
Molecular Effects of Low-Intensity Shock Wave Therapy on L6 Dorsal Root Ganglion/Spinal Cord and Blood Oxygenation Level-Dependent (BOLD) Functional Magnetic Resonance Imaging (fMRI) Changes in Capsaicin-Induced Prostatitis Rat Models


Abstract
:1. Introduction
2. Results
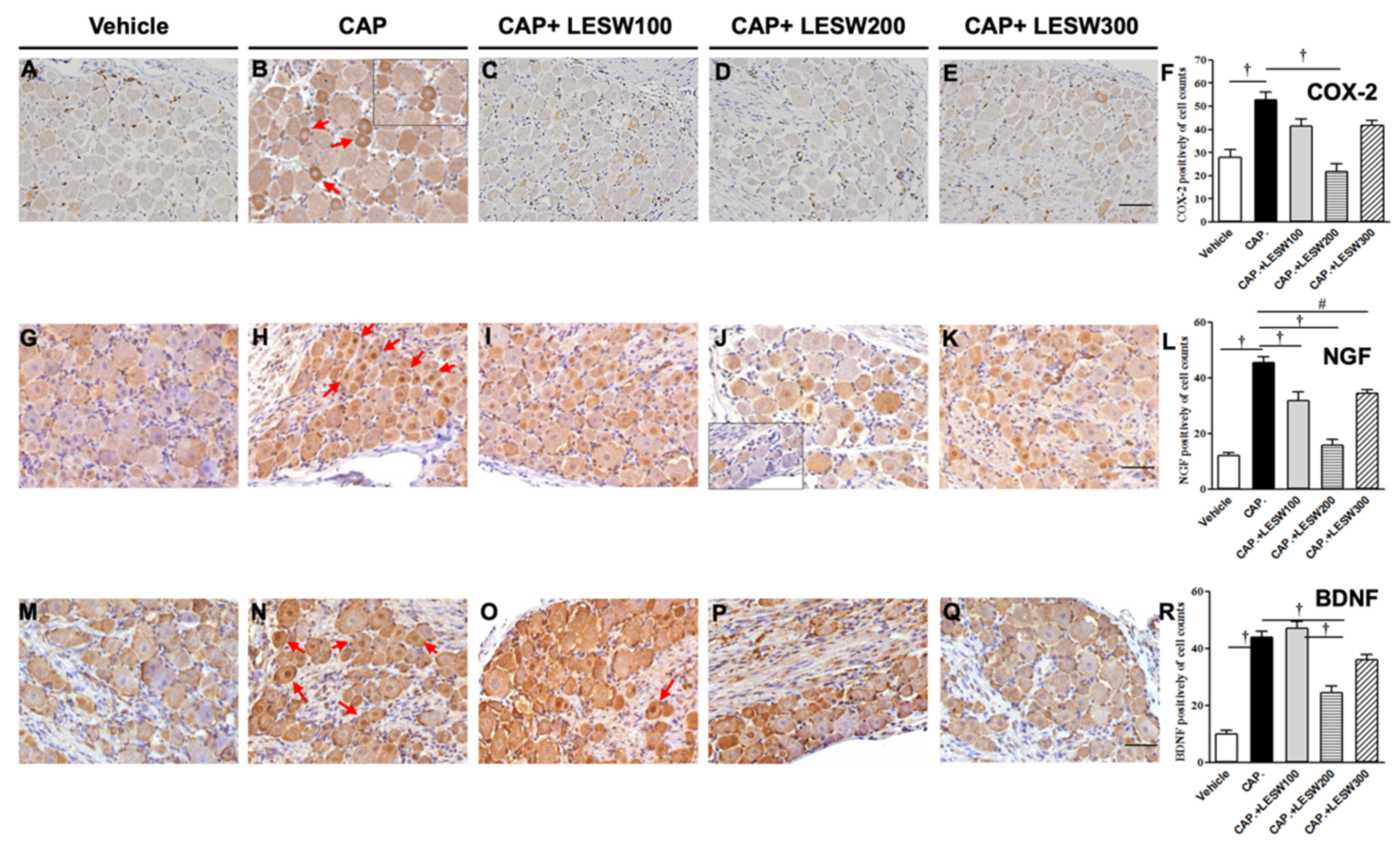
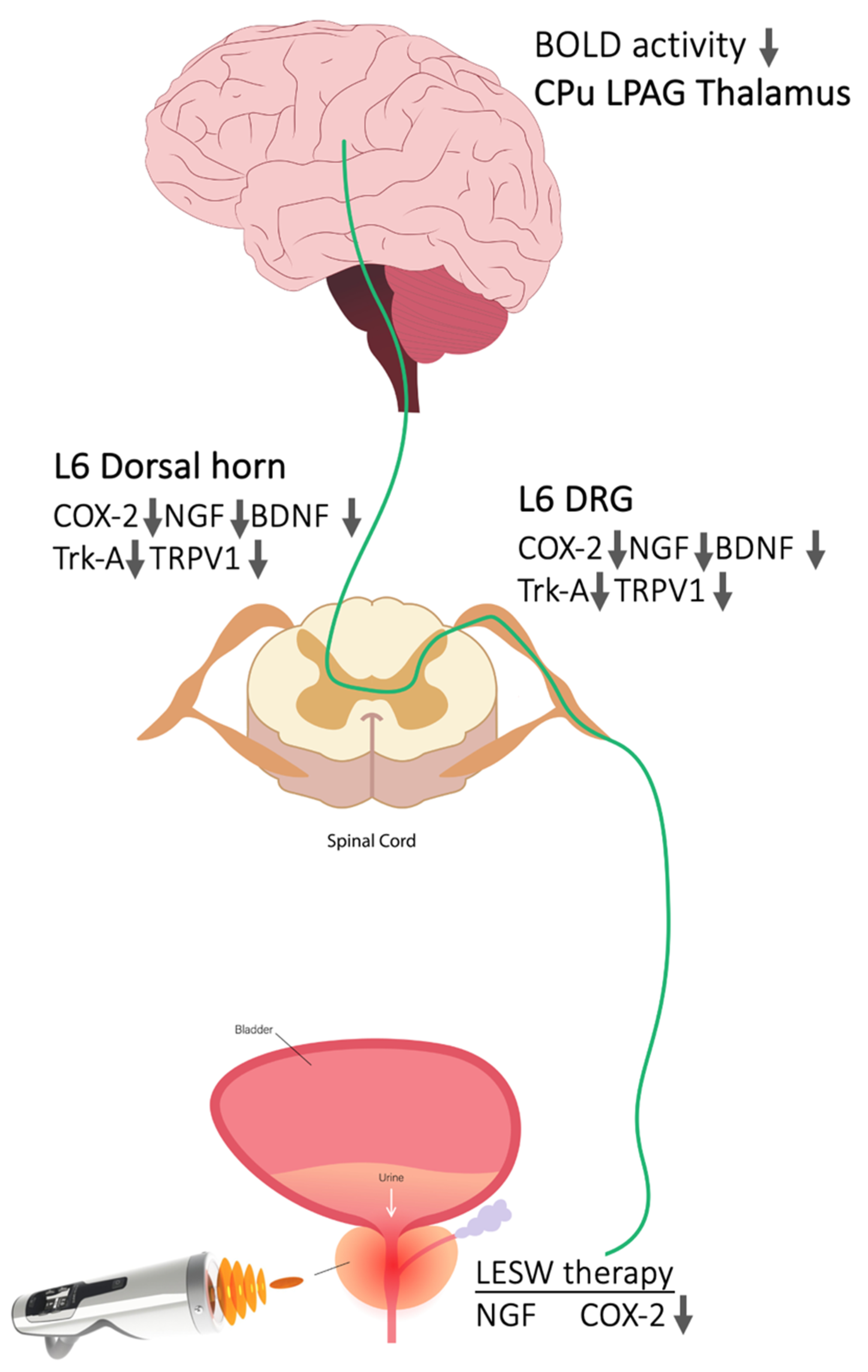
2.1. Immunostaining—Li-ESWT Decreased L6 DRG COX-2-, NGF-, and BDNF-Positive Neurons Induced by Intraprostatic Capsaicin Injection
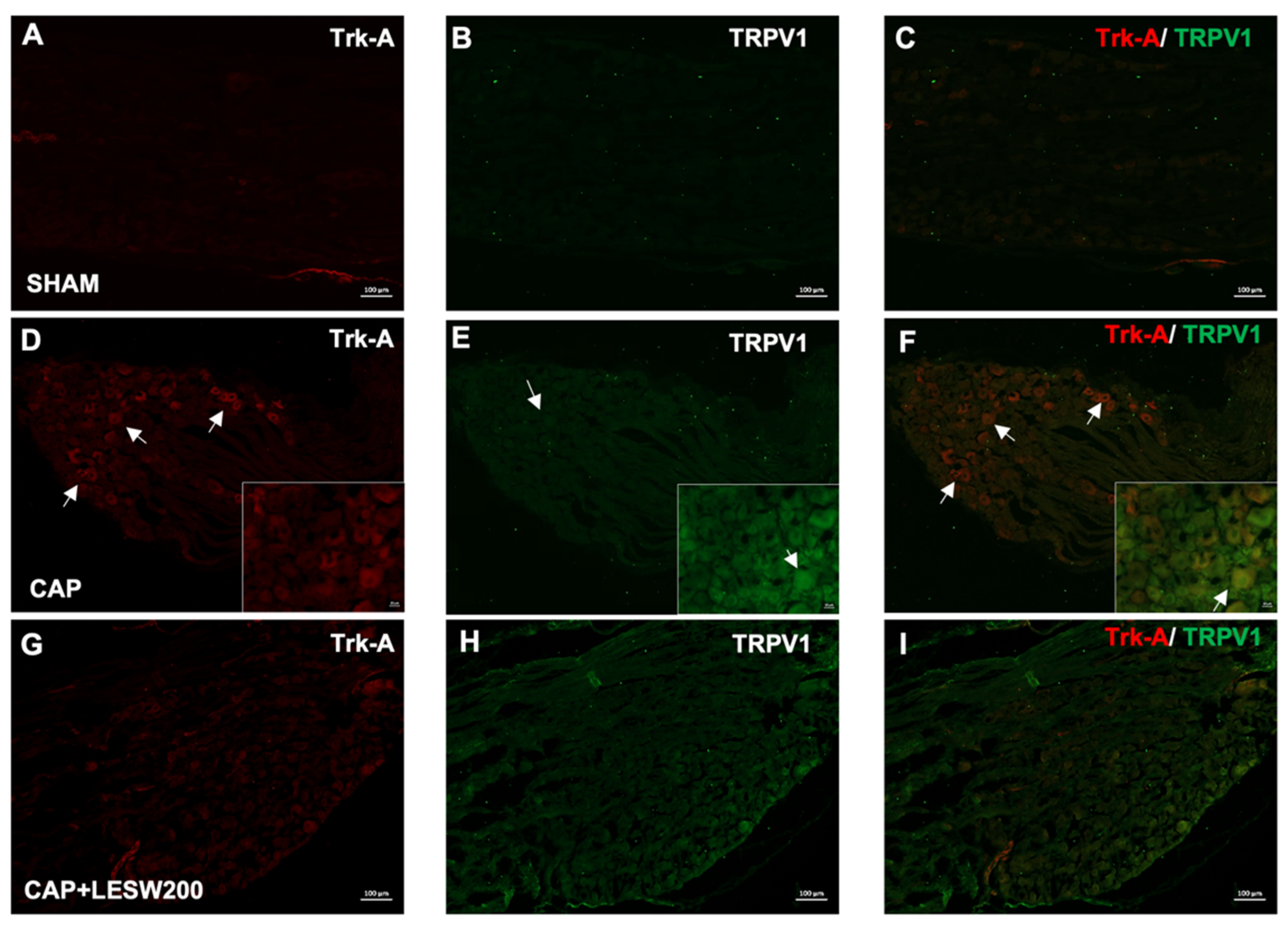
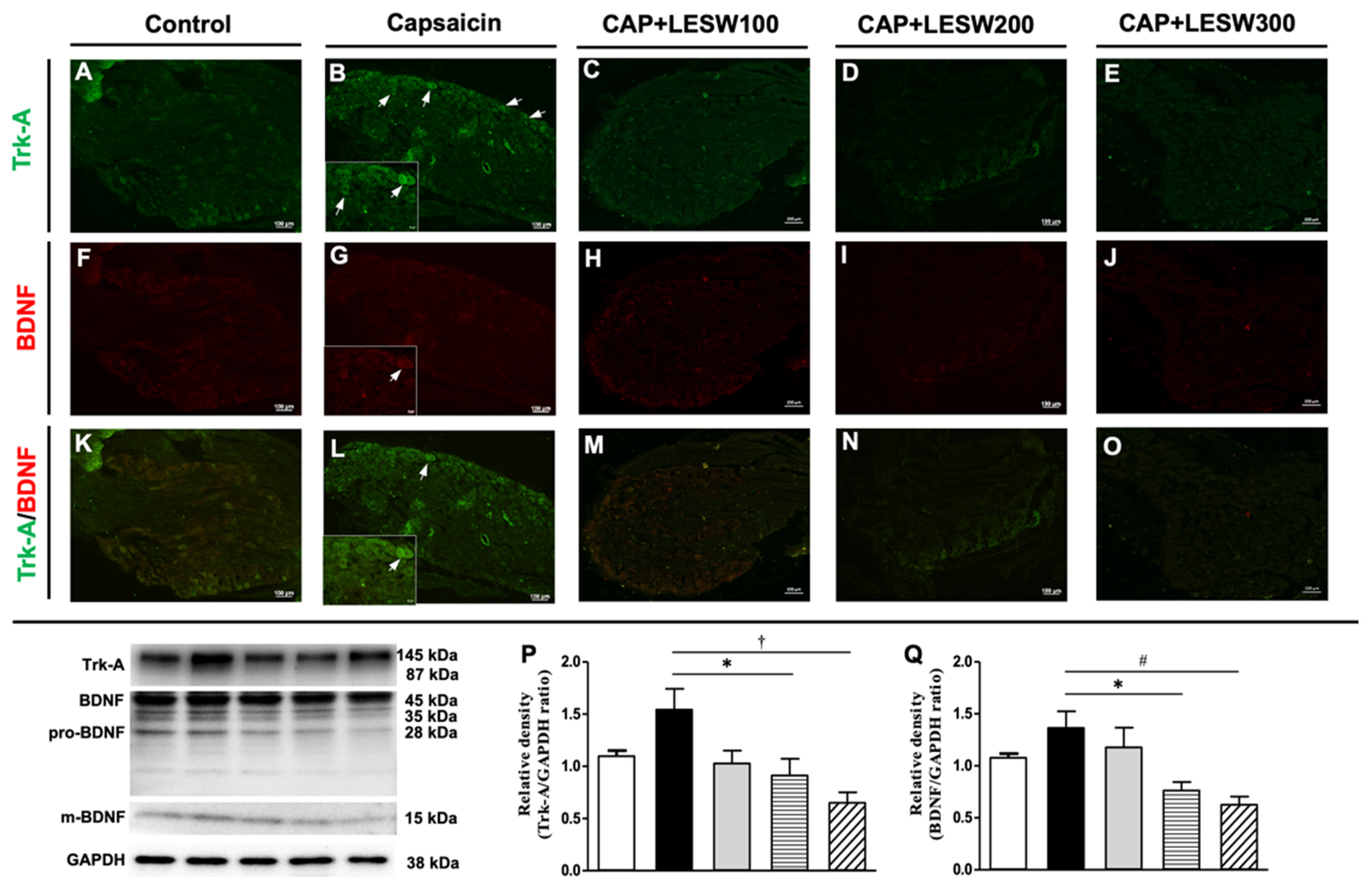
2.2. Immunostaining—Li-ESWT Decreased Upregulation of Trk-A and TRPV1 Immunoreactivity in L6 DRG Induced by Intraprostatic Capsaicin Injection
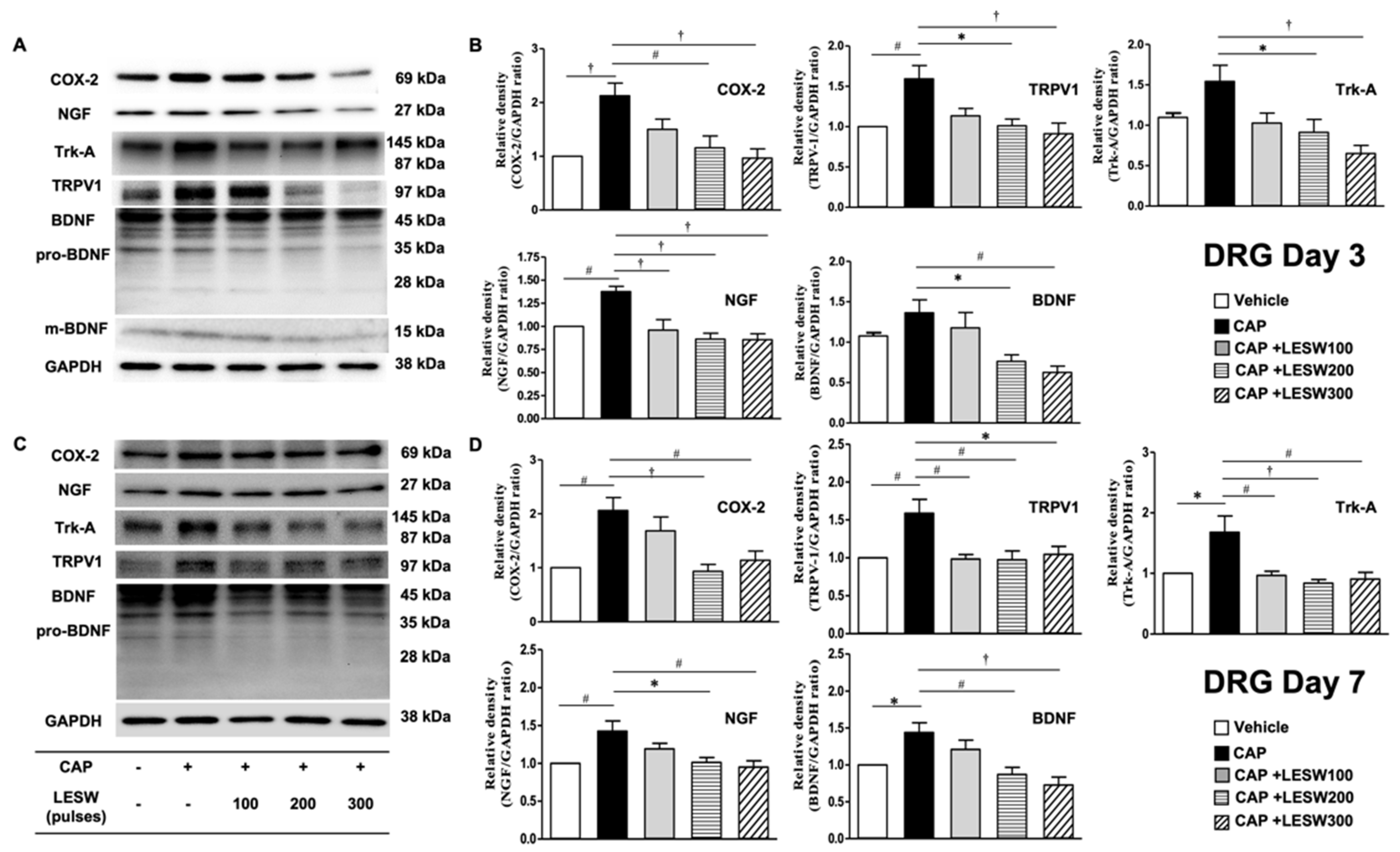
2.3. Western Blotting—Li-ESWT Suppressed Upregulation of COX-2, NGF, and BDNF and Trk-A and TRPV1 in L6 DRG/Spinal Cord Induced by Intraprostatic Capsaicin Injection
2.4. Li-ESWT Suppressed the Progression of BOLD fMRI Signal Changes That Increased with Time in Prostatitis Rats
3. Discussion
4. Materials and Methods
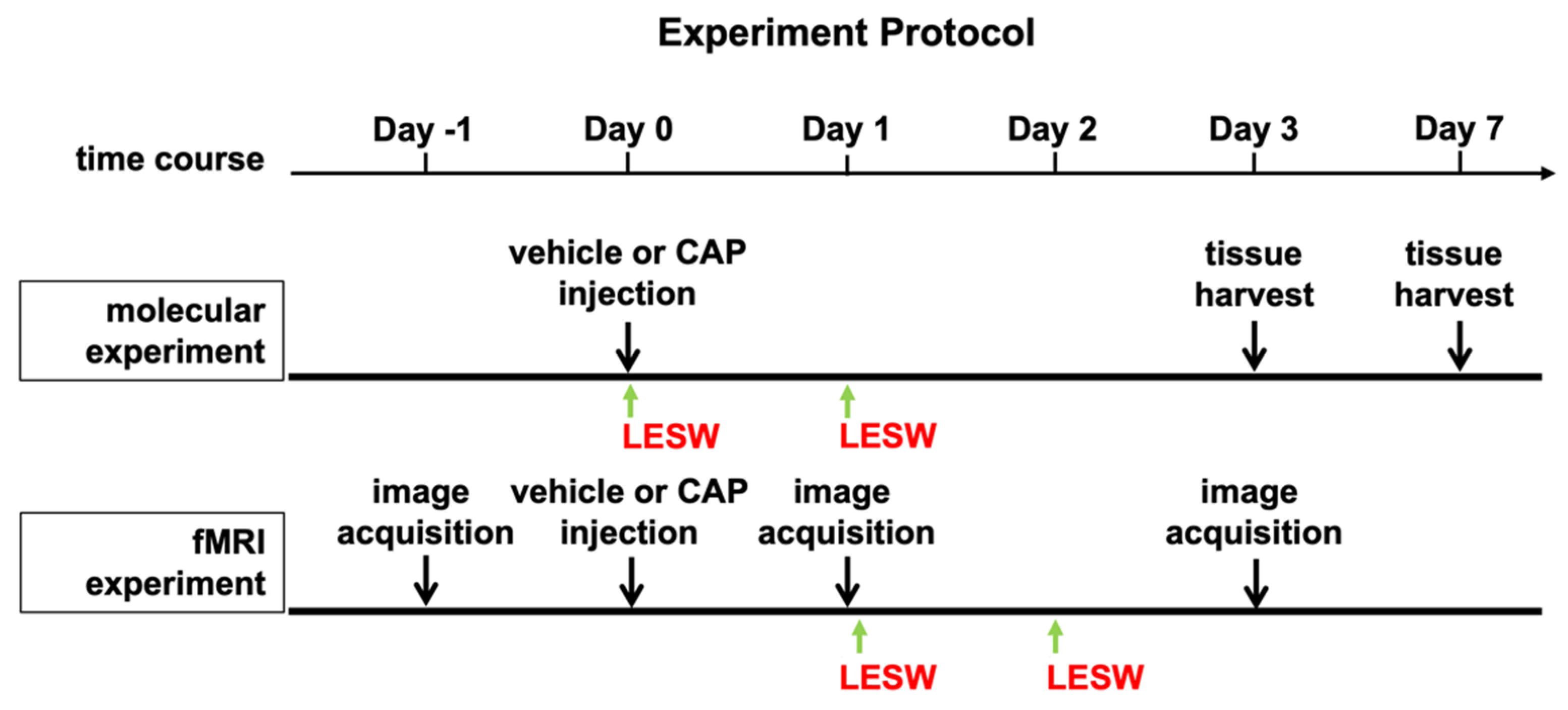
4.1. Experimental Animals and Study Design
4.2. Capsaicin Injection-Induced Prostatitis
4.3. Low-Energy Shock Wave Treatments
4.4. Immunohistochemistry and Immunofluorescent Studies for the L6 Spinal Cord and L6 Dorsal Root Ganglia
4.5. Western Blotting
4.6. MR Image Acquisition and Parameters
4.7. Post-Processing of Images Data Analysis
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nickel, J.C.; Downey, J.; Hunter, D.; Clark, J. Prevalence of prostatitis-like symptoms in a population based study using the National Institutes of Health chronic prostatitis symptom index. J. Urol. 2001, 165, 842–845. [Google Scholar] [CrossRef]
- Strauss, A.C.; Dimitrakov, J.D. New treatments for chronic prostatitis/chronic pelvic pain syndrome. Nat. Rev. Urol. 2010, 7, 127–135. [Google Scholar] [CrossRef]
- Pontari, M.A.; Ruggieri, M.R. Mechanisms in prostatitis/chronic pelvic pain syndrome. J. Urol. 2004, 172, 839–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Reed-Maldonado, A.B.; Ly, K.; Lin, G.; Lue, T.F. Potential applications of low-intensity extracorporeal shock-wave therapy in urological diseases via activation of tissue resident stem cells. Urol. Sci. 2022, 33, 3–8. [Google Scholar]
