
Acanthopanax senticosus Polysaccharide Enhances the Pathogen Resistance of Radiation-Damaged Caenorhabditis elegans through Intestinal p38 MAPK-SKN-1/ATF-7 Pathway and Stress Response

 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. ASPS Enhanced the Pathogen Resistance of Radiation-Damaged Nematodes
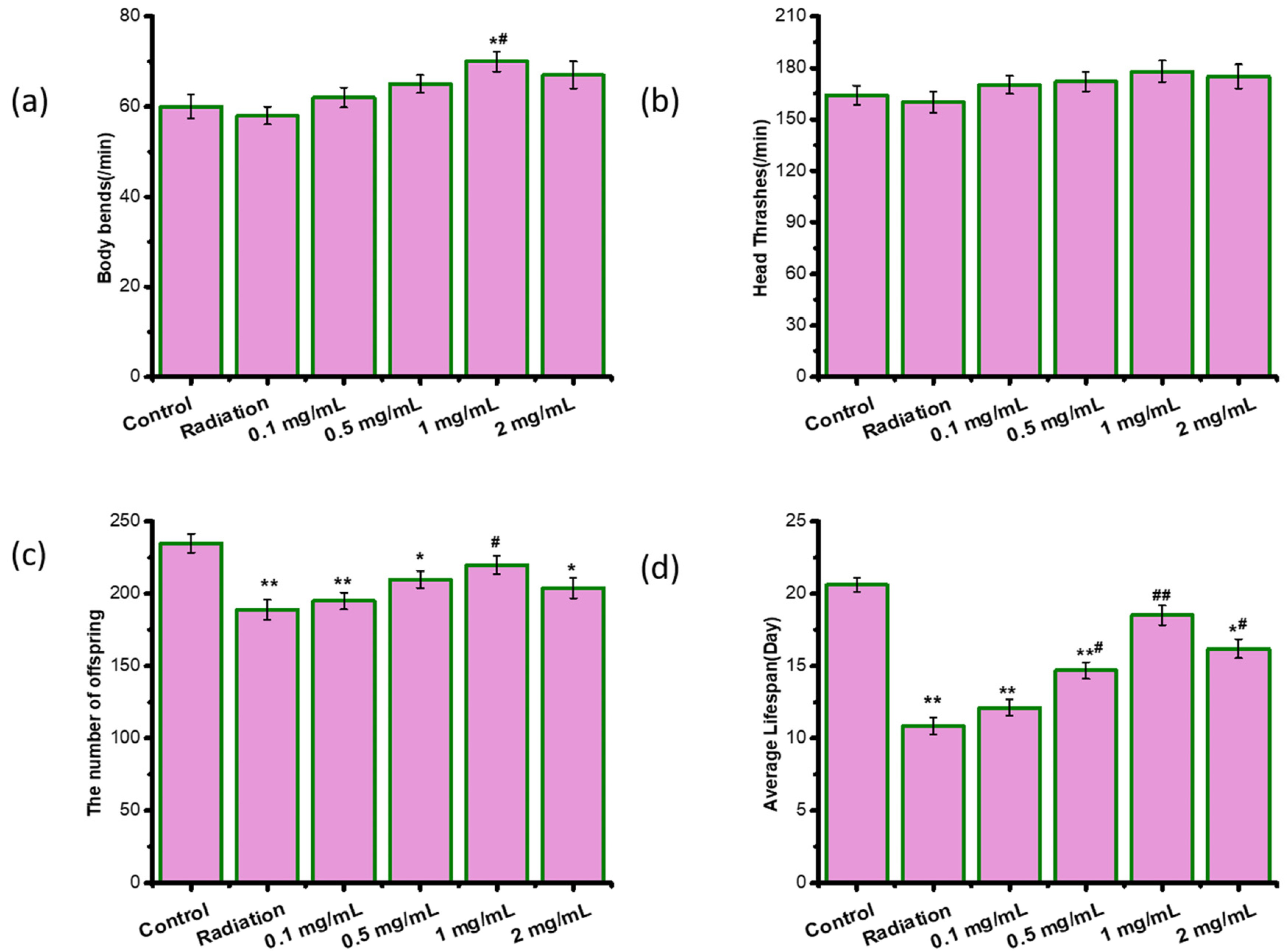
2.2. ASPS Did Not Cause Toxic Effects on Radiation-Damaged C. elegans
2.3. ASPS Protected Radiation-Damaged Nematodes against PA14 by Elevating Innate Immune Response
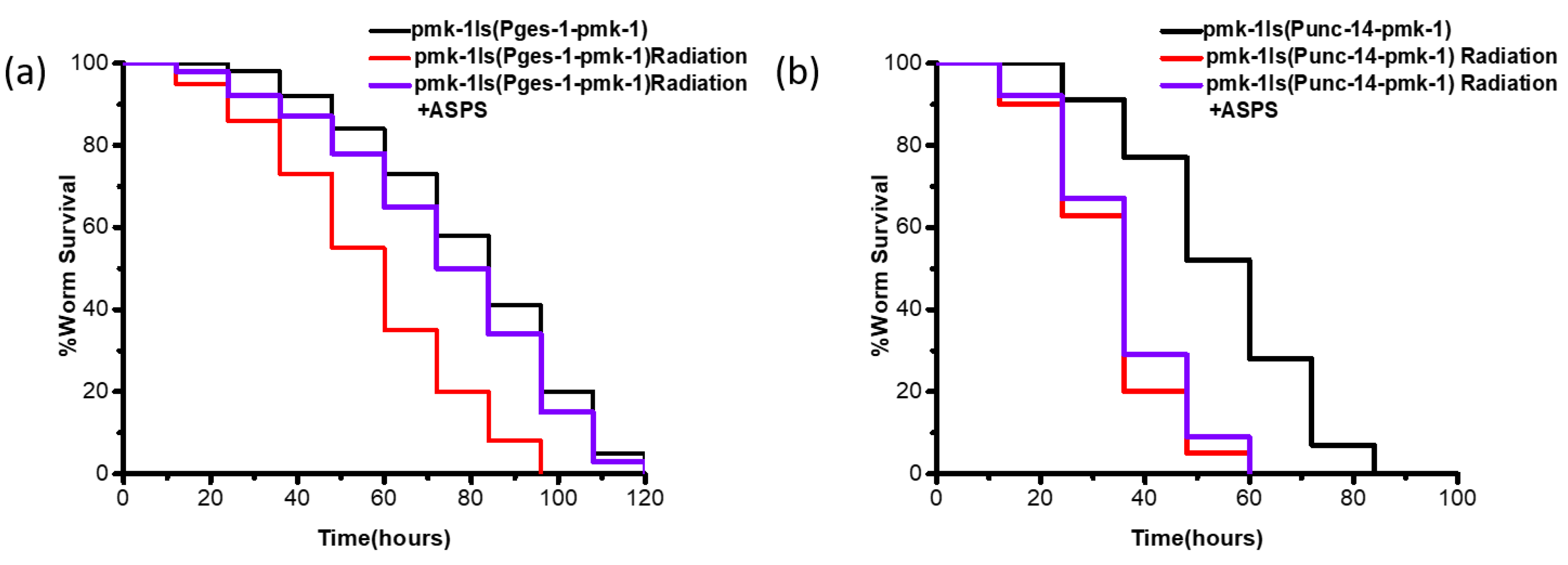
2.4. ASPS Promoted Innate Immunity of Radiation-Damaged Nematodes Through Intestinal p38 MAPK Pathway
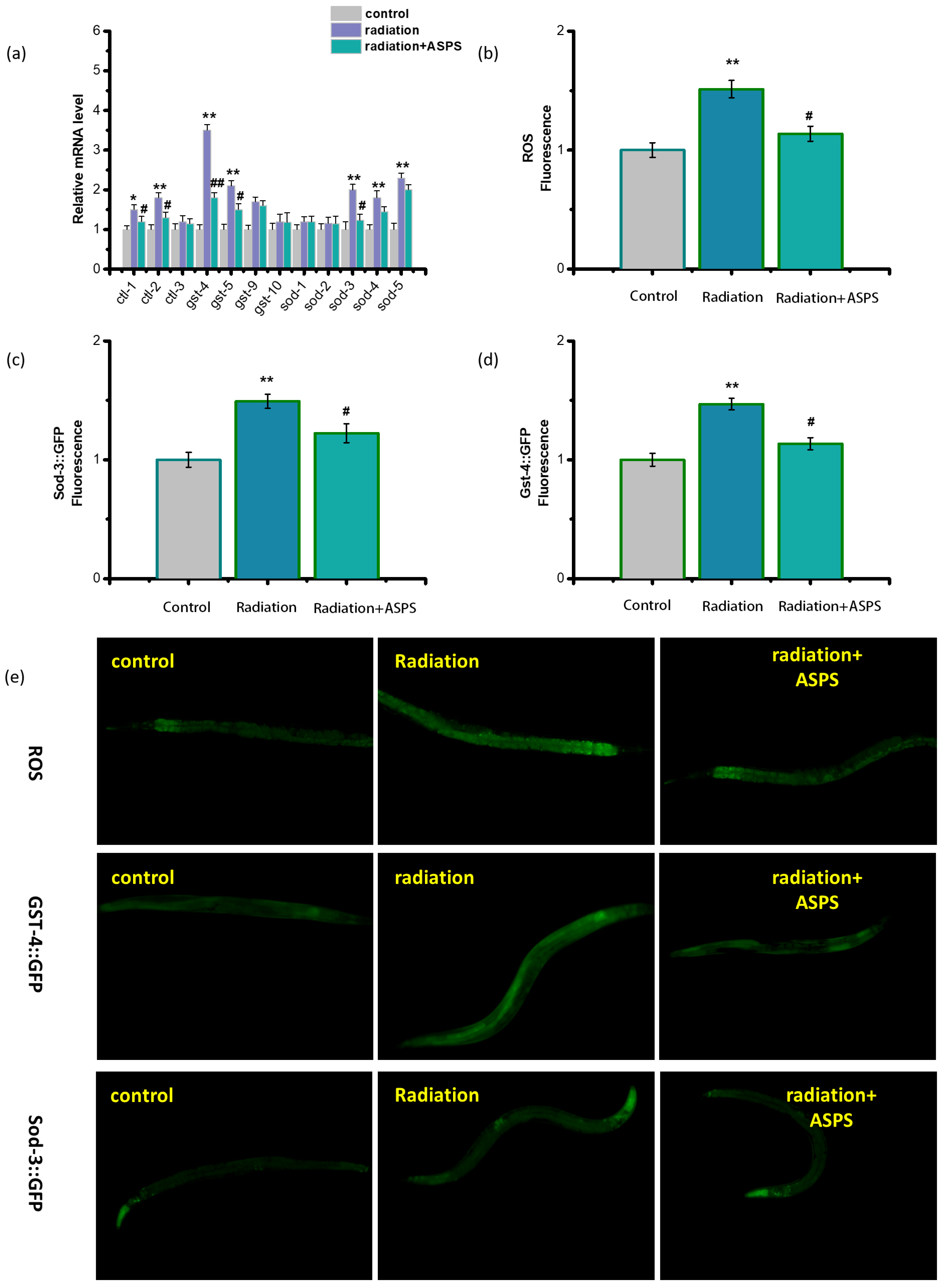
2.5. ASPS Affected Downstream Genes of p38 MAPK Signaling Pathway
2.6. ASPS Regulated Oxidative Stress
3. Discussion
4. Materials and Methods
4.1. Nematode Strains and Bacterial
4.2. Preparation of A. senticosus Extract
4.3. Radiation Treatment and the Usage of Extract
4.4. PA14 Infection Assay
4.5. Pharynx Pump Determination
4.6. Colony-Forming Unit (Cfu) Measurement
4.7. Protease Activity, Elastase Activity and Biofilm Assays
4.8. Measurement of ROS
4.9. Microscopic Imaging
4.10. Quantitative RT-PCR Assay
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reisz, J.A.; Bansal, N.; Qian, J.; Zhao, W.; Furdui, C.M. Effects of Ionizing Radiation on Biological Molecules - Mechanisms of Damage and Emerging Methods of Detection. Antioxidants Redox Signal. 2014, 21, 260–292. [Google Scholar] [CrossRef] [PubMed]
- Manda, K.; Glasow, A.; Paape, D.; Hildebrandt, G. Effects of Ionizing Radiation on the Immune System with Special Emphasis on the Interaction of Dendritic and T Cells. Front. Oncol. 