
Early Life Events and Maturation of the Dentate Gyrus: Implications for Neurons and Glial Cells



{kind=link}
Abstract
:1. Introduction
2. The Structure of the DG in the Adult Brain
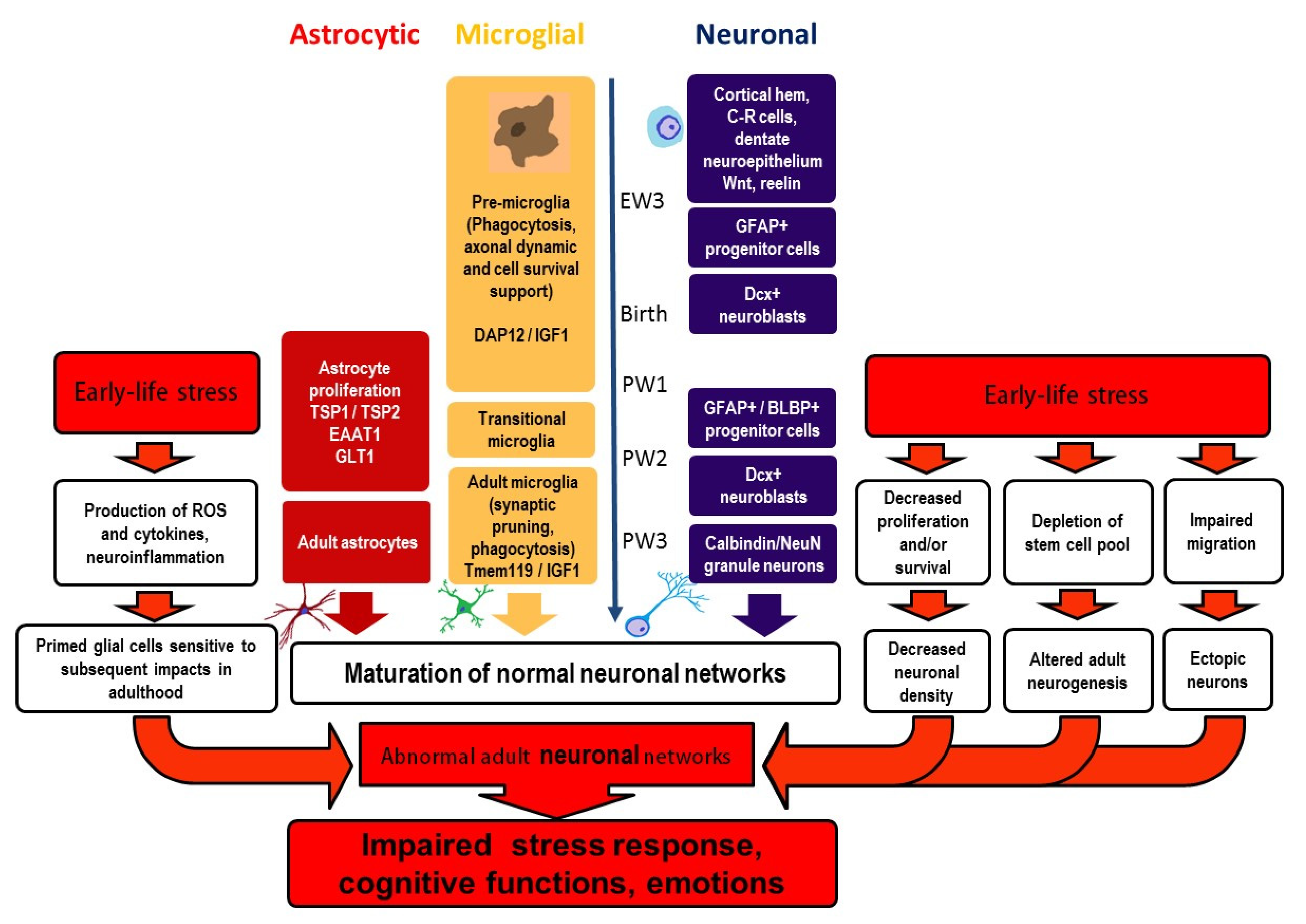
3. Pre- and Postnatal Development of the DG
4. Effects of Early Life Events on Neuronal Structure of the DG
4.1. Inflammation
4.2. Early Life Stress
4.3. Hormones
4.4. Hypoxia
4.5. Seizures
5. Maturation of Microglial and Astroglial Cells in the DG
5.1. Postnatal Maturation of Astroglia
5.2. Microglia Maturation in Early Ontogenesis
5.3. Involvement of Microglia in Neurogenesis in the DG during Ontogenesis
6. Responses of Microglial and Astroglial Cells to Aversive Events during Early Ontogenesis
6.1. Immediate Effects of Aversive Events in Early Ontogenesis on Glial Cells
6.2. Delayed Effects of Early Aversive Events on Glial Cells
7. Effects of Early and Late Aversive Events Combination on Glial
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carpenter, M.B. Chapter 18: Olfactory Pathways, Hippocampal Formation and Amygdala. In Human Neuroanatomy, 7th ed.; Williams & Wilkins: Baltimore, MD, USA, 1976; pp. 521–546. [Google Scholar]
- Amaral, D.G.; Witter, M.P. The three-dimensional organization of the hippocampal formation: A review of anatomical data. Neuroscience 1989, 31, 571–591. [Google Scholar] [CrossRef]
- Treves, A.; Tashiro, A.; Witter, M.P.; Moser, E.I. What is the mammalian dentate gyrus good for? Neuroscience 2008, 154, 1155–1172. [Google Scholar] [CrossRef] [PubMed]
- Gall, C. Comparative anatomy of the hippocampus: With special reference to differences in the distributions of neuroactive peptides. In Cerebral Cortex; Jones, E.G., Peters, A., Eds.; Plenum: New York, NY, USA, 1990; Volume 8B, pp. 167–213. [Google Scholar]
- Hodge, R.D.; Garcia, A.J., 3rd; Elsen, G.E.; Nelson, B.R.; Mussar, K.E.; Reiner, S.L.; Ramirez, J.M.; Hevner, R.F. Tbr2 expression in Cajal-Retzius cells and intermediate neuronal progenitors is required for morphogenesis of the dentate gyrus. J. Neurosci. 2013, 33, 4165–4180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, D.K.; Ming, G.L.; Song, H. Glial influences on neural stem cell development: Cellular niches for adult neurogenesis. Curr. Opin. Neurobiol. 2005, 15, 514–520. [Google Scholar] [CrossRef]
- Christian, K.M.; Song, H.; Ming, G.L. Functions and dysfunctions of adult hippocampal neurogenesis. Annu. Rev. Neurosci. 2014, 37, 243–262. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.X.; Marchetto, M.C.; Gage, F.H. How to make a hippocampal dentate gyrus granule neuron. Development 2014, 141, 2366–2375. [Google Scholar] [CrossRef] [Green Version]
- Altman, J.; Bayer, S.A. Mosaic organization of the hippocampal neuroepithelium and the multiple germinal sources of dentate granule cells. J. Comp. Neurol. 1990, 301, 325–342. [Google Scholar] [CrossRef]
- Aimone, J.B.; Li, Y.; Lee, S.W.; Clemenson, G.D.; Deng, W.; Gage, F.H. Regulation and function of adult neurogenesis: From genes to cognition. Physiol. Rev. 2014, 94, 991–1026. [Google Scholar] [CrossRef] [Green Version]
- Galceran, J.; Miyashita-Lin, E.M.; Devaney, E.; Rubenstein, J.L.; Grosschedl, R. Hippocampus development and generation of dentate gyrus granule cells is regulated by LEF1. Development 2000, 127, 469–482. [Google Scholar] [CrossRef]
- Rickmann, M.; Amaral, D.G.; Cowan, W.M. Organization of radial glial cells during the development of the rat dentate gyrus. J. Comp. Neurol. 1987, 264, 449–479. [Google Scholar] [CrossRef]
- Li, G.; Kataoka, H.; Coughlin, S.R.; Pleasure, S.J. Identification of a transient subpial neurogenic zone in the developing dentate gyrus and its regulation by Cxcl12 and reelin signaling. Development 2009, 136, 327–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Pleasure, S.J.; Collins, A.E.; Noebels, J.L.; Naya, F.J.; Tsai, M.J.; Lowenstein, D.H. Loss of BETA2/NeuroD leads to malformation of the dentate gyrus and epilepsy. Proc. Natl. Acad. Sci. USA 2000, 97, 865–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwano, T.; Masuda, A.; Kiyonari, H.; Enomoto, H.; Matsuzaki, F. Prox1 postmitotically defines dentate gyrus cells by specifying granule cell identity over CA3 pyramidal cell fate in the hippocampus. Development 2012, 139, 3051–3062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miquelajáuregui, A.; Varela-Echavarría, A.; Ceci, M.L.; García-Moreno, F.; Ricaño, I.; Hoang, K.; Frade-Pérez, D.; Portera-Cailliau, C.; Tamariz, E.; De Carlos, J.A.; et al. LIM-homeobox gene Lhx5 is required for normal development of Cajal-Retzius cells. J. Neurosci. 2010, 30, 10551–10562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muzio, L.; Mallamaci, A. Foxg1 confines Cajal-Retzius neuronogenesis and hippocampal morphogenesis to the dorsomedial pallium. J. Neurosci. 2005, 25, 4435–4441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanfield, B.B.; Cowan, W.M. The development of the hippocampus and dentate gyrus in normal and reeler mice. J. Comp. Neurol. 1979, 185, 423–459. [Google Scholar] [CrossRef]
