
Context Dependent Sulf1/Sulf2 Functional Divergence in Endothelial Cell Activity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
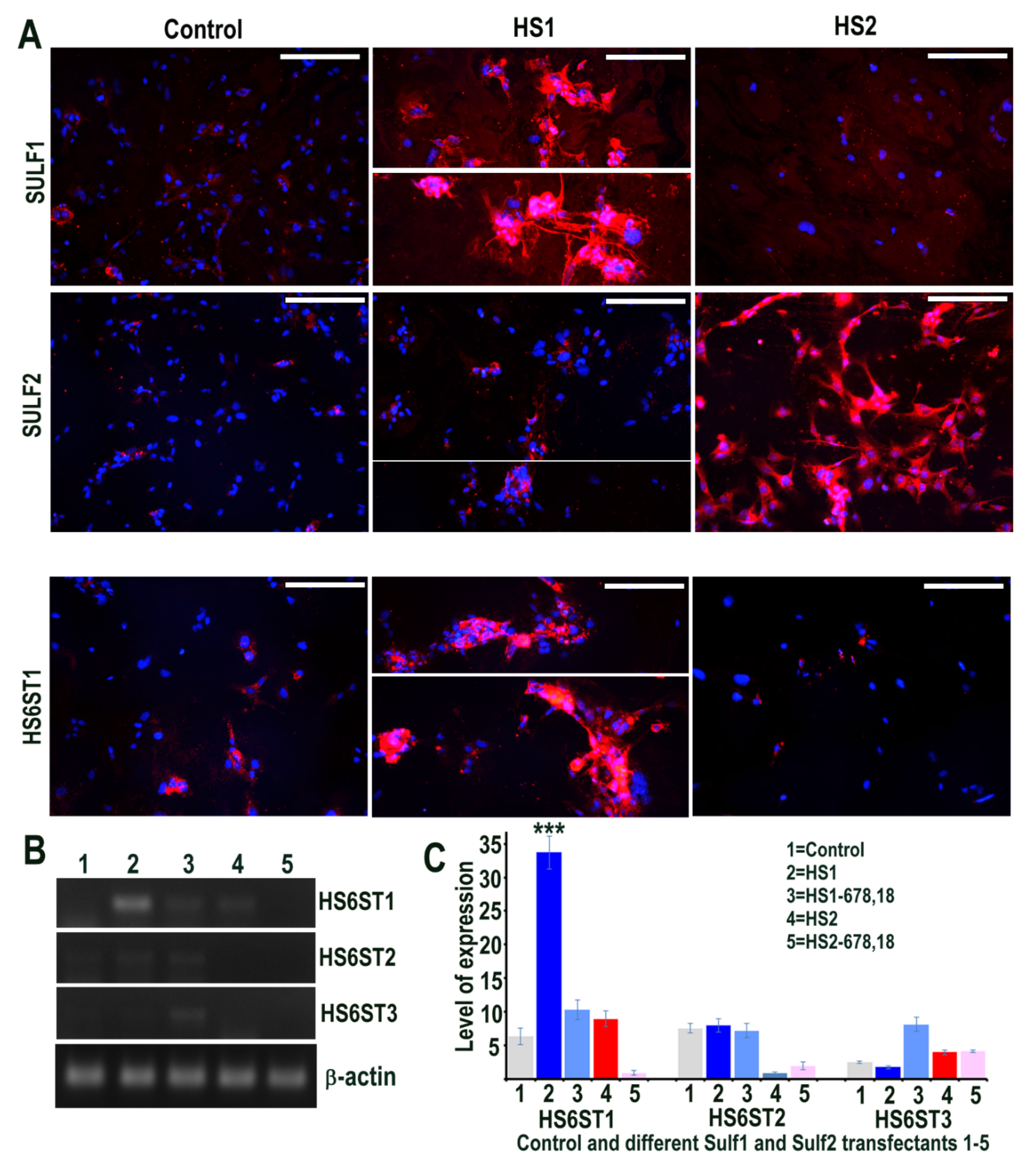
2.1. Sulf Over-Expression in HMec1 Cells Defines Distinct Sulf1 and Sulf2 Regulatory Roles
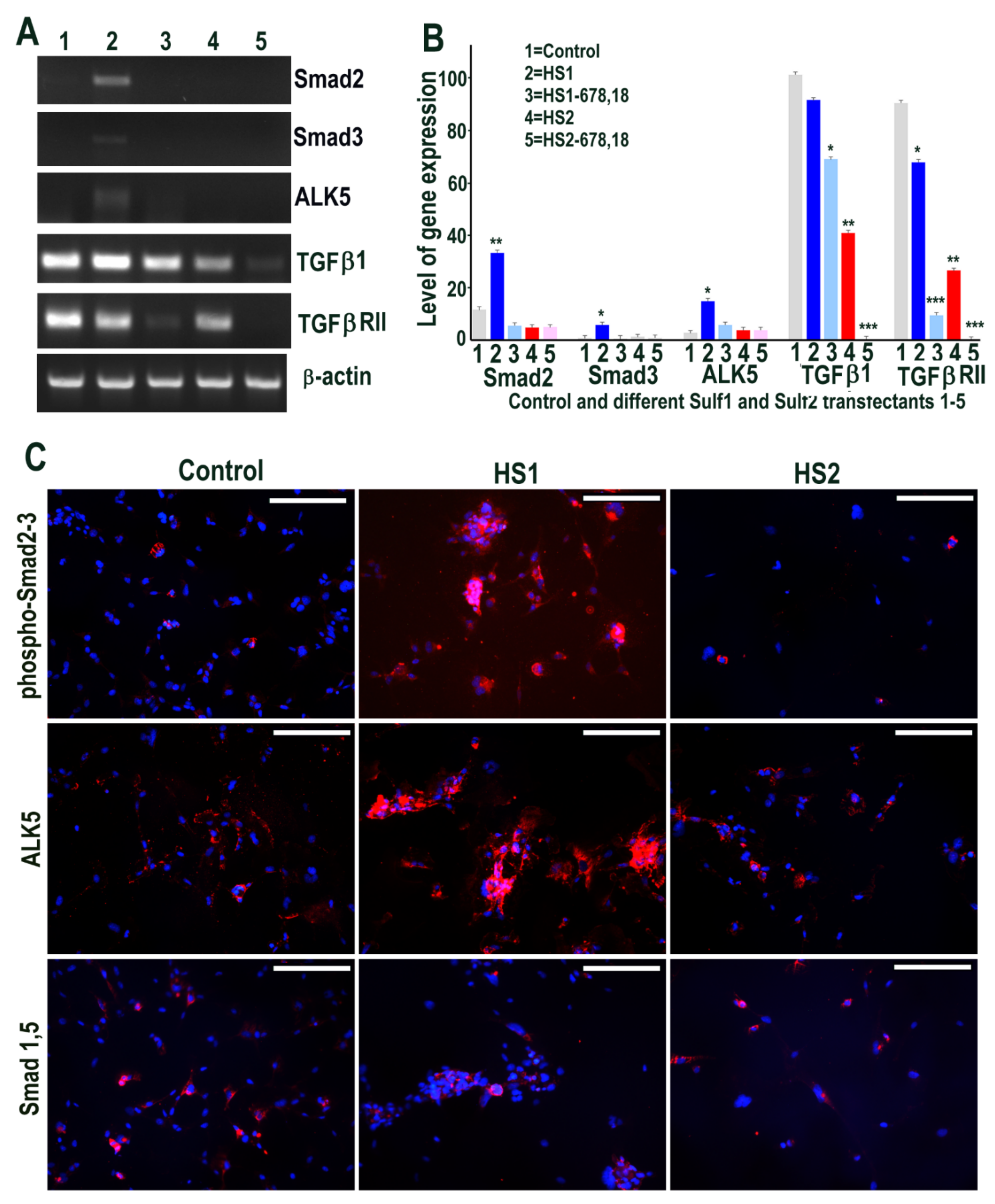
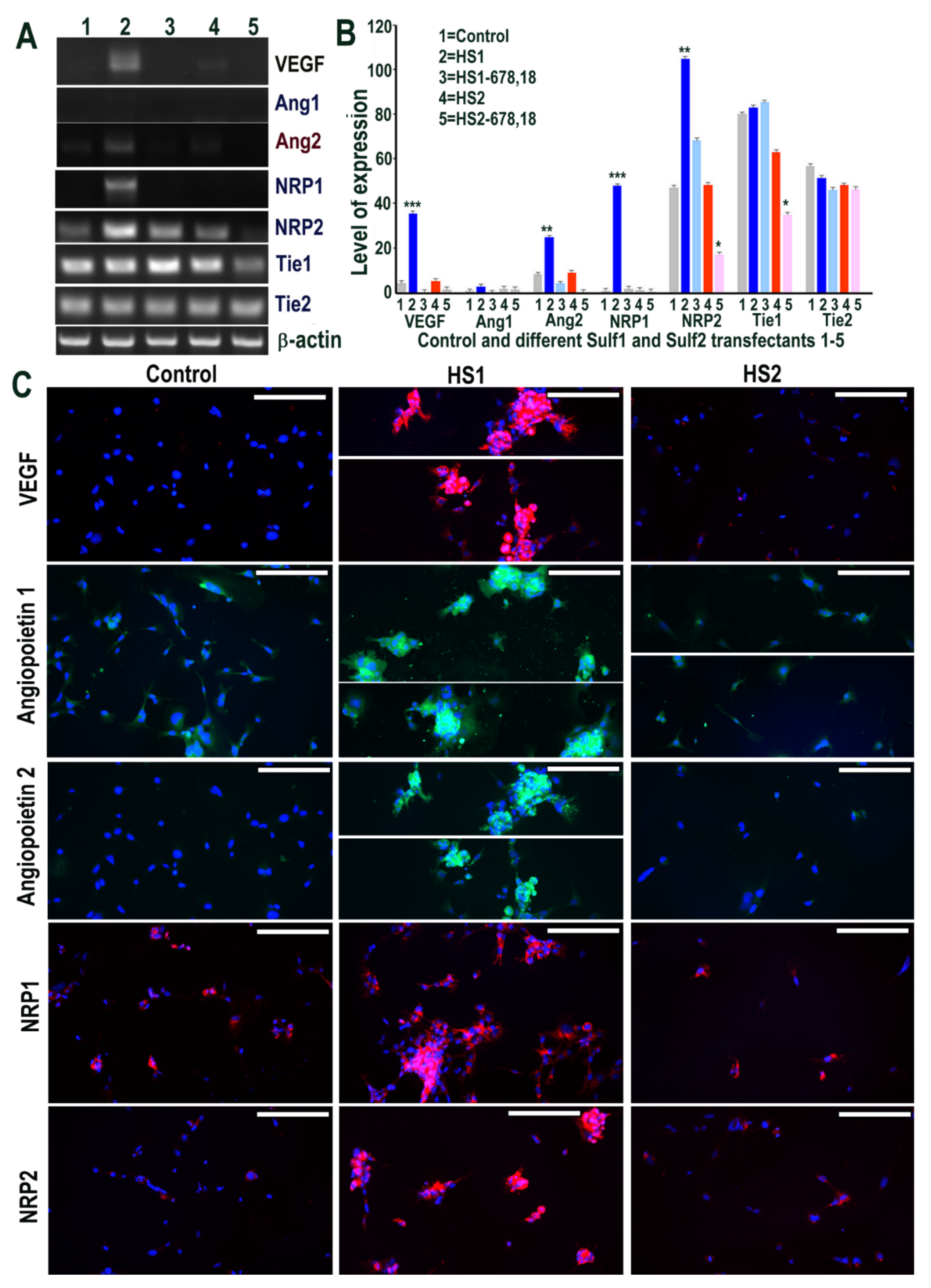
2.2. Over-Expression of Sulf1, but Not Sulf2, in HMec1 Cells Augments TGFβ and VEGF-A Cell Signalling
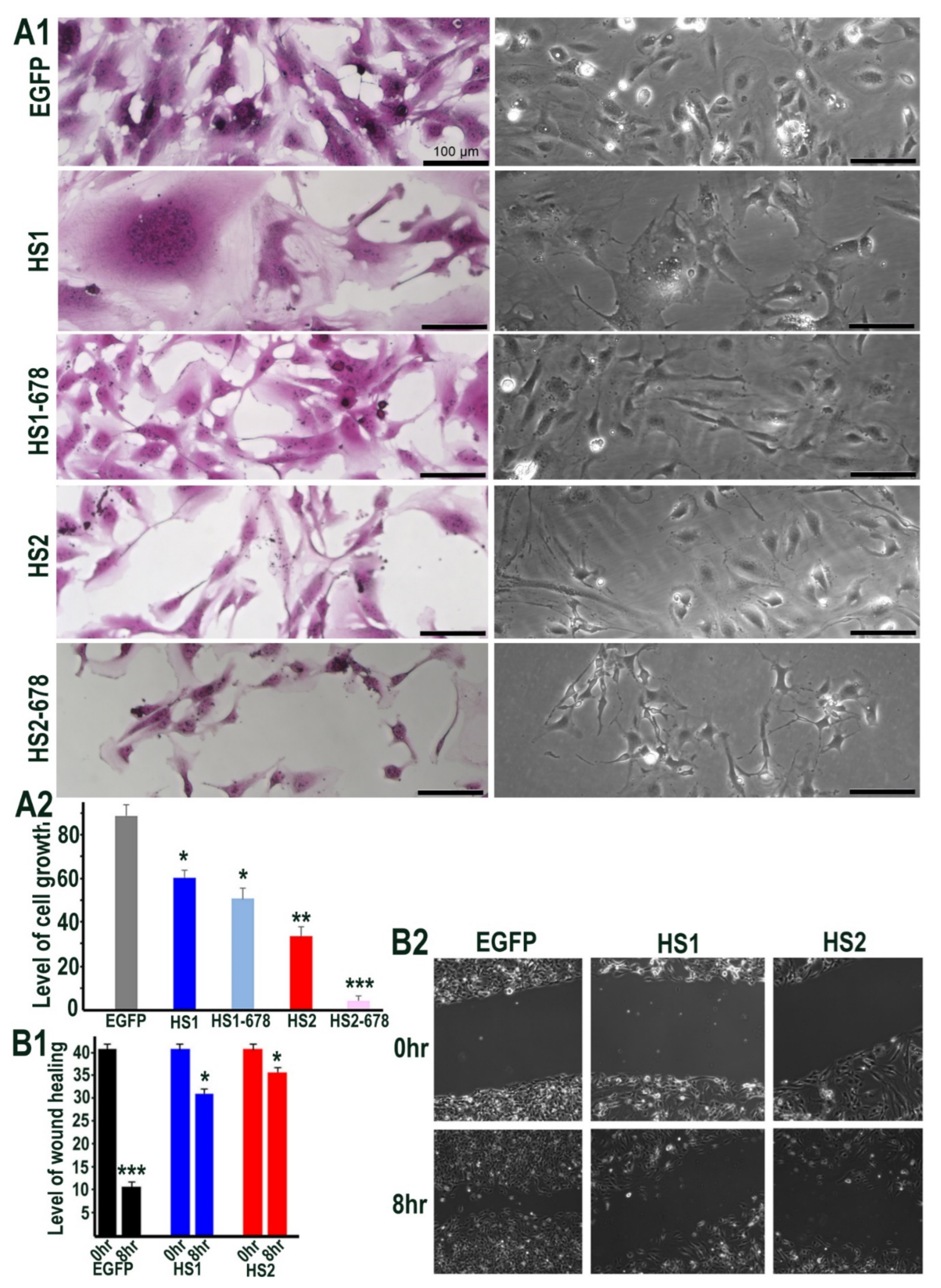
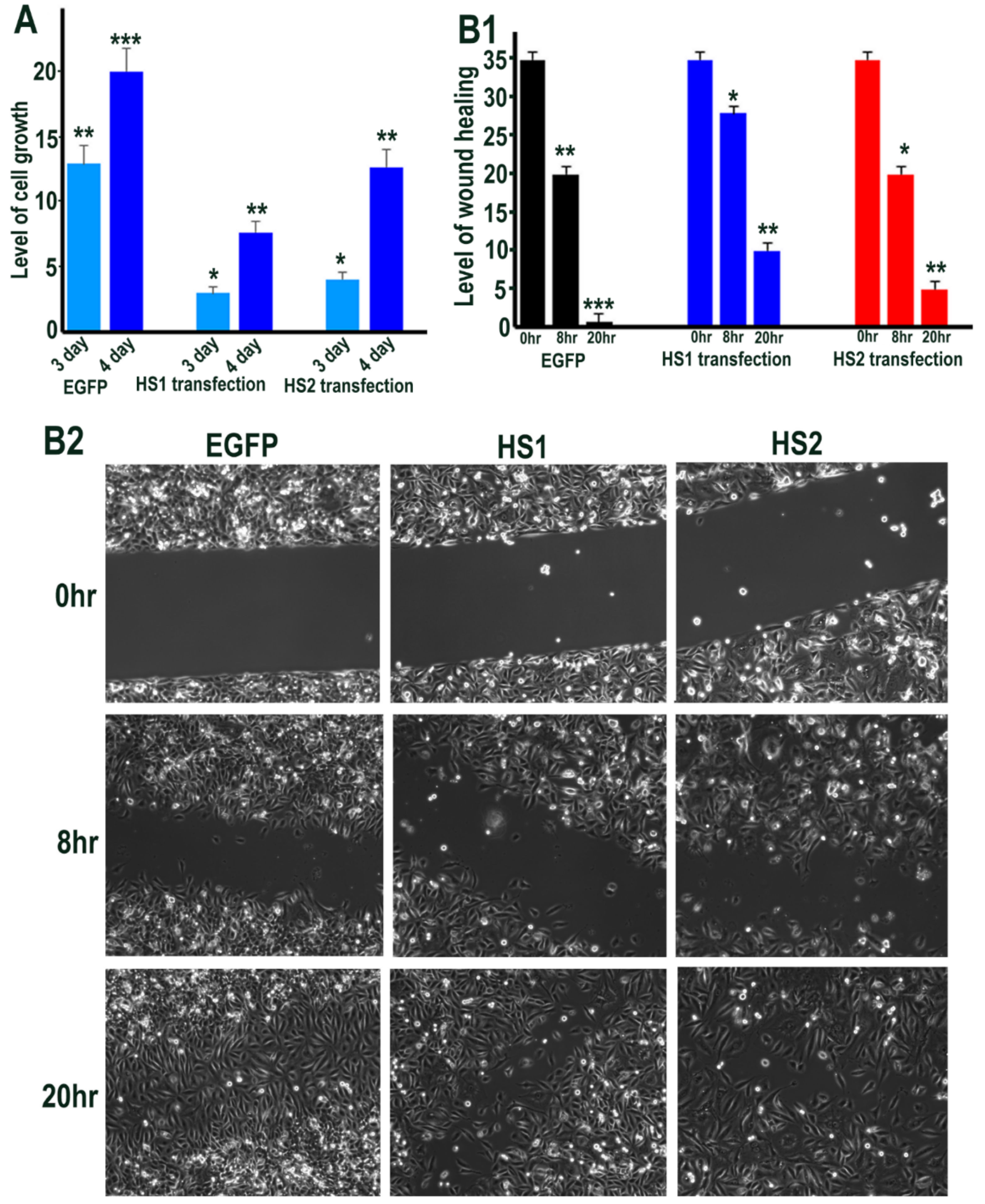
2.3. Sulf1 and Sulf2 Over-Expression Leads to Diminished Proliferation and Cell Migration in HMec1 Cells
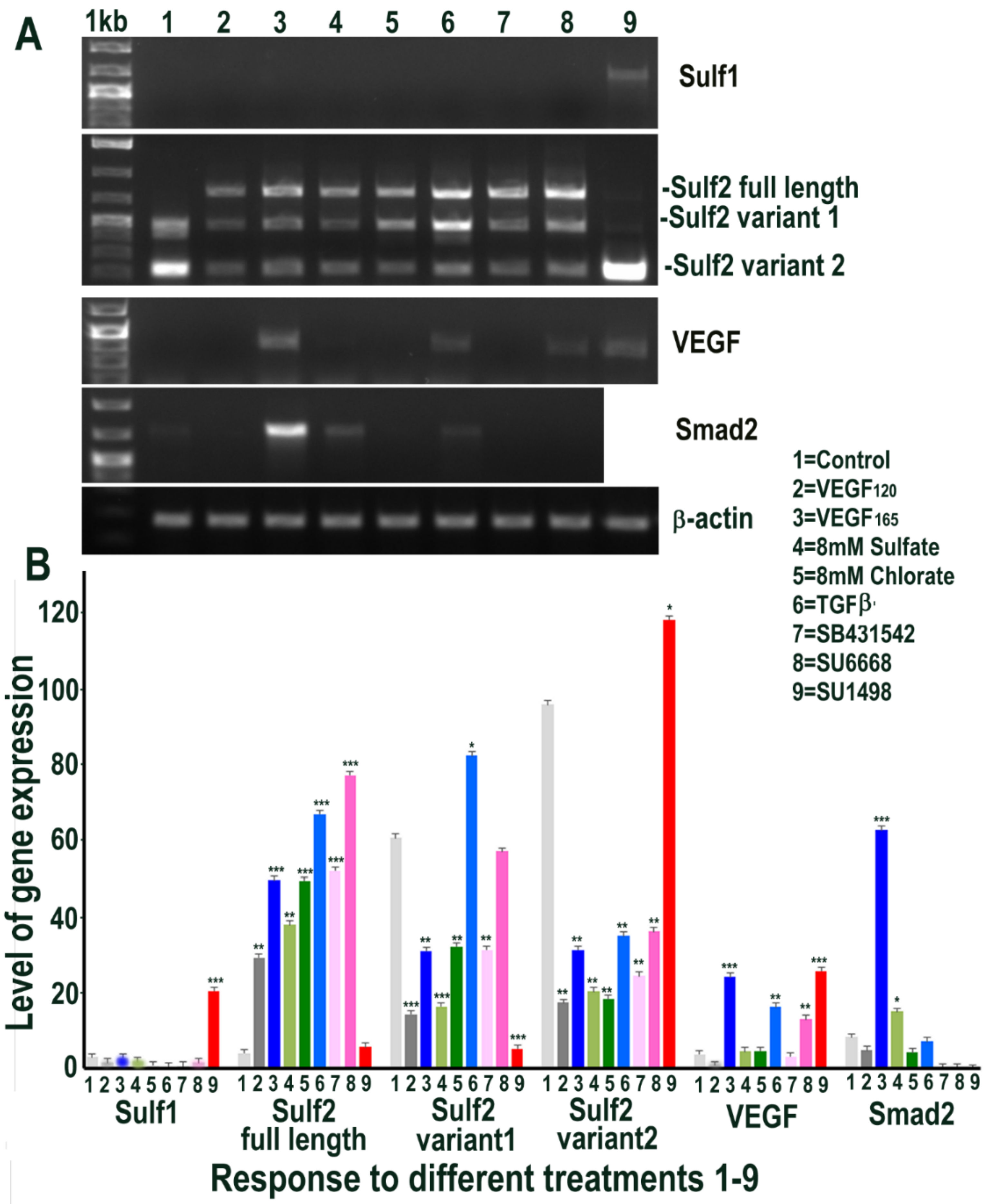
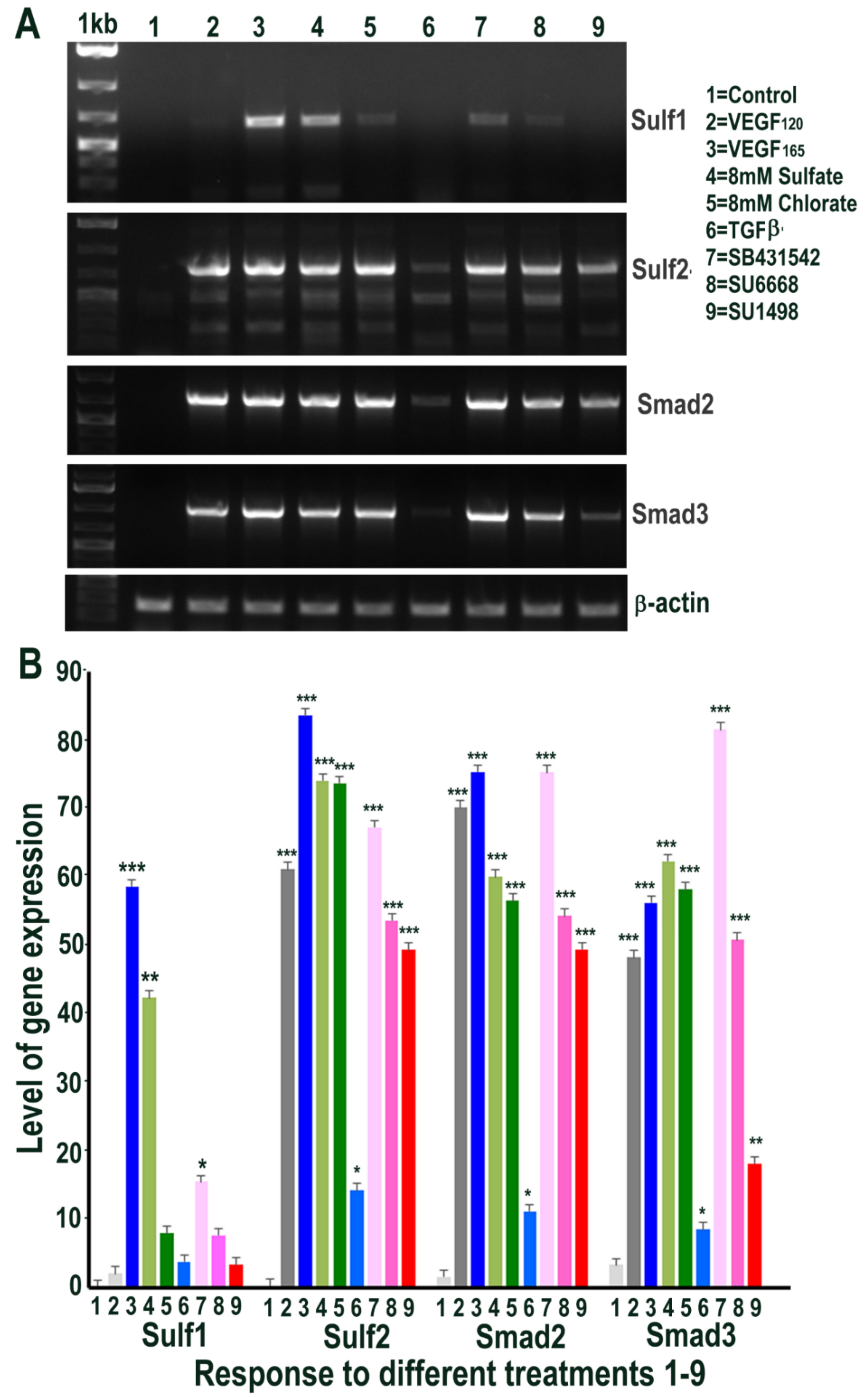
2.4. HMec1 Cell Responses to Increased VEGF, TGFβ or Their Inhibitors Highlights Their Interdependent Cell Signalling and Sulf1, Sulf2 Regulation
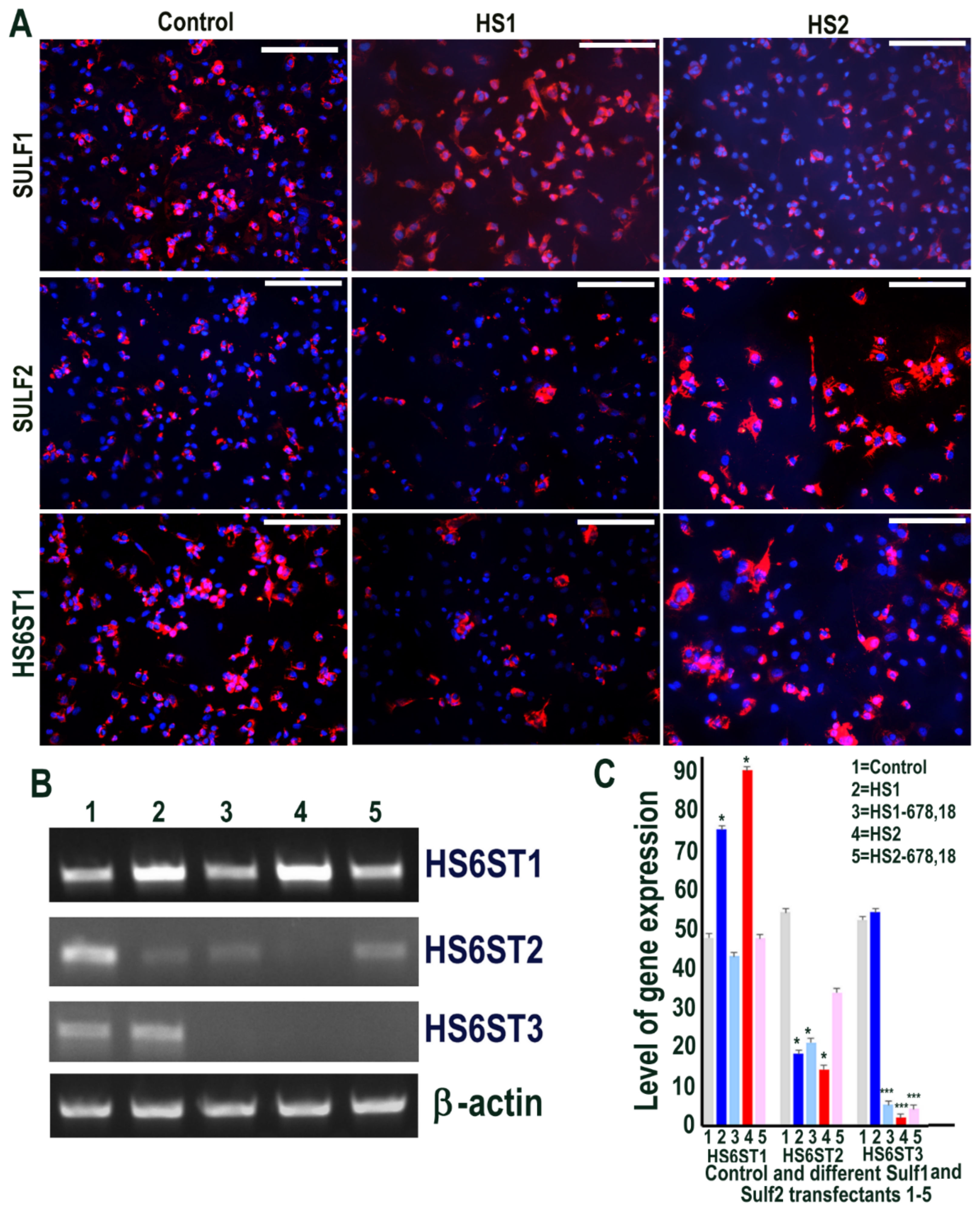
2.5. Sulf1/Sulf2 Over-Expression in ea926 Cells and Their Effect on HS 6-O Sulfotransferases 1,2,3
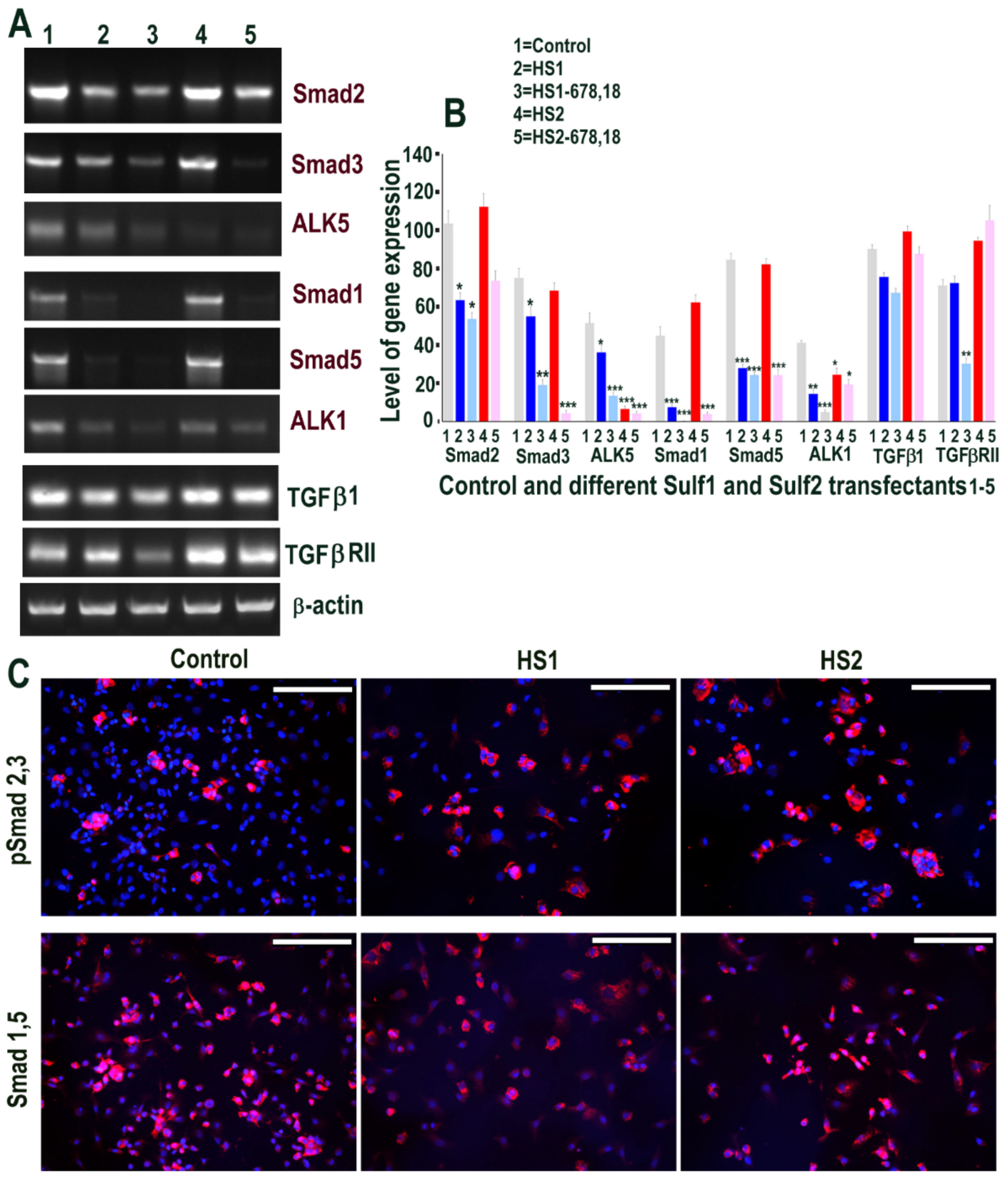
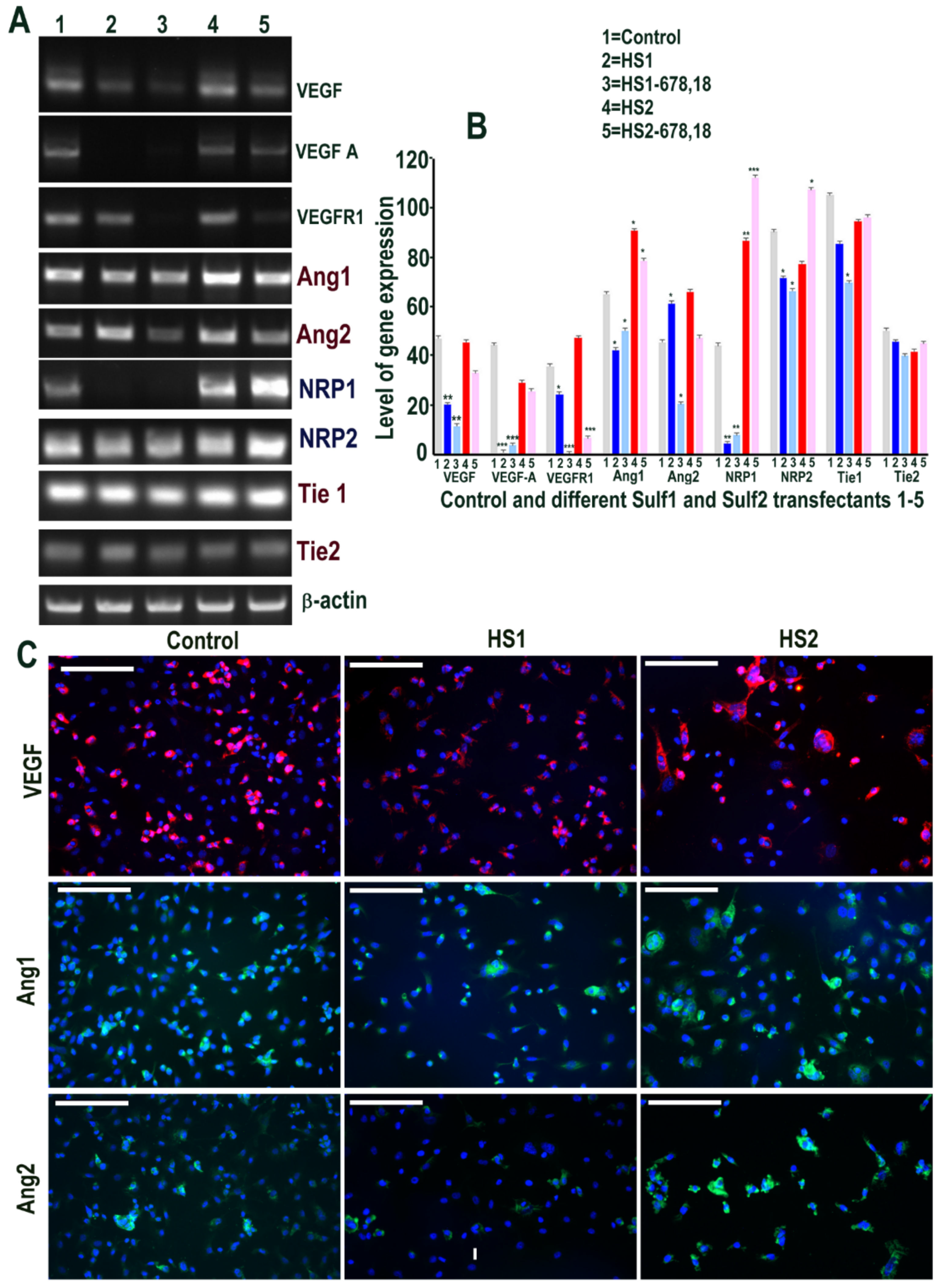
2.6. Sulf1 Over-Expression, but Not Sulf2, Reduces TGFβ and VEGF Cell Signalling in ea926 Cells
2.7. The Effect of Sulf1 or Sulf2 Over-Expression in ea926 Cells on Growth and Cell Migration