- Zimmermann, R.; Cumpanas, A.; Miclea, F.; Janetschek, G. Extracorporeal shock wave therapy for the treatment of chronic pelvic pain syndrome in males: A randomised, double-blind, placebo-controlled study. Eur. Urol. 2009, 56, 418–424. [Google Scholar] [CrossRef]
- Mykoniatis, I.; Kalyvianakis, D.; Zilotis, F.; Kapoteli, P.; Fournaraki, A.; Poulios, E.; Hatzichristou, D. Evaluation of a low-intensity shockwave therapy for chronic prostatitis type IIIb/chronic pelvic pain syndrome: A double-blind randomized sham-controlled clinical trial. Prostate Cancer Prostatic Dis. 2021, 24, 370–379. [Google Scholar] [CrossRef]
- Mykoniatis, I.; Pyrgidis, N.; Sokolakis, I.; Sountoulides, P.; Hatzichristodoulou, G.; Apostolidis, A.; Hatzichristou, D. Low-intensity shockwave therapy for the management of chronic prostatitis/chronic pelvic pain syndrome: A systematic review and meta-analysis. JAMA 2011, 305, 78–86. [Google Scholar] [CrossRef]
- Wess, O.J. A neural model for chronic pain and pain relief by extracorporeal shock wave treatment. Urol. Res. 2008, 36, 327–334. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.J.; Tyagi, P.; Chen, Y.M.; Chancellor, M.B.; Chuang, Y.C. Low Energy Shock Wave Therapy Inhibits Inflammatory Molecules and Suppresses Prostatic Pain and Hypersensitivity in a Capsaicin Induced Prostatitis Model in Rats. Int. J. Mol. Sci 2019, 20, 4777. [Google Scholar] [CrossRef] [Green Version]
- Ishigooka, M.; Zermann, D.H.; Doggweiler, R.; Schmidt, R.A. Similarity of distributions of spinal c-Fos and plasma extravasation after acute chemical irritation of the bladder and the prostate. J. Urol. 2000, 164, 1751–1756. [Google Scholar] [CrossRef]
- Schwartz, E.S.; La, J.H.; Young, E.E.; Feng, B.; Joyce, S.; Gebhart, G.F. Chronic Prostatitis Induces Bladder Hypersensitivity and Sensitizes Bladder Afferents in the Mouse. J. Urol. 2016, 196, 892–901. [Google Scholar] [CrossRef] [PubMed]
- Kaya, S.; Hermans, L.; Willems, T.; Roussel, N.; Meeus, M. Central sensitization in urogynecological chronic pelvic pain: A systematic literature review. Pain Physician 2013, 16, 291–308. [Google Scholar] [CrossRef] [PubMed]
