
MicroRNA 3928 Suppresses Glioblastoma through Downregulation of Several Oncogenes and Upregulation of p53

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
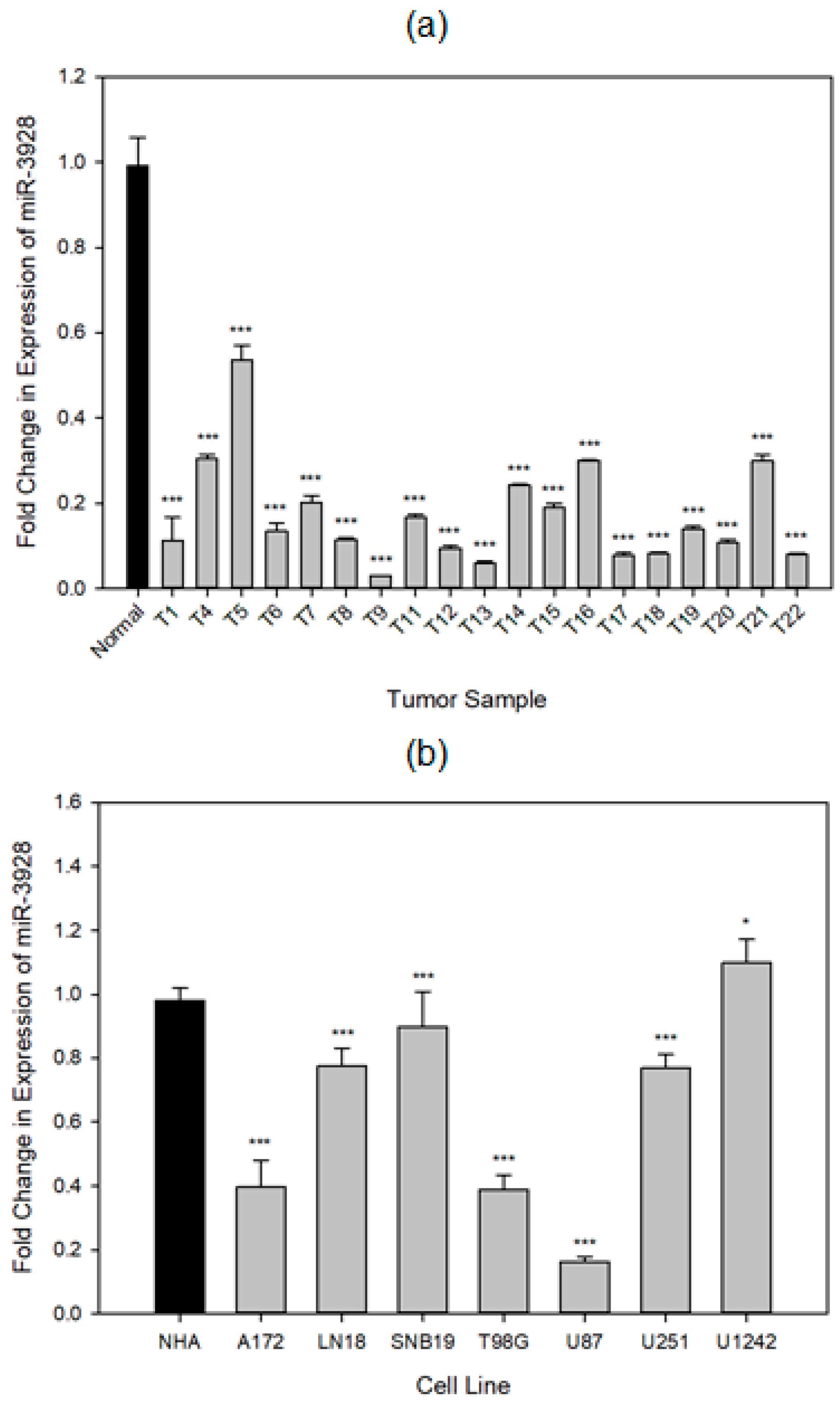
2.1. miR-3928 Expression Is Downregulated in GBM Tumors and Cell Lines
2.2. miR-3928 Inhibits GBM Cell Proliferation and Invasion
2.3. miR-3928 inhibits In Vivo GBM Xenograft Growth
2.4. miR-3298 Inhibits the Expressions of Several Oncogenes, including MDM2 and CD44, Leading to Upregulation and Activation of p53 in GBM Cells
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Tumor Specimens
4.2. Reagents
4.3. Vectors
4.4. Quantitative RT-PCR
4.5. Cell Growth Assay
4.6. Invasion Assay
4.7. Cell Cycle Assay
4.8. Animal Experiments
4.9. Immunoblot Analysis
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fromm, B.; Billipp, T.; Peck, L.E.; Johansen, M.; Tarver, J.E.; King, B.L.; Newcomb, J.M.; Sempere, L.F.; Flatmark, K.; Hovig, E.; et al. A Uniform System for the Annotation of Vertebrate microRNA Genes and the Evolution of the Human microRNAome. Annu. Rev. Genet. 2015, 49, 213–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macfarlane, L.A.; Murphy, P.R. MicroRNA: Biogenesis, Function and Role in Cancer. Curr. Genom. 2010, 11, 537–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Dodbele, S.; Park, T.; Glass, R.; Bhat, K.; Sulman, E.P.; Zhang, Y.; Abounader, R. MicroRNA-29a inhibits glioblastoma stem cells and tumor growth by regulating the PDGF pathway. J. Neurooncol. 2019, 145, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Dutta, A.; Abounader, R. The role of microRNAs in glioma initiation and progression. Front. Biosci. 2012, 17, 700–712. [Google Scholar] [CrossRef] [Green Version]
- Lamichhane, S.R.; Thachil, T.; De Ieso, P.; Gee, H.; Moss, S.A.; Milic, N. Prognostic Role of MicroRNAs in Human Non-Small-Cell Lung Cancer: A Systematic Review and Meta-Analysis. Dis. Markers 2018, 2018, 8309015. [Google Scholar] [CrossRef] [Green Version]
- Møller, H.G.; Rasmussen, A.P.; Andersen, H.H.; Johnsen, K.B.; Henriksen, M.; Duroux, M. A systematic review of microRNA in glioblastoma multiforme: Micro-modulators in the mesenchymal mode of migration and invasion. Mol. Neurobiol. 2013, 47, 131–144. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Croce, C.M. The role of MicroRNAs in human cancer. Signal Transduct. Target. Ther. 2016, 1, 15004. [Google Scholar] [CrossRef] [Green Version]
- Svoronos, A.A.; Engelman, D.M.; Slack, F.J. OncomiR or Tumor Suppressor? The Duplicity of MicroRNAs in Cancer. Cancer Res. 2016, 76, 3666–3670. [Google Scholar]
