
Mechanism of Action of an Environmentally Relevant Organochlorine Mixture in Repressing Steroid Hormone Biosynthesis in Leydig Cells †

 , , and
, , and 
Abstract
:1. Introduction
2. Results
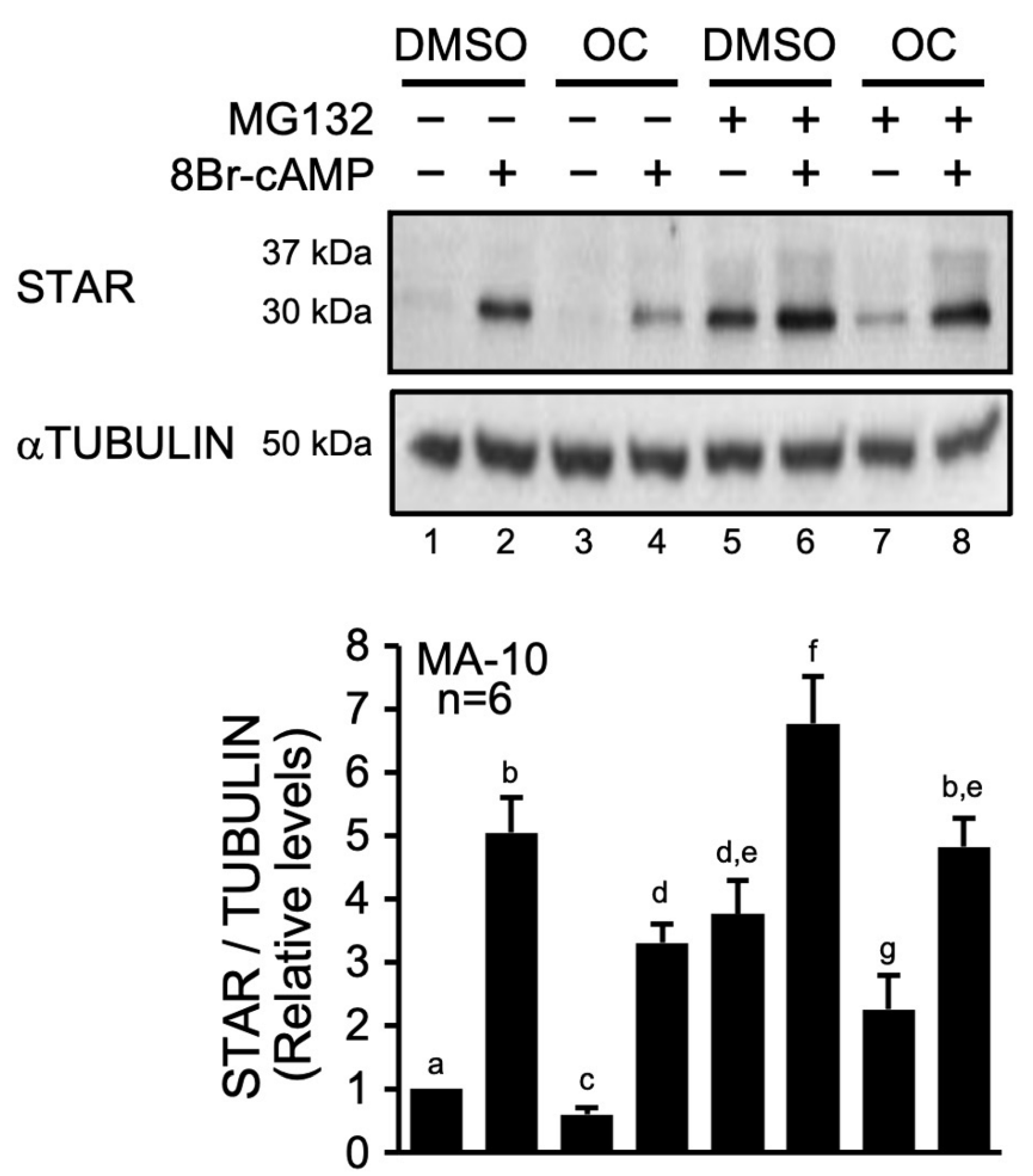
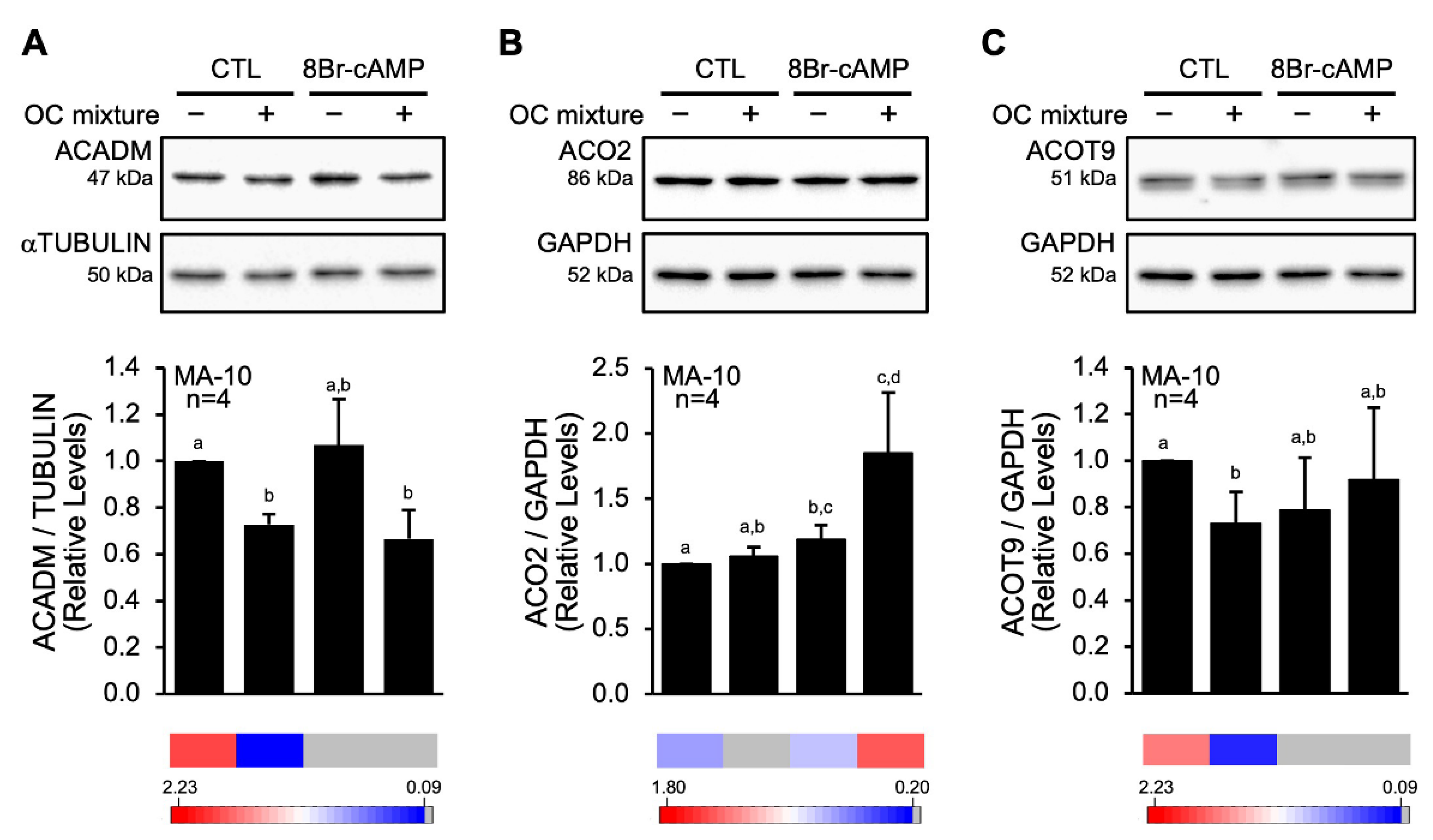
2.1. The OC Mixture Affects STAR Protein Degradation
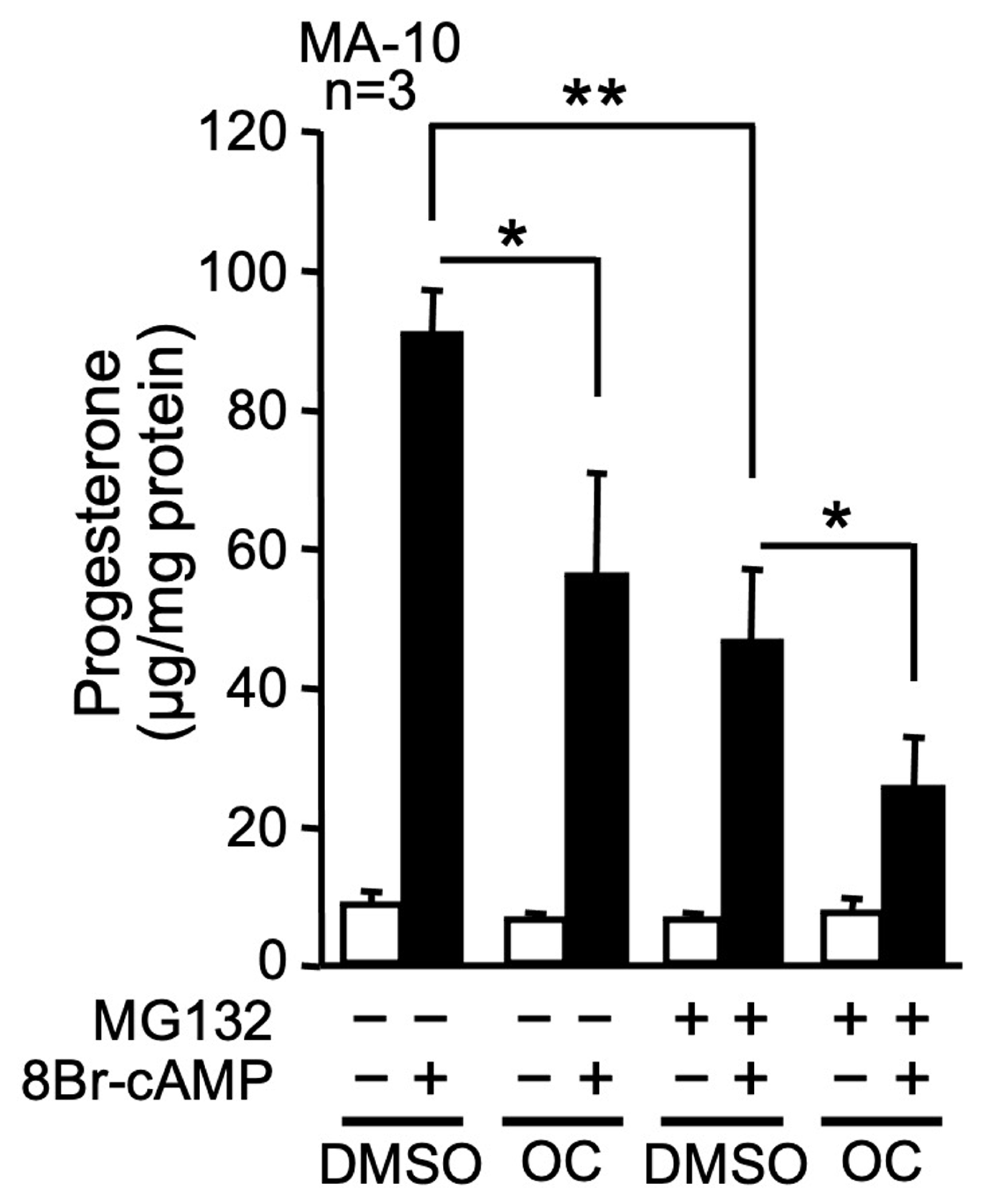
2.2. Inhibition of Protein Degradation and the OC Mixture Both Repress Progesterone Synthesis
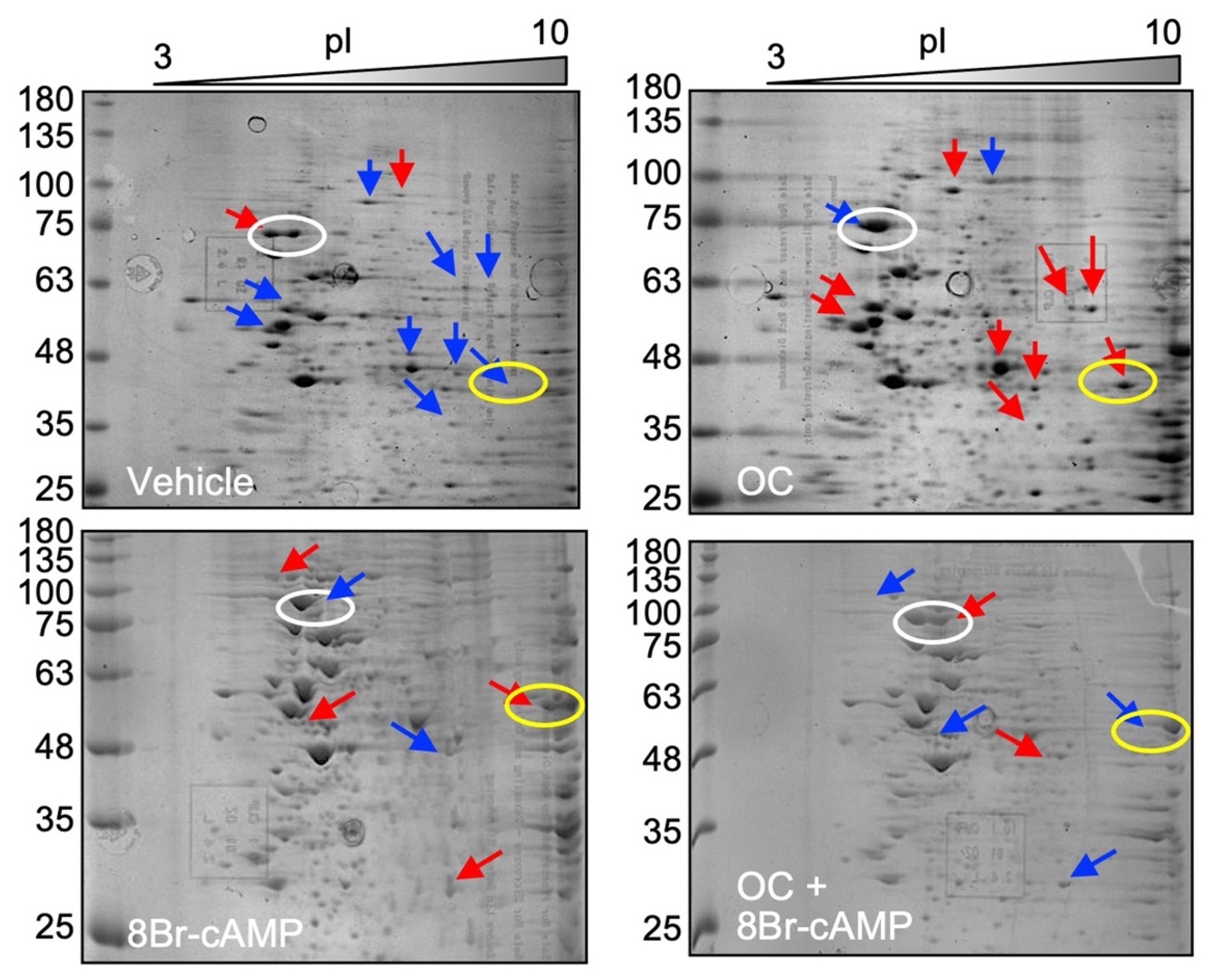
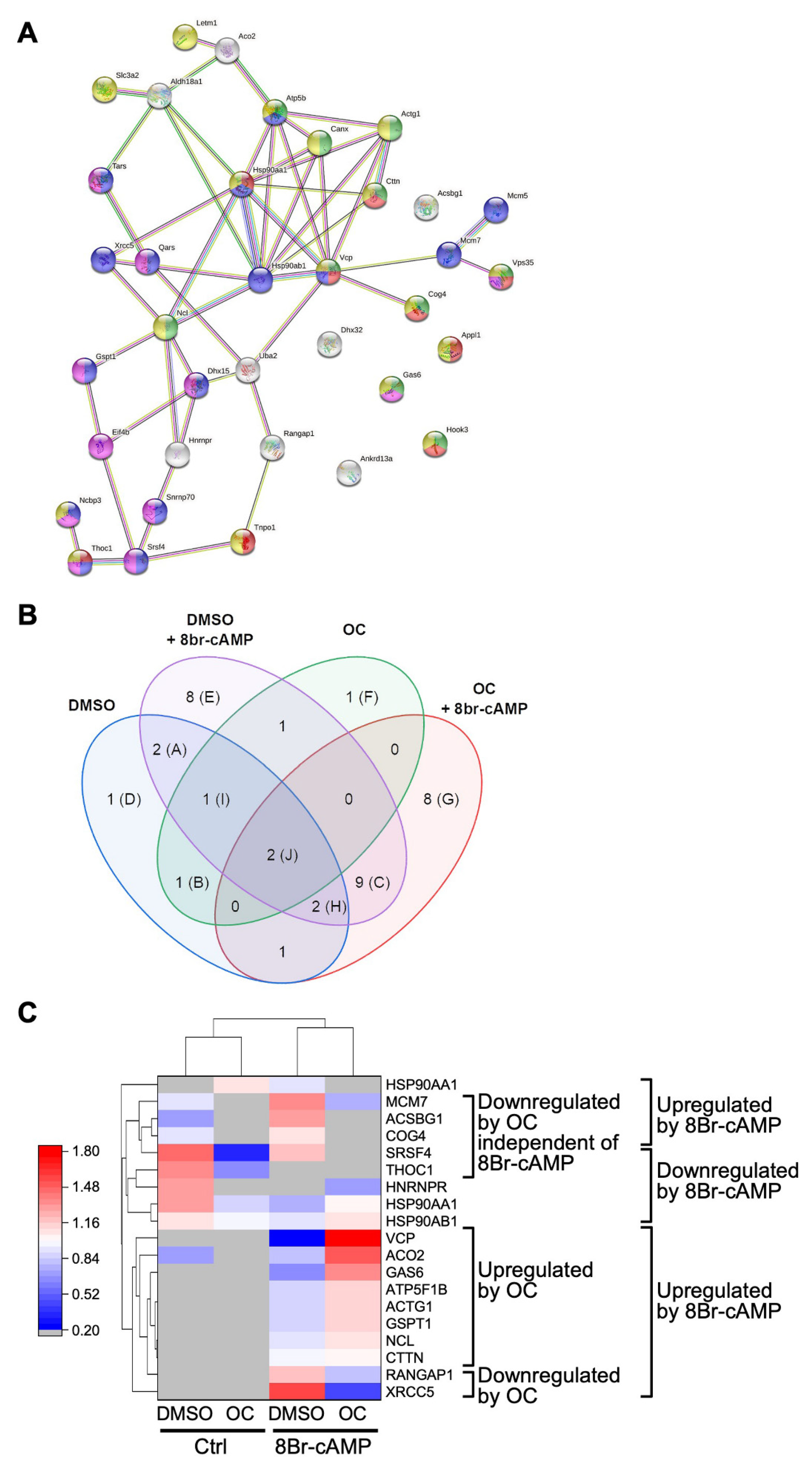
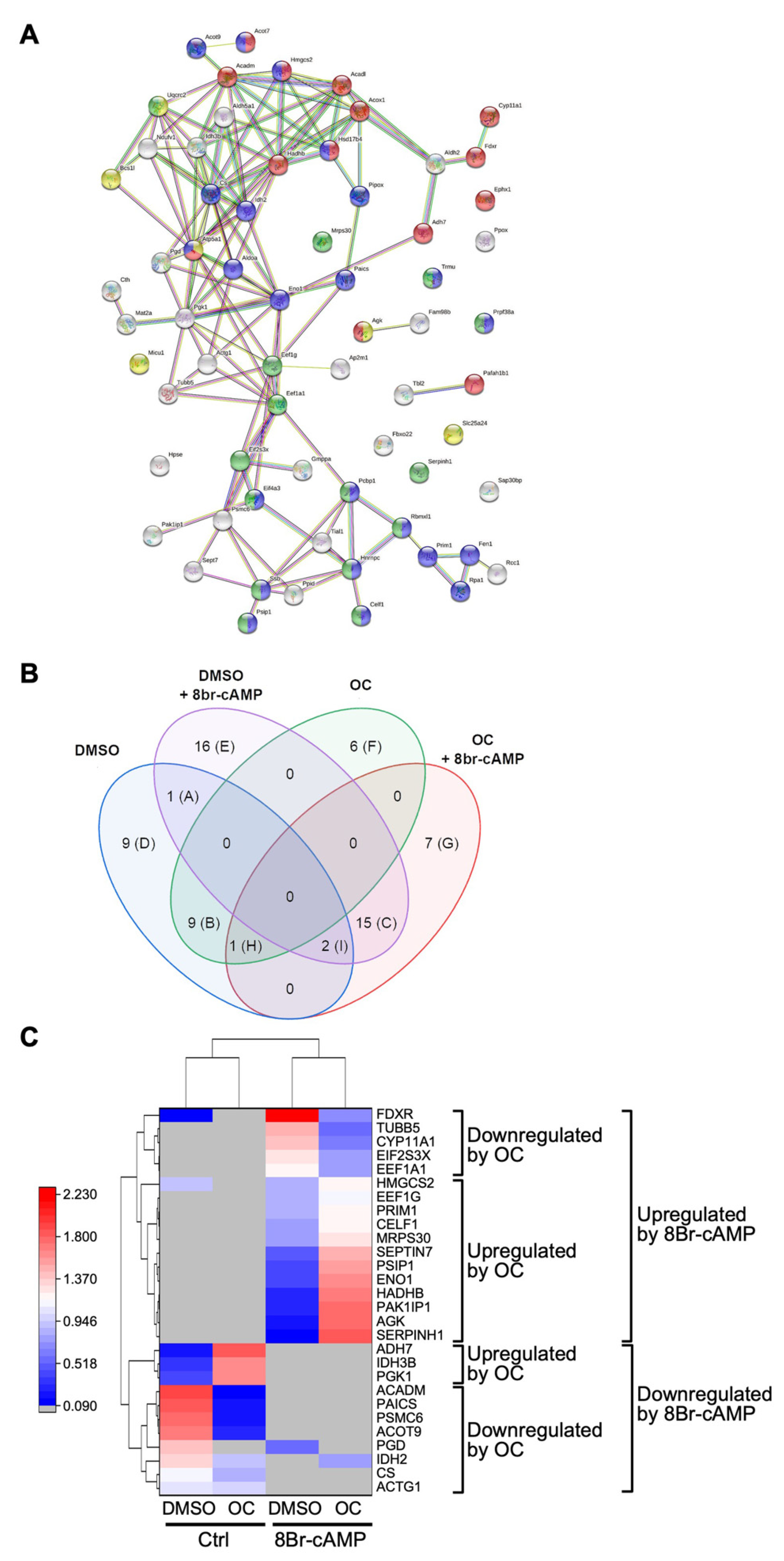
2.3. Exposure to the OC Mixture Affects Several Proteins and Biological Processes in Leydig Cells
3. Discussion
3.1. The OC Mixture Modulates STAR Protein Processing
3.2. The OC Mixture Globally Affects Proteins Involved in Multiple Biological Processes in Leydig Cells
4. Materials and Methods
4.1. Chemicals
4.2. Cell Culture
4.3. Protein Purification and Western Blot
4.4. Progesterone Quantification
4.5. Protein Purification and Two-Dimensional SDS-PAGE
4.6. Gel Fixation, Staining and Analysis
4.7. LC-MS/MS and Bioinformatic Data Analysis
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hiort, O. Androgens and puberty. Best Pract. Res. Clin. Endocrinol. Metab. 2002, 16, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Walker, W.H. Testosterone signaling and the regulation of spermatogenesis. Spermatogenesis 2011, 1, 116–120. [Google Scholar] [CrossRef] [Green Version]
- Verhoeven, G.; Willems, A.; Denolet, E.; Swinnen, J.V.; De Gendt, K. Androgens and spermatogenesis: Lessons from transgenic mouse models. Philos. Trans. R Soc. Lond. B Biol. Sci. 2010, 365, 1537–1556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haider, S.G. Cell biology of Leydig cells in the testis. Int. Rev. Cytol. 2004, 233, 181–241. [Google Scholar]
- Stocco, D.M. The role of the StAR protein in steroidogenesis: Challenges for the future. J. Endocrinol. 2000, 164, 247–253. [Google Scholar] [CrossRef]
- Issop, L.; Rone, M.B.; Papadopoulos, V. Organelle plasticity and interactions in cholesterol transport and steroid biosynthesis. Mol. Cell Endocrinol. 2013, 371, 34–46. [Google Scholar] [CrossRef] [PubMed]
- Manna, P.R.; Stetson, C.L.; Slominski, A.T.; Pruitt, K. Role of the steroidogenic acute regulatory protein in health and disease. Endocrine 2016, 51, 7–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aghazadeh, Y.; Zirkin, B.R.; Papadopoulos, V. Pharmacological regulation of the cholesterol transport machinery in steroidogenic cells of the testis. Vitam. Horm. 2015, 98, 189–227. [Google Scholar]
- Stocco, D.M.; Wang, X.; Jo, Y.; Manna, P.R. Multiple signaling pathways regulating steroidogenesis and steroidogenic acute regulatory protein expression: More complicated than we thought. Mol. Endocrinol. 2005, 19, 2647–2659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tremblay, J.J. Molecular regulation of steroidogenesis in endocrine Leydig cells. Steroids 2015, 103, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Flück, C.E.; Pandey, A.V.; Dick, B.; Camats, N.; Fernandez-Cancio, M.; Clemente, M.; Gussinye, M.; Carrascosa, A.; Mullis, P.E.; Audi, L. Characterization of novel StAR (steroidogenic acute regulatory protein) mutations causing non-classic lipoid adrenal hyperplasia. PLoS ONE 2011, 6, e20178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, W.L. Disorders in the initial steps of steroid hormone synthesis. J. Steroid Biochem. Mol. Biol. 2017, 165, 18–37. [Google Scholar] [CrossRef] [PubMed]
- Caron, K.M.; Soo, S.C.; Wetsel, W.C.; Stocco, D.M.; Clark, B.J.; Parker, K.L. Targeted disruption of the mouse gene encoding steroidogenic acute regulatory protein provides insights into congenital lipoid adrenal hyperplasia. Proc. Natl. Acad. Sci. USA 1997, 94, 11540–11545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasegawa, T.; Zhao, L.; Caron, K.M.; Majdic, G.; Suzuki, T.; Shizawa, S.; Sasano, H.; Parker, K.L. Developmental roles of the steroidogenic acute regulatory protein (StAR) as revealed by StAR knockout mice. Mol. Endocrinol. 2000, 14, 1462–1471. [Google Scholar] [CrossRef] [PubMed]
- Svechnikov, K.; Izzo, G.; Landreh, L.; Weisser, J.; Soder, O. Endocrine disruptors and Leydig cell function. J. Biomed. Biotechnol. 2010, 2010, 684504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, B.J.; Cochrum, R.K. The steroidogenic acute regulatory protein as a target of endocrine disruption in male reproduction. Drug Metab. Rev. 2007, 39, 353–370. [Google Scholar] [CrossRef] [PubMed]
- Enangue Njembele, A.N.; Tremblay, J.J. Mechanisms of MEHP Inhibitory Action and Analysis of Potential Replacement Plasticizers on Leydig Cell Steroidogenesis. Int. J. Mol. Sci. 2021, 22, 11456. [Google Scholar] [CrossRef] [PubMed]
- Enangue Njembele, A.N.; Bailey, J.L.; Tremblay, J.J. In vitro exposure of Leydig cells to an environmentally relevant mixture of organochlorines represses early steps of steroidogenesis. Biol. Reprod. 2014, 90, 118. [Google Scholar] [CrossRef] [PubMed]