- Delcroix, J.D.; Valletta, J.S.; Wu, C.; Hunt, S.J.; Kowal, A.S.; Mobley, W.C. NGF signaling in sensory neurons: Evidence that early endosomes carry NGF retrograde signals. Neuron 2003, 39, 69–84. [Google Scholar] [CrossRef] [Green Version]
- Miller, L.J.; Fischer, K.A.; Goralnick, S.J.; Litt, M.; Burleson, J.A.; Albertsen, P.; Kreutzer, D.L. Nerve growth factor and chronic prostatitis/chronic pelvic pain syndrome. Urology 2002, 59, 603–608. [Google Scholar] [CrossRef]
- Yoshida, K.; Gage, F.H. Cooperative regulation of nerve growth factor synthesis and secretion in fibroblasts and astrocytes by fibroblast growth factor and other cytokines. Brain Res. 1992, 569, 14–25. [Google Scholar] [CrossRef]
- Woolf, C.J.; Safieh-Garabedian, B.; Ma, Q.P.; Crilly, P.; Winter, J. Nerve growth factor contributes to the generation of inflammatory sensory hypersensitivity. Neuroscience 1994, 62, 327–331. [Google Scholar] [CrossRef]
- Woolf, C.J.; Allchorne, A.; Safieh-Garabedian, B.; Poole, S. Cytokines, nerve growth factor and inflammatory hyperalgesia: The contribution of tumour necrosis factor alpha. Br. J. Pharmacol. 1997, 121, 417–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, B.; Dong, Z.; Wang, Y.; Yan, G.; Yang, E.; Cheng, H.; Liang, C.; Hao, Z.; Zhang, X.; Song, Z.; et al. Li-ESWT treatment reduces inflammation, oxidative stress, and pain via the PI3K/AKT/FOXO1 pathway in autoimmune prostatitis rat models. Andrology 2021, 9, 1593–1602. [Google Scholar] [CrossRef] [PubMed]
- Leon, A.; Buriani, A.; Dal Toso, R.; Fabris, M.; Romanello, S.; Aloe, L.; Levi-Montalcini, R. Mast cells synthesize, store, and release nerve growth factor. Proc. Natl. Acad. Sci. USA 1994, 91, 3739–3743. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.; Jin, C.; Bian, Z.; Liang, C. Extracorporeal shock wave therapy decreases the number of total and degranulated mast cells and alleviates pelvic pain in a rat model of prostatitis. Mol. Cell. Biochem. 2021, 476, 1905–1913. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.J.; Tyagi, P.; Lin, T.K.; Huang, C.C.; Lee, W.C.; Chancellor, M.B.; Chuang, Y.C. Low Energy Shock Wave Therapy Attenuates Mitochondrial Dysfunction and Improves Bladder Function in HCl induced Cystitis in Rats. Biomed. J. 2021. [Google Scholar] [CrossRef] [PubMed]