2.8. Exogenous VEGF and TGFβ Exposure also Regulates Sulf1 and Sulf2 Expression in ea926 Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Sulf1/Sulf2 Transfection
4.2. RT PCR Analysis
4.3. Immunocytochemistry
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carmeliet, P.J.R. Molecular mechanisms and clinical applications of angiogenesis. Nature 2011, 473, 298–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasut, A.B.L.; Cuypers, A.; Carmeliet, P. Endothelial cell plasticity at the single-cell level. Angiogenesis 2021, 24, 311–326. [Google Scholar] [CrossRef] [PubMed]
- El Masri, R.S.A.; Lortat-Jacob, H.; Vivès, R.R. The “in and out” of glucosamine 6-O-sulfation: The 6th sense of heparan sulfate. Glycoconj. J. 2017, 34, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Dhoot, G.K. Recent Progress and Related Patents on the Applications of SULF1/SULF2 Enzymes in Regenerative Medicine and Cancer Therapies. Recent Pat. Regen. Med. 2012, 2, 137–145. [Google Scholar] [CrossRef]
- Xie, M.L.J. Heparan sulfate proteoglycan—A common receptor for diverse cytokines. Cell Signal. 2019, 54, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Narita, K.; Staub, J.; Chien, J.; Meyer, K.; Bauer, M.; Friedl, A.; Ramakrishnan, S.; Shridhar, V. HSulf-1 inhibits angiogenesis and tumorigenesis in vivo. Cancer Res. 2006, 66, 6025–6032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosen, S.D.; Lemjabbar-Alaoui, H. Sulf-2: An extracellular modulator of cell signaling and a cancer target candidate. Expert Opin. Ther. Targets 2010, 14, 935–949. [Google Scholar] [CrossRef] [Green Version]
- Dhoot, G.K.; Gustafsson, M.K.; Ai, X.; Sun, W.; Standiford, D.M.; Emerson, C.P., Jr. Regulation of Wnt signaling and embryo patterning by an extracellular sulfatase. Science 2001, 293, 1663–1666. [Google Scholar] [CrossRef]
- Morimoto-Tomita, M.; Uchimura, K.; Werb, Z.; Hemmerich, S.; Rosen, S.D. Cloning and characterization of two extracellular heparin-degrading endosulfatases in mice and humans. J. Biol. Chem. 2002, 277, 49175–49185. [Google Scholar] [CrossRef] [Green Version]
- Ai, X.; Do, A.T.; Lozynska, O.; Kusche-Gullberg, M.; Lindahl, U.; Emerson, C.P., Jr. QSulf1 remodels the 6-O sulfation states of cell surface heparan sulfate proteoglycans to promote Wnt signaling. J. Cell Biol. 2003, 162, 341–351. [Google Scholar] [CrossRef] [Green Version]