2012, 2, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Xiong, Y.; Shan, S.; Zhu, Y.; Zeng, D.; Shi, Y.; Zhang, Y.; Lu, W. Eleutheroside E Enhances the Long-Term Memory of Radiation-Damaged C. elegans through G-Protein-Coupled Receptor and Neuropeptide Signaling Pathways. J. Nat. Prod. 2020, 83, 3315–3323. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Song, B.W.; Fu, C.Y.; Baranenko, D.D.; Wang, E.J.; Li, F.Y.; Lu, G.W. Acanthopanax Senticosus Reduces Brain Injury in Mice Exposed to Low Linear Energy Transfer Radiation. Biomed. Pharmacother. 2018, 99, 781–790. [Google Scholar] [CrossRef]
- Xia, Y.G.; Huang, Y.X.; Liang, J.; Kuang, H.X. Comparable Studies of Two Polysaccharides from Leaves of Acanthopanax Senticosus: Structure and Antioxidation. Int. J. Biol. Macromol. 2020, 147, 350–362. [Google Scholar] [CrossRef]
- Mahajan-Miklos, S.; Tan, M.W.; Rahme, L.G.; Ausubel, F.M. Molecular Mechanisms of Bacterial Virulence Elucidated Using a Pseudomonas Aeruginosa-Caenorhabditis Elegans Pathogenesis Model. Cell 1999, 96, 47–56. [Google Scholar] [CrossRef] [Green Version]
- Hwang, I.Y.; Koh, E.; Wong, A.; March, J.C.; Bentley, W.E.; Lee, Y.S.; Chang, M.W. Engineered Probiotic Escherichia Coli Can Eliminate and Prevent Pseudomonas Aeruginosa Gut Infection in Animal Models. Nat. Commun. 2017, 8, 15028. [Google Scholar] [CrossRef] [Green Version]
- Estes, K.A.; Dunbar, T.L.; Powell, J.R.; Ausubel, F.M.; Troemel, E.R. BZIP Transcription Factor Zip-2 Mediates an Early Response to Pseudomonas Aeruginosa Infection in Caenorhabditis Elegans. Proc. Natl. Acad. Sci. USA 2010, 107, 2153–2158. [Google Scholar] [CrossRef] [Green Version]
- Kandasamy, S.; Khan, W.; Evans, F.; Critchley, A.T.; Prithiviraj, B. Tasco®: A Product of Ascophyllum Nodosum Enhances Immune Response of Caenorhabditis Elegans against Pseudomonas Aeruginosa Infection. Mar. Drugs 2012, 10, 84–105. [Google Scholar] [CrossRef] [Green Version]