- Chuang, Y.C.; Yoshimura, N.; Huang, C.C.; Wu, M.; Chiang, P.H.; Chancellor, M.B. Intraprostatic botulinum toxin a injection inhibits cyclooxygenase-2 expression and suppresses prostatic pain on capsaicin induced prostatitis model in rat. J. Urol. 2008, 180, 742–748. [Google Scholar] [CrossRef] [PubMed]
- Chuang, Y.C.; Yoshimura, N.; Wu, M.; Huang, C.C.; Chiang, P.H.; Tyagi, P.; Chancellor, M.B. Intraprostatic capsaicin injection as a novel model for nonbacterial prostatitis and effects of botulinum toxin A. Eur. Urol. 2007, 51, 1119–1127. [Google Scholar] [CrossRef] [PubMed]
- Dinis, P.; Charrua, A.; Avelino, A.; Nagy, I.; Quintas, J.; Ribau, U.; Cruz, F. The distribution of sensory fibers immunoreactive for the TRPV1 (capsaicin) receptor in the human prostate. Eur. Urol. 2005, 48, 162–167. [Google Scholar] [CrossRef]
- Richardson, J.D.; Vasko, M.R. Cellular mechanisms of neurogenic inflammation. J. Pharmacol. Exp. Ther. 2002, 302, 839–845. [Google Scholar] [CrossRef] [Green Version]
- Bonnington, J.K.; McNaughton, P.A. Signalling pathways involved in the sensitisation of mouse nociceptive neurones by nerve growth factor. J. Physiol. 2003, 551, 433–446. [Google Scholar] [CrossRef]
- Zhang, J.; Yi, Q.T.; Gong, M.; Zhang, Y.Q.; Liu, D.; Zhu, R.J. Upregulation of TRPV1 in spinal dorsal root ganglion by activating NGF-TrkA pathway contributes to pelvic organ cross-sensitisation in rats with experimental autoimmune prostatitis. Andrologia 2019, 51, 13302. [Google Scholar] [CrossRef]
- Roman, K.; Hall, C.; Schaeffer, A.J.; Thumbikat, P. TRPV1 in experimental autoimmune prostatitis. Prostate 2020, 80, 28–37. [Google Scholar] [CrossRef]
- Dimitriadis, F.; Papaioannou, M.; Sokolakis, I.; Fragou, A.; Hatzichristou, D.; Apostolidis, A. The Effect of Low-Intensity Extracorporeal Shockwave Treatment on the Urinary Bladder in an Experimental Diabetic Rat Model. Int. Neurourol. J. 2021, 25, 34–41. [Google Scholar] [CrossRef]
