
Circ003429 Regulates Unsaturated Fatty Acid Synthesis in the Dairy Goat Mammary Gland by Interacting with miR-199a-3p, Targeting the YAP1 Gene

 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
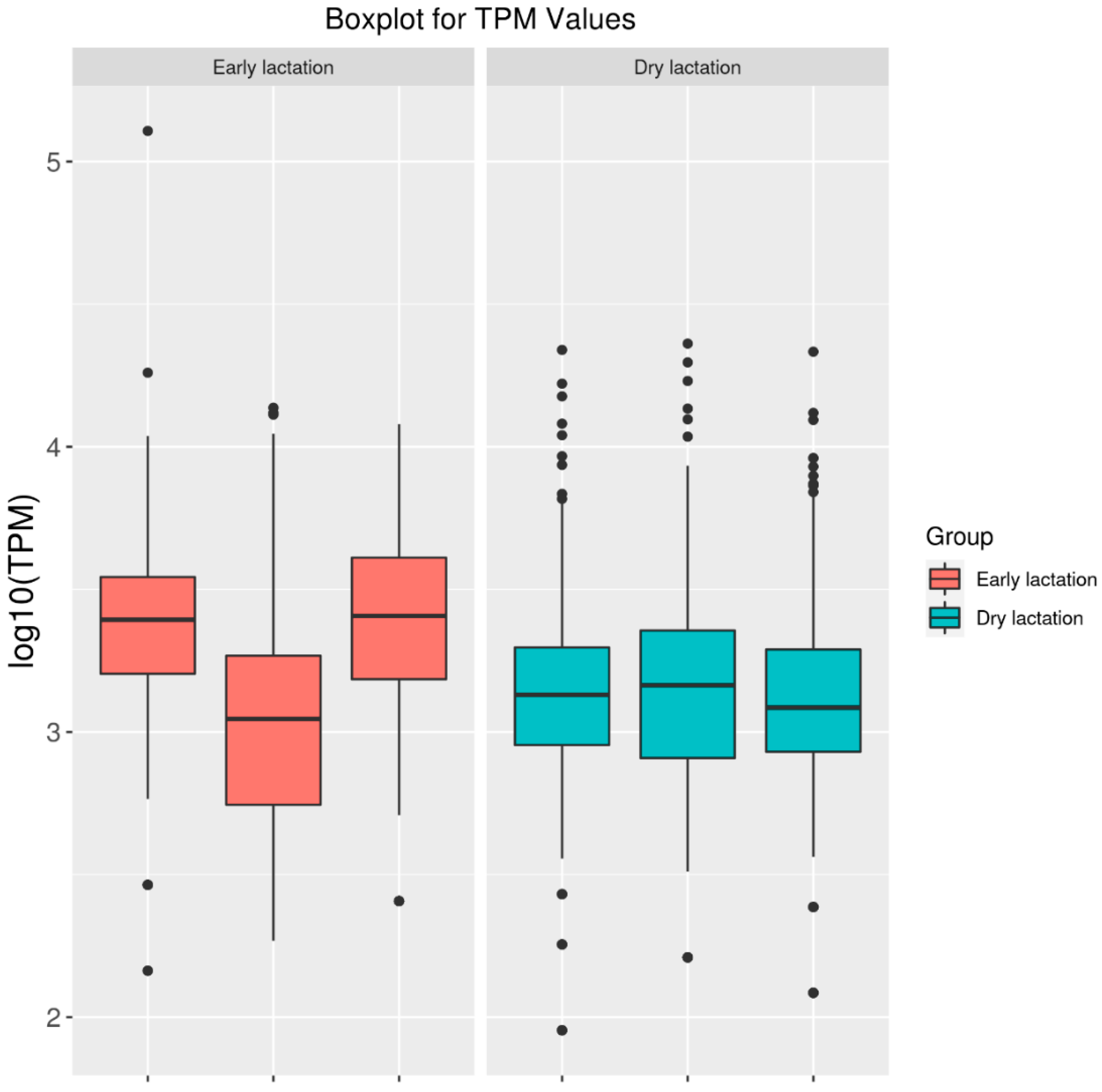
2.1. High-Throughput Sequencing of Mammary Gland Tissue
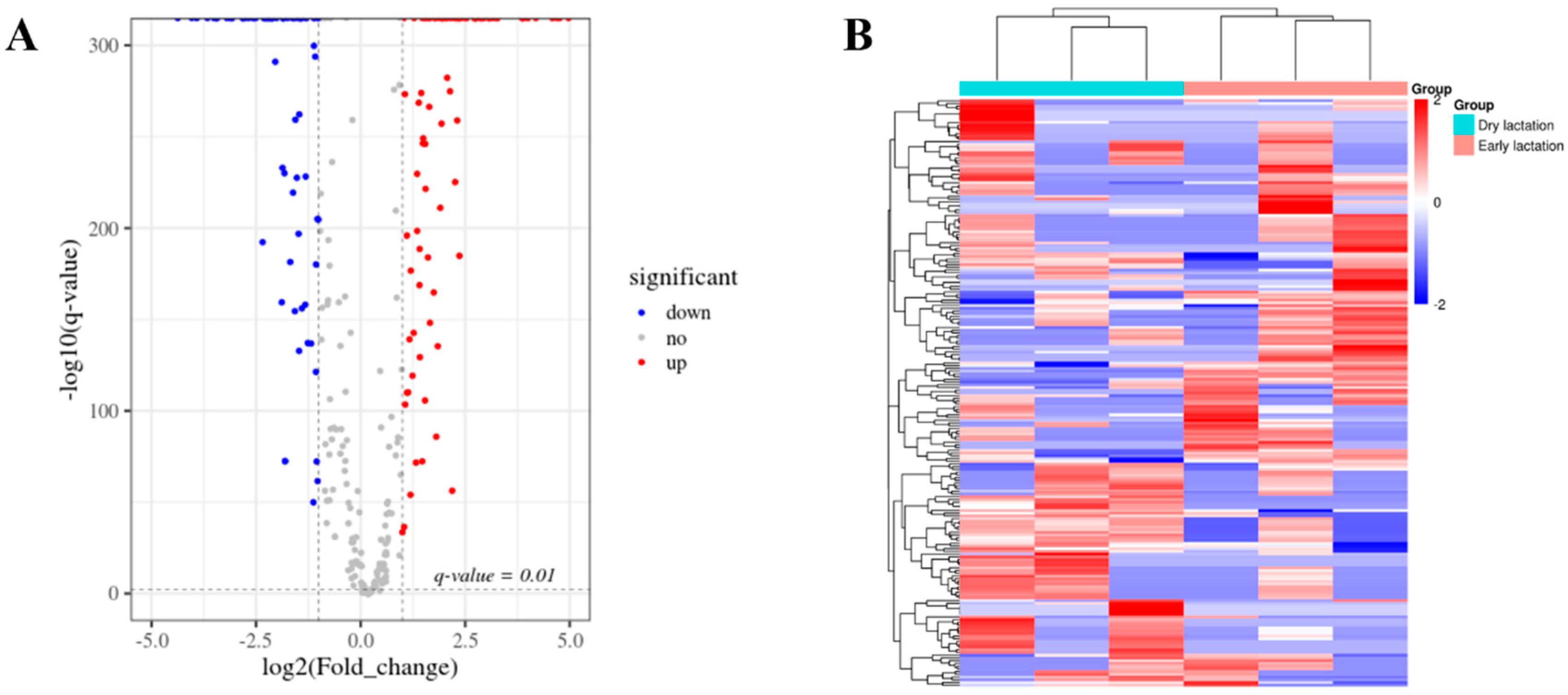
2.2. Differentially Expressed circRNAs between Stages of Lactation
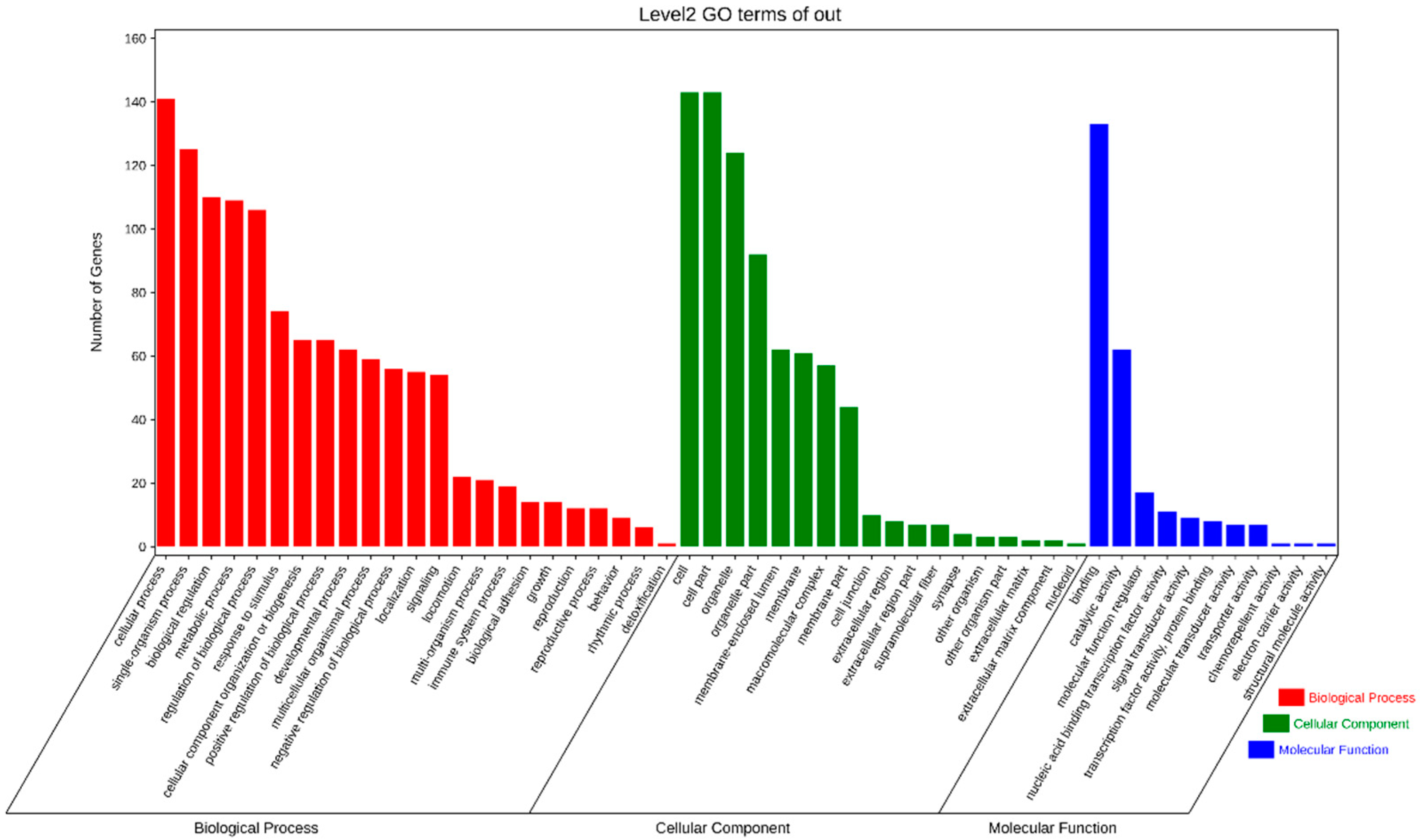
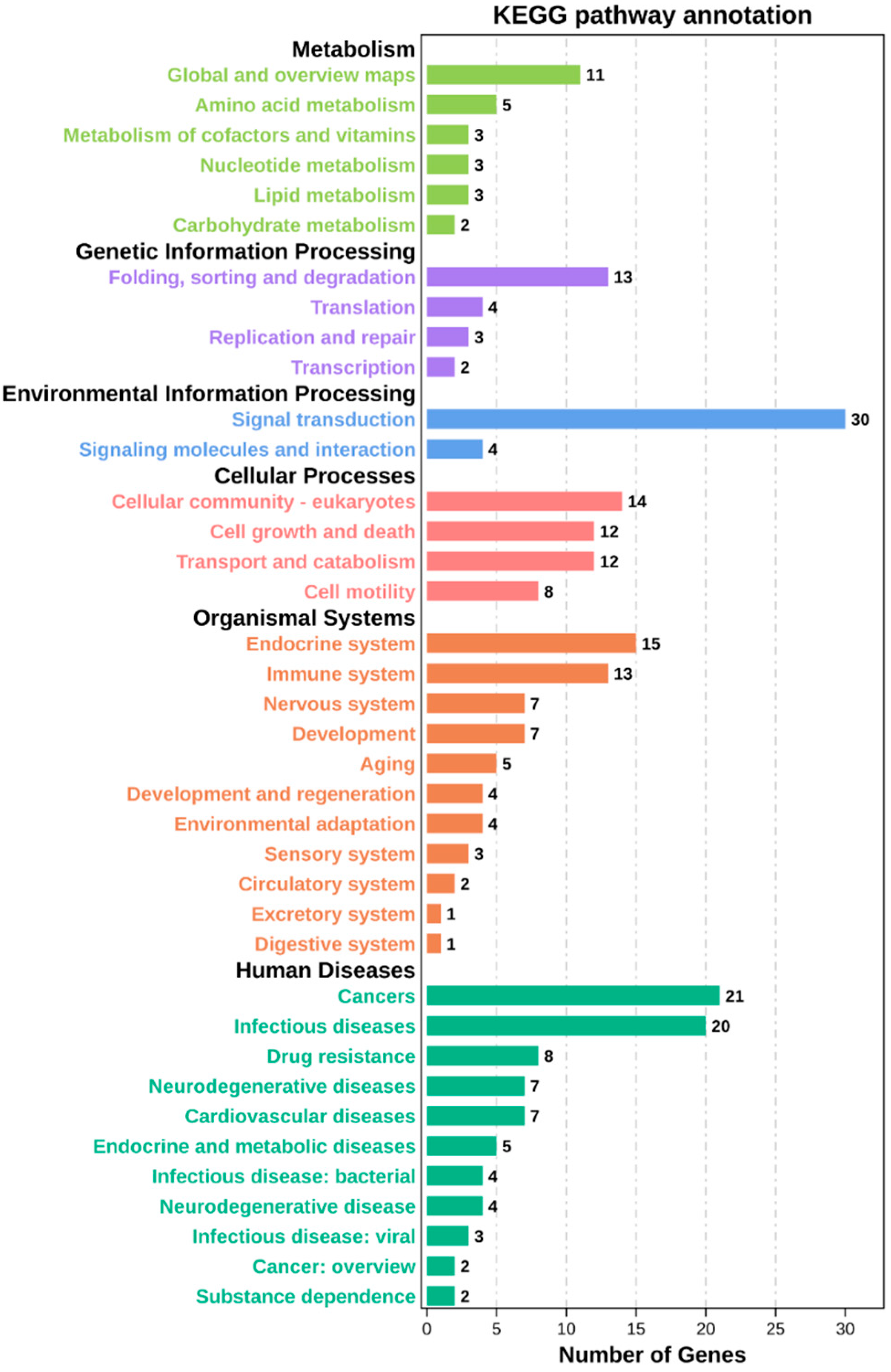
2.3. Enrichment Analysis
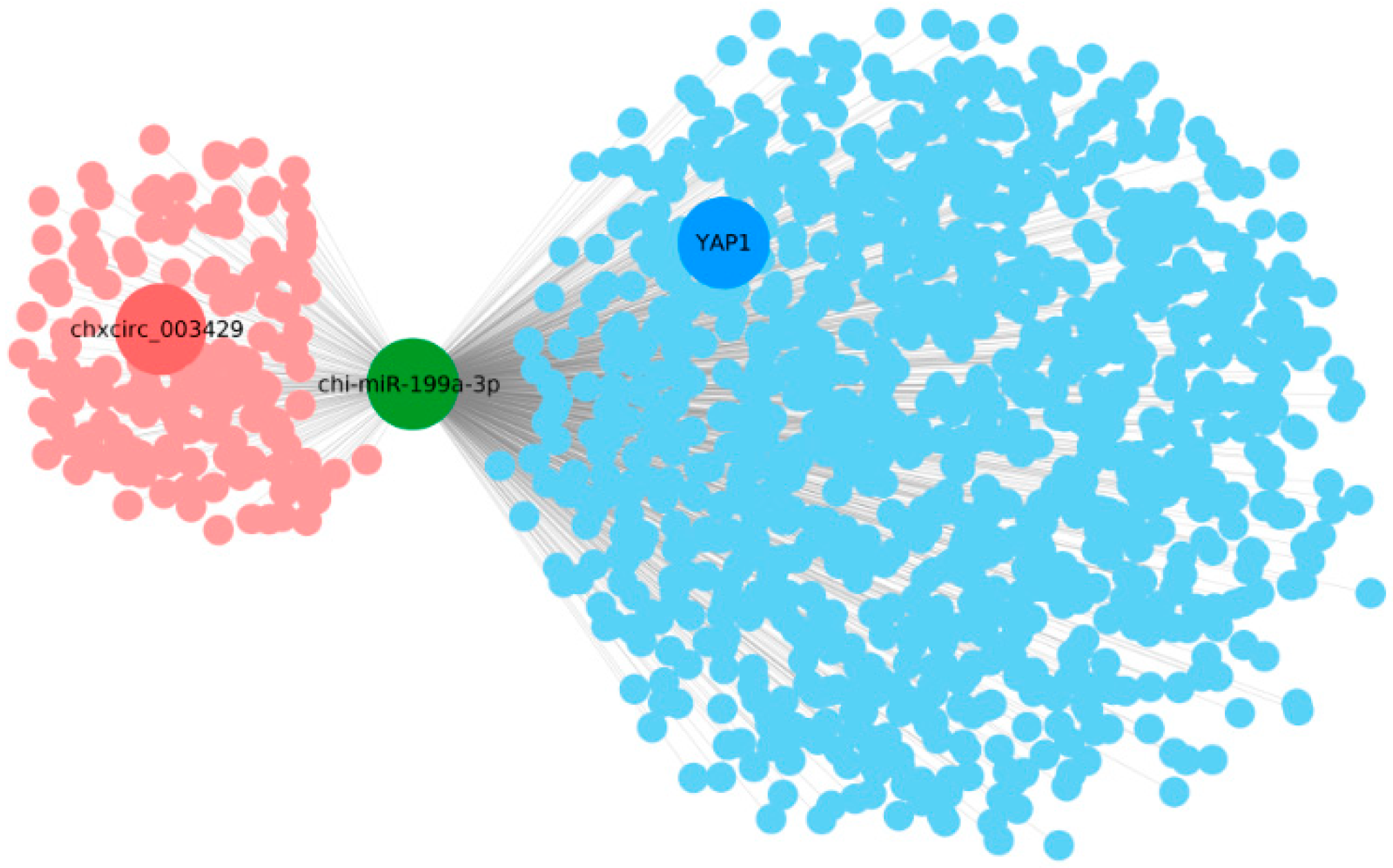
2.4. Analysis of the Interaction between circRNA and miRNA
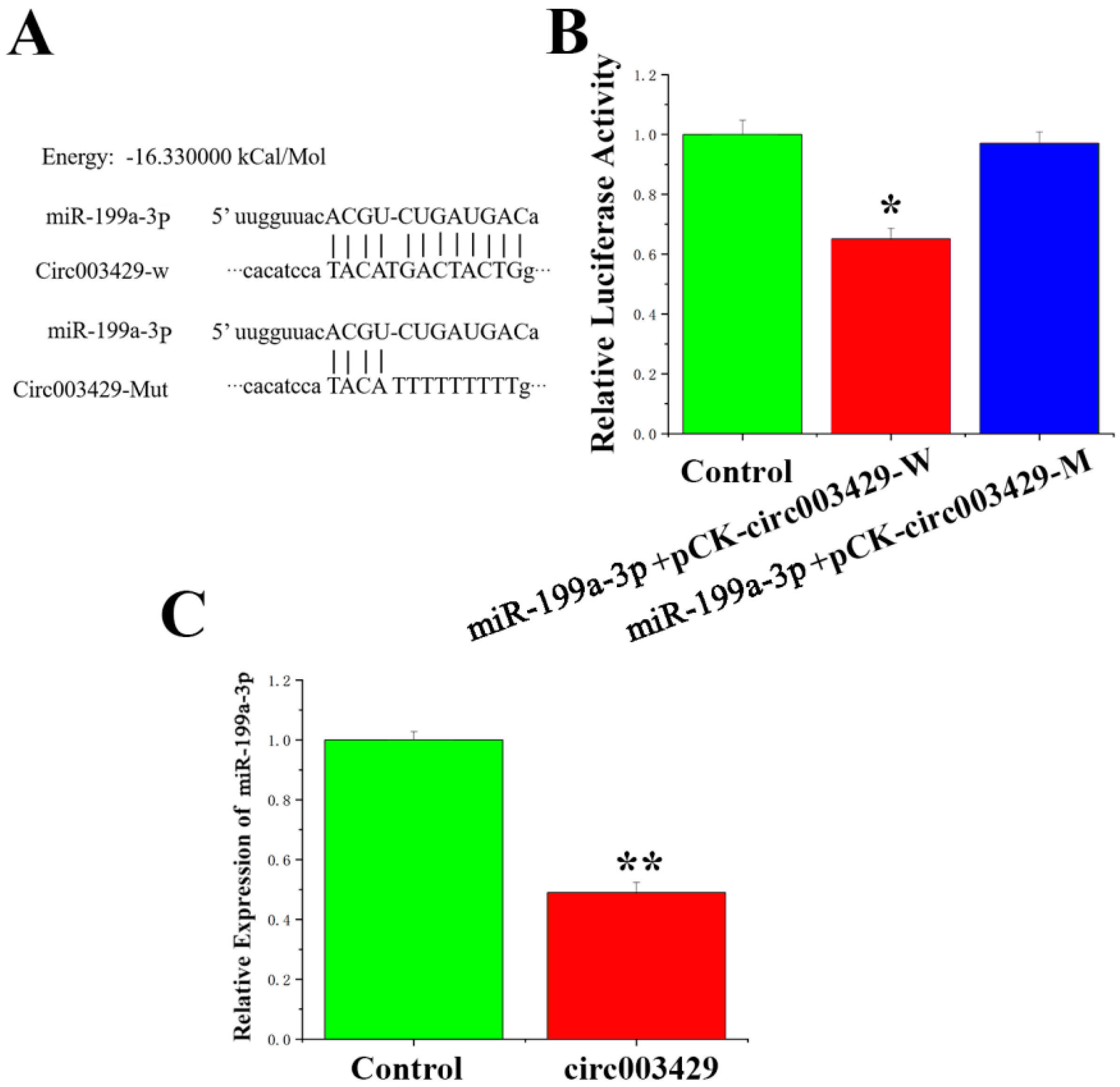
2.5. Circ003429 Adsorption and Binding of miR-199a-3p
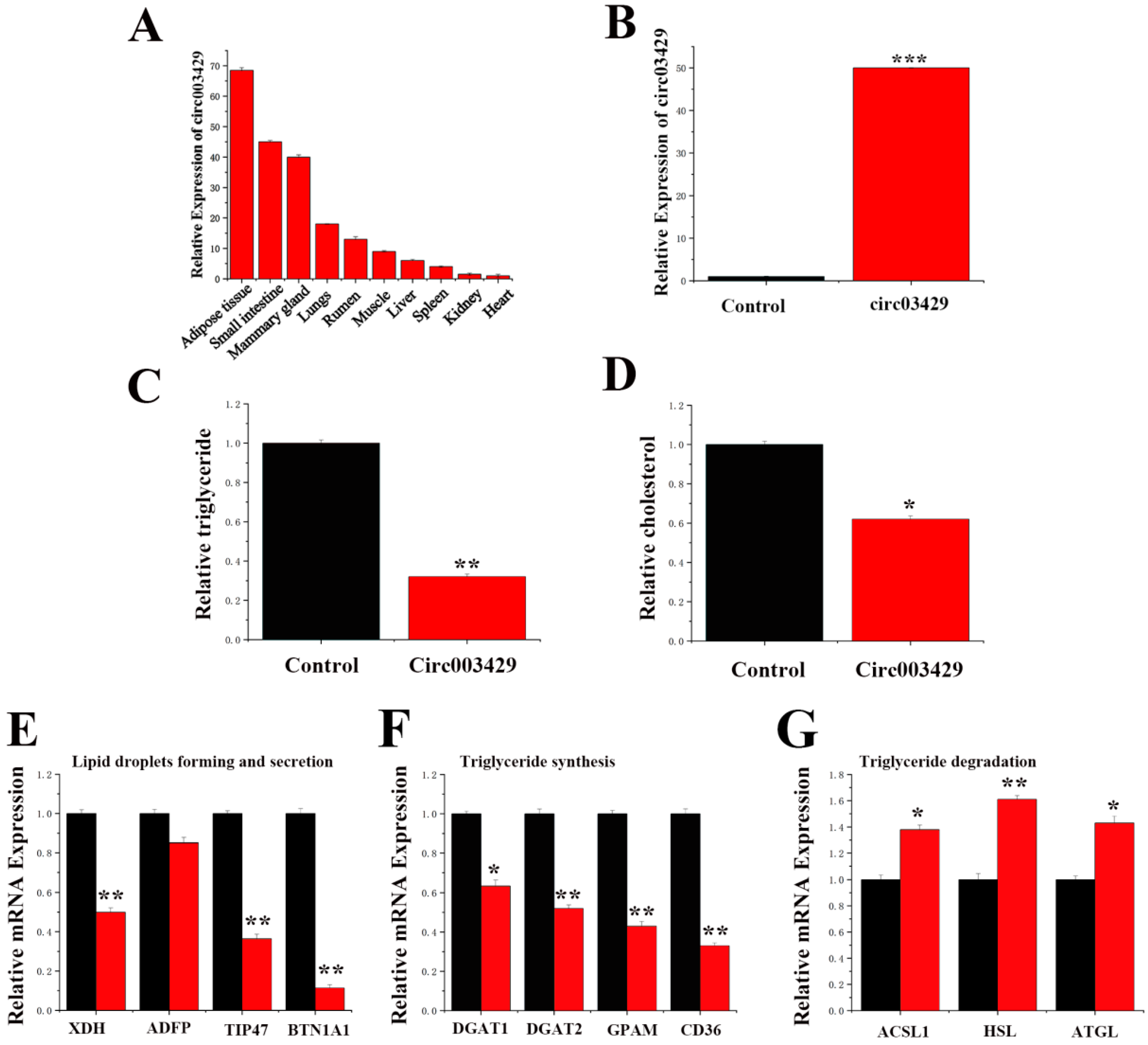
2.6. Functional Verification of circ003429 in GMECs
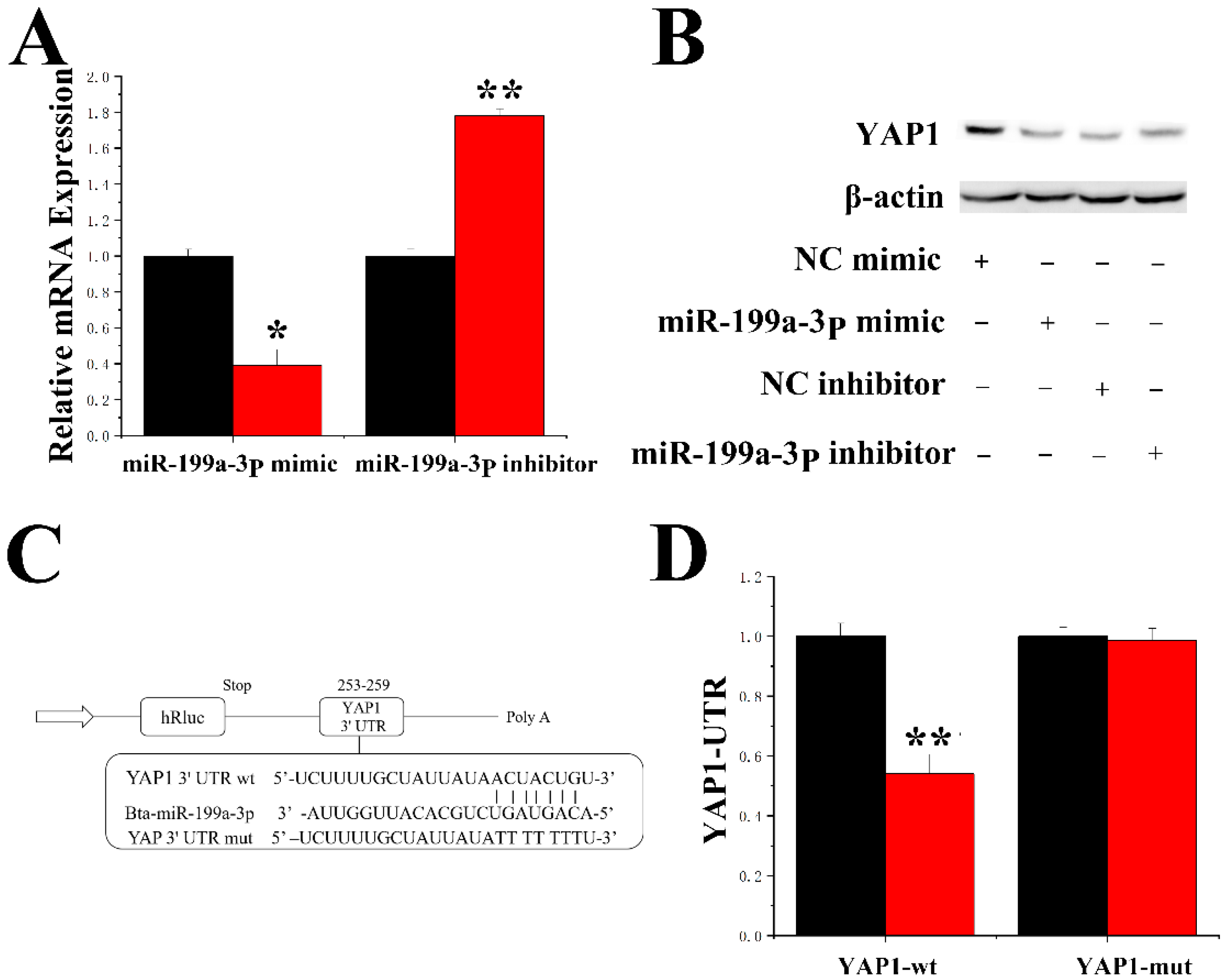
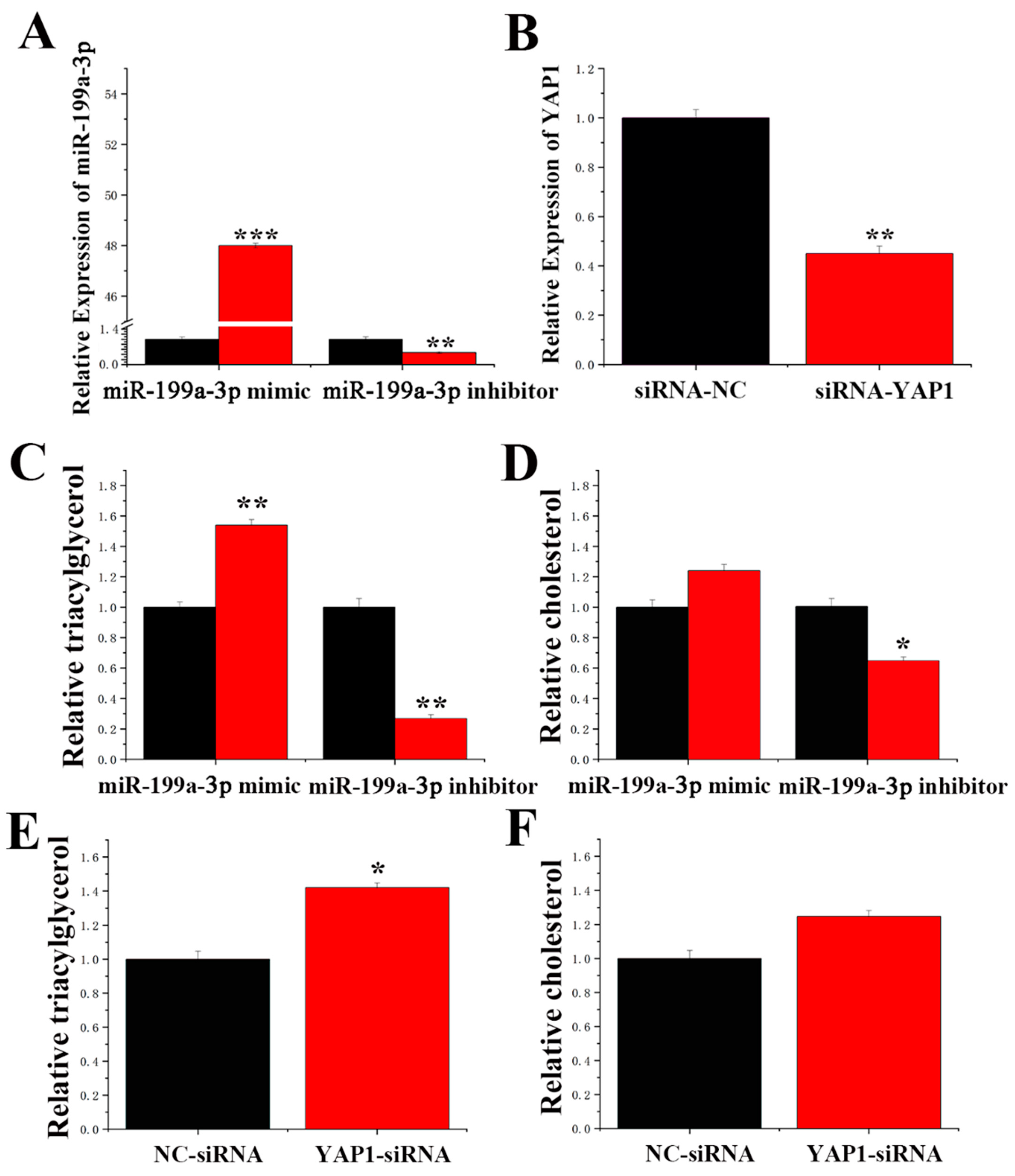
2.7. MiR-199a-3p Specifically Targets YAP1 in GMECs
2.8. Function of miR-199a-3p and YAP1 in GMECs
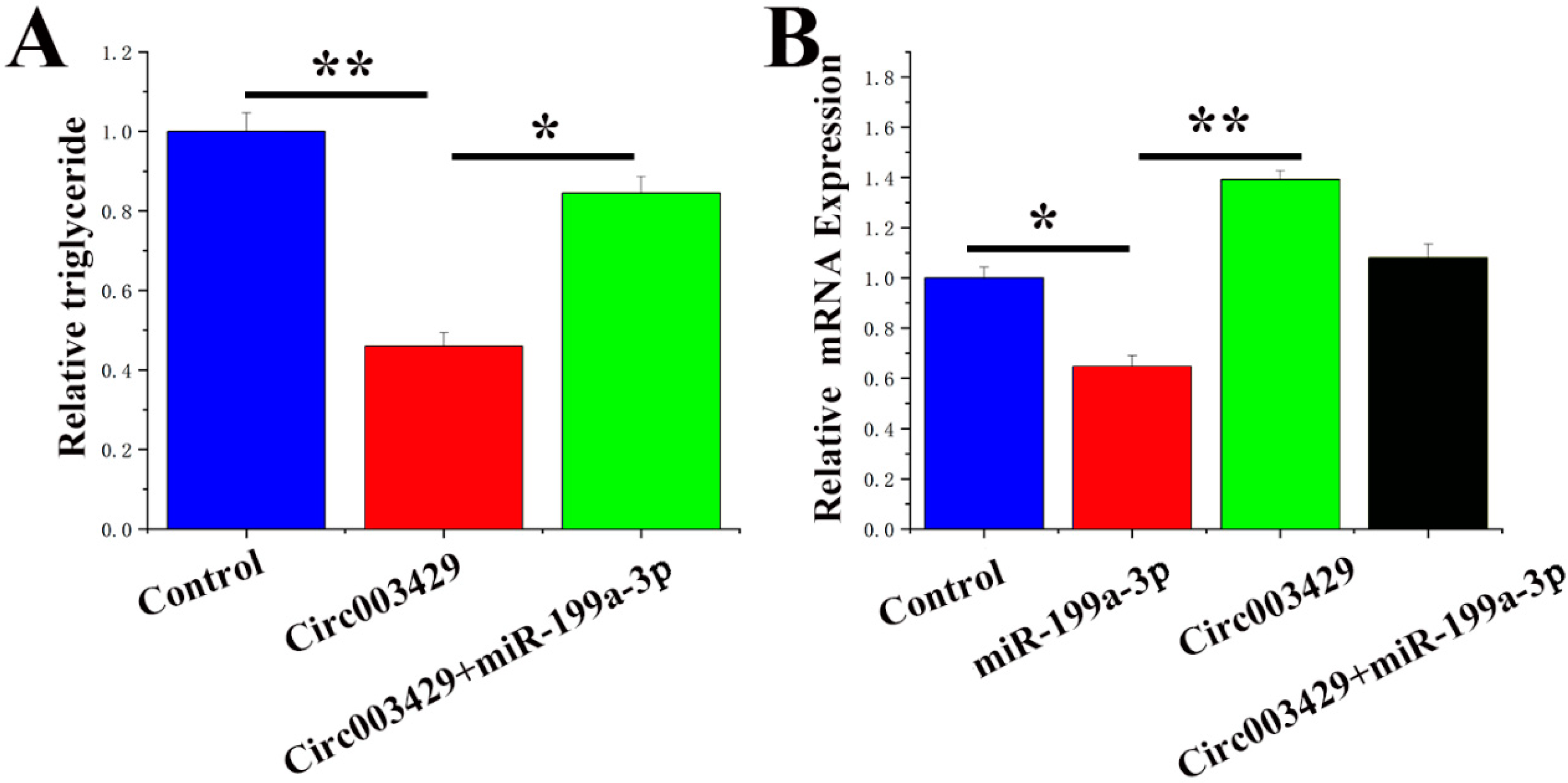
2.9. Circ003429 Combined with miR-199a-3p Relieves YAP1 Inhibition
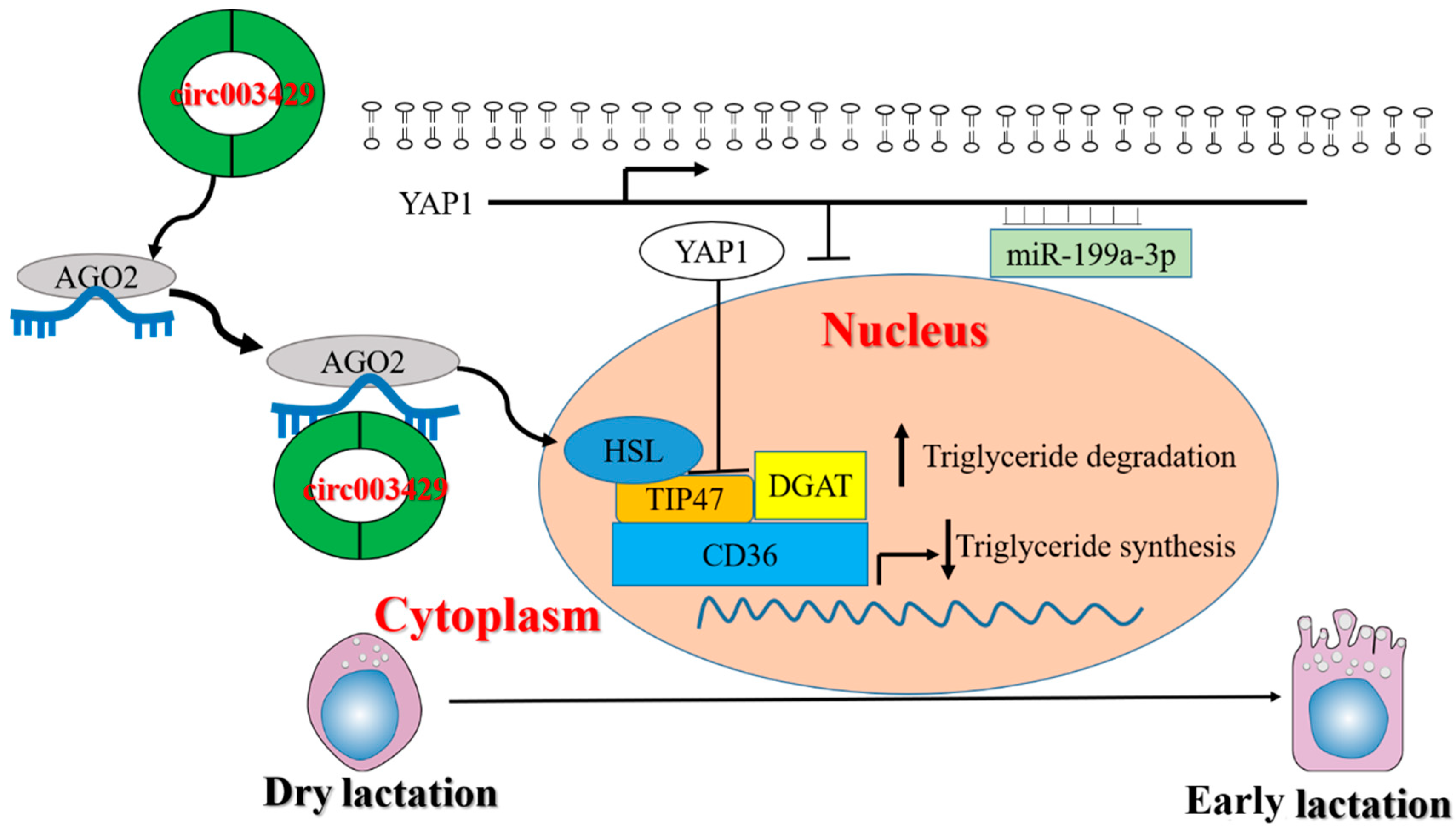
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Sequencing Library Construction and High-Throughput Sequencing
4.3. Acquisition of circRNAs
4.4. Expression Level Analysis
4.5. Analysis of Differential Expression of circRNAs
4.6. Analysis of circRNA and miRNA Interactions
4.7. Quantitative Real-Time Polymerase Chain Reaction (RT–qPCR)
4.8. Cell Culture
4.9. Construction of circRNA Expression Vectors
4.10. Detection of TAG and Cholesterol Contents
4.11. Western Blotting
4.12. Determination of Fatty Acid Content in GMECs
4.13. Analysis of Dual-Luciferase Activity
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rudolph, M.C.; McManaman, J.L.; Hunter, L.; Phang, T.; Neville, M.C. Functional development of the mammary gland: Use of expression profiling and trajectory clustering to reveal changes in gene expression during pregnancy, lactation, and involution. J. Mammary Gland Biol. 2003, 8, 287–307. [Google Scholar] [CrossRef] [PubMed]
- Kompan, D.; Komprej, A. The effect of fatty acids in goat milk on health. In Milk Production: An Up-to-Date Overview of Animal Nutrition, Management and Health; IntechOpen: London, UK, 2012. [Google Scholar] [CrossRef] [Green Version]
