [Google Scholar] [CrossRef]
- Kim, Y.-A.; Moon, H.; Par, C.-J. CRISPR/Cas9-targeted mutagenesis of Os8N3 in rice to confer resistance to Xanthomonas oryzae pv. oryzae. Rice 2019, 12, 67. [Google Scholar] [CrossRef]
- Schenke, D.; Cai, D. Applications of CRISPR/Cas to Improve Crop Disease Resistance: Beyond Inactivation of Susceptibility Factors. iScience 2020, 23, 101478. [Google Scholar] [CrossRef]
- Kieu, N.P.; Lenman, M.; Wang, E.S.; Peterson, B.L.; Andreasson, E. Mutations introduced in susceptibility genes through CRISPR/Cas9 genome editing confer increased late blight resistance in potatoes. Sci. Rep. 2021, 11, 4487. [Google Scholar] [CrossRef]
- Sun, K.; Wolters, A.M.; Vossen, J.H.; Rouwet, M.E.; Loonen, A.E.; Jacobsen, E.; Visser, R.G.; Bai, Y. Silencing of six susceptibility genes results in potato late blight resistance. Transgenic Res. 2016, 25, 731–742. [Google Scholar] [CrossRef] [Green Version]
- Engelhardt, S.; Stam, R.; Hückelhoven, R. Good riddance? Breaking disease susceptibility in the era of new breeding technologies. Agronomy 2018, 8, 114. [Google Scholar] [CrossRef] [Green Version]
- Dong, O.X.; Ronald, P.C. Genetic Engineering for Disease Resistance in Plants: Recent Progress and Future Perspectives. Plant Physiol. 2019, 180, 26–38. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Zhang, J.; Jia, H.; Sosso, D.; Li, T.; Frommer, W.B.; Wang, N. CsLOB1 is a disease susceptibility gene. Proc. Natl. Acad. Sci. USA 2014, 111, E521–E529. [Google Scholar] [CrossRef] [Green Version]
- Low, Y.C.; Lawton, M.A.; Di, R. Validation of barley 2OGO gene as a functional orthologue of Arabidopsis DMR6 gene in Fusarium head blight susceptibility. Sci. Rep. 2020, 10, 9935. [Google Scholar] [CrossRef]
- Zhang, Y.; Bai, Y.; Wu, G.; Zou, S.; Chen, Y.; Gao, C.; Tang, D. Simultaneous modification of three homoeologs of TaEDR1 by genome editing enhances powdery mildew resistance in wheat. Plant J. 2017, 91, 714–724. [Google Scholar] [CrossRef] [Green Version]
- Santillán Martínez, M.I.; Bracuto, V.; Koseoglou, E.; Appiano, M.; Jacobsen, E.; Visser, R.G.F.; Walters, A.M.; Bai, Y. CRISPR/Cas9-targeted mutagenesis of the tomato susceptibility gene PMR4 for resistance against powdery mildew. BMC Plant Biol. 2020, 20, 284. [Google Scholar] [CrossRef]
- Yin, K.; Qiu, J.L. Genome editing for plant disease resistance: Applications and perspectives. Philos. Trans. R. Soc. Biol. Sci. 2019, 374, 20180322. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Qi, L.S. Repurposing CRISPR System for Transcriptional Activation. Adv. Exp. Med. Biol. 2017, 983, 147–157. [Google Scholar] [CrossRef]
- Di-Maria, V.; Moindrot, M.; Ryde, M.; Bono, A.; Quintino, L.; Ledri, M. Development and Validation of CRISPR Activator Systems for Overexpression of CB1 Receptors in Neurons. Front. Mol. Neurosci. 2020, 13, 168. [Google Scholar] [CrossRef]
- Shakirova, K.M.; Ovchinnikova, V.Y.; Dashinimaev, E.B. Cell Reprogramming with CRISPR/Cas9 based transcriptional regulation systems. Front. Bioeng. Biotechnol. 2020, 8, 882. [Google Scholar] [CrossRef]
- Gilbert, L.A.; Larson, M.H.; Morsut, L.; Liu, Z.; Brar, G.A.; Torres, S.E.; Stern-Ginossar, N.; Brandman, O.; Whitehead, E.H.; Doudna, J.A.; et al. CRISPR-mediated modular RNA-guided regulation of transcription in eukaryotes. Cell 2013, 154, 442–451. [Google Scholar] [CrossRef] [Green Version]
- Hilton, I.B.; D’Ippolito, A.M.; Vockley, C.M.; Thakore, P.I.; Crawford, G.E.; Reddy, T.E.; Gersbach, C.A. Epigenome editing by a CRISPR-Cas9-based acetyltransferase activates genes from promoters and enhancers. Nat. Biotechnol. 2015, 33, 510–517. [Google Scholar] [CrossRef] [Green Version]
- Cheng, A.W.; Wang, H.; Yang, H.; Shi, L.; Katz, Y.; Theunissen, T.W.; Rangarajan, S.; Shivalila, C.S.; Dadon, D.B.; Jaenisch, R. Multiplexed activation of endogenous genes by CRISPR-on, an RNA-guided transcriptional activator system. Cell Res. 2013, 23, 1163–1171. [Google Scholar] [CrossRef]