- Zhang, W.; Liu, J.; Wang, G. The role of microRNAs in human breast cancer progression. Tumour Biol. 2014, 35, 6235–6244. [Google Scholar] [CrossRef]
- Zheng, Q.; Chen, C.; Guan, H.; Kang, W.; Yu, C. Prognostic role of microRNAs in human gastrointestinal cancer: A systematic review and meta-analysis. Oncotarget 2017, 8, 46611–46623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omuro, A.; DeAngelis, L.M. Glioblastoma and other malignant gliomas: A clinical review. JAMA 2013, 310, 1S842–1850. [Google Scholar] [CrossRef] [PubMed]
- Lehrer, S.; Green, S.; Ramanathan, L.; Rosenzweig, K.; LaBombardi, V. No consistent relationship of glioblastoma incidence and cytomegalovirus seropositivity in whites, blacks, and Hispanics. Anticancer Res. 2012, 32, 1113–1115. [Google Scholar] [PubMed]
- Huang, S.-W.; Ali, N.-D.; Zhong, L.; Shi, J. MicroRNAs as biomarkers for human glioblastoma: Progress and potential. Acta Pharmacol. Sin. 2018, 39, 1405–1413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, O.G.; Brzozowski, J.S.; Skelding, K.A. Glioblastoma Multiforme: An Overview of Emerging Therapeutic Targets. Front. Oncol. 2019, 9, 963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, H.; LeBrun, D.; Yang, J.; Zhu, V.F.; Li, M. Deregulated signaling pathways in glioblastoma multiforme: Molecular mechanisms and therapeutic targets. Cancer Investig. 2012, 30, 48–56. [Google Scholar] [CrossRef] [Green Version]
- Henriksen, M.; Johnsen, K.B.; Andersen, H.H.; Pilgaard, L.; Duroux, M. MicroRNA expression signatures determine prognosis and survival in glioblastoma multiforme—A systematic overview. Mol. Neurobiol. 2014, 50, 896–913. [Google Scholar] [CrossRef] [Green Version]
- Banelli, B.; Forlani, A.; Allemanni, G.; Morabito, A.; Pistillo, M.P.; Romani, M. MicroRNA in Glioblastoma: An Overview. Int. J. Genom. 2017, 2017, 7639084. [Google Scholar] [CrossRef] [Green Version]
- Buruiană, A.; Florian, Ș.I.; Florian, A.I.; Timiș, T.L.; Mihu, C.M.; Miclăuș, M.; Oșan, S.; Hrapșa, I.; Cataniciu, R.C.; Farcaș, M.; et al. The Roles of miRNA in Glioblastoma Tumor Cell Communication: Diplomatic and Aggressive Negotiations. Int. J. Mol. Sci. 2020, 21, 1950. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Zhang, Y.; Skalski, M.; Hayes, J.; Kefas, B.; Schiff, D.; Purow, B.; Parsons, S.; Lawler, S.; Abounader, R. microRNA-148a is a prognostic oncomiR that targets MIG6 and BIM to regulate EGFR and apoptosis in glioblastoma. Cancer Res. 2014, 74, 1541–1553. [Google Scholar] [CrossRef] [Green Version]
- Kefas, B.; Floyd, D.H.; Comeau, L.; Frisbee, A.; Dominguez, C.; Dipierro, C.G.; Guessous, F.; Abounader, R.; Purow, B. A miR-297/hypoxia/DGK-alpha axis regulating glioblastoma survival. Neuro-Oncology 2013, 15, 1652–1663. [Google Scholar] [CrossRef] [PubMed]