- Mather, I.H.; Keenan, T.W. Origin and secretion of milk lipids. J. Mammary Gland Biol. 1998, 3, 259–273. [Google Scholar] [CrossRef] [PubMed]
- Chong, B.M.; Reigan, P.; Mayle-Combs, K.D.; Orlicky, D.J.; McManaman, J.L. Determinants of adipophilin function in milk lipid formation and secretion. Trends Endocrinol. Metab. 2011, 22, 211–217. [Google Scholar] [CrossRef] [Green Version]
- Clegg, R.A.; Barber, M.C.; Pooley, L.; Ernens, I.; Larondelle, Y.; Travers, M.T. Milk fat synthesis and secretion: Molecular and cellular aspects. Livest. Prod. Sci. 2001, 70, 3–14. [Google Scholar] [CrossRef]
- Kang, W.; Tong, J.H.; Lung, R.W.; Dong, Y.; Zhao, J.; Liang, Q.; Zhang, L.; Pan, Y.; Yang, W.; Pang, J.C.; et al. Targeting of YAP1 by microRNA-15a and microRNA-16-1 exerts tumor suppressor function in gastric adenocarcinoma. Mol. Cancer 2015, 14, 52. [Google Scholar] [CrossRef] [Green Version]
- Verdelli, C.; Morotti, A.; Tavanti, G.S.; Silipigni, R.; Guerneri, S.; Ferrero, S.; Vicentini, L.; Vaira, V.; Corbetta, S. The core stem genes SOX2, POU5F1/OCT4, and NANOG are expressed in human parathyroid tumors and modulated by MEN1, YAP1, and beta-catenin pathways activation. Biomedicines 2021, 9, 637. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Shi, H.; Sun, S.; Xu, H.; Cao, D.; Luo, J. MicroRNA-181b suppresses TAG via target IRS2 and regulating multiple genes in the Hippo pathway. Exp. Cell Res. 2016, 348, 66–74. [Google Scholar] [CrossRef]
- Tudek, A.; Krawczyk, P.S.; Mroczek, S.; Tomecki, R.; Turtola, M.; Matylla-Kulinska, K.; Jensen, T.H.; Dziembowski, A. Global view on the metabolism of RNA poly(A) tails in yeast Saccharomyces cerevisiae. Nat. Commun. 2021, 12, 4951. [Google Scholar] [CrossRef]
- Xu, T.; Xie, M.; Jing, X.; Cui, J.; Wu, X.; Shu, Y. Crosstalk between environmental inflammatory stimuli and non-coding RNA in cancer occurrence and development. Cancers 2021, 13, 4436. [Google Scholar] [CrossRef]
- Cen, L.; Liu, R.; Liu, W.; Li, Q.; Cui, H. Competing endogenous RNA networks in glioma. Front. Genet. 2021, 12, 675498. [Google Scholar] [CrossRef]