- Nawroth, R.; van Zante, A.; Cervantes, S.; McManus, M.; Hebrok, M.; Rosen, S.D. Extracellular sulfatases, elements of the Wnt signaling pathway, positively regulate growth and tumorigenicity of human pancreatic cancer cells. PLoS ONE 2007, 2, e392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhanasekaran, R.N.I.; Hu, C.; Chen, G.; Oseini, A.M.; Seven, E.S.; Miamen, A.G.; Moser, C.D.; Zhou, W.; van Kuppevelt, T.H.; van Deursen, J.; et al. Activation of the TGFβ/SMAD Transcriptional Pathway Underlies a Novel Tumor Promoting Role of Sulfatase1 in Hepatocellular Carcinoma. Hepatology 2015, 61, 1269–1283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascale, R.M.; Calvisi, D.F.; Feo, F. Sulfatase 1: A new Jekyll and Hyde in hepatocellular carcinoma? Transl. Gastroenterol. Hepatol. 2016, 1, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, X.; Nguyen, H.T.; Chin, D.R.; Sullivan, D.E.; Lasky, J.A. Transforming Growth Factor-β1 Induces Heparan Sulfate 6-O-Endosulfatase 1 Expression in Vitro and in Vivo*. J. Biol. Chem. 2008, 283, 20397–20407. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.D.; Sun, Z.; Hu, C.; Lai, J.; Dove, R.; Nakamura, I.; Lee, J.S.; Thorgeirsson, S.S.; Kang, K.J.; Chu, I.S.; et al. Sulfatase 1 and sulfatase 2 in hepatocellular carcinoma: Associated signaling pathways, tumor phenotypes, and survival. Genes Chromosomes Cancer 2011, 59, 122–135. [Google Scholar] [CrossRef] [Green Version]