- Troemel, E.R.; Chu, S.W.; Reinke, V.; Lee, S.S.; Ausubel, F.M.; Kim, D.H. P38 MAPK Regulates Expression of Immune Response Genes and Contributes to Longevity in C. elegans. PLoS Genet. 2006, 2, 1725–1739. [Google Scholar] [CrossRef]
- Shivers, R.P.; Pagano, D.J.; Kooistra, T.; Richardson, C.E.; Reddy, K.C.; Whitney, J.K.; Kamanzi, O.; Matsumoto, K.; Hisamoto, N.; Kim, D.H. Phosphorylation of the Conserved Transcription Factor ATF-7 by PMK-1 P38 MAPK Regulates Innate Immunity in Caenorhabditis Elegans. PLoS Genet. 2010, 6, 1000892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colitti, M.; Stefanon, B.; Gabai, G.; Gelain, M.E.; Bonsembiante, F. Oxidative Stress and Nutraceuticals in the Modulation of the Immune Function: Current Knowledge in Animals of Veterinary Interest. Antioxidants 2019, 8, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Raamsdonk, J.M.; Hekimi, S. Deletion of the Mitochondrial Superoxide Dismutase Sod-2 Extends Lifespan in Caenorhabditis Elegans. PLoS Genet. 2009, 5, 1344–1350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Arriola, E.; Cárdenas-Rodríguez, N.; Coballase-Urrutia, E.; Pedraza-Chaverri, J.; Carmona-Aparicio, L.; Ortega-Cuellar, D. Caenorhabditis Elegans: A Useful Model for Studying Metabolic Disorders in Which Oxidative Stress Is a Contributing Factor. Oxid. Med. Cell. Longev. 2014, 18, 293–306. [Google Scholar] [CrossRef] [Green Version]
- Park, S.K.; Tedesco, P.M.; Johnson, T.E. Oxidative Stress and Longevity in Caenorhabditis Elegans as Mediated by SKN-1. Aging Cell 2009, 8, 258–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, M.W.; Rahme, L.G.; Sternberg, J.A.; Tompkins, R.G.; Ausubel, F.M. Pseudomonas Aeruginosa Killing of Caenorhabditis Elegans Used to Identify P. Aeruginosa Virulence Factors. Proc. Natl. Acad. Sci. USA 1999, 96, 2408–2413. [Google Scholar] [CrossRef] [Green Version]