- Choi, S.; Sathe, A.; Mathe, E.; Xing, C.; Pan, Z. Identification of a putative enhancer RNA for EGFR in hyper-accessible regions in esophageal squamous cell carcinoma cells by analysis of chromatin accessibility landscapes. Front. Oncol. 2021, 11, 724687. [Google Scholar] [CrossRef] [PubMed]
- Yue, B.; Wang, J.; Ru, W.; Wu, J.; Cao, X.; Yang, H.; Huang, Y.; Lan, X.; Lei, C.; Huang, B.; et al. The circular RNA circHUWE1 sponges the miR-29b-AKT3 axis to regulate myoblast development. Mol. Ther.-Nucleic Acids 2020, 19, 1086–1097. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Cao, X.; Lu, Q.; Zhou, J.; Wang, Y.; Wu, Y.; Mao, Y.; Xu, H.V.; Yang, Z. Circ01592 regulates unsaturated fatty acid metabolism through adsorbing miR-218 in bovine mammary epithelial cells. Food Funct. 2021, 12, 12047–12058. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Song, C.; Li, H.; Cao, X.; Ma, Y.; Wang, X.; Huang, Y.; Lan, X.; Lei, C.; Chaogetu, B.; et al. Circular RNA SNX29 sponges miR-744 to regulate proliferation and differentiation of myoblasts by activating the Wnt5a/Ca2+ signaling pathway. Mol. Ther.-Nucleic Acids 2019, 16, 481–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Liu, J.; Liu, S.; Jiao, W.; Wang, X. Mesenchymal stem cell-derived extracellular vesicles prevent the development of osteoarthritis via the circHIPK3/miR-124-3p/MYH9 axis. J. Nanobiotechnol. 2021, 19, 194. [Google Scholar] [CrossRef] [PubMed]
- Swerdlow, N.R.; Lipska, B.K.; Weinberger, D.R.; Braff, D.L.; Jaskiw, G.E.; Geyer, M.A. Increased sensitivity to the sensorimotor gating-disruptive effects of apomorphine after lesions of medial prefrontal cortex or ventral hippocampus in adult rats. Psychopharmacology 1995, 122, 27–34. [Google Scholar] [CrossRef]
- Justice, R.W.; Zilian, O.; Woods, D.F.; Noll, M.; Bryant, P.J. The Drosophila tumor suppressor gene warts encodes a homolog of human myotonic dystrophy kinase and is required for the control of cell shape and proliferation. Gene Dev. 1995, 9, 534–546. [Google Scholar] [CrossRef] [Green Version]
- Dong, Z.; Haines, S.; Coates, D. Proteomic profiling of stem cell tissues during regeneration of deer antler: A model of mammalian organ regeneration. J. Proteome Res. 2020, 19, 1760–1775. [Google Scholar] [CrossRef]