- Otsuki, S.; Hanson, S.; Miyaki, S.; Grogan, S.P.; Kinoshita, M.; Asahara, H.; Wong, C.H.; Lotz, M.K. Extracellular sulfatases support cartilage homeostasis by regulating BMP and FGF signaling pathways. Proc. Natl. Acad. Sci. USA 2010, 107, 10202–10207. [Google Scholar] [CrossRef] [Green Version]
- Buczek-Thomas, J.A.; Rich, C.; Nugent, M.A. Hypoxia Induced Heparan Sulfate Primes the Extracellular Matrix for Endothelial Cell Recruitment by Facilitating VEGF-Fibronectin Interactions. Int. J. Mol. Sci. 2019, 20, 5065. [Google Scholar] [CrossRef] [Green Version]
- Justo, T.; Martiniuc, A.; Dhoot, G.K. Modulation of cell signalling and sulfation in cardiovascular development and disease. Sci. Rep. 2021, 11, 22424. [Google Scholar] [CrossRef]
- Wang, S.A.X.; Freeman, S.D.; Pownall, M.E.; Lu, Q.; Kessler, D.S.; Emerson, C.P., Jr. QSulf1, a heparan sulfate 6-O-endosulfatase, inhibits fibroblast growth factor signaling in mesoderm induction and angiogenesis. Proc. Natl. Acad. Sci. USA 2004, 101, 4833–4838. [Google Scholar] [CrossRef] [Green Version]
- Kalus, K.; Salmen, B.; Viebahn, C.; von Figura, K.; Schmitz, D.; D’Hooge, R.; Dierks, T. Differential involvement of the extracellular 6-O-endosulfatases Sulf1 and Sulf2 in brain development and neuronal and behavioural plasticity. J. Cell Mol. Med. 2009, 13, 4505–4521. [Google Scholar] [CrossRef] [Green Version]