- Walsh, L.P.; Stocco, D.M. Effects of lindane on steroidogenesis and steroidogenic acute regulatory protein expression. Biol. Reprod. 2000, 63, 1024–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akingbemi, B.T.; Ge, R.S.; Klinefelter, G.R.; Gunsalus, G.L.; Hardy, M.P. A metabolite of methoxychlor, 2,2-bis(p-hydroxyphenyl)-1,1, 1-trichloroethane, reduces testosterone biosynthesis in rat leydig cells through suppression of steady-state messenger ribonucleic acid levels of the cholesterol side-chain cleavage enzyme. Biol. Reprod. 2000, 62, 571–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murono, E.P.; Derk, R.C.; Akgul, Y. In vivo exposure of young adult male rats to methoxychlor reduces serum testosterone levels and ex vivo Leydig cell testosterone formation and cholesterol side-chain cleavage activity. Reprod. Toxicol. 2006, 21, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Campagna, C.; Guillemette, C.; Ayotte, P.; Bailey, J.L. Effects of an environmentally relevant organochlorine mixture and a metabolized extract of this mixture on porcine sperm parameters in vitro. J. Androl. 2009, 30, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Granot, Z.; Silverman, E.; Friedlander, R.; Melamed-Book, N.; Eimerl, S.; Timberg, R.; Hales, K.H.; Hales, D.B.; Stocco, D.M.; Orly, J. The life cycle of the steroidogenic acute regulatory (StAR) protein: From transcription through proteolysis. Endocr. Res. 2002, 28, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Granot, Z.; Geiss-Friedlander, R.; Melamed-Book, N.; Eimerl, S.; Timberg, R.; Weiss, A.M.; Hales, K.H.; Hales, D.B.; Stocco, D.M.; Orly, J. Proteolysis of normal and mutated steroidogenic acute regulatory proteins in the mitochondria: The fate of unwanted proteins. Mol. Endocrinol. 2003, 17, 2461–2476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tugaeva, K.V.; Sluchanko, N.N. Steroidogenic Acute Regulatory Protein: Structure, Functioning, and Regulation. Biochemistry 2019, 84, S233–S253. [Google Scholar] [CrossRef] [PubMed]
- Clark, B.J.; Wells, J.; King, S.R.; Stocco, D.M. The purification, cloning, and expression of a novel luteinizing hormone-induced mitochondrial protein in MA-10 mouse Leydig tumor cells. Characterization of the steroidogenic acute regulatory protein (StAR). J. Biol. Chem. 1994, 269, 28314–28322. [Google Scholar] [CrossRef]
- Arakane, F.; Sugawara, T.; Nishino, H.; Liu, Z.; Holt, J.A.; Pain, D.; Stocco, D.M.; Miller, W.L.; Strauss, J.F. 3rd Steroidogenic acute regulatory protein (StAR) retains activity in the absence of its mitochondrial import sequence: Implications for the mechanism of StAR action. Proc. Natl. Acad. Sci. USA 1996, 93, 13731–13736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, S.R.; Ronen-Fuhrmann, T.; Timberg, R.; Clark, B.J.; Orly, J.; Stocco, D.M. Steroid production after in vitro transcription, translation, and mitochondrial processing of protein products of complementary deoxyribonucleic acid for steroidogenic acute regulatory protein. Endocrinology 1995, 136, 5165–5176. [Google Scholar] [CrossRef] [PubMed]
- Granot, Z.; Kobiler, O.; Melamed-Book, N.; Eimerl, S.; Bahat, A.; Lu, B.; Braun, S.; Maurizi, M.R.; Suzuki, C.K.; Oppenheim, A.B.; et al. Turnover of mitochondrial steroidogenic acute regulatory (StAR) protein by Lon protease: The unexpected effect of proteasome inhibitors. Mol. Endocrinol. 2007, 21, 2164–2177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artemenko, I.P.; Zhao, D.; Hales, D.B.; Hales, K.H.; Jefcoate, C.R. Mitochondrial processing of newly synthesized steroidogenic acute regulatory protein (StAR), but not total StAR, mediates cholesterol transfer to cytochrome P450 side chain cleavage enzyme in adrenal cells. J. Biol. Chem. 2001, 276, 46583–46596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epstein, L.F.; Orme-Johnson, N.R. Regulation of steroid hormone biosynthesis. Identification of precursors of a phosphoprotein targeted to the mitochondrion in stimulated rat adrenal cortex cells. J. Biol. Chem. 1991, 266, 19739–19745. [Google Scholar] [CrossRef]
- Granot, Z.; Melamed-Book, N.; Bahat, A.; Orly, J. Turnover of StAR protein: Roles for the proteasome and mitochondrial proteases. Mol. Cell Endocrinol. 2007, 265–266, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Lessard, M.; Herst, P.M.; Charest, P.L.; Navarro, P.; Joly-Beauparlant, C.; Droit, A.; Kimmins, S.; Trasler, J.; Benoit-Biancamano, M.O.; MacFarlane, A.J.; et al. Prenatal Exposure to Environmentally-Relevant Contaminants Perturbs Male Reproductive Parameters Across Multiple Generations that are Partially Protected by Folic Acid Supplementation. Sci. Rep. 2019, 9, 13829. [Google Scholar] [CrossRef] [Green Version]
- Anas, M.K.; Guillemette, C.; Ayotte, P.; Pereg, D.; Giguere, F.; Bailey, J.L. In utero and lactational exposure to an environmentally relevant organochlorine mixture disrupts reproductive development and function in male rats. Biol. Reprod. 2005, 73, 414–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurice, C.; Kaczmarczyk, M.; Cote, N.; Tremblay, Y.; Kimmins, S.; Bailey, J.L. Prenatal exposure to an environmentally relevant mixture of Canadian Arctic contaminants decreases male reproductive function in an aging rat model. J. Dev. Orig. Health Dis. 2018, 9, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Payne, A.H. Steroidogenic enzymes in Leydig cells. In The Leydig Cell in Health and Disease; Payne, A., Hardy, M., Eds.; Humana Press: Totowa, NJ, USA, 2007; pp. 157–171. [Google Scholar]