- Yu, F.; Jiang, R.; Han, W.; Zhan, G.; Xu, X.; Jiang, X.; Wang, L.; Xiang, S.; Zhou, Q.; Liu, C.; et al. Gut microbiota transplantation from db/db mice induces diabetes-like phenotypes and alterations in Hippo signaling in pseudo germ-free mice. Aging 2020, 12, 24156–24167. [Google Scholar] [CrossRef]
- Fabretti, F.; Tschernoster, N.; Erger, F.; Hedergott, A.; Buescher, A.K.; Dafinger, C.; Reusch, B.; Kontges, V.K.; Kohl, S.; Bartram, M.P.; et al. Expanding the spectrum of FAT1 nephropathies by novel mutations that affect Hippo Signaling. Kidney Int. Rep. 2021, 6, 1368–1378. [Google Scholar] [CrossRef]
- Deng, K.; Ren, C.; Fan, Y.; Pang, J.; Zhang, G.; Zhang, Y.; You, P.; Wang, F. YAP1 regulates PPARG and RXR alpha expression to affect the proliferation and differentiation of ovine preadipocyte. J. Cell. Biochem. 2019, 120, 19578–19589. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Chen, J.H.; Oh, H.R.; Park, J.H. Transcription factor SOX2 contributes to nonalcoholic fatty liver disease development by regulating the expression of the fatty acid transporter CD36. FEBS Lett. 2021, 595, 2493–2503. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Wang, Z.; Li, Y.; Ma, L.; Jin, J.; Scott, A.W.; Xu, Y.; Estrella, J.S.; Song, Y.; Liu, B.; et al. PPARδ interacts with the Hippo coactivator YAP1 to promote SOX9 expression and gastric cancer progression. Mol. Cancer Res. 2020, 18, 390–402. [Google Scholar] [CrossRef] [PubMed]
- Rudolf, M.A.; Andreeva, A.; Kozlowski, M.M.; Kim, C.E.; Moskowitz, B.A.; Anaya-Rocha, A.; Kelley, M.W.; Corwin, J.T. YAP mediates hair cell regeneration in balance organs of chickens, but LATS kinases suppress its activity in mice. J. Neurosci. 2020, 40, 3915–3932. [Google Scholar] [CrossRef]
- Chen, H.C.; Zhu, Y.T.; Chen, S.Y.; Tseng, S.C. Wnt signaling induces epithelial-mesenchymal transition with proliferation in ARPE-19 cells upon loss of contact inhibition. Lab. Investig. 2012, 92, 676–687. [Google Scholar] [CrossRef]