- Mertenskötter, A.; Keshet, A.; Gerke, P.; Paul, R.J. The P38 MAPK PMK-1 Shows Heat-Induced Nuclear Translocation, Supports Chaperone Expression, and Affects the Heat Tolerance of Caenorhabditis Elegans. Cell Stress Chaperones 2013, 18, 293–306. [Google Scholar] [CrossRef] [Green Version]
- Tullet, J.M.A.; Hertweck, M.; An, J.H.; Baker, J.; Hwang, J.Y.; Liu, S.; Oliveira, R.P.; Baumeister, R.; Blackwell, T.K. Direct Inhibition of the Longevity-Promoting Factor SKN-1 by Insulin-like Signaling in C. elegans. Cell 2008, 132, 1025–1038. [Google Scholar] [CrossRef] [Green Version]
- Sakashita, T.; Takanami, T.; Yanase, S.; Hamada, N.; Suzuki, M.; Kimura, T.; Kobayashi, Y.; Ishii, N.; Higashitani, A. Radiation Biology of Caenorhabditis Elegans: Germ Cell Response, Aging and Behavior. J. Radiat. Res. 2010, 51, 107–121. [Google Scholar] [CrossRef] [Green Version]
- van Haaften, G.; Romeijn, R.; Pothof, J.; Koole, W.; Mullenders, L.H.F.; Pastink, A.; Plasterk, R.H.A.; Tijsterman, M. Identification of Conserved Pathways of DNA-Damage Response and Radiation Protection by Genome-Wide RNAi. Curr. Biol. 2006, 16, 1344–1350. [Google Scholar] [CrossRef] [Green Version]
- Schulenburg, H.; Kurz, C.L.; Ewbank, J.J. Evolution of the Innate Immune System: The Worm Perspective. Immunol. Rev. 2004, 198, 36–58. [Google Scholar] [CrossRef] [PubMed]
- JebaMercy, G.; Vigneshwari, L.; Balamurugan, K. A MAP Kinase Pathway in Caenorhabditis Elegans Is Required for Defense against Infection by Opportunistic Proteus Species. Microbes Infect. 2013, 15, 550–568. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Liu, F.; Li, S.; Jiang, N.; Yu, C.; Zhu, X.; Qin, Y.; Hui, J.; Meng, L.; Song, C.; et al. Metformin Promotes Innate Immunity through a Conserved PMK-1/P38 MAPK Pathway. Virulence 2020, 11, 39–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakabayashi, T.; Nakano, Y. Stress Responses Against Rare Earth Ions Are Mediated by the JNK and P38 MAPK Pathways in Caenorhabditis Elegans. Biol. Trace Elem. Res. 2019, 190, 550–555. [Google Scholar] [CrossRef]
- Li, L.; Chen, Y.; Chenzhao, C.; Fu, S.; Xu, Q.; Zhao, J. Glucose Negatively Affects Nrf2/SKN-1-Mediated Innate Immunity in C. elegans. Aging 2018, 10, 3089–3103. [Google Scholar] [CrossRef]
- Fu, J.; Yuan, J.; Tu, Y.; Fu, J.; Zhang, N.; Gao, B.; Fu, G.; Zhang, Y. A Polysaccharide from Acanthopanax Senticosus Improves the Antioxidant Status in Alloxan-Induced Diabetic Mice. Carbohydr. Polym. 2012, 88, 517–521. [Google Scholar] [CrossRef]
- Chen, M.; Yin, J.; Liang, Y.; Yuan, S.; Wang, F.; Song, M.; Wang, H. Oxidative Stress and Immunotoxicity Induced by Graphene Oxide in Zebrafish. Aquat. Toxicol. 2016, 174, 54–60. [Google Scholar] [CrossRef]
- Brenner, S. The Genetics of Caenorhabditis Elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef]
- Tan, M.W.; Mahajan-Miklos, S.; Ausubel, F.M. Killing of Caenorhabditis Elegans by Pseudomonas Aeruginosa Used to Model Mammalian Bacterial Pathogenesis. Proc. Natl. Acad. Sci. USA 1999, 96, 715–720. [Google Scholar] [CrossRef] [Green Version]
- Lazakovitch, E.; Kalb, J.M.; Gronostajski, R.M. Lifespan Extension and Increased Pumping Rate Accompany Pharyngeal Muscle-Specific Expression of Nfi-1 in C. elegans. Dev. Dyn. 2008, 237, 2100–2107. [Google Scholar] [CrossRef]
- Dow, J.M.; Clarker, B.; Milligan, D.E.; Tang, J.L.; Daniels, M.J. Extracellular Proteases from Xanthomonas Campestris Pv. Campestris, the Black Rot Pathogen. Appl. Environ. Microbiol. 