
Genome-Wide Identification and Characterization of the CC-NBS-LRR Gene Family in Cucumber (Cucumis sativus L.)

Abstract
:1. Introduction
2. Results
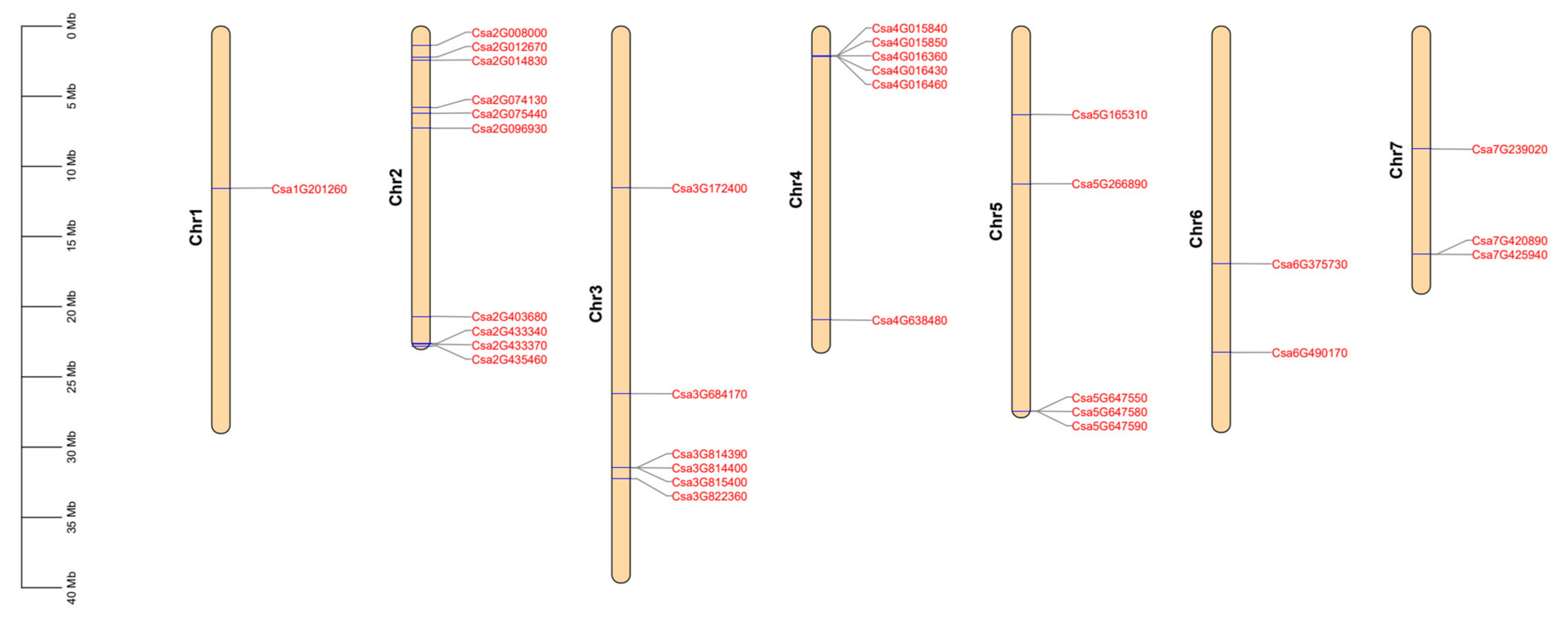
2.1. Identification and Characterization of the CNL Family of Genes in Cucumber Plants
2.2. Cis-Acting Element Analysis of the Promoters of the CNL Genes in Cucumber Plants
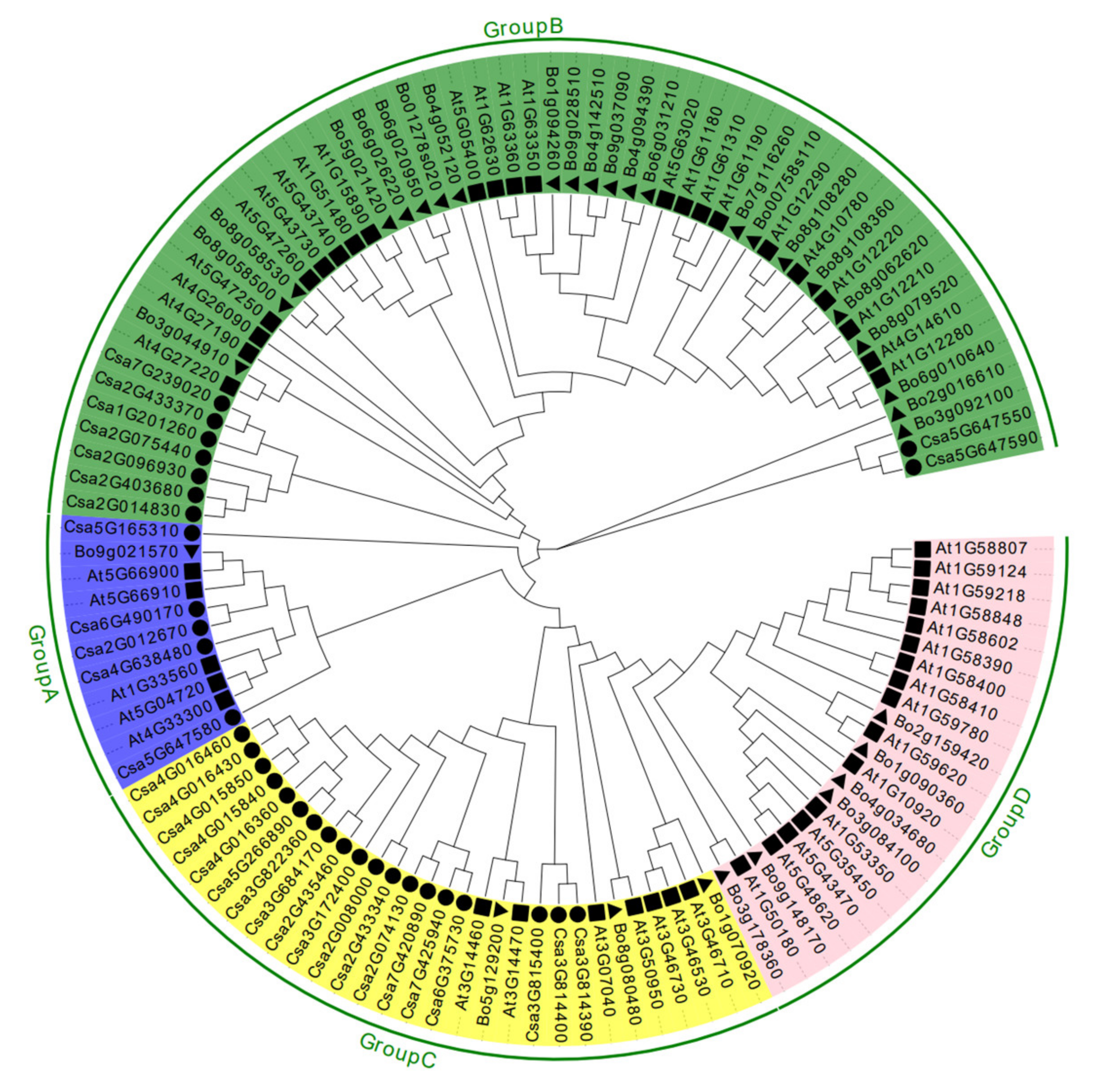
2.3. Analysis of the Phylogenetic Relationship and the Conserved Motifs of CNL Proteins in Cucumber Plants
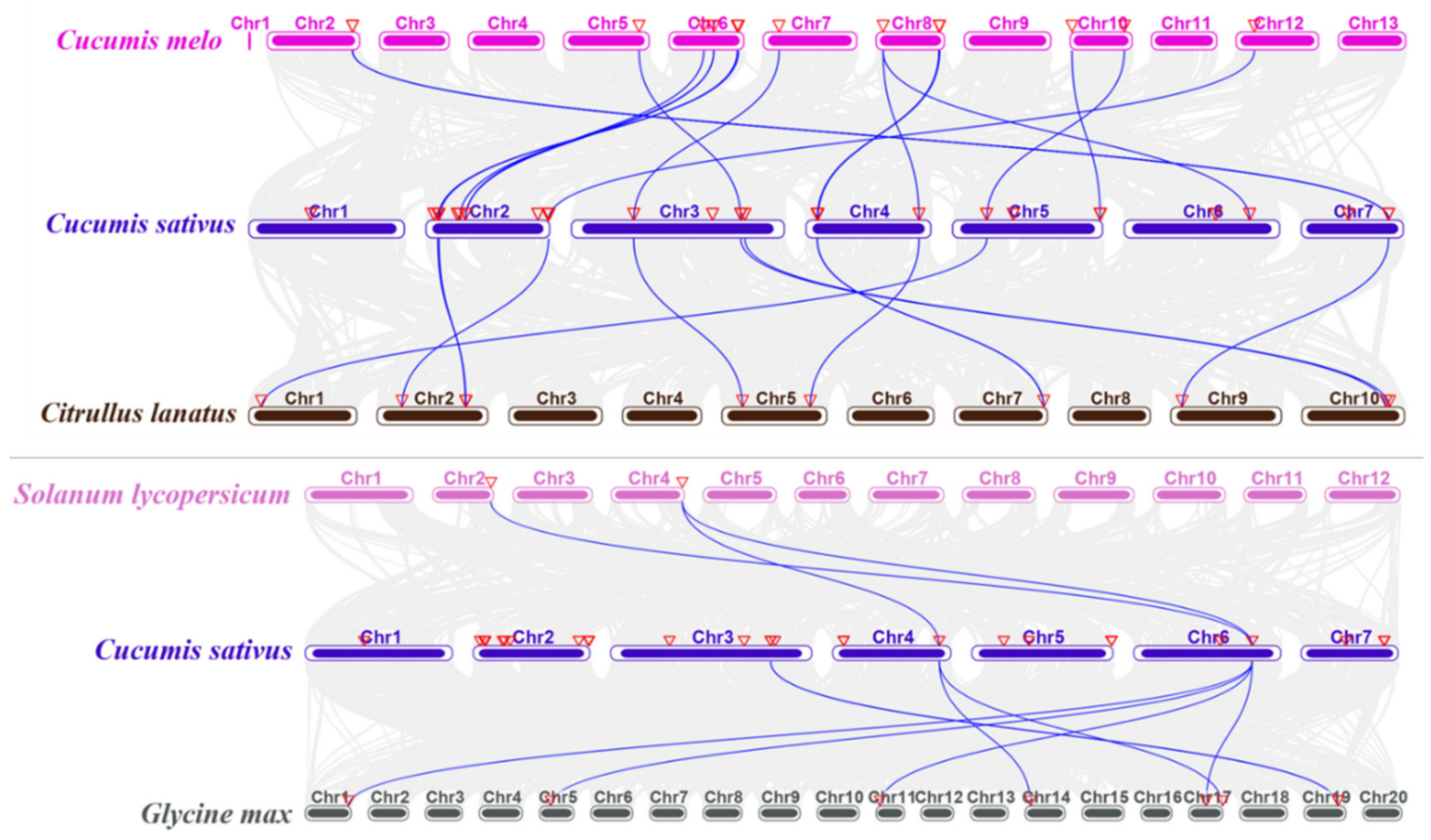
2.4. Synteny Analysis of the CNL Genes of Cucumber
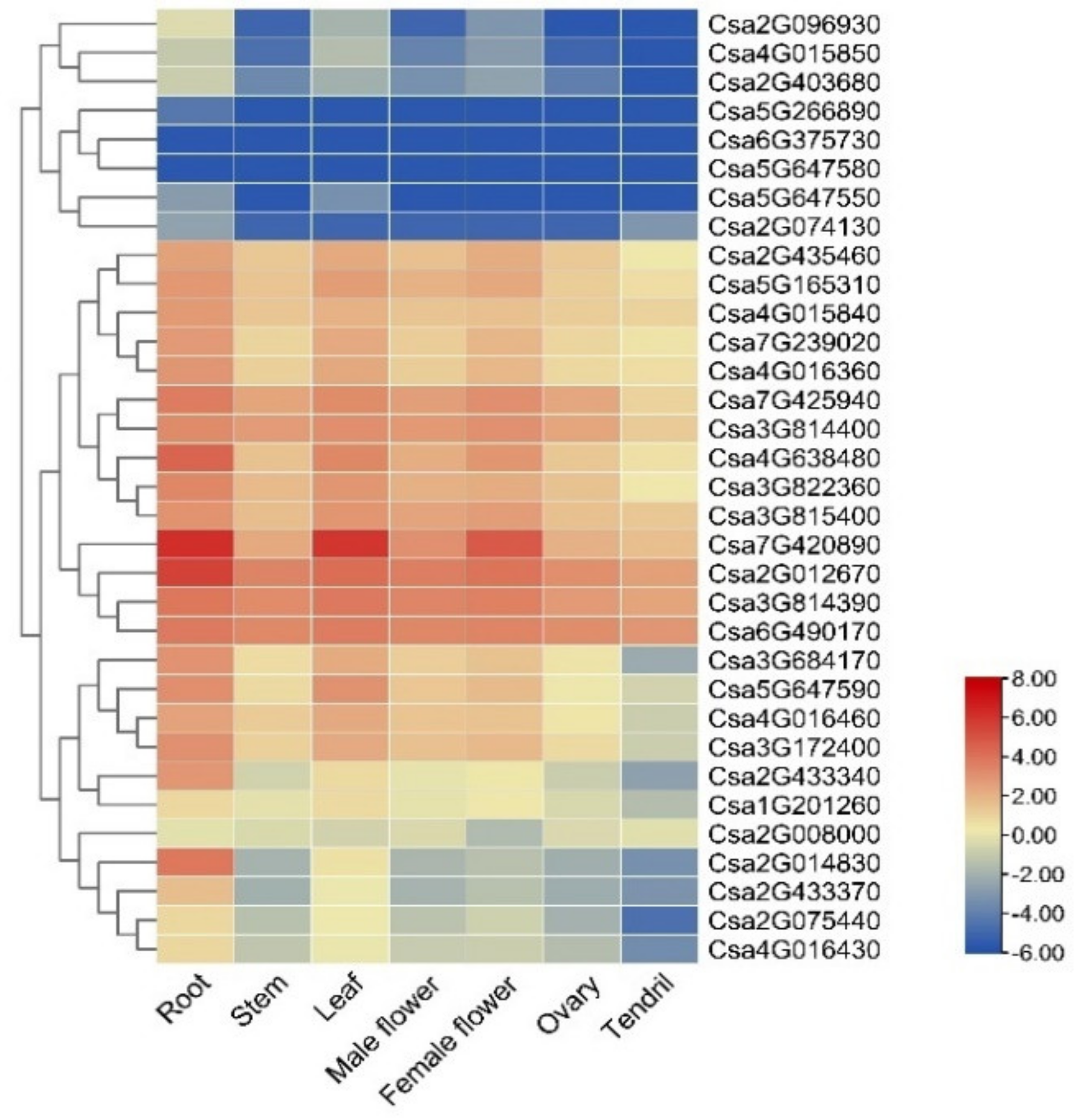
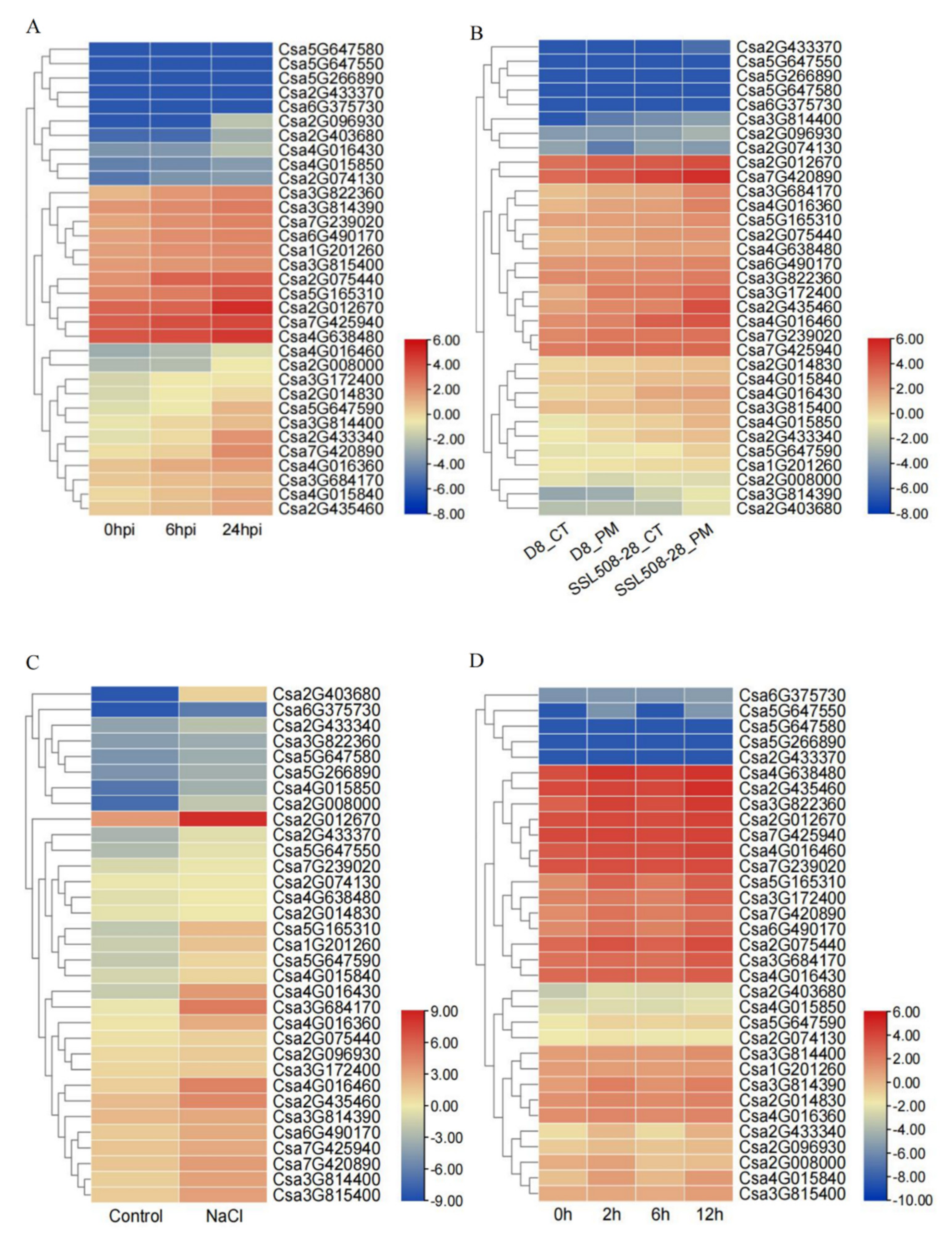
2.5. Expression Analysis of the CNL Family Genes in Cucumber Plants
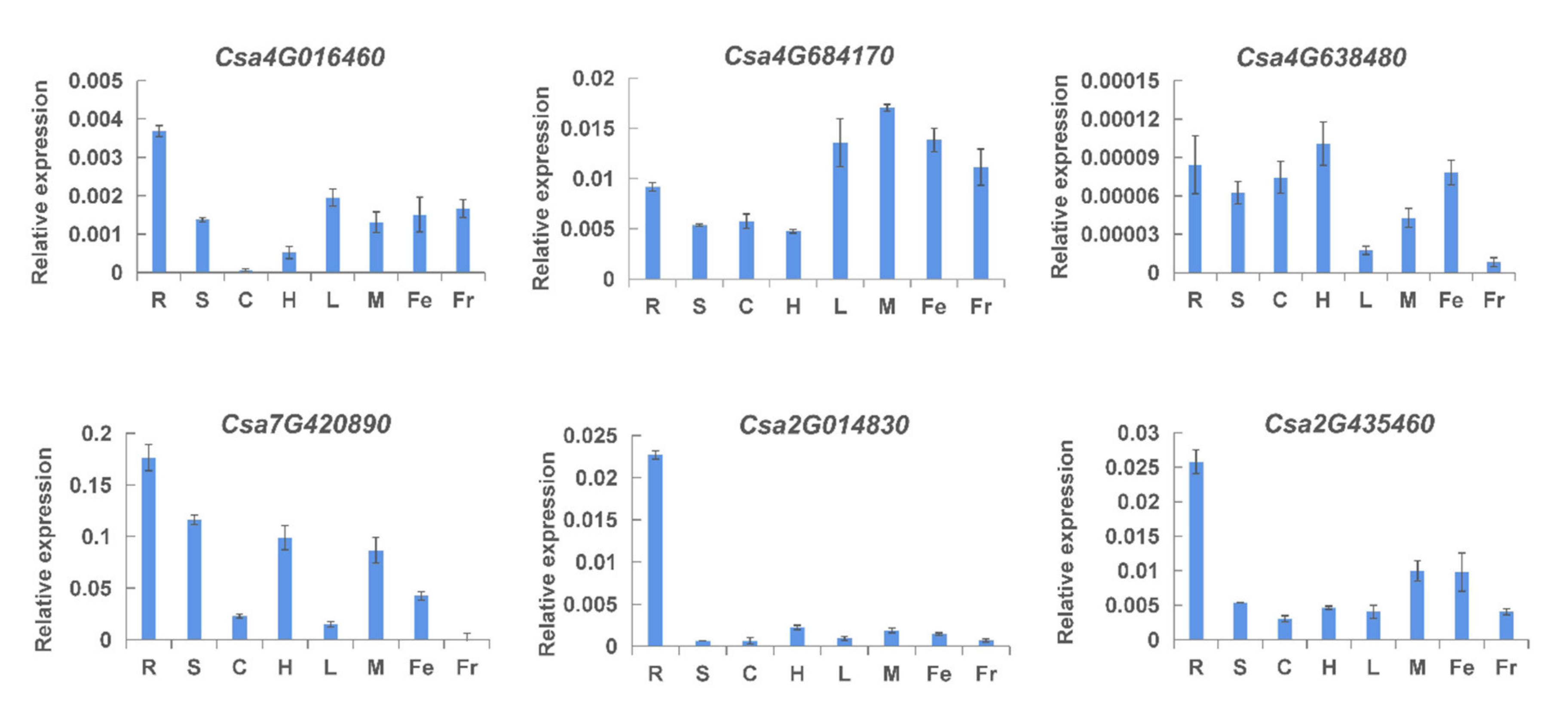
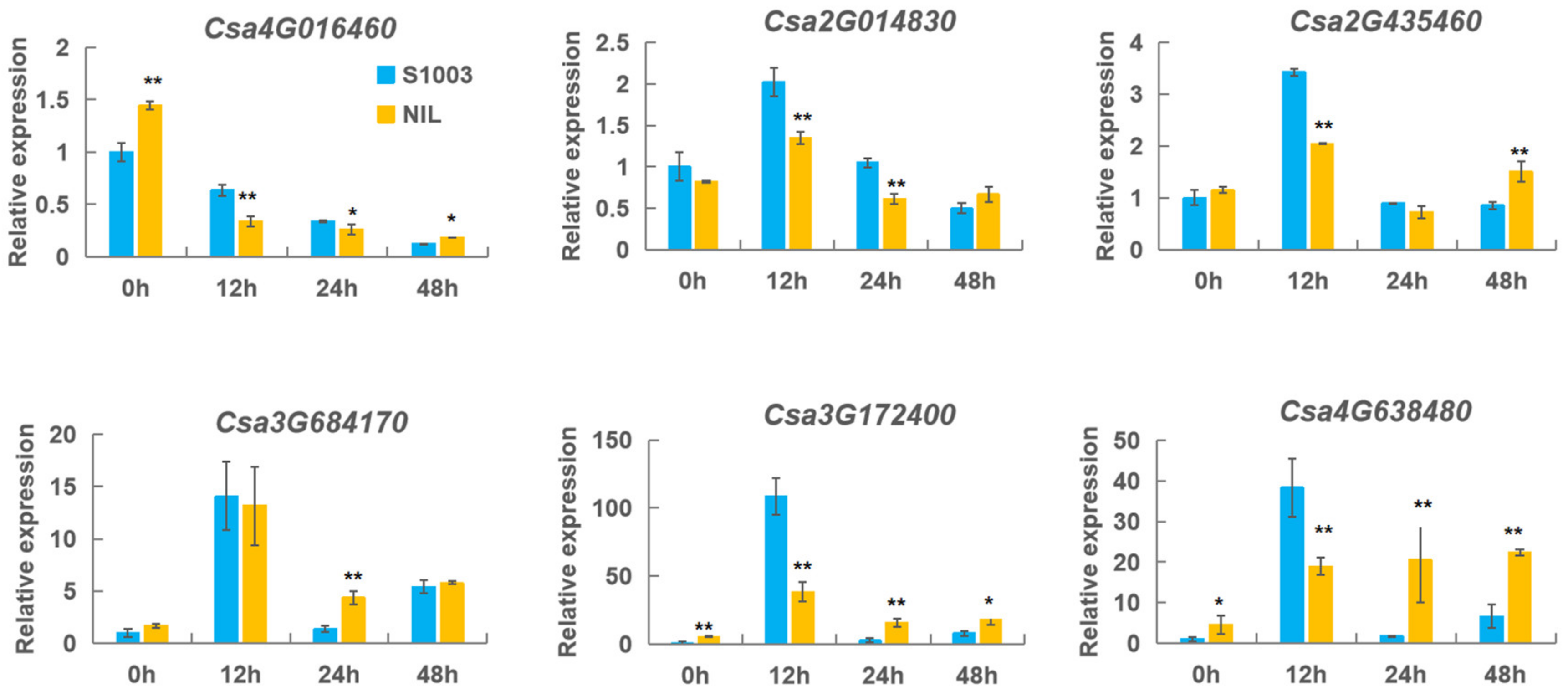
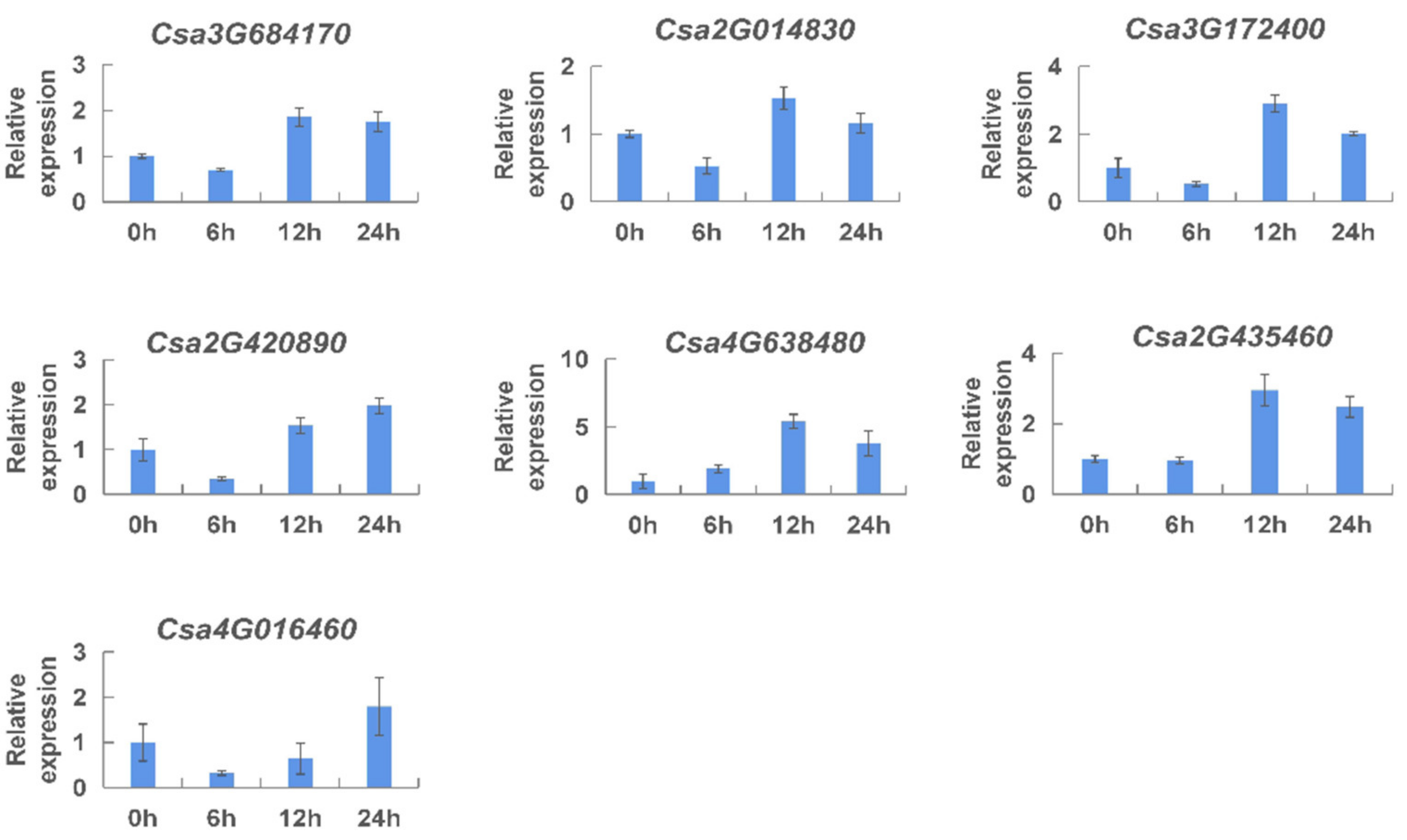
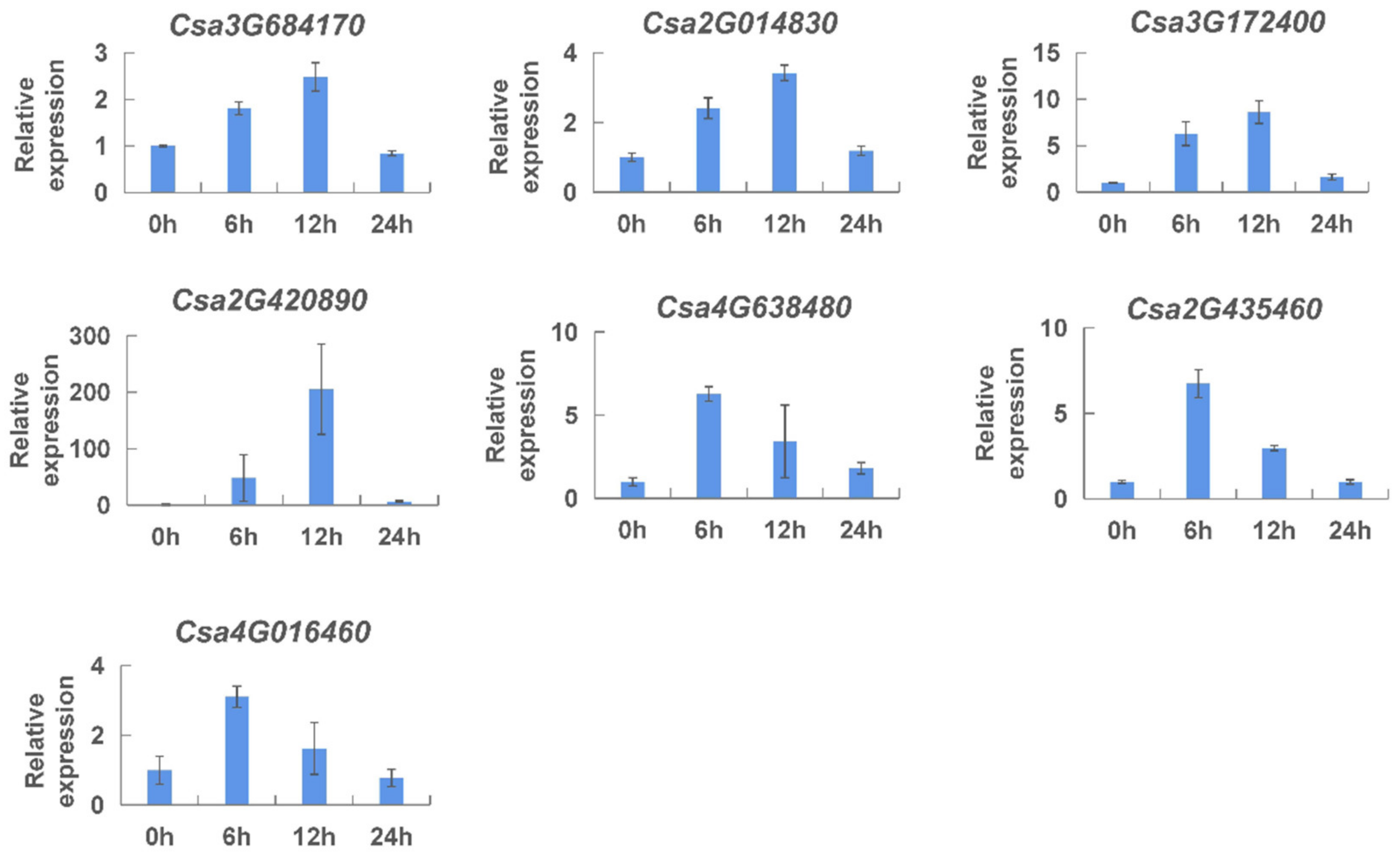
2.6. Analysis of the Expression of CNL Genes by qRT-PCR
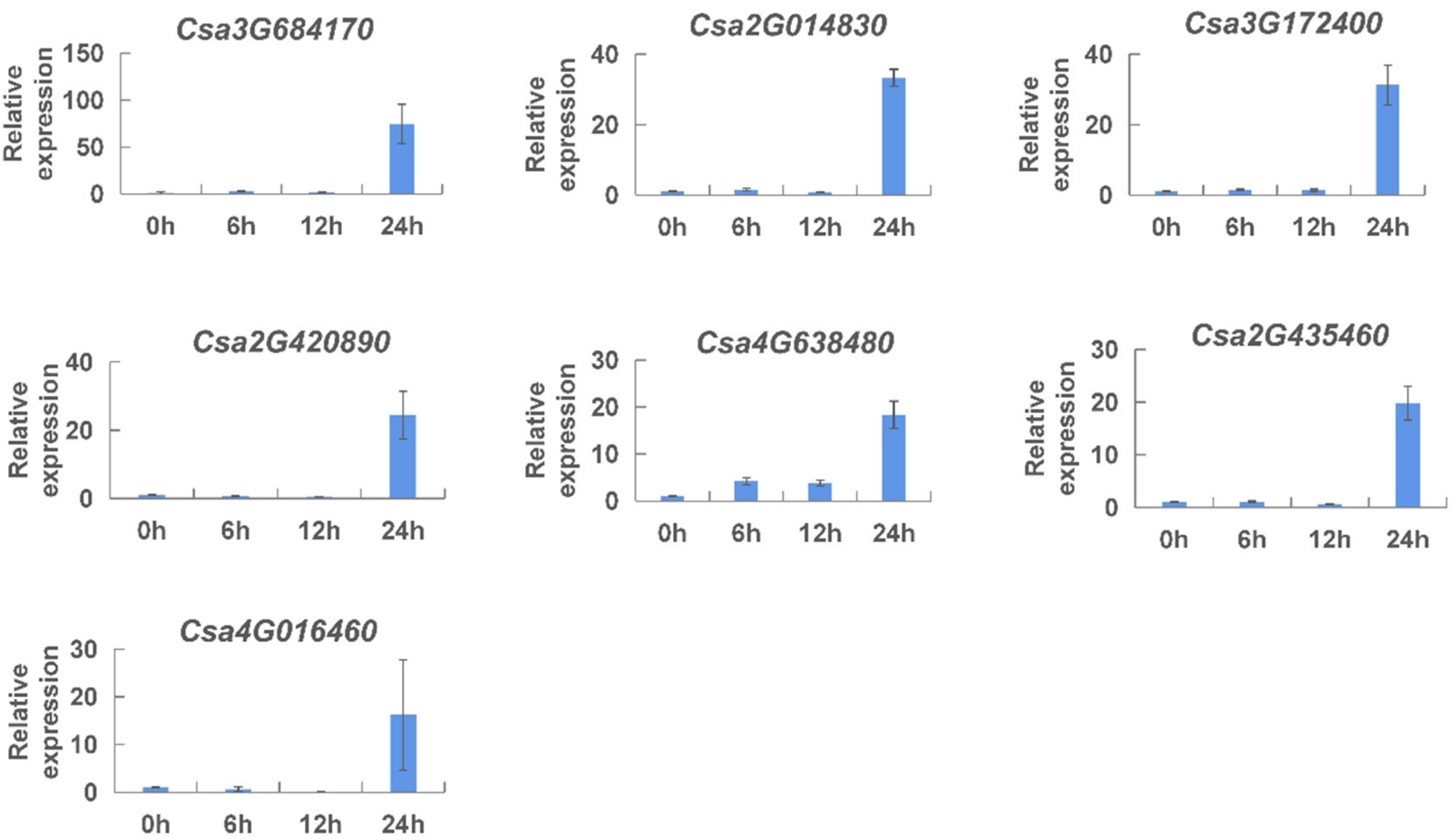
2.7. Expression Patterns of the CNL Genes Responding to Hormones in Cucumber Plants
2.8. Prediction Analysis of the Binding Site of CNL Genes Targeted by miRNA in Cucumber Plants
3. Discussion
4. Materials and Methods
4.1. Identification of Cucumber CNL Family Genes