- Fu, J.; Zheng, M.; Zhang, X.; Zhang, Y.; Chen, Y.; Li, H.; Wang, X.; Zhang, J. Fibulin-5 promotes airway smooth muscle cell proliferation and migration via modulating Hippo-YAP/TAZ pathway. Biochem. Biophys. Res. Commun. 2017, 493, 985–991. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.H.; Lan, G.T.; Zhou, Y.; Qu, T.H.; Xiong, Q. Effect of yes associated protein on osteogenic/adipogenic differentiation of bone marrow mesenchymal stem cells under oxidative stress. J. Biomater. Tissue Eng. 2021, 11, 1636–1642. [Google Scholar] [CrossRef]
- Guo, F.S.; Xu, S.S.; Zhu, Y.L.; Zheng, X.; Lu, Y.; Tu, J.; He, Y.; Jin, L.H.; Li, Y. PPAR gamma transcription deficiency exacerbates high-fat diet-induced adipocyte hypertrophy and insulin resistance in mice. Front. Pharmacol. 2020, 11, 1285. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Luo, J.; Zhu, J.; Li, J.; Sun, Y.; Lin, X.; Zhang, L.; Yao, D.; Shi, H. PPAR gamma regulates genes involved in triacylglycerol synthesis and secretion in mammary gland epithelial cells of dairy goats. PPAR Res. 2013, 2013, 310948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.B.; Luo, J.; Yao, D.W.; Zhu, J.J.; Xu, H.F.; Shi, H.P.; Loor, J.J. Peroxisome proliferator-activated receptor-Y stimulates the synthesis of monounsaturated fatty acids in dairy goat mammary epithelial cells via the control of stearoyl-coenzyme A desaturase. J. Dairy Sci. 2013, 96, 7844–7853. [Google Scholar] [CrossRef] [Green Version]
- Rosset, C.; Vieira, I.A.; Salzano, F.M.; Bandinelli, E. A germline variant affects putative miRNA-binding sites at the F8 3’UTR and acts as a potential haemophilia A phenotype modifier in Southern Brazilian patients. Haemophilia 2016, 22, e327–e329. [Google Scholar] [CrossRef] [PubMed]
- Rowley, J.W.; Chappaz, S.; Corduan, A.; Chong, M.M.; Campbell, R.; Khoury, A.; Manne, B.K.; Wurtzel, J.G.; Michael, J.V.; Goldfinger, L.E.; et al. Dicer1-mediated miRNA processing shapes the mRNA profile and function of murine platelets. Blood 2016, 127, 1743–1751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, C.A.; Haas, J.T.; Streeper, R.S.; Stone, S.J.; Kumari, M.; Yang, K.; Han, X.; Brownell, N.; Gross, R.W.; Zechner, R.; et al. DGAT enzymes are required for triacylglycerol synthesis and lipid droplets in adipocytes. J. Lipid Res. 2011, 52, 657–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganji, S.H.; Tavintharan, S.; Zhu, D.; Xing, Y.; Kamanna, V.S.; Kashyap, M.L. Niacin noncompetitively inhibits DGAT2 but not DGAT1 activity in HepG2 cells. J. Lipid Res. 2004, 45, 1835–1845. [Google Scholar] [CrossRef] [Green Version]