- Gill, R.M.; Michael, A.; Westley, L.; Kocher, H.M.; Murphy, J.I.; Dhoot, G.K. SULF1/SULF2 splice variants differentially regulate pancreatic tumour growth progression. Exp. Cell Res. 2014, 324, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Gill, R.M.; Mehra, V.; Milford, E.; Dhoot, G.K. Short SULF1/SULF2 splice variants predominate in mammary tumours with a potential to facilitate receptor tyrosine kinase-mediated cell signalling. Histochem. Cell Biol. 2016, 146, 431–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, K.; Murphy, J.I.; Dhoot, G.K. SULF1/SULF2 reactivation during liver damage and tumour growth. Histochem. Cell Biol. 2016, 146, 85–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morimoto-Tomita, M.; Uchimura, K.; Bistrup, A.; Lum, D.H.; Egeblad, M.; Boudreau, N.; Werb, Z.; Rosen, S.D. Sulf-2, a Proangiogenic Heparan Sulfate Endosulfatase, Is Upregulated in Breast Cancer. Neoplasia 2005, 7, 1001–1010. [Google Scholar] [CrossRef] [Green Version]
- Korf-Klingebiel, M.; Reboll, M.R.; Grote, K.; Schleiner, H.; Wang, Y.; Wu, X.; Klede, S.; Mikhed, Y.; Bauersachs, J.; Klintschar, M.; et al. Heparan Sulfate-Editing Extracellular Sulfatases Enhance VEGF Bioavailability for Ischemic Heart Repair. Circ. Res. 2019, 125, 787–801. [Google Scholar] [CrossRef]