- Rone, M.B.; Midzak, A.S.; Martinez-Arguelles, D.B.; Fan, J.; Ye, X.; Blonder, J.; Papadopoulos, V. Steroidogenesis in MA-10 mouse Leydig cells is altered via fatty acid import into the mitochondria. Biol. Reprod. 2014, 91, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dees, J.H.; Gazouli, M.; Papadopoulos, V. Effect of mono-ethylhexyl phthalate on MA-10 Leydig tumor cells. Reprod. Toxicol. 2001, 15, 171–187. [Google Scholar] [CrossRef]
- Muir, D.; Braune, B.; DeMarch, B.; Norstrom, R.; Wagemann, R.; Lockhart, L.; Hargrave, B.; Bright, D.; Addison, R.; Payne, J.; et al. Spatial and temporal trends and effects of contaminants in the Canadian Arctic marine ecosystem: A review. Sci. Total Environ. 1999, 230, 83–144. [Google Scholar] [CrossRef]
- Bjerregaard, P.; Hansen, J.C. Organochlorines and heavy metals in pregnant women from the Disko Bay area in Greenland. Sci. Total Environ. 2000, 245, 195–202. [Google Scholar] [CrossRef]
- Ascoli, M. Characterization of several clonal lines of cultured Leydig tumor cells: Gonadotropin receptors and steroidogenic responses. Endocrinology 1981, 108, 88–95. [Google Scholar] [CrossRef]
- Rahman, N.A.; Huhtaniemi, I.T. Testicular cell lines. Mol. Cell Endocrinol. 2004, 228, 53–65. [Google Scholar] [CrossRef]
- Trbovich, A.M.; Martinelle, N.; O’Neill, F.H.; Pearson, E.J.; Donahoe, P.K.; Sluss, P.M.; Teixeira, J. Steroidogenic activities in MA-10 Leydig cells are differentially altered by cAMP and Mullerian inhibiting substance. J. Steroid Biochem. Mol. Biol. 2004, 92, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Abdou, H.S.; Bergeron, F.; Tremblay, J.J. A cell-autonomous molecular cascade initiated by AMP-activated protein kinase represses steroidogenesis. Mol. Cell Biol. 2014, 34, 4257–4271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdou, H.S.; Villeneuve, G.; Tremblay, J.J. The calcium signaling pathway regulates leydig cell steroidogenesis through a transcriptional cascade involving the nuclear receptor NR4A1 and the steroidogenic acute regulatory protein. Endocrinology 2013, 154, 511–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R Stat. Soc. Ser. B Stat. Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | CAS no. | % Weight |
|---|---|---|
| PCB mixture * | 32.6 | |
| Technical chlordane | 57-74-9 | 21.3 |
| P,P’-DDE | 72-55-9 | 19.3 |
| P,P’-DDT | 50-29-3 | 6.79 |
| Technical toxaphene | 8001-35-2 | 6.54 |
| α-HCH | 319-84-6 | 6.17 |
| Aldrin | 309-00-2 | 2.52 |
| Dieldrin | 60-57-1 | 2.09 |
| 1,2,4,5-Tetrachlorobenzene | 95-94-3 | 0.86 |
| P,P’-DDD | 72-54-8 | 0.49 |
| β-HCH | 319-85-7 | 0.46 |
| Hexachlorobenzene | 118-74-1 | 0.35 |
| Mirex | 2385-85-5 | 0.23 |
| γ-HCH | 58-89-9 | 0.20 |
| Pentachlorobenzene | 608-93-5 | 0.18 |
| GO Term ID | Term Description-Biological Process | Observed Gene Count | Background Gene Count | Strength | FDR | Matching Proteins in Your Network (Labels) |
|---|---|---|---|---|---|---|
| GO:0009987 | Cellular process | 33 | 13330 | 0.18 | 0.0196 | Mcm7,Letm1,Qars,Canx,Ncbp3,Tars,Aco2,Hsp90ab1,Thoc1,Aldh18a1,Atp5b,Xrcc5,Ncl,Vcp,Dhx15,Gas6,Vps35,Cog4,Acsbg1,Appl1,Hook3,Srsf4,Actg1,Snrnp70,Gspt1,Hsp90aa1,Cttn,Uba2,Tnpo1,Mcm5,Rangap1,Eif4b,Slc3a2 |
| GO:0044238 | Primary metabolic process | 23 | 6369 | 0.34 | 0.0145 | Mcm7,Qars,Ncbp3,Tars,Aco2,Hsp90ab1,Thoc1,Aldh18a1,Atp5b,Xrcc5,Vcp,Dhx15,Gas6,Vps35,Acsbg1,Srsf4,Snrnp70,Gspt1,Hsp90aa1,Uba2,Mcm5,Eif4b,Slc3a2 |
| GO:0071704 | Organic substance metabolic process | 23 | 6824 | 0.31 | 0.0271 | Mcm7,Qars,Ncbp3,Tars,Aco2,Hsp90ab1,Thoc1,Aldh18a1,Atp5b,Xrcc5,Vcp,Dhx15,Gas6,Vps35,Acsbg1,Srsf4,Snrnp70,Gspt1,Hsp90aa1,Uba2,Mcm5,Eif4b,Slc3a2 |
| GO:0044237 | Cellular metabolic process | 22 | 6445 | 0.32 | 0.0350 | Mcm7,Qars,Ncbp3,Tars,Aco2,Hsp90ab1,Thoc1,Aldh18a1,Atp5b,Xrcc5,Vcp,Dhx15,Gas6,Vps35,Acsbg1,Srsf4,Snrnp70,Gspt1,Hsp90aa1,Uba2,Mcm5,Eif4b |
| GO:0051179 | Localization | 18 | 4646 | 0.38 | 0.0445 | Letm1,Canx,Ncbp3,Thoc1,Atp5b,Ncl,Vcp,Gas6,Vps35,Cog4,Appl1,Hook3,Actg1,Hsp90aa1,Cttn,Tnpo1,Rangap1,Slc3a2 |
| GO:0006810 | Transport | 17 | 3382 | 0.49 | 0.0116 | Letm1,Canx,Ncbp3,Thoc1,Atp5b,Ncl,Vcp,Gas6,Vps35,Cog4,Appl1,Hook3,Actg1,Hsp90aa1,Cttn,Tnpo1,Slc3a2 |
| GO:0046483 | Heterocycle metabolic process | 16 | 2347 | 0.62 | 0.0044 | Mcm7,Qars,Ncbp3,Tars,Hsp90ab1,Thoc1,Aldh18a1,Atp5b,Xrcc5,Vcp,Dhx15,Srsf4,Snrnp70,Gspt1,Hsp90aa1,Mcm5 |
| GO:1901360 | Organic cyclic compound metabolic process | 16 | 2614 | 0.57 | 0.0048 | Mcm7,Qars,Ncbp3,Tars,Hsp90ab1,Thoc1,Aldh18a1,Atp5b,Xrcc5,Vcp,Dhx15,Srsf4,Snrnp70,Gspt1,Hsp90aa1,Mcm5 |
| GO:0034641 | Cellular nitrogen compound metabolic process | 16 | 2805 | 0.54 | 0.0070 | Mcm7,Qars,Ncbp3,Tars,Hsp90ab1,Thoc1,Atp5b,Xrcc5,Vcp,Dhx15,Srsf4,Snrnp70,Gspt1,Hsp90aa1,Mcm5,Eif4b |
| GO:0006139 | Nucleobase-containing compound metabolic process | 15 | 2205 | 0.62 | 0.0044 | Mcm7,Qars,Ncbp3,Tars,Hsp90ab1,Thoc1,Atp5b,Xrcc5,Vcp,Dhx15,Srsf4,Snrnp70,Gspt1,Hsp90aa1,Mcm5 |