- Cases, S.; Stone, S.J.; Zhou, P.; Yen, E.; Tow, B.; Lardizabal, K.D.; Voelker, T.; Farese, R.V., Jr. Cloning of DGAT2, a second mammalian diacylglycerol acyltransferase, and related family members. J. Biol. Chem. 2001, 276, 38870–38876. [Google Scholar] [CrossRef] [Green Version]
- Buhman, K.K.; Smith, S.J.; Stone, S.J.; Repa, J.J.; Wong, J.S.; Knapp, F.F., Jr.; Burri, B.J.; Hamilton, R.L.; Abumrad, N.A.; Farese, R.V., Jr. DGAT1 is not essential for intestinal triacylglycerol absorption or chylomicron synthesis. J. Biol. Chem. 2002, 277, 25474–25479. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.J.; Cases, S.; Jensen, D.R.; Chen, H.C.; Sande, E.; Tow, B.; Sanan, D.A.; Raber, J.; Eckel, R.H.; Farese, R.V., Jr. Obesity resistance and multiple mechanisms of triglyceride synthesis in mice lacking Dgat. Nat. Genet. 2000, 25, 87–90. [Google Scholar] [CrossRef]
- Meegalla, R.L.; Billheimer, J.T.; Cheng, D. Concerted elevation of acyl-coenzyme A: Diacylglycerol acyltransferase (DGAT) activity through independent stimulation of mRNA expression of DGAT1 and DGAT2 by carbohydrate and insulin. Biochem. Biophys. Res. Commun. 2002, 298, 317–323. [Google Scholar] [CrossRef]
- Kolakofsky, D. Isolation and characterization of Sendai virus DI-RNAs. Cell 1976, 8, 547–555. [Google Scholar] [CrossRef]
- Hsu, M.T.; Coca-Prados, M. Electron microscopic evidence for the circular form of RNA in the cytoplasm of eukaryotic cells. Nature 1979, 280, 339–340. [Google Scholar] [CrossRef]
- Wang, T.; Meng, S.; Chen, P.; Wei, L.; Liu, C.; Tang, D.; Liu, D.; Jiang, Z.; Hong, X. Comprehensive analysis of differentially expressed mRNA and circRNA in Ankylosing spondylitis patients’ platelets. Exp. Cell Res. 2021, 409, 112895. [Google Scholar] [CrossRef] [PubMed]