4.2. Analysis of Gene Characteristics, Genomic Distribution, and Cis-Acting Elements in Promoters
4.3. Analysis of the Conserved Motif and the Synteny of the CNL
4.4. Transcriptome Analysis of the Genes Expression of the CNLs
4.5. Tissue-Expression Analysis
4.6. Cucumber Materials and the Treatment of PM and Hormones
4.7. Total RNA Extraction and qRT-PCR Analysis
4.8. Prediction of Binding Sites of the CNL Genes Targeted by miRNA
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, Y.; Li, D.; Yang, N.; Zhu, X.; Han, K.; Gu, R.; Bai, J.; Wang, A.; Zhang, Y. Genome-Wide Identification and Analysis of CC-NBS-LRR Family in Response to Downy Mildew and Black Rot in Chinese Cabbage. Int. J. Mol. Sci. 2021, 22, 4266. [Google Scholar] [CrossRef] [PubMed]
- Boller, T.; He, S.Y. Innate Immunity in Plants: An Arms Race Between Pattern Recognition Receptors in Plants and Effectors in Microbial Pathogens. Science 2009, 324, 742–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, M.; Ngou, B.P.M.; Ding, P.; Xin, X.-F. PTI-ETI crosstalk: An integrative view of plant immunity. Curr. Opin. Plant Biol. 2021, 62, 102030. [Google Scholar] [CrossRef] [PubMed]
- Van der Hoorn, R.A.L.; Kamoun, S. From Guard to Decoy: A New Model for Perception of Plant Pathogen Effectors. The Plant Cell 2008, 20, 2009–2017. [Google Scholar] [CrossRef] [Green Version]