- Newell, A.J.; Chung, S.H.; Wagner, C.K. Inhibition of progesterone receptor activity during development increases reelin-immunoreactivity in Cajal-Retzius cells, alters synaptic innervation in neonatal dentate gyrus, and impairs episodic-like memory in adulthood. Horm. Behav. 2021, 127, 104887. [Google Scholar] [CrossRef]
- Hevner, R.F.; Hodge, R.D.; Daza, R.A.; Englund, C. Transcription factors in glutamatergic neurogenesis: Conserved programs in neocortex, cerebellum, and adult hippocampus. Neurosci. Res. 2006, 55, 223–233. [Google Scholar] [CrossRef]
- Sugiyama, T.; Osumi, N.; Katsuyama, Y. The germinal matrices in the developing dentate gyrus are composed of neuronal progenitors at distinct differentiation stages. Dev. Dyn. 2013, 242, 1442–1453. [Google Scholar] [CrossRef]
- Seki, T.; Sato, T.; Toda, K.; Osumi, N.; Imura, T.; Shioda, S. Distinctive population of Gfap-expressing neural progenitors arising around the dentate notch migrate and form the granule cell layer in the developing hippocampus. J. Comp. Neurol. 2014, 522, 261–283. [Google Scholar] [CrossRef]
- Pleasure, S.J.; Anderson, S.; Hevner, R.; Bagri, A.; Marin, O.; Lowenstein, D.H.; Rubenstein, J.L. Cell migration from the ganglionic eminences is required for the development of hippocampal GABAergic interneurons. Neuron 2000, 28, 727–740. [Google Scholar] [CrossRef]
- McDonald, H.Y.; Wojtowicz, J.M. Dynamics of neurogenesis in the dentate gyrus of adult rats. Neurosci. Lett. 2005, 385, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Muramatsu, R.; Ikegaya, Y.; Matsuki, N.; Koyama, R. Neonatally born granule cells numerically dominate adult mice dentate gyrus. Neuroscience 2007, 148, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Arnold, S.E.; Trojanowski, J.Q. Human fetal hippocampal development: I. Cytoarchitecture, myeloarchitecture, and neuronal morphologic features. J. Comp. Neurol. 1996, 367, 274–292. [Google Scholar] [CrossRef]
- Holland, D.; Chang, L.; Ernst, T.M.; Curran, M.; Buchthal, S.D.; Alicata, D.; Skranes, J.; Johansen, H.; Hernandez, A.; Yamakawa, R.; et al. Structural growth trajectories and rates of change in the first 3 months of infant brain development. JAMA Neurol. 2014, 71, 1266–1274. [Google Scholar] [CrossRef]
- Matsue, K.; Minakawa, S.; Kashiwagi, T.; Toda, K.; Sato, T.; Shioda, S.; Seki, T. Dentate granule progenitor cell properties are rapidly altered soon after birth. Brain Struct. Funct. 2018, 223, 357–369. [Google Scholar] [CrossRef]
- Cameron, H.A.; Woolley, C.S.; McEwen, B.S.; Gould, E. Differentiation of newly born neurons and glia in the dentate gyrus of the adult rat. Neuroscience 1993, 56, 337–344. [Google Scholar] [CrossRef]
- Von Bohlen und Halbach, O. Immunohistological markers for staging neurogenesis in adult hippocampus. Cell Tissue Res. 2007, 329, 409–420. [Google Scholar] [CrossRef]
- Encinas, J.M.; Michurina, T.V.; Peunova, N.; Park, J.H.; Tordo, J.; Peterson, D.A.; Fishell, G.; Koulakov, A.; Enikolopov, G. Division-coupled astrocytic differentiation and age-related depletion of neural stem cells in the adult hippocampus. Cell Stem Cell 2011, 8, 566–579. [Google Scholar] [CrossRef] [Green Version]
- Deng, W.; Aimone, J.B.; Gage, F.H. New neurons and new memories: How does adult hippocampal neurogenesis affect learning and memory? Nat. Rev. Neurosci. 2010, 11, 339–350. [Google Scholar] [CrossRef]
- Green, H.F.; Nolan, Y.M. Inflammation and the developing brain: Consequences for hippocampal neurogenesis and behavior. Neurosci. Biobehav. Rev. 2014, 40, 20–34. [Google Scholar] [CrossRef] [PubMed]
- Mu, Y.; Gage, F.H. Adult hippocampal neurogenesis and its role in Alzheimer’s disease. Mol. Neurodegener. 2011, 6, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergami, M.; Berninger, B.; Canossa, M. Conditional deletion of TrkB alters adult hippocampal neurogenesis and anxiety-related behavior. Commun. Integr. Biol. 2009, 2, 14–16. [Google Scholar] [CrossRef] [PubMed]
- Revest, J.M.; Dupret, D.; Koehl, M.; Funk-Reiter, C.; Grosjean, N.; Piazza, P.V.; Abrous, D.N. Adult hippocampal neurogenesis is involved in anxiety-related behaviors. Mol. Psychiatry 2009, 14, 959–967. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.C.; Wang, S. Adult neurogenesis is reduced in the dorsal hippocampus of rats displaying learned helpless behavior. Neuroscience 2010, 171, 153–161. [Google Scholar] [CrossRef]