- Thompson, S.W.; Bennett, D.L.; Kerr, B.J.; Bradbury, E.J.; McMahon, S.B. Brain-derived neurotrophic factor is an endogenous modulator of nociceptive responses in the spinal cord. Proc. Natl. Acad. Sci. USA 1999, 96, 7714–7718. [Google Scholar] [CrossRef] [Green Version]
- Michael, G.J.; Averill, S.; Nitkunan, A.; Rattray, M.; Bennett, D.L.; Yan, Q.; Priestley, J.V. Nerve growth factor treatment increases brain-derived neurotrophic factor selectively in TrkA-expressing dorsal root ganglion cells and in their central terminations within the spinal cord. J. Neurosci. 1997, 17, 8476–8490. [Google Scholar] [CrossRef]
- Wang, B.; Ning, H.; Reed-Maldonado, A.B.; Zhou, J.; Ruan, Y.; Zhou, T.; Wang, H.S.; Oh, B.S.; Banie, L.; Lin, G.; et al. Low-Intensity Extracorporeal Shock Wave Therapy Enhances Brain-Derived Neurotrophic Factor Expression through PERK/ATF4 Signaling Pathway. Int. J. Mol. Sci. 2017, 18, 433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, M.; Kanno, H.; Sugaya, T.; Yamaya, S.; Yahata, K.; Handa, K.; Shindo, T.; Shimokawa, H.; Ozawa, H.; Itoi, E. Low-energy extracorporeal shock wave therapy promotes BDNF expression and improves functional recovery after spinal cord injury in rats. Exp. Neurol. 2020, 328, 113251. [Google Scholar] [CrossRef]
- Araldi, D.; Ferrari, L.F.; Lotufo, C.M.; Vieira, A.S.; Athie, M.C.; Figueiredo, J.G.; Duarte, D.B.; Tambeli, C.H.; Ferreira, S.H.; Parada, C.A. Peripheral inflammatory hyperalgesia depends on the COX increase in the dorsal root ganglion. Proc. Natl. Acad. Sci. USA 2013, 110, 3603–3608. [Google Scholar] [CrossRef] [Green Version]
- Jang, Y.; Kim, M.; Hwang, S.W. Molecular mechanisms underlying the actions of arachidonic acid-derived prostaglandins on peripheral nociception. J. Neuroinflamm. 2020, 17, 30. [Google Scholar] [CrossRef] [PubMed]
- Amirmohseni, S.; Segelcke, D.; Reichl, S.; Wachsmuth, L.; Gorlich, D.; Faber, C.; Pogatzki-Zahn, E. Characterization of incisional and inflammatory pain in rats using functional tools of MRI. Neuroimage 2015, 127, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Yee, J.R.; Kenkel, W.; Caccaviello, J.C.; Gamber, K.; Simmons, P.; Nedelman, M.; Kulkarni, P.; Ferris, C.F. Identifying the integrated neural networks involved in capsaicin-induced pain using fMRI in awake TRPV1 knockout and wild-type rats. Front. Syst. Neurosci. 