1990, 56, 2994–2998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prithiviraj, B.; Bais, H.P.; Weir, T.; Suresh, B.; Najarro, E.H.; Dayakar, B.V.; Schweizer, H.P.; Vivanco, J.M. Down Regulation of Virulence Factors of Pseudomonas Aeruginosa by Salicylic Acid Attenuates Its Virulence on Arabidopsis Thaliana and Caenorhabditis Elegans. Infect. Immun. 2005, 73, 5319–5328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Toole, G.A. Microtiter Dish Biofilm Formation Assay. J. Vis. Exp. 2010, 47, e2437. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, M.; Li, N.; Shan, S.; Shi, Y.; Zhu, Y.; Lu, W. Acanthopanax senticosus Polysaccharide Enhances the Pathogen Resistance of Radiation-Damaged Caenorhabditis elegans through Intestinal p38 MAPK-SKN-1/ATF-7 Pathway and Stress Response. Int. J. Mol. Sci. 2022, 23, 5034. https://doi.org/10.3390/ijms23095034
Liu M, Li N, Shan S, Shi Y, Zhu Y, Lu W. Acanthopanax senticosus Polysaccharide Enhances the Pathogen Resistance of Radiation-Damaged Caenorhabditis elegans through Intestinal p38 MAPK-SKN-1/ATF-7 Pathway and Stress Response. International Journal of Molecular Sciences. 2022; 23(9):5034. https://doi.org/10.3390/ijms23095034
Chicago/Turabian StyleLiu, Mengyao, Nana Li, Shan Shan, Yudong Shi, Yuanbing Zhu, and Weihong Lu. 2022. "Acanthopanax senticosus Polysaccharide Enhances the Pathogen Resistance of Radiation-Damaged Caenorhabditis elegans through Intestinal p38 MAPK-SKN-1/ATF-7 Pathway and Stress Response" International Journal of Molecular Sciences 23, no. 9: 5034. https://doi.org/10.3390/ijms23095034
APA StyleLiu, M., Li, N., Shan, S., Shi, Y., Zhu, Y., & Lu, W. (2022). Acanthopanax senticosus Polysaccharide Enhances the Pathogen Resistance of Radiation-Damaged Caenorhabditis elegans through Intestinal p38 MAPK-SKN-1/ATF-7 Pathway and Stress Response. International Journal of Molecular Sciences, 23(9), 5034. https://doi.org/10.3390/ijms23095034