- D’Ambrosi, S.; Visser, A.; Antunes-Ferreira, M.; Poutsma, A.; Giannoukakos, S.; Sol, N.; Sabrkhany, S.; Bahce, I.; Kuijpers, M.J.E.; Oude Egbrink, M.G.A.; et al. The Analysis of platelet-derived circRNA repertoire as potential diagnostic biomarker for non-small cell lung cancer. Cancers 2021, 13, 4644. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Chen, Z.; Zhuang, X.; Luo, J.; Chen, T.; Xi, Q.; Zhang, Y.; Sun, J. Identification of circRNA-associated-ceRNA networks involved in milk fat metabolism under heat stress. Int. J. Mol. Sci. 2020, 21, 4162. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wu, H.; Wang, Y.; Zhao, Y.; Fang, X.; Chen, C.; Chen, H. Expression patterns of circular RNAs from primary kinase transcripts in the mammary glands of lactating rats. J. Breast Cancer 2015, 18, 235–241. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Wu, H.; Wang, Y.; Zhu, S.; Liu, J.; Fang, X.; Chen, H. Circular RNA of cattle casein genes are highly expressed in bovine mammary gland. J. Dairy Sci. 2016, 99, 4750–4760. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Zhou, J.; Wang, M.; Liu, J.; Zhang, L.; Loor, J.J.; Liang, Y.; Wu, H.; Yang, Z. Circ09863 regulates unsaturated fatty acid metabolism by adsorbing miR-27a-3p in bovine mammary epithelial cells. J. Agric. Food Chem. 2020, 68, 8589–8601. [Google Scholar] [CrossRef]
- Chen, Z.; Lu, Q.; Liang, Y.; Cui, X.; Wang, X.; Mao, Y.; Yang, Z. Circ11103 interacts with miR-128/PPARGC1A to regulate milk fat metabolism in dairy cows. J. Agric. Food Chem. 2021, 69, 4490–4500. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Read No. | Bases (bp) | Clean_Read No. | Clean Data (bp) | Q30 | Q30 (%) |
|---|---|---|---|---|---|---|
| EL1 | 112,447,802 | 16,867,170,300 | 103,061,092 | 15,459,163,800 | 15,705,576,121 | 93.11 |
| EL2 | 119,900,238 | 17,985,035,700 | 102,779,038 | 15,416,855,700 | 17,048,097,687 | 94.79 |
| EL3 | 135,189,948 | 20,278,492,200 | 101,520,494 | 15,228,074,100 | 19,299,211,421 | 95.17 |
| DL1 | 126,893,190 | 19,033,978,500 | 108,906,480 | 16,335,972,000 | 17,756,274,866 | 93.28 |
| DL2 | 124,554,188 | 18,683,128,200 | 104,354,388 | 15,653,158,200 | 17,736,653,054 | 94.93 |
| DL3 | 127,449,174 | 19,117,376,100 | 108,510,024 | 16,276,503,600 | 17,869,615,977 | 93.47 |
| Item | EL1 | EL2 | EL3 | DL1 | DL2 | DL3 |
|---|---|---|---|---|---|---|
| Clean_Reads | 103,061,092 | 102,779,038 | 101,520,494 | 108,906,480 | 104,354,388 | 108,510,024 |
| Total_Mapped (%) | 97.20 | 97.64 | 92.11 | 96.72 | 97.72 | 96.74 |