| GO:0010033 | Response to organic substance | 15 | 2742 | 0.53 | 0.0141 | Mcm7,Canx,Hsp90ab1,Atp5b,Xrcc5,Ncl,Vcp,Dhx15,Gas6,Acsbg1,Appl1,Actg1,Hsp90aa1,Rangap1,Slc3a2 |
| GO:0090304 | Nucleic acid metabolic process | 14 | 1777 | 0.68 | 0.0044 | Mcm7,Qars,Ncbp3,Tars,Hsp90ab1,Thoc1,Xrcc5,Vcp,Dhx15,Srsf4,Snrnp70,Gspt1,Hsp90aa1,Mcm5 |
| GO:0051641 | Cellular localization | 14 | 2115 | 0.61 | 0.0070 | Letm1,Canx,Thoc1,Atp5b,Vcp,Gas6,Vps35,Appl1,Hook3,Actg1,Hsp90aa1,Cttn,Tnpo1,Rangap1 |
| GO:0022607 | Cellular component assembly | 13 | 1983 | 0.6 | 0.0130 | Mcm7,Letm1,Hsp90ab1,Xrcc5,Vcp,Gas6,Vps35,Cog4,Actg1,Hsp90aa1,Cttn,Mcm5,Eif4b |
| GO:0044249 | Cellular biosynthetic process | 13 | 2025 | 0.59 | 0.0140 | Mcm7,Qars,Tars,Hsp90ab1,Thoc1,Aldh18a1,Atp5b,Vcp,Acsbg1,Gspt1,Hsp90aa1,Mcm5,Eif4b |
| GO:1901576 | Organic substance biosynthetic process | 13 | 2121 | 0.57 | 0.0164 | Mcm7,Qars,Tars,Hsp90ab1,Thoc1,Aldh18a1,Atp5b,Vcp,Acsbg1,Gspt1,Hsp90aa1,Mcm5,Eif4b |
| GO:0033036 | Macromolecule localization | 13 | 2198 | 0.56 | 0.0194 | Ncbp3,Thoc1,Vcp,Gas6,Vps35,Cog4,Appl1,Hook3,Actg1,Hsp90aa1,Cttn,Tnpo1,Rangap1 |
| GO:0051649 | Establishment of localization in cell | 12 | 1518 | 0.68 | 0.0070 | Letm1,Canx,Thoc1,Atp5b,Vcp,Vps35,Appl1,Hook3,Actg1,Hsp90aa1,Cttn,Tnpo1 |
| GO:0008104 | Protein localization | 12 | 1890 | 0.59 | 0.0194 | Thoc1,Vcp,Gas6,Vps35,Cog4,Appl1,Hook3,Actg1,Hsp90aa1,Cttn,Tnpo1,Rangap1 |
| GO:0043933 | Protein-containing complex subunit organization | 11 | 1164 | 0.76 | 0.0057 | Mcm7,Letm1,Hsp90ab1,Xrcc5,Vcp,Cog4,Gspt1,Hsp90aa1,Cttn,Mcm5,Eif4b |
| GO:0071705 | Nitrogen compound transport | 11 | 1534 | 0.64 | 0.0194 | Ncbp3,Thoc1,Vcp,Vps35,Cog4,Appl1,Hook3,Hsp90aa1,Cttn,Tnpo1,Slc3a2 |
| GO:0010467 | Gene expression | 11 | 1743 | 0.59 | 0.0350 | Qars,Ncbp3,Tars,Thoc1,Dhx15,Gas6,Vps35,Srsf4,Snrnp70,Gspt1,Eif4b |
| GO:0071702 | Organic substance transport | 11 | 1836 | 0.56 | 0.0445 | Ncbp3,Thoc1,Vcp,Vps35,Cog4,Appl1,Hook3,Hsp90aa1,Cttn,Tnpo1,Slc3a2 |
| GO:0016192 | Vesicle-mediated transport | 10 | 1130 | 0.73 | 0.0140 | Canx,Atp5b,Ncl,Vcp,Gas6,Vps35,Cog4,Hook3,Actg1,Cttn |
| GO:0034645 | Cellular macromolecule biosynthetic process | 10 | 1233 | 0.7 | 0.0188 | Mcm7,Qars,Tars,Hsp90ab1,Thoc1,Vcp,Gspt1,Hsp90aa1,Mcm5,Eif4b |
| GO:0034613 | Cellular protein localization | 10 | 1392 | 0.64 | 0.0312 | Thoc1,Vcp,Gas6,Vps35,Appl1,Hook3,Hsp90aa1,Cttn,Tnpo1,Rangap1 |
| GO:0065003 | Protein-containing complex assembly | 9 | 1025 | 0.73 | 0.0196 | Mcm7,Letm1,Hsp90ab1,Xrcc5,Cog4,Hsp90aa1,Cttn,Mcm5,Eif4b |
| GO:0046907 | Intracellular transport | 9 | 1166 | 0.67 | 0.0390 | Thoc1,Atp5b,Vcp,Vps35,Appl1,Hook3,Hsp90aa1,Cttn,Tnpo1 |
| GO:0015031 | Protein transport | 9 | 1204 | 0.66 | 0.0430 | Thoc1,Vcp,Vps35,Cog4,Appl1,Hook3,Hsp90aa1,Cttn,Tnpo1 |
| GO:0034622 | Cellular protein-containing complex assembly | 8 | 683 | 0.86 | 0.0141 | Mcm7,Hsp90ab1,Xrcc5,Cog4,Hsp90aa1,Cttn,Mcm5,Eif4b |
| GO:1903827 | Regulation of cellular protein localization | 7 | 576 | 0.87 | 0.0216 | Hsp90ab1,Gas6,Vps35,Appl1,Hsp90aa1,Ankrd13a,Rangap1 |
| GO:0006259 | DNA metabolic process | 7 | 638 | 0.83 | 0.0350 | Mcm7,Hsp90ab1,Thoc1,Xrcc5,Vcp,Hsp90aa1,Mcm5 |
| GO:0071345 | Cellular response to cytokine stimulus | 7 | 698 | 0.79 | 0.0455 | Hsp90ab1,Atp5b,Xrcc5,Ncl,Gas6,Appl1,Actg1 |
| GO:0046822 | Regulation of nucleocytoplasmic transport | 4 | 129 | 1.28 | 0.0328 | Hsp90ab1,Gas6,Hsp90aa1,Rangap1 |
| GO:0009651 | Response to salt stress | 3 | 31 | 1.77 | 0.0194 | Hsp90ab1,Xrcc5,Hsp90aa1 |
| GO:0034605 | Cellular response to heat | 3 | 57 | 1.51 | 0.0466 | Hsp90ab1,Vcp,Hsp90aa1 |
| GO:1905323 | Telomerase holoenzyme complex assembly | 2 | 3 | 2.61 | 0.0194 | Hsp90ab1,Hsp90aa1 |
| GO:0006267 | Pre-replicative complex assembly involved in nuclear cell cycle DNA replication | 2 | 7 | 2.24 | 0.0415 | Mcm7,Mcm5 |
| GO:0019062 | Virion attachment to host cell | 2 | 7 | 2.24 | 0.0415 | Hsp90ab1,Gas6 |
| A | B | C | D | E | F | G | H | I | J | Other |
|---|---|---|---|---|---|---|---|---|---|---|
| ACSBG1 | THOC1 | CTTN | TNPO1 | EIF4B | ANKRD13A | DHX15 | ACO2 | SRSF4 | HSP90AB1 | HSP90AA1 |
| COG4 | GSPT1 | HOOK3 | TARS | MCM7 | HSP90AA1 | HNRNPR | ||||
| ACTG1 | VPS35 | MCM5 | ||||||||
| RANGAP1 | NCBP3 | DHX32 | ||||||||
| NCL | SLC3A2 | UBA2 | ||||||||
| VCP | APPL1 | ALDH18A1 | ||||||||
| XRCC5 | CANX | QARS | ||||||||
| GAS6 | LETM1 | SNRNP70 | ||||||||
| ATP5F1B |
| Upregulated by 8Br-cAMP and Downregulated by the OC Mixture: | |
| RANGAP1: | Ran GTPase-activating protein 1. Activation of GTPase activity |
| XRCC5: | X-ray repair cross-complementing protein 5. Single-stranded DNA-dependent ATP-dependent helicase that plays a key role in DNA non-homologous end joining (NHEJ) by recruiting DNA-PK to DNA |
| Upregulated by 8Br-cAMP and Upregulated by the OC Mixture: | |
| VCP: | Transitional endoplasmic reticulum ATPase. Involved in DNA damage response, apoptosis, ER stress, autophagy. |
| ACO2: | Aconitate hydratase. Catalyzes the isomerization of citrate to isocitrate via cis-aconitate. TCA/Krebs cycle. Energy metabolism. |