- Yue, J.; Meyers, B.C.; Chen, J.; Tian, D.; Yang, S. Tracing the origin and evolutionary history of plant nucleotide-binding site–leucine-rich repeat (NBS-LRR) genes. New Phytologist 2012, 193, 1049–1063. [Google Scholar] [CrossRef]
- Jones, J.D.G.; Vance, R.E.; Dangl, J.L. Intracellular innate immune surveillance devices in plants and animals. Science 2016, 354, aaf6395. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Dong, S.; Gu, F.; Liu, W.; Yang, G.; Huang, M.; Xiao, W.; Liu, Y.; Guo, T.; Wang, H.; et al. NBS-LRR Protein Pik-H4 Interacts with OsBIHD1 to Balance Rice Blast Resistance and Growth by Coordinating Ethylene-Brassinosteroid Pathway. Front. Plant Sci. 2017, 8, 127. [Google Scholar] [CrossRef] [Green Version]
- Cai, H.; Wang, W.; Rui, L.; Han, L.; Luo, M.; Liu, N.; Tang, D. The TIR-NBS protein TN13 associates with the CC-NBS-LRR resistance protein RPS5 and contributes to RPS5-triggered immunity in Arabidopsis. Plant J 2021, 107, 775–786. [Google Scholar] [CrossRef]
- Chen, Y.; Zhong, G.; Cai, H.; Chen, R.; Liu, N.; Wang, W.; Tang, D. A Truncated TIR-NBS Protein TN10 Pairs with Two Clustered TIR-NBS-LRR Immune Receptors and Contributes to Plant Immunity in Arabidopsis. Int. J. Mol. Sci. 2021, 22, 4004. [Google Scholar] [CrossRef]