- Duellman, T.; Doll, A.; Chen, X.; Wakamiya, R.; Yang, J. dCas9-mediated transcriptional activation of tissue inhibitor of metalloproteinases. Met. Med. 2017, 4, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhang, D.; Xiong, X.; Yan, B.; Xie, W.; Sheen, J.; Li, J.F. A potent Cas9-derived gene activator for plant and mammalian cells. Nat. Plants 2017, 3, 930–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haenfler, J.M.; Skariah, G.; Rodriguez, C.M.; Monteiro da Rocha, A.; Parent, J.M.; Smith, G.D.; Todd, P.K. Targeted reactivation of FMR1 transcription in fragile X syndrome embryonic stem cells. Front. Mol. Neurosci. 2018, 11, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowder, L.G.; Zhang, Y.; Baltes, N.J.; Paul, J.W.; Tang, X.; Zheng, X.; Jaenisch, R.; Voytas, D.F.; Hsieh, T.; Zhang, Y.; et al. A CRISPR/Cas9 toolbox for multiplexed plant genome editing and transcriptional regulation. Plant Physiol. 2015, 169, 971–985. [Google Scholar] [CrossRef] [Green Version]
- Lowder, L.G.; Malzahn, A.; Qi, Y. Plant Gene Regulation Using Multiplex CRISPR-dCas9 Artificial Transcription Factors. In Maize. Methods in Molecular Biology; Lagrimini, L., Ed.; Humana Press: New York, NY, USA, 2018; Volume 1676. [Google Scholar] [CrossRef]
- Selma, S.; Bernabé-Orts, J.M.; Vazquez-Vilar, M.; Diego-Martin, B.; Ajenjo, M.; Garcia-Carpintero, V.; Granell, A.; Orzaez, D. Cas9-based programmable transcriptional activator. Plant Biotechnol. J. 2019, 17, 1703–1705. [Google Scholar] [CrossRef] [Green Version]
- Pan, C.; Wu, X.; Markel, K.; Malzahn, A.A.; Kundagrami, N.; Sretenovic, S.; Zhang, Y.; Cheng, Y.; Shih, P.M.; Yiping Qi, Y. CRISPR–Act3.0 for highly efficient multiplexed gene activation in plants. Nat. Plants 2021, 7, 942–953. [Google Scholar] [CrossRef]
- Ganapathi, T.R.; Higgs, N.S.; Balint-Kurti, P.J.; Arntzen, C.J.; May, G.D.; Van Eck, J.M. Agrobacterium-mediated transformation of the embryogenic cell suspensions of the banana cultivars Rasthali (AAB). Plant Cell Rep. 2001, 20, 157–162. [Google Scholar] [CrossRef]
- Khanna, H.; Becker, D.; Kleidon, J.; Dale, J. Centrifugation assisted Agrobacterium tumefaciens mediated transformation (CAA) of embryogenic cell suspensions of banana (Musa spp.) Cavendish AAA and Lady finger AAB. Mol. Breed. 2004, 14, 239–252. [Google Scholar] [CrossRef]
- Tripathi, J.N.; Muwonge, A.; Tripathi, L. Efficient regeneration and transformation protocol for plantain cv. ‘Gonja Manjaya’ (Musa spp. AAB) using embryogenic cell suspension. Vitr. Cell. Dev. Biol. Plant 2012, 48, 216–224. [Google Scholar] [CrossRef]
- Tripathi, J.N.; Oduor, R.O.; Tripathi, L. A high-throughput regeneration and transformation platform for production of genetically modified banana. Front. Plant Sci. 2015, 6, 1025. [Google Scholar] [CrossRef] [Green Version]
- Gordon-Kamm, B.; Sardesai, N.; Arling, M.; Lowe, K.; Hoerster, G.; Betts, S.; Jones, T. Using Morphogenic Genes to Improve Recovery and Regeneration of Transgenic Plants. Plants 2019, 8, 38. [Google Scholar] [CrossRef] [Green Version]
- Masters, A.; Kang, M.; McCaw, M.; Zobrist, J.D.; Gordon-Kamm, W.; Jones, T.; Wang, K. Agrobacterium-mediated immature embryo transformation of recalcitrant maize inbred lines using morphogenic genes. J. Vis. Exp. 2020, 156, e60782. [Google Scholar] [CrossRef] [Green Version]
- Lowe, K.; Wu, E.; Wang, N.; Hoerster, G.; Hastings, C.; Cho, M.J.; Scelonge, C.; Lenderts, B.; Chamberlin, M.; Cushatt, J.; et al. Morphogenic Regulators Baby boom and Wuschel Improve Monocot Transformation. Plant Cell 2016, 28, 1998–2015. [Google Scholar] [CrossRef] [Green Version]