| Multiple_Mapped (%) | 2.09 | 1.92 | 3.12 | 4.13 | 1.54 | 3.69 |
| Uniquely_Mapped (%) | 97.91 | 98.08 | 96.88 | 95.87 | 98.46 | 96.31 |
| Map_Events | 98,081,017 | 98,425,684 | 90,599,563 | 100,986,515 | 100,406,806 | 101,099,167 |
| Mapped_to_Gene (%) | 73.50 | 72.42 | 76.07 | 74.13 | 72.81 | 74.46 |
| Mapped_to_InterGene (%) | 26.50 | 27.58 | 23.93 | 25.87 | 27.19 | 25.54 |
| Mapped_to_Exon (%) | 42.83 | 37.12 | 41.90 | 42.20 | 31.82 | 40.52 |
| Fatty Acid | NC | circ003429 |
|---|---|---|
| C16:0 (%) | 26.34 + 0.54 | 34.56 + 0.57 ** |
| C16:1 (%) | 18.34 + 0.65 | 5.47 + 1.038 ** |
| C18:0 (%) | 12.04 + 1.01 | 32.16 + 0.87 ** |
| C18:1 (%) | 34.71 + 1.54 | 25.36 + 1.67 * |
| C18:2 (%) | 8.57 + 0.38 | 2.45 + 0.97 * |
| SFAs (%) | 38.38 | 66.72 |
| UFAs (%) | 61.62 | 33.28 |
| UFAs/SFAs | 1.97 | 0.96 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiao, P.; Zhang, M.; Wang, Z.; Liang, G.; Xie, X.; Zhang, Y.; Chen, Z.; Jiang, Q.; Loor, J.J. Circ003429 Regulates Unsaturated Fatty Acid Synthesis in the Dairy Goat Mammary Gland by Interacting with miR-199a-3p, Targeting the YAP1 Gene. Int. J. Mol. Sci. 2022, 23, 4068. https://doi.org/10.3390/ijms23074068
Jiao P, Zhang M, Wang Z, Liang G, Xie X, Zhang Y, Chen Z, Jiang Q, Loor JJ. Circ003429 Regulates Unsaturated Fatty Acid Synthesis in the Dairy Goat Mammary Gland by Interacting with miR-199a-3p, Targeting the YAP1 Gene. International Journal of Molecular Sciences. 2022; 23(7):4068. https://doi.org/10.3390/ijms23074068
Chicago/Turabian StyleJiao, Peixin, Meimei Zhang, Ziwei Wang, Gege Liang, Xiaolai Xie, Yonggen Zhang, Zhi Chen, Qianming Jiang, and Juan J. Loor. 2022. "Circ003429 Regulates Unsaturated Fatty Acid Synthesis in the Dairy Goat Mammary Gland by Interacting with miR-199a-3p, Targeting the YAP1 Gene" International Journal of Molecular Sciences 23, no. 7: 4068. https://doi.org/10.3390/ijms23074068
APA StyleJiao, P., Zhang, M., Wang, Z., Liang, G., Xie, X., Zhang, Y., Chen, Z., Jiang, Q., & Loor, J. J. (2022). Circ003429 Regulates Unsaturated Fatty Acid Synthesis in the Dairy Goat Mammary Gland by Interacting with miR-199a-3p, Targeting the YAP1 Gene. International Journal of Molecular Sciences, 23(7), 4068. https://doi.org/10.3390/ijms23074068