- Inoue, H.; Hayashi, N.; Matsushita, A.; Xinqiong, L.; Nakayama, A.; Sugano, S.; Jiang, C.-J.; Takatsuji, H. Blast resistance of CC-NB-LRR protein Pb1 is mediated by WRKY45 through protein–protein interaction. Proc. Natl. Acad. Sci. USA 2013, 110, 9577–9582. [Google Scholar] [CrossRef] [Green Version]
- Takken, F.L.; Albrecht, M.; Tameling, W.I. Resistance proteins: Molecular switches of plant defence. Curr. Opin. Plant Biol. 2006, 9, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Xie, J.; Wang, H.; Zhong, X.; Li, H.; Yu, J.; Kang, J. Identification and expression profiling analysis of NBS–LRR genes involved in Fusarium oxysporum f.sp. conglutinans resistance in cabbage. 3 Biotech 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Miller, R.N.G.; Bertioli, D.J.; Baurens, F.C.; Santos, C.M.R.; Alves, P.C.; Martins, N.F.; Togawa, R.C.; Souza Júnior, M.T.; Pappas Júnior, G.J. Analysis of non-TIR NBS-LRR resistance gene analogs in Musa acuminata Colla: Isolation, RFLP marker development, and physical mapping. BMC Plant Biology 2008, 8, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saraste, M.; Sibbald, P.R.; Wittinghofer, A. The P-loop—A common motif in ATP- and GTP-binding proteins. Trends Biochem. Sci. 1990, 15, 430–434. [Google Scholar] [CrossRef]