| GAS6: | Growth arrest-specific protein 6. Ligand for tyrosine-protein kinase receptors AXL, TYRO3, and MER, whose signaling is implicated in cell growth and survival, cell adhesion, and cell migration. |
| ATP5F1B: | ATP synthase subunit beta. Component of the mitochondrial membrane ATP synthase. |
| ACTG1: | Actin. |
| GSPT1: | Eukaryotic peptide chain release factor GTP-binding subunit ERF3A. Involved in translation termination, cell growth. |
| NCL: | Nucleolin. The major nucleolar protein of growing eukaryotic cells. It induces chromatin decondensation by binding to histone H1. |
| CTTN: | Src substrate cortactin. Contributes to the organization of the actin cytoskeleton and cell shape. |
| Downregulated by the OC Mixture Independent of 8Br-cAMP: | |
| MCM7: | DNA replication licensing factor MCM7. Component of the MCM2-7 complex (MCM complex). |
| ACSBG1: | Long-chain-fatty-acid--CoA ligase. Catalyzes the conversion of fatty acids such as long-chain and very long-chain fatty acids to their active form acyl-CoAs for both synthesis of cellular lipids and degradation via beta-oxidation. |
| COG4: | Conserved oligomeric Golgi complex subunit 4. Required for normal Golgi function. |
| SRSF4: | Serine/arginine-rich splicing factor 4. Plays a role in alternative splice site selection during pre-mRNA splicing. |
| THOC1: | THO complex subunit 1. Required for efficient export of polyadenylated RNA. Participates in an apoptotic pathway. Essential for early embryonic development. Required for normal gene expression during postnatal testis development. |
| GO Term ID | Term Description-Biological Process | Observed Gene Count | Background Gene Count | Strength | FDR | Matching Proteins in Your Network (Labels) |
|---|---|---|---|---|---|---|
| GO:0009987 | Cellular process | 63 | 13330 | 0.2 | 1.37 × 10−7 | Rpa1,Tubb5,Cs,Ap2m1,Pipox,Fdxr,Pafah1b1,Mrps30,Psmc6,Trmu,Hsd17b4,Fen1,Acot9,Prim1,Atp5a1,Eif4a3,Acadl,Bcs1l,Fam98b,Ppid,Slc25a24,Psip1,Paics,Aldh2,Agk,Uqcrc2,Fbxo22,Cyp11a1,Aldh5a1,Pak1ip1,Eef1a1,Ndufv1,Hpse,Ephx1,Rbmxl1,Pcbp1,Eif2s3x,Acox1,Celf1,Actg1,Acadm,Ppox,Acot7,Prpf38a,Eno1,Pgd,Rcc1,Aldoa,Mat2a,Adh7,Hmgcs2,Ssb,Eef1g,Tial1,Idh2,Hnrnpc,Hadhb,Sept7,Cth,Sap30bp,Tbl2,Serpinh1,Micu1 |
| GO:0008152 | Metabolic process | 53 | 7331 | 0.38 | 1.06 × 10−10 | Rpa1,Cs,Ap2m1,Pipox,Fdxr,Pafah1b1,Mrps30,Psmc6,Trmu,Hsd17b4,Fen1,Acot9,Prim1,Atp5a1,Eif4a3,Acadl,Fam98b,Ppid,Psip1,Paics,Aldh2,Agk,Uqcrc2,Fbxo22,Cyp11a1,Gmppa,Aldh5a1,Eef1a1,Ndufv1,Hpse,Ephx1,Rbmxl1,Pcbp1,Eif2s3x,Acox1,Celf1,Acadm,Ppox,Acot7,Prpf38a,Eno1,Pgd,Aldoa,Mat2a,Adh7,Hmgcs2,Ssb,Eef1g,Idh2,Hnrnpc,Hadhb,Cth,Serpinh1 |
| GO:0071704 | Organic substance metabolic process | 51 | 6824 | 0.4 | 1.06 × 10−10 | Rpa1,Cs,Ap2m1,Pipox,Fdxr,Pafah1b1,Mrps30,Psmc6,Trmu,Hsd17b4,Fen1,Acot9,Prim1,Atp5a1,Eif4a3,Acadl,Fam98b,Ppid,Psip1,Paics,Aldh2,Agk,Uqcrc2,Fbxo22,Cyp11a1,Aldh5a1,Eef1a1,Hpse,Ephx1,Rbmxl1,Pcbp1,Eif2s3x,Acox1,Celf1,Acadm,Ppox,Acot7,Prpf38a,Eno1,Pgd,Aldoa,Mat2a,Adh7,Hmgcs2,Ssb,Eef1g,Idh2,Hnrnpc,Hadhb,Cth,Serpinh1 |
| GO:0044237 | Cellular metabolic process | 50 | 6445 | 0.41 | 1.06 × 10−10 | Rpa1,Cs,Ap2m1,Pipox,Fdxr,Mrps30,Psmc6,Trmu,Hsd17b4,Fen1,Acot9,Prim1,Atp5a1,Eif4a3,Acadl,Fam98b,Ppid,Psip1,Paics,Aldh2,Agk,Uqcrc2,Fbxo22,Cyp11a1,Aldh5a1,Eef1a1,Ndufv1,Hpse,Ephx1,Rbmxl1,Pcbp1,Eif2s3x,Acox1,Celf1,Acadm,Ppox,Acot7,Prpf38a,Eno1,Pgd,Aldoa,Mat2a,Adh7,Hmgcs2,Ssb,Eef1g,Idh2,Hnrnpc,Hadhb,Cth |
| GO:0044238 | Primary metabolic process | 47 | 6369 | 0.39 | 6.80 × 10−9 | Rpa1,Cs,Pipox,Fdxr,Pafah1b1,Mrps30,Psmc6,Trmu,Hsd17b4,Fen1,Acot9,Prim1,Atp5a1,Eif4a3,Acadl,Fam98b,Ppid,Psip1,Paics,Agk,Uqcrc2,Fbxo22,Cyp11a1,Aldh5a1,Eef1a1,Hpse,Ephx1,Rbmxl1,Pcbp1,Eif2s3x,Acox1,Celf1,Acadm,Acot7,Prpf38a,Eno1,Pgd,Aldoa,Adh7,Hmgcs2,Ssb,Eef1g,Idh2,Hnrnpc,Hadhb,Cth,Serpinh1 |
| GO:0006807 | Nitrogen compound metabolic process | 40 | 5878 | 0.36 | 7.55 × 10−6 | Rpa1,Cs,Pipox,Mrps30,Psmc6,Trmu,Hsd17b4,Fen1,Acot9,Prim1,Atp5a1,Eif4a3,Acadl,Fam98b,Ppid,Psip1,Paics,Agk,Uqcrc2,Fbxo22,Aldh5a1,Eef1a1,Hpse,Rbmxl1,Pcbp1,Eif2s3x,Celf1,Acadm,Ppox,Acot7,Prpf38a,Eno1,Aldoa,Hmgcs2,Ssb,Eef1g,Idh2,Hnrnpc,Cth,Serpinh1 |
| GO:0034641 | Cellular nitrogen compound metabolic process | 32 | 2805 | 0.58 | 7.04 × 10−9 | Rpa1,Cs,Pipox,Mrps30,Trmu,Hsd17b4,Fen1,Acot9,Prim1,Atp5a1,Eif4a3,Acadl,Psip1,Paics,Agk,Eef1a1,Rbmxl1,Pcbp1,Eif2s3x,Celf1,Acadm,Ppox,Acot7,Prpf38a,Eno1,Aldoa,Hmgcs2,Ssb,Eef1g,Idh2,Hnrnpc,Cth |
| GO:1901564 | Organonitrogen compound metabolic process | 29 | 4475 | 0.34 | 4.1 × 10−3 | Cs,Pipox,Mrps30,Psmc6,Hsd17b4,Acot9,Atp5a1,Acadl,Fam98b,Ppid,Paics,Agk,Uqcrc2,Fbxo22,Aldh5a1,Eef1a1,Hpse,Rbmxl1,Eif2s3x,Acadm,Ppox,Acot7,Eno1,Aldoa,Hmgcs2,Eef1g,Idh2,Cth,Serpinh1 |
| GO:1901360 | Organic cyclic compound metabolic process | 27 | 2614 | 0.54 | 2.61 × 10−6 | Rpa1,Cs,Pipox,Fdxr,Trmu,Hsd17b4,Fen1,Acot9,Prim1,Atp5a1,Eif4a3,Psip1,Paics,Cyp11a1,Ephx1,Rbmxl1,Pcbp1,Celf1,Ppox,Acot7,Prpf38a,Eno1,Aldoa,Hmgcs2,Ssb,Idh2,Hnrnpc |
| GO:0009058 | Biosynthetic process | 25 | 2176 | 0.58 | 1.90 × 10−6 | Rpa1,Fdxr,Mrps30,Fen1,Prim1,Atp5a1,Paics,Agk,Cyp11a1,Gmppa,Eef1a1,Ephx1,Rbmxl1,Eif2s3x,Acadm,Ppox,Acot7,Pgd,Aldoa,Mat2a,Hmgcs2,Eef1g,Idh2,Cth,Serpinh1 |