- Snyder, J.S.; Soumier, A.; Brewer, M.; Pickel, J.; Cameron, H.A. Adult hippocampal neurogenesis buffers stress responses and depressive behavior. Nature 2011, 476, 458–461. [Google Scholar] [CrossRef]
- Burd, I.; Bentz, A.I.; Chai, J.; Gonzalez, J.; Monnerie, H.; Le Roux, P.D.; Cohen, A.S.; Yudkoff, M.; Elovitz, M.A. Inflammation-induced preterm birth alters neuronal morphology in the mouse fetal brain. J. Neurosci. Res. 2010, 88, 1872–1881. [Google Scholar] [CrossRef] [Green Version]
- Elovitz, M.A.; Mrinalini, C.; Sammel, M.D. Elucidating the early signal transduction pathways leading to fetal brain injury in preterm birth. Pediatr. Res. 2006, 59, 50–55. [Google Scholar] [CrossRef] [Green Version]
- Hester, M.S.; Tulina, N.; Brown, A.; Barila, G.; Elovitz, M.A. Intrauterine inflammation reduces postnatal neurogenesis in the hippocampal subgranular zone and leads to accumulation of hilar ectopic granule cells. Brain Res. 2018, 1685, 51–59. [Google Scholar] [CrossRef]
- Lin, Y.L.; Wang, S. Prenatal lipopolysaccharide exposure increases depression-like behaviors and reduces hippocampal neurogenesis in adult rats. Behav. Brain Res. 2014, 259, 24–34. [Google Scholar] [CrossRef]
- Järlestedt, K.; Naylor, A.S.; Dean, J.; Hagberg, H.; Mallard, C. Decreased survival of newborn neurons in the dorsal hippocampus after neonatal LPS exposure in mice. Neuroscience 2013, 253, 21–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, P.L.; Hagberg, H.; Naylor, A.S.; Mallard, C. Neonatal peripheral immune challenge activates microglia and inhibits neurogenesis in the developing murine hippocampus. Dev. Neurosci. 2014, 36, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Pang, Y.; Dai, X.; Roller, A.; Carter, K.; Paul, I.; Bhatt, A.J.; Lin, R.C.; Fan, L.W. Early postnatal lipopolysaccharide exposure leads to enhanced neurogenesis and impaired communicative functions in rats. PLoS ONE 2016, 11, e0164403. [Google Scholar] [CrossRef] [Green Version]
- Domínguez-Rivas, E.; Ávila-Muñoz, E.; Schwarzacher, S.W.; Zepeda, A. Adult hippocampal neurogenesis in the context of lipopolysaccharide-induced neuroinflammation: A molecular, cellular and behavioral review. Brain Behav. Immun. 2021, 97, 286–302. [Google Scholar] [CrossRef]
- Girard, S.; Sébire, H.; Brochu, M.E.; Briota, S.; Sarret, P.; Sébire, G. Postnatal administration of IL-1Ra exerts neuroprotective effects following perinatal inflammation and/or hypoxic-ischemic injuries. Brain Behav. Immun. 2012, 26, 1331–1339. [Google Scholar] [CrossRef] [Green Version]
- Dinel, A.L.; Joffre, C.; Trifilieff, P.; Aubert, A.; Foury, A.; Le Ruyet, P.; Layé, S. Inflammation early in life is a vulnerability factor for emotional behavior at adolescence and for lipopolysaccharide-induced spatial memory and neurogenesis alteration at adulthood. J. Neuroinflammation 2014, 11, 155. [Google Scholar] [CrossRef]
- Veerasammy, S.; Van Steenwinckel, J.; Le Charpentier, T.; Seo, J.H.; Fleiss, B.; Gressens, P.; Levison, S.W. Perinatal IL-1β-induced inflammation suppresses Tbr2+ intermediate progenitor cell proliferation in the developing hippocampus accompanied by long-term behavioral deficits. Brain Behav. Immun. Health 2020, 7, 100106. [Google Scholar] [CrossRef]
- Correa, F.; Ljunggren, E.; Patil, J.; Wang, X.; Hagberg, H.; Mallard, C.; Sandberg, M. Time-dependent effects of systemic lipopolysaccharide injection on regulators of antioxidant defence Nrf2 and PGC-1α in the neonatal rat brain. Neuroimmunomodulation 2013, 20, 185–193. [Google Scholar] [CrossRef] [Green Version]
- da Silva, C.S.; Calió, M.L.; Mosini, A.C.; Pires, J.M.; Rêgo, D.D.S.B.; Mello, L.E.; Leslie, A.T.F.S. LPS-Induced systemic neonatal inflammation: Blockage of P2X7R by BBG decreases mortality on rat pups and oxidative stress in hippocampus of adult rats. Front. Behav. Neurosci. 2019, 13, 240. [Google Scholar] [CrossRef]
- Youssef, M.; Atsak, P.; Cardenas, J.; Kosmidis, S.; Leonardo, E.D.; Dranovsky, A. Early life stress delays hippocampal development and diminishes the adult stem cell pool in mice. Sci. Rep. 2019, 9, 4120. [Google Scholar] [CrossRef]
- Naninck, E.F.; Hoeijmakers, L.; Kakava-Georgiadou, N.; Meesters, A.; Lazic, S.E.; Lucassen, P.J.; Korosi, A. Chronic early life stress alters developmental and adult neurogenesis and impairs cognitive function in mice. Hippocampus 2015, 25, 309–328. [Google Scholar] [CrossRef] [PubMed]
- Suri, D.; Veenit, V.; Sarkar, A.; Thiagarajan, D.; Kumar, A.; Nestler, E.J.; Galande, S.; Vaidya, V.A. Early stress evokes age-dependent biphasic changes in hippocampal neurogenesis, BDNF expression, and cognition. Biol. Psychiatry 2013, 73, 658–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kight, K.E.; McCarthy, M.M. Androgens and the developing hippocampus. Biol. Sex Differ. 2020, 11, 30. [Google Scholar] [CrossRef]