- Meyers, B.C.; Dickerman, A.W.; Michelmore, R.W.; Sivaramakrishnan, S.; Sobral, B.W.; Young, N.D. Plant disease resistance genes encode members of an ancient and diverse protein family within the nucleotide-binding superfamily. Plant J. 1999, 20, 317–332. [Google Scholar] [CrossRef]
- Lozano, R.; Ponce, O.P.; Ramírez, M.; Mostajo, N.; Orjeda, G. Genome-Wide Identification and Mapping of NBS-Encoding Resistance Genes in Solanum tuberosum Group Phureja. PLoS ONE 2012, 7, e34775. [Google Scholar] [CrossRef]
- Wulff, B.B.; Thomas, C.M.; Smoker, M.; Grant, M.; Jones, J.D. Domain swapping and gene shuffling identify sequences required for induction of an Avr-dependent hypersensitive response by the tomato Cf-4 and Cf-9 proteins. Plant Cell 2001, 13, 255–272. [Google Scholar] [CrossRef] [Green Version]
- Ray, S.K.; Macoy, D.M.; Kim, W.-Y.; Lee, S.Y.; Kim, M.G. Role of RIN4 in Regulating PAMP-Triggered Immunity and Effector-Triggered Immunity: Current Status and Future Perspectives. Mol. Cells 2019, 42, 503–511. [Google Scholar] [CrossRef]