- Kefas, B.; Comeau, L.; Floyd, D.H.; Seleverstov, O.; Godlewski, J.; Schmittgen, T.; Jiang, J.; Dipierro, C.G.; Li, Y.; Chiocca, E.A.; et al. The neuronal microRNA miR-326 acts in a feedback loop with notch and has therapeutic potential against brain tumors. J. Neurosci. 2009, 29, 15161–15168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guessous, F.; Alvarado-Velez, M.; Marcinkiewicz, L.; Zhang, Y.; Kim, J.; Heister, S.; Kefas, B.; Godlewski, J.; Schiff, D.; Purow, B.; et al. Oncogenic effects of miR-10b in glioblastoma stem cells. J. Neurooncol. 2013, 112, 153–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramaniam, S.; Jeet, V.; Gunter, J.H.; Clements, J.A.; Batra, J. Allele-Specific MicroRNA-Mediated Regulation of a Glycolysis Gatekeeper PDK1 in Cancer Metabolism. Cancers 2021, 13, 3582. [Google Scholar] [CrossRef]
- Xu, H.; Liu, X.; Zhao, J. Down-regulation of miR-3928 promoted osteosarcoma growth. Cell Physiol. Biochem. 2014, 33, 1547–1556. [Google Scholar] [CrossRef]
- Xia, E.; Kanematsu, S.; Suenaga, Y.; ElZawahry, A.; Kondo, H.; Otsuka, N.; Moriya, Y.; Iizasa, T.; Kato, M.; Yoshino, I.; et al. MicroRNA induction by copy number gain is associated with poor outcome in squamous cell carcinoma of the lung. Sci. Rep. 2018, 8, 15363. [Google Scholar] [CrossRef] [Green Version]
- Fadhil, R.S.; Wei, M.Q.; Nikolarakos, D.; Good, D.; Nair, R.G. Salivary microRNA miR-let-7a-5p and miR-3928 could be used as potential diagnostic bio-markers for head and neck squamous cell carcinoma. PLoS ONE 2020, 15, e0221779. [Google Scholar] [CrossRef] [Green Version]
- Ding, N.; Wu, X.; He, J.; Chang, L.; Hu, W.; Li, W.; Wang, J.; Wang, T.; Zhou, G. Detection of novel human MiRNAs responding to X-ray irradiation. J. Radiat. Res. 2011, 52, 425–432. [Google Scholar] [CrossRef] [Green Version]
- Chene, P. Inhibiting the p53-MDM2 interaction: An important target for cancer therapy. Nat. Rev. Cancer 2003, 3, 102–109. [Google Scholar] [CrossRef]
- Hueng, D.-Y.; Tsai, W.-C.; Chiou, H.Y.; Feng, S.-W.; Lin, C.; Li, Y.-F.; Huang, L.-C.; Lin, M.-H. DDX3X Biomarker Correlates with Poor Survival in Human Gliomas. Int. J. Mol. Sci. 2015, 16, 15578–15591. [Google Scholar] [CrossRef] [Green Version]
- Cemeli, T.; Guasch-Vallés, M.; Nàger, M.; Felip, I.; Cambray, S.; Santacana, M.; Gatius, S.; Pedraza, N.; Dolcet, X.; Ferrezuelo, F.; et al. Cytoplasmic cyclin D1 regulates glioblastoma dissemination. J. Pathol. 2019, 248, 501–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, H.; Ali, S.Z.; Huey, L.; Hütt-Cabezas, M.; Taylor, I.; Mao, X.