- Wimer, R.E.; Wimer, C. Three sex dimorphisms in the granule cell layer of the hippocampus in house mice. Brain Res. 1985, 328, 105–109. [Google Scholar] [CrossRef]
- Zhang, J.M.; Konkle, A.T.; Zup, S.L.; McCarthy, M.M. Impact of sex and hormones on new cells in the developing rat hippocampus: A novel source of sex dimorphism? Eur. J. Neurosci. 2008, 27, 791–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowers, J.M.; Waddell, J.; McCarthy, M.M. A developmental sex difference in hippocampal neurogenesis is mediated by endogenous oestradiol. Biol. Sex Differ. 2010, 1, 8. [Google Scholar] [CrossRef] [Green Version]
- Waddell, J.; Bowers, J.; Edwards, N.; Jordan, C.; McCarthy, M. Dysregulation of neonatal hippocampal cell genesis in the androgen insensitive Tfm rat. Horm. Behav. 2013, 64, 144–152. [Google Scholar] [CrossRef] [Green Version]
- Roof, R.L.; Havens, M.D. Testosterone improves maze performance and induces development of a male hippocampus in females. Brain Res. 1992, 572, 310–313. [Google Scholar] [CrossRef]
- Quadros, P.S.; Pfau, J.L.; Wagner, C.K. Distribution of progesterone receptor immunoreactivity in the fetal and neonatal rat forebrain. J. Comp. Neurol. 2007, 56, 42–56. [Google Scholar] [CrossRef]
- Newell, A.J.; Lalitsasivimol, D.; Willing, J.; Gonzales, K.; Waters, E.M.; Milner, T.A.; McEwen, B.S.; Wagner, C.K. Progesterone receptor expression in cajal-retzius cells of the developing rat dentate gyrus: Potential role in hippocampus-dependent memory. J. Comp. Neurol. 2018, 526, 2285–2300. [Google Scholar] [CrossRef]
- Sanchez, R.M.; Koh, S.; Rio, C.; Wang, C.; Lamperti, E.D.; Sharma, D.; Corfas, G.; Jensen, F.E. Decreased glutamate receptor 2 expression and enhanced epileptogenesis in immature rat hippocampus after perinatal hypoxia-induced seizures. J. Neurosci. 2001, 21, 8154–8163. [Google Scholar] [CrossRef] [PubMed]
- Wolfart, J.; Laker, D. Homeostasis or channelopathy? Acquired cell type-specific ion channel changes in temporal lobe epilepsy and their antiepileptic potential. Front. Physiol. 2015, 6, 168. [Google Scholar] [CrossRef] [PubMed]
- Cilio, M.R.; Sogawa, Y.; Cha, B.H.; Liu, X.; Huang, L.T.; Holmes, G.L. Long-term effects of status epilepticus in the immature brain are specific for age and model. Epilepsia 2003, 44, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Nitecka, L.; Tremblay, E.; Charton, G.; Bouillot, J.P.; Berger, M.L.; Ben-Ari, Y. Maturation of kainic acid seizure-brain damage syndrome in the rat. II. Histopathological sequelae. Neuroscience 1984, 13, 1073–1094. [Google Scholar] [CrossRef]
- Holmes, G.L.; Gairsa, J.-L.; Chevassus-Au-Louis, N.; Ben-Ari, Y. Consequences of neonatal seizures in the rat: Morphological and behavioral effects. Ann. Neurol. 1998, 44, 845–857. [Google Scholar] [CrossRef]
- Porter, B.E.; Cui, X.N.; Brooks-Kayal, A.R. Status epilepticus differentially alters AMPA and kainate receptor subunit expression in mature and immature dentate granule neurons. Eur. J. Neurosci. 2006, 23, 2857–2863. [Google Scholar] [CrossRef]
- Bender, R.A.; Baram, T.Z. Epileptogenesis in the developing brain: What can we learn from animal models? Epilepsia 2007, 48, 2–6. [Google Scholar] [CrossRef] [Green Version]
- Schafer, D.P.; Stevens, B. Phagocytic glial cells: Sculpting synaptic circuits in the developing nervous system. Curr. Opin. Neurobiol. 2013, 23, 1034–1040. [Google Scholar] [CrossRef] [Green Version]
- Polazzi, E.; Contestabile, A. Reciprocal interactions between microglia and neurons: From survival to neuropathology. Rev. Neurosci. 2002, 13, 221–242. [Google Scholar] [CrossRef]
- Tremblay, M.-Ѐ.; Stevens, B.; Sierra, A.; Wake, H.; Bessis, A.; Nimmerjahn, A. The role of microglia in the healthy brain. J. Neurosci. 2011, 31, 16064–16069. [Google Scholar] [CrossRef]
- Tay, T.L.; Savage, J.C.; Hui, C.W.; Bisht, K.; Tremblay, M.-Ѐ. Microglia across the lifespan: From origin to function in brain development, plasticity and cognition. J. Physiol. 2017, 595, 1929–1945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paolicelli, R.C.; Bolasco, G.; Pagani, F.; Maggi, L.; Scianni, M.; Panzanelli, P.; Giustetto, M.; Ferreira, T.A.; Guiducci, E.; Dumas, L.; et al. Synaptic pruning by microglia is necessary for normal brain development. Science 2011, 333, 1456–1458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freeman, M.R. Specification and morphogenesis of astrocytes. Science 2010, 330, 774–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felix, L.; Stephan, J.; Rose, C.R. Astrocytes of the early postnatal brain. Eur. J. Neurosci. 2021, 54, 5649–5672. [Google Scholar] [CrossRef] [PubMed]