- Andersen, E.J.; Ali, S.; Byamukama, E.; Yen, Y.; Nepal, M.P. Disease Resistance Mechanisms in Plants. Genes 2018, 9, 339. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Wu, X.; Sun, K.; Gao, Z. Structure and function analysis of a CC-NBS-LRR protein AT1G12290. Biochem. Biophys. Res. Commun. 2021, 534, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Wu, X.; Gao, Z. The RING-type protein BOI negatively regulates the protein level of a CC-NBS-LRR in Arabidopsis. Biochem. Biophys. Res. Commun. 2021, 578, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Si, Y.; Zheng, S.; Niu, J.; Tian, S.; Gu, M.; Lu, Q.; He, Y.; Zhang, J.; Shi, X.; Li, Y.; et al. Ne2, a typical CC–NBS–LRR-type gene, is responsible for hybrid necrosis in wheat. New Phytol. 2021, 232, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Fang, L.; Zhu, X.; Zhou, B.; Zhang, T. A CC-NBS-LRR gene induces hybrid lethality in cotton. J. Exp. Bot. 2019, 70, 5145–5156. [Google Scholar] [CrossRef]
- Li, T.; Zhang, Q.; Jiang, X.; Li, R.; Dhar, N. Cotton CC-NBS-LRR Gene GbCNL130 Confers Resistance to Verticillium Wilt Across Different Species. Front. Plant Sci. 2021, 12, 695691. [Google Scholar] [CrossRef]
- Li, N.-Y.; Zhou, L.; Zhang, D.-D.; Klosterman, S.J.; Li, T.-G.; Gui, Y.-J.; Kong, Z.-Q.; Ma, X.-F.; Short, D.P.G.; Zhang, W.-Q.; et al. Heterologous Expression of the Cotton NBS-LRR Gene GbaNA1 Enhances Verticillium Wilt Resistance in Arabidopsis. Front. Plant Sci. 2018, 9, 119. [Google Scholar] [CrossRef] [Green Version]