2015, 9, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubbard, C.S.; Khan, S.A.; Xu, S.; Cha, M.; Masri, R.; Seminowicz, D.A. Behavioral, metabolic and functional brain changes in a rat model of chronic neuropathic pain: A longitudinal MRI study. Neuroimage 2015, 107, 333–344. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. Paxino’s and Watson’s the Rat Brain in Stereotaxic Coordinates, 7th ed.; Elsevier/AP, Academic Press: Amsterdam, The Netherlands; Boston, MA, USA, 2014; p. 1. [Google Scholar]
- Brevard, M.E.; Duong, T.Q.; King, J.A.; Ferris, C.F. Changes in MRI signal intensity during hypercapnic challenge under conscious and anesthetized conditions. Magn. Reson. Imaging 2003, 21, 995–1001. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Adjusted p-Value (Bonferroni’s Multiple Comparisons Test) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Sham Control | Capsaicin (Cap.) | +Cap. LESW100 | +Cap. LESW200 | +Cap. LESW300 | Cap.vs. Sham | +Cap. LESW100 vs. Cap. | +Cap. LESW200 vs. Cap. | +Cap. LESW300 vs. Cap. | ||
| Day 3 | COX-2 | 1.00 ± 0.00 | 2.13 ± 0.24 | 1.50 ± 0.19 | 1.16 ± 0.22 | 0.97 ± 0.17 | † <0.001 | NS. | # <0.01 | † <0.001 |
| NGF | 1.00 ± 0.00 | 1.38 ± 0.06 | 0.96 ± 0.11 | 0.86 ± 0.06 | 0.86 ± 0.06 | # <0.01 | † <0.001 | † <0.001 | † <0.001 | |
| Trk-A | 1.00 ± 0.00 | 1.56 ± 0.21 | 1.01 ± 0.11 | 0.83 ± 0.12 | 0.69 ± 0.095 | * <0.05 | * <0.05 | # <0.01 | † <0.001 | |
| TRPV1 | 1.00 ± 0.00 | 1.60 ± 0.17 | 1.14 ± 0.09 | 1.01 ± 0.08 | 0.91 ± 0.13 | # <0.01 | NS. | * <0.05 | † <0.001 | |
| BDNF | 1.00 ± 0.00 | 1.47 ± 0.18 | 1.02 ± 0.09 | 0.82 ± 0.08 | 0.68 ± 0.10 | * <0.05 | * <0.05 | † <0.001 | † <0.001 | |
| Day 7 | COX-2 | 1.00 ± 0.00 | 2.06 ± 0.24 | 1.68 ± 0.26 | 0.93 ± 0.13 | 1.14 ± 0.17 | <0.01 | NS. | <0.001 | <0.01 |
| NGF | 1.00 ± 0.00 | 1.43 ± 0.14 | 1.19 ± 0.07 | 1.01 ± 0.07 | 0.95 ± 0.08 | <0.01 | NS. | <0.05 | <0.01 | |
| Trk-A | 1.00 ± 0.00 | 1.68 ± 0.27 | 0.97 ± 0.07 | 0.84 ± 0.06 | 0.91 ± 0.11 | <0.05 | <0.01 | <0.001 | <0.01 | |
| TRPV1 | 1.00 ± 0.00 | 1.51 ± 0.18 | 0.98 ± 0.06 | 0.97 ± 0.12 | 1.04 ± 0.11 | <0.05 | <0.05 | <0.05 | <0.05 | |