- Ge, W.P.; Miyawaki, A.; Gage, F.H.; Jan, Y.N.; Jan, L.Y. Local generation of glia is a major astrocyte source in postnatal cortex. Nature 2012, 484, 376–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelhardt, B. Development of the blood-brain barrier. Cell Tissue Res. 2003, 314, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Daneman, R.; Zhou, L.; Kebede, A.A.; Barres, B.A. Pericytes are required for blood-brain barrier integrity during embryogenesis. Nature 2010, 468, 562–566. [Google Scholar] [CrossRef] [Green Version]
- Christopherson, K.S.; Ullian, E.M.; Stokes, C.C.; Mullowney, C.E.; Hell, J.W.; Agah, A.; Lawler, J.; Mosher, D.F.; Bornstein, P.; Barres, B.A. Thrombospondins are astrocyte-secreted proteins that promote CNS synaptogenesis. Cell 2005, 120, 421–433. [Google Scholar] [CrossRef] [Green Version]
- Ullian, E.M.; Christopherson, K.S.; Barres, B.A. Role for glia in synaptogenesis. Glia 2004, 47, 209–216. [Google Scholar] [CrossRef]
- Regan, M.R.; Huang, Y.H.; Kim, Y.S.; Dykes-Hoberg, M.I.; Jin, L.; Watkins, A.M.; Bergles, D.E.; Rothstein, J.D. Variations in promoter activity reveal a differential expression and physiology of glutamate transporters by glia in the developing and mature CNS. J. Neurosci. 2007, 27, 6607–6619. [Google Scholar] [CrossRef] [Green Version]
- Schools, G.P.; Kimelberg, H.K. mGluR3 and mGluR5 are the predominant metabotropic glutamate receptor mRNAs expressed in hippocampal astrocytes acutely isolated from young rats. J. Neurosci. Res. 1999, 58, 533–543. [Google Scholar] [CrossRef]
- Morel, L.; Higashimori, H.; Tolman, M.; Yang, Y. VGluT1+ neuronal glutamatergic signaling regulates postnatal developmental maturation of cortical protoplasmic astroglia. J. Neurosci. 2014, 34, 10950–10962. [Google Scholar] [CrossRef] [PubMed]
- Clarke, L.E.; Barres, B.A. Emerging roles of astrocytes in neural circuit development. Nat. Rev. Neurosci. 2013, 14, 311–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, T.; Vukelic, J.; Hertzberg, E.L.; Nagy, J.I. Differential anatomical and cellular patterns of connexin43 expression during postnatal development of rat brain. Brain Res. Dev. Brain Res. 1992, 66, 165–180. [Google Scholar] [CrossRef]
- Bushong, E.A.; Martone, M.E.; Ellisman, M.H. Maturation of astrocyte morphology and the establishment of astrocyte domains during postnatal hippocampal development. Int. J. Dev. Neurosci. 2004, 22, 73–86. [Google Scholar] [CrossRef]
- Amateau, S.K.; McCarthy, M.M. Sexual differentiation of astrocyte morphology in the developing rat preoptic area. J. Neuroendocrinol. 2002, 14, 904–910. [Google Scholar] [CrossRef]
- Mong, J.A.; Glaser, E.; McCarthy, M.M. Gonadal steroids promote glial differentiation and alter neuronal morphology in the developing hypothalamus in a regionally specific manner. J. Neurosci. 1999, 19, 1464–1472. [Google Scholar] [CrossRef]
- Conejo, N.M.; González-Pardo, H.; Cimadevilla, J.M.; Argüelles, J.A.; Díaz, F.; Vallejo-Seco, G.; Arias, J.L. Influence of gonadal steroids on the glial fibrillary acidic protein-immunoreactive astrocyte population in young rat hippocampus. J. Neurosci. Res. 2005, 79, 488–494. [Google Scholar] [CrossRef]
- Bennett, M.L.; Bennett, F.C.; Liddelow, S.A.; Ajami, B.; Zamanian, J.L.; Fernhoff, N.B.; Mulinyawe, S.B.; Bohlen, C.J.; Adil, A.; Tucker, A.; et al. New tools for studying microglia in the mouse and human CNS. Proc. Natl. Acad. Sci. USA 2016, 113, E1738–E1746. [Google Scholar] [CrossRef] [Green Version]
- Matcovitch-Natan, O.; Winter, D.R.; Giladi, A.; Vargas Aguilar, S.; Spinrad, A.; Sarrazin, S.; Ben-Yehuda, H.; David, E.; Zelada González, F.; Perrin, P.; et al. Microglia development follows a stepwise program to regulate brain homeostasis. Science 2016, 353, aad8670. [Google Scholar] [CrossRef]
- Pont-Lezica, L.; Beumer, W.; Colasse, S.; Drexhage, H.; Versnel, M.; Bessis, A. Microglia shape corpus callosum axon tract fasciculation: Functional impact of prenatal inflammation. Eur. J. Neurosci. 2014, 39, 1551–1557. [Google Scholar] [CrossRef] [PubMed]
- Ueno, M.; Fujita, Y.; Tanaka, T.; Nakamura, Y.; Kikuta, J.; Ishii, M.; Yamashita, T. Layer V cortical neurons require microglial support for survival during postnatal development. Nat. Neurosci. 2013, 16, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Wakselman, S.; Bechade, C.; Roumier, A.; Bernard, D.; Triller, A.; Bessis, A. Developmental neuronal death in hippocampus requires the microglial CD11b integrin and DAP12 immunoreceptor. J. Neurosci. 2008, 28, 8138–8143. [Google Scholar] [CrossRef]