- Jones, T.; Lowe, K.; Hoerster, G.; Anand, A.; Wu, E.; Wang, N.; Arling, M.; Lenderts, B.; Gordon-Kamm, W. Maize Transformation Using the Morphogenic Genes Baby Boom and Wuschel2. In Transgenic Plants; Kumar, S., Barone, P., Smith, M., Eds.; Humana Press: New York, NY, USA, 2019; p. 1864. [Google Scholar] [CrossRef]
- Debernardi, J.M.; Tricoli, D.M.; Ercoli, M.F.; Hayta, S.; Ronald, P.; Palatnik, J.F.; Dubcovsky, J. A GRF-GIF chimeric protein improves the regeneration efficiency of transgenic plants. Nat. Biotechnol. 2020, 38, 1274–1279. [Google Scholar] [CrossRef]
- Woo, J.W.; Kim, J.; Kwon, S.I.; Corvalan, C.; Cho, S.W.; Kim, H.; Kim, S.-G.; Kim, S.-T.; Choe, S.; Kim, J.-S. DNA-free genome editing in plants with preassembled CRISPR-Cas9 ribonucleoproteins. Nat. Biotechnol. 2015, 33, 1162–1164. [Google Scholar] [CrossRef]
- Svitashev, S.; Schwartz, C.; Lenderts, B.; Young, J.K.; Cigan, M.A. Genome editing in maize directed by CRISPR-Cas9 ribonucleoprotein complexes. Nat. Commun. 2016, 7, 13274. [Google Scholar] [CrossRef]
- Liang, Z.; Chen, K.L.; Li, T.; Zhang, Y.; Wang, Y.P.; Zhao, Q.; Liu, J.; Zhang, H.; Liu, C.; Ran, Y.; et al. Efficient DNA-free genome editing of bread wheat using CRISPR/Cas9 ribonucleoprotein complexes. Nat. Commun. 2017, 8, 14261. [Google Scholar] [CrossRef]
- Kanchiswamy, C.N.; Malnoy, M.; Velasco, R.; Kim, J.-S.; Viola, R. Non-GMO genetically edited crop plants. Trends Biotechnol. 2015, 33, 489–491. [Google Scholar] [CrossRef]
- Panis, B.; Wauwe, A.V.; Swennen, R. Plant regeneration through direct somatic embryogenesis from protoplasts of banana (Musa spp.). Plant Cell Rep. 1993, 12, 403–407. [Google Scholar] [CrossRef]
- Matsumoto, K.; Oks, S. Plant regeneration from protoplasts of a Brazilian dessert banana (Musa spp. AAB group). Acta Hortic. 1998, 490, 455–462. [Google Scholar] [CrossRef]
- Assani, A.; Haicour, R.; Wenzel, G.; Cote, F.; Bakry, F.; Foroughi-Wehr, B.; Ducreux, G.; Aguillar, M.-E.; Grapin, A. Plant regeneration from protoplasts of dessert banana cv. Grande Naine (Musa spp., Cavendish sub-group AAA) via somatic embryogenesis. Plant Cell Rep. 2001, 20, 482–488. [Google Scholar] [CrossRef]
- Chen, L.; Li, W.; Katin-Grazzini, L.; Ding, J. A method for the production and expedient screening of CRISPR/Cas9- mediated non-transgenic mutant plants. Hortic. Res. 2018, 5, 13. [Google Scholar] [CrossRef]
- Veillet, F.; Perrot, L.; Chauvin, L.; Kermarrec, M.-P.; Guyon-Debast, A.; Chauvin, J.-E.; Nogue, F.; Mazier, M. Transgene-Free Genome Editing in Tomato and Potato Plants Using Agrobacterium-Mediated Delivery of a CRISPR/Cas9 Cytidine Base Editor. Int. J. Mol. Sci. 2019, 20, 402. [Google Scholar] [CrossRef] [Green Version]
- Costa, L.D.; Piazza, S.; Pompili, V.; Salvagnin, U.; Cestaro, A.; Moffa, L.; Vittani, L.; Moser, C.; Malnoy, M. Strategies to produce T-DNA free CRISPRed fruit trees via Agrobacterium tumefaciens stable gene transfer. Sci. Rep. 2020, 10, 20155. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tripathi, L.; Ntui, V.O.; Tripathi, J.N. Control of Bacterial Diseases of Banana Using CRISPR/Cas-Based Gene Editing. Int. J. Mol. Sci. 2022, 23, 3619. https://doi.org/10.3390/ijms23073619
Tripathi L, Ntui VO, Tripathi JN. Control of Bacterial Diseases of Banana Using CRISPR/Cas-Based Gene Editing. International Journal of Molecular Sciences. 2022; 23(7):3619. https://doi.org/10.3390/ijms23073619
Chicago/Turabian StyleTripathi, Leena, Valentine O. Ntui, and Jaindra N. Tripathi. 2022. "Control of Bacterial Diseases of Banana Using CRISPR/Cas-Based Gene Editing" International Journal of Molecular Sciences 23, no. 7: 3619. https://doi.org/10.3390/ijms23073619
APA StyleTripathi, L., Ntui, V. O., & Tripathi, J. N. (2022). Control of Bacterial Diseases of Banana Using CRISPR/Cas-Based Gene Editing. International Journal of Molecular Sciences, 23(7), 3619. https://doi.org/10.3390/ijms23073619