| BDNF | 1.00 ± 0.00 | 1.44 ± 0.13 | 1.21 ± 0.13 | 0.87 ± 0.10 | 0.73 ± 0.11 | <0.05 | NS. | <0.01 | <0.001 | |
| Adjusted p-Value (Bonferroni’s Multiple Comparisons Test) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Sham Control | Capsaicin (Cap.) | +Cap. LESW100 | +Cap. LESW200 | +Cap. LESW300 | Cap.vs. Sham | +Cap. LESW100 vs. Cap. | +Cap. LESW200 vs. Cap. | +Cap. LESW300 vs. Cap. | ||
| Day 3 | COX-2 | 1.00 ± 0.00 | 2.20 ± 0.29 | 1.42 ± 0.25 | 1.02 ± 0.14 | 0.95 ± 0.15 | † <0.001 | NS. | # <0.01 | † <0.001 |
| NGF | 1.00 ± 0.00 | 1.33 ± 0.095 | 1.01 ± 0.09 | 0.76 ± 0.05 | 0.83 ± 0.06 | * <0.05 | * <0.05 | † <0.001 | † <0.001 | |
| Trk-A | 1.00 ± 0.00 | 1.70 ± 0.12 | 1.34 ± 0.11 | 0.87 ± 0.23 | 0.90 ± 0.12 | # <0.01 | NS. | † <0.001 | † <0.001 | |
| TRPV1 | 1.00 ± 0.00 | 1.63 ± 0.17 | 1.42 ± 0.18 | 0.87 ± 0.10 | 0.97 ± 0.16 | * <0.05 | NS. | # <0.01 | * <0.05 | |
| BDNF | 1.00 ± 0.00 | 1.99 ± 0.19 | 1.60 ± 0.32 | 0.94 ± 0.12 | 0.96 ± 0.18 | * <0.05 | NS. | * <0.05 | # <0.01 | |
| Day 7 | COX-2 | 1.00 ± 0.00 | 2.25 ± 0.35 | 1.13 ± 0.18 | 0.84 ± 0.05 | 1.17 ± 0.30 | <0.01 | <0.01 | <0.001 | <0.05 |
| NGF | 1.00 ± 0.00 | 2.10 ± 0.43 | 1.09 ± 0.10 | 1.00 ± 0.12 | 0.91 ± 0.16 | <0.01 | <0.05 | <0.01 | <0.01 | |
| Trk-A | 1.00 ± 0.00 | 2.58 ± 0.70 | 1.04 ± 0.054 | 1.00 ± 0.10 | 1.52 ± 0.43 | <0.05 | <0.05 | <0.05 | NS. | |
| TRPV1 | 1.00 ± 0.00 | 1.91 ± 0.29 | 0.84 ± 0.081 | 0.85 ± 0.06 | 1.21 ± 0.23 | <0.01 | <0.001 | <0.001 | NS. | |
| BDNF | 1.00 ± 0.00 | 1.68 ± 0.23 | 1.04 ± 0.099 | 0.91 ± 0.08 | 0.91 ± 0.14 | <0.01 | <0.01 | <0.001 | <0.01 | |
| Average Number of Active Voxels | Significant (Tukey’s Multiple Comparisons Test) | |||||
|---|---|---|---|---|---|---|
| Sham Control | Capsaicin (CAP) | CAP + LESW200 | Sham vs. Cap | CAP vs. CAP + LESW200 | CAP + LESW200 vs. Sham | |
| Thalamus; Th | ||||||
| pre-injection | 62.25 ± 46.29 | 66.08 ± 43.93 | 94.00 ± 41.59 | NS | NS | NS |
| postcapsaicininjection, day1 | 209.20 ± 70.59 | 259.60 ± 82.05 | 410.83 ± 38.06 | NS | NSNS | |
| postcapsaicininjection, day3 | 217.71 ± 102.77 | 459.830 ± 89.98 | 83.50 ± 43.04 | * | # | NS |
| Lateral PAG; LPAG | ||||||