- Hoshiko, M.; Arnoux, I.; Avignone, E.; Yamamoto, N.; Audinat, E. Deficiency of the microglial receptor CX3CR1 impairs postnatal functional development of thalamocortical synapses in the barrel cortex. J. Neurosci. 2012, 32, 15106–15111. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.; Paolicelli, R.C.; Sforazzini, F.; Weinhard, L.; Bolasco, G.; Pagani, F.; Vyssotski, A.L.; Bifone, A.; Gozzi, A.; Ragozzino, D.; et al. Deficient neuron-microglia signaling results in impaired functional brain connectivity and social behavior. Nat. Neurosci. 2014, 17, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Lenz, K.M.; Nelson, L.H. Microglia and beyond: Innate immune cells as regulators of brain development and behavioral function. Front. Immunol. 2018, 9, 698. [Google Scholar] [CrossRef] [Green Version]
- Hagemeyer, N.; Hanft, K.M.; Akriditou, M.A.; Unger, N.; Park, E.S.; Stanley, E.R.; Staszewski, O.; Dimou, L.; Prinz, M. Microglia contribute to normal myelinogenesis and to oligodendrocyte progenitor maintenance during adulthood. Acta Neuropathol. 2017, 134, 441–458. [Google Scholar] [CrossRef] [Green Version]
- Wlodarczyk, A.; Holtman, I.R.; Krueger, M.; Yogev, N.; Bruttger, J.; Khorooshi, R.; Benmamar-Badel, A.; de Boer-Bergsma, J.J.; Martin, N.A.; Karram, K.; et al. A novel microglial subset plays a key role in myelinogenesis in developing brain. EMBO J. 2017, 36, 3292–3308. [Google Scholar] [CrossRef]
- Oikawa, T.; Yamada, T.; Kihara-Negishi, F.; Yamamoto, H.; Kondoh, N.; Hitomi, Y.; Hashimoto, Y. The role of Ets family transcription factor PU.1 in hematopoietic cell differentiation, proliferation and apoptosis. Cell Death Differ. 1999, 6, 599–608. [Google Scholar] [CrossRef] [Green Version]
- Delpech, J.C.; Wei, L.; Hao, J.; Yu, X.; Madore, C.; Butovsky, O.; Kaffman, A. Early life stress perturbs the maturation of microglia in the developing hippocampus. Brain Behav. Immun. 2016, 57, 79–93. [Google Scholar] [CrossRef] [Green Version]
- Ma, D.K.; Kim, W.R.; Ming, G.L.; Song, H. Activity-dependent extrinsic regulation of adult olfactory bulb and hippocampal neurogenesis. Ann. N. Y. Acad. Sci. 2009, 1170, 664–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sierra, A.; Encinas, J.M.; Deudero, J.J.; Chancey, J.H.; Enikolopov, G.; Overstreet-Wadiche, L.S.; Tsirka, S.E.; Maletic-Savatic, M. Microglia shape adult hippocampal neurogenesis through apoptosis-coupled phagocytosis. Cell Stem Cell 2010, 7, 483–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekdahl, C.T.; Claasen, J.H.; Bonde, S.; Kokaia, Z.; Lindvall, O. Inflammation is detrimental for neurogenesis in adult brain. Proc. Natl. Acad. Sci. USA 2003, 100, 13632–13637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziv, Y.; Schwartz, M. Immune-based regulation of adult neurogenesis: Implications for learning and memory. Brain Behav. Immun. 2008, 22, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Aarum, J.; Sandberg, K.; Haeberlein, S.L.; Persson, M.A. Migration and differentiation of neural precursor cells can be directed by microglia. Proc. Natl. Acad. Sci. USA 2003, 100, 15983–15988. [Google Scholar] [CrossRef] [Green Version]
- Walton, N.M.; Sutter, B.M.; Laywell, E.D.; Levkoff, L.H.; Kearns, S.M.; Marshall, G.P., 2nd; Scheffler, B.; Steindler, D.A. Microglia instruct subventricular zone neurogenesis. Glia 2006, 54, 815–825. [Google Scholar] [CrossRef]
- Vukovic, J.; Colditz, M.J.; Blackmore, D.G.; Ruitenberg, M.J.; Bartlett, P.F. Microglia modulate hippocampal neural precursor activity in response to exercise and aging. J. Neurosci. 2012, 32, 6435–6443. [Google Scholar] [CrossRef] [Green Version]
- Gemma, C.; Bachstetter, A.D. The role of microglia in adult hippocampal neurogenesis. Front. Cell Neurosci. 2013, 7, 229. [Google Scholar] [CrossRef] [Green Version]
- Ekdahl, C.T.; Kokaia, Z.; Lindvall, O. Brain inflammation and adult neurogenesis: The dual role of microglia. Neuroscience 2009, 158, 1021–1029. [Google Scholar] [CrossRef]
- Cacci, E.; Ajmone-Cat, M.A.; Anelli, T.; Biagioni, S.; Minghetti, L. In vitro neuronal and glial differentiation from embryonic or adult neural precursor cells are differently affected by chronic or acute activation of microglia. Glia 2008, 56, 412–425. [Google Scholar] [CrossRef]
- Koo, J.W.; Duman, R.S. IL-1beta is an essential mediator of the antineurogenic and anhedonic effects of stress. Proc. Natl. Acad. Sci. USA 2008, 105, 751–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cacci, E.; Claasen, J.H.; Kokaia, Z. Microglia-derived tumor necrosis factor-alpha exaggerates death of newborn hippocampal progenitor cells in vitro. J. Neurosci. Res. 2005, 80, 789–797. [Google Scholar] [CrossRef] [PubMed]
- Jakubs, K.; Bonde, S.; Iosif, R.E.; Ekdahl, C.T.; Kokaia, Z.; Kokaia, M.; Lindvall, O. Inflammation regulates functional integration of neurons born in adult brain. J. Neurosci. 2008, 28, 12477–12488. [Google Scholar] [CrossRef] [PubMed]