| GO:0046483 | Heterocycle metabolic process | 25 | 2347 | 0.55 | 6.8 × 10−6 | Rpa1,Cs,Pipox,Trmu,Hsd17b4,Fen1,Acot9,Prim1,Atp5a1,Eif4a3,Psip1,Paics,Ephx1,Rbmxl1,Pcbp1,Celf1,Ppox,Acot7,Prpf38a,Eno1,Aldoa,Hmgcs2,Ssb,Idh2,Hnrnpc |
| GO:0006725 | Cellular aromatic compound metabolic process | 25 | 2412 | 0.54 | 1 × 10−5 | Rpa1,Cs,Pipox,Trmu,Hsd17b4,Fen1,Acot9,Prim1,Atp5a1,Eif4a3,Psip1,Paics,Ephx1,Rbmxl1,Pcbp1,Celf1,Ppox,Acot7,Prpf38a,Eno1,Aldoa,Hmgcs2,Ssb,Idh2,Hnrnpc |
| GO:0044281 | Small molecule metabolic process | 24 | 1450 | 0.74 | 7.04 × 10−9 | Cs,Pipox,Fdxr,Hsd17b4,Acot9,Atp5a1,Acadl,Paics,Aldh2,Cyp11a1,Aldh5a1,Ephx1,Acox1,Acadm,Acot7,Eno1,Pgd,Aldoa,Mat2a,Adh7,Hmgcs2,Idh2,Hadhb,Cth |
| GO:1901576 | Organic substance biosynthetic process | 23 | 2121 | 0.56 | 1.52 × 10−5 | Rpa1,Fdxr,Mrps30,Fen1,Prim1,Atp5a1,Paics,Agk,Cyp11a1,Eef1a1,Ephx1,Rbmxl1,Eif2s3x,Acadm,Ppox,Acot7,Pgd,Aldoa,Mat2a,Hmgcs2,Eef1g,Idh2,Cth |
| GO:0006139 | Nucleobase-containing compound metabolic process | 23 | 2205 | 0.54 | 2.84 × 10−5 | Rpa1,Cs,Pipox,Trmu,Hsd17b4,Fen1,Acot9,Prim1,Atp5a1,Eif4a3,Psip1,Paics,Rbmxl1,Pcbp1,Celf1,Acot7,Prpf38a,Eno1,Aldoa,Hmgcs2,Ssb,Idh2,Hnrnpc |
| GO:0044249 | Cellular biosynthetic process | 21 | 2025 | 0.54 | 1.2 × 10−4 | Rpa1,Fdxr,Mrps30,Fen1,Prim1,Atp5a1,Paics,Agk,Cyp11a1,Eef1a1,Rbmxl1,Eif2s3x,Acadm,Ppox,Acot7,Aldoa,Mat2a,Hmgcs2,Eef1g,Idh2,Cth |
| GO:0009056 | Catabolic process | 20 | 1680 | 0.6 | 3.22 × 10−5 | Pipox,Pafah1b1,Psmc6,Hsd17b4,Fen1,Eif4a3,Acadl,Aldh2,Aldh5a1,Hpse,Ephx1,Acox1,Acadm,Acot7,Eno1,Pgd,Aldoa,Adh7,Ssb,Hadhb |
| GO:1901575 | Organic substance catabolic process | 19 | 1425 | 0.65 | 1.52 × 10−5 | Pipox,Pafah1b1,Psmc6,Hsd17b4,Fen1,Eif4a3,Acadl,Aldh2,Aldh5a1,Hpse,Acox1,Acadm,Acot7,Eno1,Pgd,Aldoa,Adh7,Ssb,Hadhb |
| GO:0055114 | Oxidation-reduction process | 18 | 917 | 0.82 | 3.49 × 10−7 | Cs,Pipox,Fdxr,Hsd17b4,Acadl,Aldh2,Uqcrc2,Cyp11a1,Aldh5a1,Ndufv1,Acox1,Acadm,Ppox,Eno1,Pgd,Adh7,Idh2,Hadhb |
| GO:0044085 | Cellular component biogenesis | 18 | 2201 | 0.44 | 1.69 × 10−2 | Tubb5,Ap2m1,Pafah1b1,Eif4a3,Bcs1l,Ppid,Psip1,Pak1ip1,Rbmxl1,Eif2s3x,Celf1,Actg1,Rcc1,Aldoa,Mat2a,Sept7,Cth,Micu1 |
| GO:0019752 | Carboxylic acid metabolic process | 16 | 754 | 0.85 | 1.18 × 10−6 | Cs,Pipox,Hsd17b4,Acadl,Aldh5a1,Ephx1,Acox1,Acadm,Acot7,Eno1,Pgd,Aldoa,Adh7,Idh2,Hadhb,Cth |
| GO:0022607 | Cellular component assembly | 16 | 1983 | 0.43 | 4.86 × 10−2 | Tubb5,Ap2m1,Pafah1b1,Bcs1l,Ppid,Psip1,Rbmxl1,Eif2s3x,Celf1,Actg1,Rcc1,Aldoa,Mat2a,Sept7,Cth,Micu1 |
| GO:0044271 | Cellular nitrogen compound biosynthetic process | 15 | 1156 | 0.64 | 5.3 × 10−4 | Rpa1,Mrps30,Prim1,Atp5a1,Paics,Agk,Eef1a1,Rbmxl1,Eif2s3x,Acadm,Ppox,Acot7,Aldoa,Eef1g,Idh2 |
| GO:0044248 | Cellular catabolic process | 15 | 1445 | 0.54 | 6.3 × 10−3 | Pipox,Psmc6,Hsd17b4,Fen1,Eif4a3,Acadl,Aldh5a1,Hpse,Ephx1,Acox1,Acadm,Acot7,Pgd,Ssb,Hadhb |
| GO:0010467 | Gene expression | 15 | 1743 | 0.46 | 4.06 × 10−2 | Mrps30,Trmu,Eif4a3,Psip1,Uqcrc2,Eef1a1,Rbmxl1,Pcbp1,Eif2s3x,Celf1,Prpf38a,Ssb,Eef1g,Hnrnpc,Serpinh1 |
| GO:0006629 | Lipid metabolic process | 14 | 1032 | 0.66 | 7.3 × 10−4 | Fdxr,Pafah1b1,Hsd17b4,Atp5a1,Acadl,Agk,Cyp11a1,Ephx1,Acox1,Acadm,Acot7,Adh7,Hmgcs2,Hadhb |
| GO:0032787 | Monocarboxylic acid metabolic process | 13 | 461 | 0.97 | 1.82 × 10−6 | Hsd17b4,Acadl,Aldh5a1,Ephx1,Acox1,Acadm,Acot7,Eno1,Pgd,Aldoa,Adh7,Idh2,Hadhb |
| GO:1901566 | Organonitrogen compound biosynthetic process | 13 | 1096 | 0.6 | 6.3 × 10−3 | Mrps30,Atp5a1,Paics,Agk,Eef1a1,Eif2s3x,Acadm,Ppox,Acot7,Aldoa,Eef1g,Idh2,Cth |
| GO:0044282 | Small molecule catabolic process | 12 | 318 | 1.1 | 4.65 × 10−7 | Pipox,Hsd17b4,Acadl,Aldh2,Aldh5a1,Acox1,Acadm,Acot7,Eno1,Pgd,Adh7,Hadhb |
| GO:0043603 | Cellular amide metabolic process | 12 | 689 | 0.76 | 3.7 × 10−4 | Cs,Pipox,Mrps30,Hsd17b4,Acot9,Agk,Eef1a1,Eif2s3x,Acot7,Hmgcs2,Eef1g,Cth |
| GO:0019637 | Organophosphate metabolic process | 12 | 724 | 0.74 | 6 × 10−4 | Cs,Pipox,Hsd17b4,Acot9,Atp5a1,Paics,Acot7,Eno1,Pgd,Aldoa,Hmgcs2,Idh2 |
| GO:1901135 | Carbohydrate derivative metabolic process | 12 | 814 | 0.69 | 1.8 × 10−3 | Cs,Pipox,Hsd17b4,Acot9,Atp5a1,Paics,Hpse,Acot7,Eno1,Pgd,Aldoa,Hmgcs2 |
| GO:0009117 | Nucleotide metabolic process | 11 | 361 | 1.01 | 1.03 × 10−5 | Cs,Pipox,Hsd17b4,Acot9,Atp5a1,Paics,Acot7,Eno1,Aldoa,Hmgcs2,Idh2 |
| GO:1901362 | Organic cyclic compound biosynthetic process | 11 | 883 | 0.62 | 1.7 × 10−2 | Rpa1,Prim1,Atp5a1,Paics,Cyp11a1,Rbmxl1,Ppox,Acot7,Aldoa,Hmgcs2,Idh2 |
| GO:0009150 | Purine ribonucleotide metabolic process | 10 | 265 | 1.1 | 7.91 × 10−6 | Cs,Pipox,Hsd17b4,Acot9,Atp5a1,Paics,Acot7,Eno1,Aldoa,Hmgcs2 |
| GO:0006091 | Generation of precursor metabolites and energy | 10 | 319 | 1.02 | 2.73 × 10−5 | Cs,Atp5a1,Ndufv1,Acox1,Acadm,Eno1,Pgd,Aldoa,Adh7,Idh2 |
| GO:0044255 | Cellular lipid metabolic process | 10 | 797 | 0.62 | 3.38 × 10−2 | Hsd17b4,Acadl,Agk,Ephx1,Acox1,Acadm,Acot7,Adh7,Hmgcs2,Hadhb |
| GO:0046395 | Carboxylic acid catabolic process | 9 | 206 | 1.16 | 1.14 × 10−5 | Pipox,Hsd17b4,Acadl,Aldh5a1,Acox1,Acadm,Acot7,Pgd,Hadhb |
| GO:0072329 | Monocarboxylic acid catabolic process | 8 | 99 | 1.43 | 1.67 × 10−6 | Hsd17b4,Acadl,Aldh5a1,Acox1,Acadm,Acot7,Pgd,Hadhb |