- Vichai, V.; Kirtikara, K. Sulforhodamine B colorimetric assay for cytotoxicity screening. Nat. Protoc. 2006, 1, 1112–1116. [Google Scholar] [CrossRef]
- Gill, R.B.; Day, A.; Barstow, A.; Liu, H.; Zaman, G.; Dhoot, G.K. Sulf2 gene is alternatively spliced in mammalian developing and tumour tissues with functional implications. Biochem. Biophys. Res. Commun. 2011, 414, 468–473. [Google Scholar] [CrossRef]
- Sahota, A.P.; Dhoot, G.K. A novel SULF1 splice variant inhibits Wnt signalling but enhances angiogenesis by opposing SULF1 activity. Exp. Cell Res. 2009, 315, 2752–2764. [Google Scholar] [CrossRef]
- Donovan, D.; Harmey, J.H.; Toomey, D.; Osborne, D.H.; Redmond, H.P.; Bouchier-Hayes, D.J. TGF beta-1 regulation of VEGF production by breast cancer cells. Ann. Surg. Oncol. 1997, 4, 621–627. [Google Scholar] [CrossRef]
- Lebrin, F.; Deckers, M.; Bertolino, P.; Ten Dijke, P. TGF-beta receptor function in the endothelium. Cardiovasc. Res. 2005, 65, 599–608. [Google Scholar] [CrossRef]