- Orre, M.; Kamphuis, W.; Osborn, L.M.; Melief, J.; Kooijman, L.; Huitinga, I.; Klooster, J.; Bossers, K.; Hol, E.M. Acute isolation and transcriptome characterization of cortical astrocytes and microglia from young and aged mice. Neurobiol. Aging 2014, 35, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Linnerbauer, M.; Wheeler, M.A.; Quintana, F.J. Astrocyte Crosstalk in CNS Inflammation. Neuron 2020, 108, 608–622. [Google Scholar] [CrossRef]
- Masuda, T.; Sankowski, R.; Staszewski, O.; Böttcher, C.; Amann, L.; Sagar; Scheiwe, C.; Nessler, S.; Kunz, P.; van Loo, G.; et al. Spatial and temporal heterogeneity of mouse and human microglia at single-cell resolution. Nature 2019, 566, 388–392. [Google Scholar] [CrossRef]
- Osborne, B.F.; Turano, A.; Caulfield, J.I.; Schwarz, J.M. Sex- and region-specific differences in microglia phenotype and characterization of the peripheral immune response following early-life infection in neonatal male and female rats. Neurosci. Lett. 2018, 692, 1–9. [Google Scholar] [CrossRef]
- Liang, M.; Zhong, H.; Rong, J.; Li, Y.; Zhu, C.; Zhou, L.; Zhou, R. Postnatal Lipopolysaccharide Exposure Impairs Adult Neurogenesis and Causes Depression-like Behaviors Through Astrocytes Activation Triggering GABAA Receptor Downregulation. Neuroscience 2019, 422, 21–31. [Google Scholar] [CrossRef]
- Saavedra, L.M.; Fenton Navarro, B.; Torner, L. Early life stress activates glial cells in the hippocampus but attenuates cytokine secretion in response to an immune challenge in rat pups. Neuroimmunomodulation 2017, 24, 242–255. [Google Scholar] [CrossRef]
- Réus, G.Z.; Silva, R.H.; de Moura, A.B.; Presa, J.F.; Abelaira, H.M.; Abatti, M.; Vieira, A.; Pescador, B.; Michels, M.; Ignácio, Z.M.; et al. Early Maternal Deprivation Induces Microglial Activation, Alters Glial Fibrillary Acidic Protein Immunoreactivity and Indoleamine 2,3-Dioxygenase during the Development of Offspring Rats. Mol. Neurobiol. 2019, 56, 1096–1108. [Google Scholar] [CrossRef]
- Hoeijmakers, L.; Ruigrok, S.R.; Amelianchik, A.; Ivan, D.; van Dam, A.M.; Lucassen, P.J.; Korosi, A. Early-life stress lastingly alters the neuroinflammatory response to amyloid pathology in an Alzheimer’s disease mouse model. Brain Behav. Immun. 2017, 63, 160–175. [Google Scholar] [CrossRef] [PubMed]
- Abbink, M.R.; Kotah, J.M.; Hoeijmakers, L.; Mak, A.; Yvon-Durocher, G.; van der Gaag, B.; Lucassen, P.J.; Korosi, A. Characterization of astrocytes throughout life in wildtype and APP/PS1 mice after early-life stress exposure. J. Neuroinflammation 2020, 17, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, M.G.; Thompson, B.M.; Watkins, L.R.; Maier, S.F. Glucocorticoids mediate stress-induced priming of microglial pro-inflammatory responses. Brain Behav. Immun. 2012, 26, 337–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, M.G.; Weber, M.D.; Fonken, L.K.; Hershman, S.A.; Watkins, L.R.; Maier, S.F. The redox state of the alarmin HMGB1 is a pivotal factor in neuroinflammatory and microglial priming: A role for the NLRP3 inflammasome. Brain Behav. Immun. 2016, 55, 215–224. [Google Scholar] [CrossRef] [Green Version]
- Bellavance, M.A.; Rivest, S. The HPA—Immune Axis and the Immunomodulatory Actions of Glucocorticoids in the Brain. Front. Immunol. 2014, 5, 136. [Google Scholar] [CrossRef] [Green Version]
- Crain, J.M.; Nikodemova, M.; Watters, J.J. Expression of P2 nucleotide receptors varies with age and sex in murine brain microglia. J. Neuroinflammation 2009, 6, 24. [Google Scholar] [CrossRef] [Green Version]
- Crain, J.M.; Nikodemova, M.; Watters, J.J. Microglia express distinct M1 and M2 phenotypic markers in the postnatal and adult central nervous system in male and female mice. J. Neurosci. Res. 2013, 91, 1143–1151. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, J.M.; Sholar, P.W.; Bilbo, S.D. Sex differences in microglial colonization of the developing rat brain. J. Neurochem. 2012, 120, 948–963. [Google Scholar] [CrossRef]
- Bolton, J.L.; Auten, R.L.; Bilbo, S.D. Prenatal air pollution exposure induces sexually dimorphic fetal programming of metabolic and neuroinflammatory outcomes in adult offspring. Brain Behav. Immun. 2014, 37, 30–44. [Google Scholar] [CrossRef]
- Roughton, K.; Boström, M.; Kalm, M.; Blomgren, K. Irradiation to the young mouse brain impaired white matter growth more in females than in males. Cell Death Dis. 2013, 4, e897. [Google Scholar] [CrossRef] [Green Version]
- Pimentel-Coelho, P.M.; Michaud, J.P.; Rivest, S. C-C chemokine receptor type 2 (CCR2) signaling protects neonatal male mice with hypoxic-ischemic hippocampal damage from developing spatial learning deficits. Behav. Brain Res. 2015, 286, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Nelson, L.H.; Warden, S.; Lenz, K.M. Sex differences in microglial phagocytosis in the neonatal hippocampus. Brain Behav. Immun. 2017, 64, 11–22. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, M.M.; Wright, C.L. Convergence of Sex Differences and the Neuroimmune System in Autism Spectrum Disorder. Biol. Psychiatry 2017, 81, 402–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, J.M.; Bilbo, S.D. Sex, glia, and development: Interactions in health and disease. Horm. Behav. 2012, 62, 243–253. [Google Scholar] [CrossRef] [Green Version]