| GO:0006790 | Sulfur compound metabolic process | 8 | 280 | 0.98 | 8 × 10−4 | Cs,Pipox,Hsd17b4,Acot9,Acot7,Mat2a,Hmgcs2,Cth |
| GO:0006631 | Fatty acid metabolic process | 8 | 310 | 0.94 | 1.6 × 10−3 | Hsd17b4,Acadl,Ephx1,Acox1,Acadm,Acot7,Adh7,Hadhb |
| GO:0016071 | mRNA metabolic process | 8 | 516 | 0.71 | 3.99 × 10−2 | Eif4a3,Psip1,Rbmxl1,Pcbp1,Celf1,Prpf38a,Ssb,Hnrnpc |
| GO:0030258 | Lipid modification | 7 | 212 | 1.04 | 1.4 × 10−3 | Hsd17b4,Acadl,Agk,Acox1,Acadm,Adh7,Hadhb |
| GO:0016042 | Lipid catabolic process | 7 | 260 | 0.95 | 4.8 × 10−3 | Pafah1b1,Hsd17b4,Acadl,Acox1,Acadm,Acot7,Hadhb |
| GO:0006066 | Alcohol metabolic process | 7 | 292 | 0.9 | 8.6 × 10−3 | Fdxr,Aldh2,Cyp11a1,Ephx1,Adh7,Hmgcs2,Idh2 |
| GO:0006397 | mRNA processing | 7 | 399 | 0.77 | 4.86 × 10−2 | Eif4a3,Psip1,Rbmxl1,Pcbp1,Celf1,Prpf38a,Hnrnpc |
| GO:0019395 | Fatty acid oxidation | 6 | 77 | 1.42 | 8.83 × 10−5 | Hsd17b4,Acadl,Acox1,Acadm,Adh7,Hadhb |
| GO:0009062 | Fatty acid catabolic process | 6 | 79 | 1.4 | 9.91 × 10−5 | Hsd17b4,Acadl,Acox1,Acadm,Acot7,Hadhb |
| GO:0006637 | acyl-CoA metabolic process | 6 | 87 | 1.36 | 1.6 × 10−4 | Cs,Pipox,Hsd17b4,Acot9,Acot7,Hmgcs2 |
| GO:0006839 | Mitochondrial transport | 6 | 184 | 1.04 | 6.6 × 10−3 | Atp5a1,Bcs1l,Slc25a24,Agk,Uqcrc2,Micu1 |
| GO:0000398 | mRNA splicing, via spliceosome | 6 | 191 | 1.02 | 8 × 10−3 | Eif4a3,Psip1,Rbmxl1,Celf1,Prpf38a,Hnrnpc |
| GO:0006635 | Fatty acid beta-oxidation | 5 | 54 | 1.49 | 3.6 × 10−4 | Hsd17b4,Acadl,Acox1,Acadm,Hadhb |
| GO:0006376 | mRNA splice site selection | 3 | 27 | 1.57 | 2.54 × 10−2 | Psip1,Rbmxl1,Celf1 |
| GO:0006084 | acetyl-CoA metabolic process | 3 | 32 | 1.5 | 3.94 × 10−2 | Cs,Pipox,Hmgcs2 |
| GO:0006739 | NADP metabolic process | 3 | 32 | 1.5 | 3.94 × 10−2 | Fdxr,Pgd,Idh2 |
| GO:0019254 | Carnitine metabolic process, coa-linked | 2 | 3 | 2.35 | 2.35 × 10−2 | Acadl,Acadm |
| GO:0036112 | Medium-chain fatty-acyl-coa metabolic process | 2 | 3 | 2.35 | 2.35 × 10−2 | Hsd17b4,Acot7 |
| A | B | C | D | E | F | G | H | I |
|---|---|---|---|---|---|---|---|---|
| PGD | ADH7 | EEF1A1 | RBMXL1 | AP2M1 | BCS1L | TRMU | IDH2 | FDXR |
| PGK1 | CYP11A1 | HNRNPC | SSB | ALDOA | HSD17B4 | HMGCS2 | ||
| ACADM | SEPTIN7 | CTH | PAFAH1B1 | TIAL1 | MICU1 | |||
| PSMC6 | AGK | PPID | ALDH5A1 | PRPF38A | TBL2 | |||
| CS | CELF1 | GMPPA | ALDH2 | UQCRC2 | FEN1 | |||
| IDH3B | HADHB | SLC25A24 | EPHX1 | ACADL | FAM98B | |||
| ACTG1 | EIF2S3X | FBXO22 | EIF4A3 | SAP30BP | ||||
| PAICS | SERPINH1 | PCBP1 | ACOX | |||||
| ACOT9 | PAK1IP1 | PIPOX | RPA1 | |||||
| TUBB5 | PPOX | |||||||
| ENO1 | ATP5F1A | |||||||
| EEF1G | MAT2A | |||||||
| MRPS30 | HPSE | |||||||
| PSIP1 | NDUFV1 | |||||||
| PRIM1 | ACOT7 | |||||||
| RCC1 |
| UPREGULATED BY 8Br-cAMP | |
| Downregulated by OC: | |
| FDXR: | NADPH:adrenodoxin oxidoreductase. Serves as the first electron transfer protein in all the mitochondrial P450 systems, including cholesterol side chain cleavage in all steroidogenic tissues. |
| TUBB5: | Tubulin beta-5 chain. The major constituent of microtubules. |
| CYP11A1: | Cholesterol side-chain cleavage enzyme. Catalyzes the side-chain hydroxylation and cleavage of cholesterol to pregnenolone, the precursor of most steroid hormones. |
| EIF2S3X: | Eukaryotic translation initiation factor 2 subunit 3, X-linked. As a subunit of eukaryotic initiation factor 2 (eIF-2), involved in the early steps of protein synthesis. |
| EEF1A1: | Elongation factor 1-alpha 1. Promotes the GTP-dependent binding of aminoacyl-tRNA to the A-site of ribosomes during protein biosynthesis. |
| Upregulated by OC: | |
| HMGCS2: | Hydroxymethylglutaryl-CoA synthase. Catalyzes the first irreversible step in ketogenesis and cholesterogenesis, condensing acetyl-CoA to acetoacetyl-CoA to form HMG-CoA. |
| EEF1G: | Elongation factor 1-gamma. Role in translation elongation. |
| PRIM1: | DNA primase small subunit. Catalytic subunit of the DNA primase complex and component of the DNA polymerase alpha complex (also known as the alpha DNA polymerase-primase complex), which play an essential role in the initiation of DNA synthesis. |
| CELF1: | CUGBP Elav-like family member 1. RNA-binding protein implicated in the regulation of several post-transcriptional events. Involved in pre-mRNA alternative splicing, mRNA translation, and stability. Mediates exon inclusion and/or exclusion in pre-mRNA that are subject to tissue-specific and developmentally regulated alternative splicing. Increases translation and controls the choice of translation initiation codon of CEBPB mRNA (C/EBPb is an important regulator of several steroidogenic genes in Leydig cells). |
| MRPS30: | 28S ribosomal protein S30. Structural constituent of ribosome. |
| SEPTIN7: | Septin-7. Filament-forming cytoskeletal GTPase. Required for normal organization of the actin cytoskeleton. Required for normal progress through mitosis. Involved in cytokinesis. |
| PSIP1: | PC4 and SFRS1-interacting protein. Transcriptional coactivator involved in neuroepithelial stem cell differentiation and neurogenesis. Involved in particular in lens epithelial cell gene regulation and stress responses. May play an important role in lens epithelial to fiber cell terminal differentiation. May play a protective role during stress-induced apoptosis. |