- Dogimont, C.; Chovelon, V.; Pauquet, J.; Boualem, A.; Bendahmane, A. The Vat locus encodes for a CC-NBS-LRR protein that confers resistance to Aphis gossypii infestation and A. gossypii-mediated virus resistance. Plant J. 2014, 80, 993–1004. [Google Scholar] [CrossRef]
- Ma, J.; Lei, C.; Xu, X.; Hao, K.; Wang, J.; Cheng, Z.; Ma, X.; Zhou, K.; Zhang, X.; Guo, X.; et al. Pi64, Encoding a Novel CC-NBS-LRR Protein, Confers Resistance to Leaf and Neck Blast in Rice. Mol. Plant-Microbe Interact. 2015, 28, 558–568. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Ji, L.; Zhu, B.; Yuan, X.; Jin, D.; Xie, G. OsCML16 interacts with a novel CC-NBS-LRR protein OsPi304 in the Ca(2+)/Mg(2+) dependent and independent manner in rice. Biochem Biophys Res Commun 2018, 504, 346–351. [Google Scholar] [CrossRef]
- Wang, Q.; Li, Y.; Kosami, K.; Liu, C.; Li, J.; Zhang, D.; Miki, D.; Kawano, Y. Three highly conserved hydrophobic residues in the predicted α2-helix of rice NLR protein Pit contribute to its localization and immune induction. Plant Cell Environ. 2022. [Google Scholar] [CrossRef]