| pre-injection | 50.25 ± 20.77 | 39.17 ± 12.61 | 44.40 ± 11.90 | NS | NS | NS |
| postcapsaicininjection, day1 | 62.60 ± 13.94 | 93.00 ± 16.24 | 92.60 ± 10.50 | NS | NS | NS |
| postcapsaicininjection, day3 | 28.8 ± 7.61 | 54.50 ± 6.54 | 12.17 ± 3.68 | NS | * | NS |
| caudate putamen; CPu | ||||||
| pre-injection | 71.00 ± 48.61 | 49.20 ± 35.03 | 69.88 ± 25.44 | NS | NS | NS |
| Postcapsaicininjection, day1 | 193.60 ± 26.34 | 302.75 ± 23.35 | 455.00 ± 25.86 | NS | NS | * |
| postcapsaicininjection, day3 | 209.71 ± 98.66 | 387.43 ± 106.39 | 49.00 ± 20.15 | NS | † | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.-J.; Su, C.-H.; Chen, Y.-M.; Yu, C.-C.; Chuang, Y.-C. Molecular Effects of Low-Intensity Shock Wave Therapy on L6 Dorsal Root Ganglion/Spinal Cord and Blood Oxygenation Level-Dependent (BOLD) Functional Magnetic Resonance Imaging (fMRI) Changes in Capsaicin-Induced Prostatitis Rat Models. Int. J. Mol. Sci. 2022, 23, 4716. https://doi.org/10.3390/ijms23094716
Wang H-J, Su C-H, Chen Y-M, Yu C-C, Chuang Y-C. Molecular Effects of Low-Intensity Shock Wave Therapy on L6 Dorsal Root Ganglion/Spinal Cord and Blood Oxygenation Level-Dependent (BOLD) Functional Magnetic Resonance Imaging (fMRI) Changes in Capsaicin-Induced Prostatitis Rat Models. International Journal of Molecular Sciences. 2022; 23(9):4716. https://doi.org/10.3390/ijms23094716
Chicago/Turabian StyleWang, Hung-Jen, Chia-Hao Su, Yu-Ming Chen, Chun-Chieh Yu, and Yao-Chi Chuang. 2022. "Molecular Effects of Low-Intensity Shock Wave Therapy on L6 Dorsal Root Ganglion/Spinal Cord and Blood Oxygenation Level-Dependent (BOLD) Functional Magnetic Resonance Imaging (fMRI) Changes in Capsaicin-Induced Prostatitis Rat Models" International Journal of Molecular Sciences 23, no. 9: 4716. https://doi.org/10.3390/ijms23094716
APA StyleWang, H. -J., Su, C. -H., Chen, Y. -M., Yu, C. -C., & Chuang, Y. -C. (2022). Molecular Effects of Low-Intensity Shock Wave Therapy on L6 Dorsal Root Ganglion/Spinal Cord and Blood Oxygenation Level-Dependent (BOLD) Functional Magnetic Resonance Imaging (fMRI) Changes in Capsaicin-Induced Prostatitis Rat Models. International Journal of Molecular Sciences, 23(9), 4716. https://doi.org/10.3390/ijms23094716