| ENO1: | Alpha-enolase. Glycolytic enzyme the catalyzes the conversion of 2-phosphoglycerate to phosphoenolpyruvate (precursor for the synthesis of ATP). |
| HADHB: | Trifunctional enzyme subunit beta. Mitochondrial trifunctional enzyme catalyzes the last three of the four reactions of the mitochondrial beta-oxidation pathway. The mitochondrial beta-oxidation pathway is the major energy-producing process in tissues and is performed through four consecutive reactions, breaking down fatty acids into acetyl-CoA. Among the enzymes involved in this pathway, the trifunctional enzyme exhibits specificity for long-chain fatty acids. |
| PAK1IP1: | p21-activated protein kinase-interacting protein 1. Negatively regulates the PAK1 kinase. Involved in cell proliferation. May be involved in ribosomal large subunit assembly. |
| AGK: | Acylglycerol kinase. Lipid kinase that can phosphorylate both monoacylglycerol and diacylglycerol to form lysophosphatidic acid (LPA) and phosphatidic acid (PA), respectively. |
| SERPINH1: | Serpin H1. Binds specifically to collagen. Could be involved as a chaperone in the biosynthetic pathway of collagen. |
| DOWNREGULATED BY 8Br-cAMP | |
| Downregulated by OC: | |
| ACADM: | Medium-chain specific acyl-CoA dehydrogenase. One of the acyl-CoA dehydrogenases that catalyze the first step of mitochondrial fatty acid beta-oxidation, an aerobic process breaking down fatty acids into acetyl-CoA and allowing the production of energy from fats. |
| PAICS: | Multifunctional protein ADE2. IMP biosynthesis (SAICAR more specifically). |
| PSMC6: | 26S proteasome regulatory subunit 10B. Component of the 26S proteasome, a multiprotein complex involved in the ATP-dependent degradation of ubiquitinated proteins. This complex plays a key role in the maintenance of protein homeostasis by removing misfolded or damaged proteins, which could impair cellular functions, and by removing proteins whose functions are no longer required. |
| ACOT9: | Acyl-coenzyme A thioesterase 9. Acyl-CoA thioesterases are a group of enzymes that catalyze the hydrolysis of acyl-CoAs to the free fatty acid and coenzyme A (CoASH), providing the potential to regulate intracellular levels of acyl-CoAs, free fatty acids, and CoASH. Active on long chain acyl-CoAs. |
| PGD: | 6-phosphogluconate dehydrogenase, decarboxylating. Catalyzes the oxidative decarboxylation of 6-phosphogluconate to ribulose 5-phosphate and CO2, with concomitant reduction of NADP to NADPH. Pentose phosphate pathway. |
| IDH2: | Isocitrate dehydrogenase. Plays a role in intermediary metabolism and energy production. |
| CS: | Citrate synthase. TCA/Krebs cycle for energy production. |
| ACTG1: | Actin, cytoplasmic 2. Actins coexist in most cell types as components of the cytoskeleton and as mediators of internal cell motility/contractility. |
| Upregulated by OC: | |
| ADH7: | All-trans-retinol dehydrogenase. Catalyzes the NAD-dependent oxidation of all-trans-retinol, alcohol, aldehyde, and omega-hydroxy fatty acids and their derivatives. Therefore, it may participate in retinoid metabolism, fatty acid omega-oxidation, and elimination of cytotoxic aldehydes produced by lipid peroxidation. |
| IDH3B: | Isocitrate dehydrogenase (NAD) subunit alpha. Catalytic subunit of the enzyme which catalyzes the decarboxylation of isocitrate (ICT) into alpha-ketoglutarate. TCA/Krebs cycle for energy production. |
| PGK1: | Phosphoglycerate kinase 1. Catalyzes one of the two ATP-producing reactions in the glycolytic pathway via the reversible conversion of 1,3-diphosphoglycerate to 3-phosphoglycerate. In addition to its role as a glycolytic enzyme, it seems that PGK-1 acts as a polymerase alpha cofactor protein (primer recognition protein). May play a role in sperm motility. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Enangue Njembele, A.N.; Demmouche, Z.B.; Bailey, J.L.; Tremblay, J.J. Mechanism of Action of an Environmentally Relevant Organochlorine Mixture in Repressing Steroid Hormone Biosynthesis in Leydig Cells . Int. J. Mol. Sci. 2022, 23, 3997. https://doi.org/10.3390/ijms23073997
Enangue Njembele AN, Demmouche ZB, Bailey JL, Tremblay JJ. Mechanism of Action of an Environmentally Relevant Organochlorine Mixture in Repressing Steroid Hormone Biosynthesis in Leydig Cells . International Journal of Molecular Sciences. 2022; 23(7):3997. https://doi.org/10.3390/ijms23073997
Chicago/Turabian StyleEnangue Njembele, Annick N., Zoheir B. Demmouche, Janice L. Bailey, and Jacques J. Tremblay. 2022. "Mechanism of Action of an Environmentally Relevant Organochlorine Mixture in Repressing Steroid Hormone Biosynthesis in Leydig Cells " International Journal of Molecular Sciences 23, no. 7: 3997. https://doi.org/10.3390/ijms23073997
APA StyleEnangue Njembele, A. N., Demmouche, Z. B., Bailey, J. L., & Tremblay, J. J. (2022). Mechanism of Action of an Environmentally Relevant Organochlorine Mixture in Repressing Steroid Hormone Biosynthesis in Leydig Cells . International Journal of Molecular Sciences, 23(7), 3997. https://doi.org/10.3390/ijms23073997