- Zaman, G.; Staines, K.A.; Farquharson, C.; Newton, P.T.; Dudhia, J.; Chenu, C.; Pitsillides, A.A.; Dhoot, G.K. Expression of Sulf1 and Sulf2 in cartilage, bone and endochondral fracture healing. Histochem. Cell Biol. 2016, 145, 67–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, R.B.; Day, A.; Barstow, A.; Zaman, G.; Chenu, C.; Dhoot, G.K. Mammalian Sulf1 RNA alternative splicing and its significance to tumour growth regulation. Tumour Biol. 2012, 33, 1669–1680. [Google Scholar] [CrossRef] [PubMed]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Justo, T.; Smart, N.; Dhoot, G.K. Context Dependent Sulf1/Sulf2 Functional Divergence in Endothelial Cell Activity. Int. J. Mol. Sci. 2022, 23, 3769. https://doi.org/10.3390/ijms23073769
Justo T, Smart N, Dhoot GK. Context Dependent Sulf1/Sulf2 Functional Divergence in Endothelial Cell Activity. International Journal of Molecular Sciences. 2022; 23(7):3769. https://doi.org/10.3390/ijms23073769
Chicago/Turabian StyleJusto, Tiago, Nicola Smart, and Gurtej K. Dhoot. 2022. "Context Dependent Sulf1/Sulf2 Functional Divergence in Endothelial Cell Activity" International Journal of Molecular Sciences 23, no. 7: 3769. https://doi.org/10.3390/ijms23073769
APA StyleJusto, T., Smart, N., & Dhoot, G. K. (2022). Context Dependent Sulf1/Sulf2 Functional Divergence in Endothelial Cell Activity. International Journal of Molecular Sciences, 23(7), 3769. https://doi.org/10.3390/ijms23073769