- Hanamsagar, R.; Bilbo, S.D. Sex differences in neurodevelopmental and neurodegenerative disorders: Focus on microglial function and neuroinflammation during development. J. Steroid Biochem. Mol. Biol. 2016, 160, 127–133. [Google Scholar] [CrossRef] [Green Version]
- Banqueri, M.; Méndez, M.; Gómez-Lázaro, E.; Arias, J.L. Early life stress by repeated maternal sepAbbinkaration induces long-term neuroinflammatory response in glial cells of male rats. Stress 2019, 22, 563–570. [Google Scholar] [CrossRef]
- van Deijk, A.F.; Camargo, N.; Timmerman, J.; Heistek, T.; Brouwers, J.F.; Mogavero, F.; Mansvelder, H.D.; Smit, A.B.; Verheijen, M.H. Astrocyte lipid metabolism is critical for synapse development and function in vivo. Glia 2017, 65, 670–682. [Google Scholar] [CrossRef]
- Ganguly, P.; Thompson, V.; Gildawie, K.; Brenhouse, H.C. Adolescent food restriction in rats alters prefrontal cortex microglia in an experience-dependent manner. Stress 2018, 21, 162–168. [Google Scholar] [CrossRef]
- Stepanichev, M.Y.; Goryakina, T.; Manolova, A.; Lazareva, N.; Kvichanskii, A.; Tretyakova, L.; Volobueva, M.; Gulyaeva, N. Neonatal proinflammatory challenge evokes a microglial response and affects the ratio between subtypes of GABAergic interneurons in the hippocampus of juvenile rats: Sex-dependent and sex-independent effects. Brain Struct. Funct. 2021, 226, 563–574. [Google Scholar] [CrossRef]
- Bayer, T.A.; Falkai, P.; Maier, W. Genetic and non-genetic vulnerability factors in schizophrenia: The basis of the “two hit hypothesis”. J. Psychiatr. Res. 1999, 33, 543–548. [Google Scholar] [CrossRef]
- Walker, A.K.; Nakamura, T.; Byrne, R.J.; Naicker, S.; Tynan, R.J.; Hunter, M.; Hodgson, D.M. Neonatal lipopolysaccharide and adult stress exposure predisposes rats to anxiety-like behaviour and blunted corticosterone responses: Implications for the double-hit hypothesis. Psychoneuroendocrinology 2009, 34, 1515–1525. [Google Scholar] [CrossRef] [PubMed]
- Berkiks, I.; Garcia-Segura, L.M.; Nassiri, A.; Mesfioui, A.; Ouichou, A.; Boulbaroud, S.; Bahbiti, Y.; Lopez-Rodriguez, A.B.; Hasnaoui, E.; El Hessni, A. The sex differences of the behavior response to early Life immune stimulation: Microglia and astrocytes involvement. Physiol. Behav. 2019, 199, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Diz-Chaves, Y.; Astiz, M.; Bellini, M.J.; Garcia-Segura, L.M. Prenatal stress increases the expression of proinflammatory cytokines and exacerbates the inflammatory response to LPS in the hippocampal formation of adult male mice. Brain Behav. Immun. 2013, 28, 196–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilbo, S.D. Early-life infection is a vulnerability factor for aging-related glial alterations and cognitive decline. Neurobiol. Learn. Mem. 2010, 94, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Walker, A.K.; Nakamura, T.; Hodgson, D.M. Neonatal lipopolysaccharide exposure alters central cytokine responses to stress in adulthood in Wistar rats. Stress 2010, 13, 506–515. [Google Scholar] [CrossRef]
- Lana, D.; Ugolini, F.; Wenk, G.L.; Giovannini, M.G.; Zecchi-Orlandini, S.; Nosi, D. Microglial distribution, branching, and clearance activity in aged rat hippocampus are affected by astrocyte meshwork integrity: Evidence of a novel cell-cell interglial interaction. FASEB J. 2019, 33, 4007–4020. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aniol, V.; Manolova, A.; Gulyaeva, N. Early Life Events and Maturation of the Dentate Gyrus: Implications for Neurons and Glial Cells. Int. J. Mol. Sci. 2022, 23, 4261. https://doi.org/10.3390/ijms23084261
Aniol V, Manolova A, Gulyaeva N. Early Life Events and Maturation of the Dentate Gyrus: Implications for Neurons and Glial Cells. International Journal of Molecular Sciences. 2022; 23(8):4261. https://doi.org/10.3390/ijms23084261
Chicago/Turabian StyleAniol, Viktor, Anna Manolova, and Natalia Gulyaeva. 2022. "Early Life Events and Maturation of the Dentate Gyrus: Implications for Neurons and Glial Cells" International Journal of Molecular Sciences 23, no. 8: 4261. https://doi.org/10.3390/ijms23084261
APA StyleAniol, V., Manolova, A., & Gulyaeva, N. (2022). Early Life Events and Maturation of the Dentate Gyrus: Implications for Neurons and Glial Cells. International Journal of Molecular Sciences, 23(8), 4261. https://doi.org/10.3390/ijms23084261