-G.; Weingart, M.; Chu, Q.; Rodriguez, F.J.; Eberhart, C.G.; et al. The transcriptional modulator HMGA2 promotes stemness and tumorigenicity in glioblastoma. Cancer Lett. 2016, 377, 55–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleinschmidt-DeMasters, B.K.; Aisner, D.L.; Birks, D.K.; Foreman, N.K. Epithelioid GBMs show a high percentage of BRAF V600E mutation. Am. J. Surg. Pathol. 2013, 37, 685–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biernat, W.; Kleihues, P.; Yonekawa, Y.; Ohgaki, H. Amplification and overexpression of MDM2 in primary (de novo) glioblastomas. J. Neuropathol. Exp. Neurol. 1997, 56, 180–185. [Google Scholar] [CrossRef]
- Mooney, K.L.; Choy, W.; Sidhu, S.; Pelargos, P.; Bui, T.T.; Voth, B.; Barnette, N.; Yang, I. The role of CD44 in glioblastoma multiforme. J. Clin. Neurosci. 2016, 34, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Ying, M.; Wang, S.; Sang, Y.; Sun, P.; Lal, B.; Goodwin, C.R.; Guerrero-Cazares, H.; Quinones-Hinojosa, A.; Laterra, J.; Xia, S. Regulation of glioblastoma stem cells by retinoic acid: Role for Notch pathway inhibition. Oncogene 2011, 30, 3454–3467. [Google Scholar] [CrossRef] [Green Version]
- Abdouh, M.; Facchino, S.; Chatoo, W.; Balasingam, V.; Ferreira, J.; Bernier, G. BMI1 sustains human glioblastoma multiforme stem cell renewal. J. Neurosci. 2009, 29, 8884–8896. [Google Scholar] [CrossRef]
- Senbanjo, L.T.; Chellaiah, M.A. CD44: A Multifunctional Cell Surface Adhesion Receptor Is a Regulator of Progression and Metastasis of Cancer Cells. Front. Cell Dev. Biol. 2017, 5, 18. [Google Scholar] [CrossRef] [Green Version]
- Burandt, E.; Grunert, M.; Lebeau, A.; Choschzick, M.; Quaas, A.; Jänicke, F.; Müller, V.; Scholz, U.; Bokemeyer, C.; Petersen, C.; et al. Cyclin D1 gene amplification is highly homogeneous in breast cancer. Breast Cancer 2016, 23, 111–119. [Google Scholar] [CrossRef]
- Wlodarska, I.; Meeus, P.; Stul, M.; Thienpont, L.; Wouters, E.; Marcelis, L.; Demuynck, H.; Rummens, J.; Madoe, V.; Hagemeijer, A. Variant t(2;11)(p11;q13) associated with the IgK-CCND1 rearrangement is a recurrent translocation in leukemic small-cell B-non-Hodgkin lymphoma. Leukemia 2004, 18, 1705–1710. [Google Scholar] [CrossRef]
- Behling, F.; Schittenhelm, J. Oncogenic BRAF Alterations and Their Role in Brain Tumors. Cancers 2019, 11, 794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.; Pan, G.; Chen, P.L.; Ma, S.K.Y.; Zeng, T.; Chan, T.H.M.; Li, L.; Lian, Q.; Chow, R.; Cai, X.; et al. Loss of ATOH8 Increases Stem Cell Features of Hepatocellular Carcinoma Cells. Gastroenterology 2015, 149, 1068–1081.e5. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Yang, S.; Zhou, Y.; Zhong, J.; Lu, G.; Zheng, J.; Cai, Z.-Z.; He, Y.; Lin, H.; Xue, Z.-X. High expression of atonal homolog 8 predicts a poor clinical outcome in patients with colorectal cancer and contributes to tumor progression. Oncol. Rep. 2017, 37, 2955–2963. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.; Hassiotou, F.; Nowak, A. Glioblastoma stem-like cells: At the root of tumor recurrence and a therapeutic target. Carcinogenesis 2015, 36, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Cancer Genome Atlas Research, N. Comprehensive genomic characterization defines human glioblastoma genes and core pathways. Nature 2008, 455, 1061–1068. [Google Scholar] [CrossRef] [PubMed]
- Si, D.; Yin, F.; Peng, J.; Zhang, G. High Expression of CD44 Predicts a Poor Prognosis in Glioblastomas. Cancer Manag. Res. 2020, 12, 769–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auffinger, B.; Spencer, D.; Pytel, P.; Ahmed, A.U.; Lesniak, M.S. The role of glioma stem cells in chemotherapy resistance and glioblastoma multiforme recurrence. Expert Rev. Neurother. 2015, 15, 741–752. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Kim, J.; Mueller, A.C.; Dey, B.; Yang, Y.; Lee, D.-H.; Hachmann, J.; Finderle, S.; Park, D.M.; Christensen, J.; et al. Multiple receptor tyrosine kinases converge on microRNA-134 to control KRAS, STAT5B, and glioblastoma. Cell Death Differ. 2014, 21, 720–734. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mulcahy, E.Q.X.; Zhang, Y.; Colόn, R.R.; Cain, S.R.; Gibert, M.K., Jr.; Dube, C.J.; Hafner, M.; Abounader, R. MicroRNA 3928 Suppresses Glioblastoma through Downregulation of Several Oncogenes and Upregulation of p53. Int. J. Mol. Sci. 2022, 23, 3930. https://doi.org/10.3390/ijms23073930
Mulcahy EQX, Zhang Y, Colόn RR, Cain SR, Gibert MK Jr., Dube CJ, Hafner M, Abounader R. MicroRNA 3928 Suppresses Glioblastoma through Downregulation of Several Oncogenes and Upregulation of p53. International Journal of Molecular Sciences. 2022; 23(7):3930. https://doi.org/10.3390/ijms23073930
Chicago/Turabian StyleMulcahy, Elizabeth Q. X., Ying Zhang, Rossymar R. Colόn, Shelby R. Cain, Myron K. Gibert, Jr., Collin J. Dube, Markus Hafner, and Roger Abounader. 2022. "MicroRNA 3928 Suppresses Glioblastoma through Downregulation of Several Oncogenes and Upregulation of p53" International Journal of Molecular Sciences 23, no. 7: 3930. https://doi.org/10.3390/ijms23073930
APA StyleMulcahy, E. Q. X., Zhang, Y., Colόn, R. R., Cain, S. R., Gibert, M. K., Jr., Dube, C. J., Hafner, M., & Abounader, R. (2022). MicroRNA 3928 Suppresses Glioblastoma through Downregulation of Several Oncogenes and Upregulation of p53. International Journal of Molecular Sciences, 23(7), 3930. https://doi.org/10.3390/ijms23073930