- He, H.; Zhu, S.; Zhao, R.; Jiang, Z.; Ji, Y.; Ji, J.; Qiu, D.; Li, H.; Bie, T. Pm21, Encoding a Typical CC-NBS-LRR Protein, Confers Broad-Spectrum Resistance to Wheat Powdery Mildew Disease. Mol. Plant 2018, 11, 879–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Liu, L.; Wang, X.; Vossen, J.; Li, G.; Li, T.; Zheng, Z.; Gao, J.; Guo, Y.; Visser, R.G.F.; et al. The Ph-3 gene from Solanum pimpinellifolium encodes CC-NBS-LRR protein conferring resistance to Phytophthora infestans. Theor. Appl. Genet. 2014, 127, 1353–1364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Chen, Q.; Huang, J.; Meng, X.; Cui, N.; Yu, Y.; Fan, H. Nucleotide-Binding Leucine-Rich Repeat Genes CsRSF1 and CsRSF2 Are Positive Modulators in the Cucumis sativus Defense Response to Sphaerotheca fuliginea. Int. J. Mol. Sci. 2021, 22, 3986. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Li, R.; Zhang, Z.; Li, L.; Gu, X.; Fan, W.; Lucas, W.J.; Wang, X.; Xie, B.; Ni, P.; et al. The genome of the cucumber, Cucumis sativus L. Nat Genet 2009, 41, 1275–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Zhang, Z.; Yan, P.; Huang, S.; Fei, Z.; Lin, K. RNA-Seq improves annotation of protein-coding genes in the cucumber genome. BMC Genom. 2011, 12, 540. [Google Scholar] [CrossRef] [Green Version]
- Nie, J.; Wang, Y.; He, H.; Guo, C.; Zhu, W.; Pan, J.; Li, D.; Lian, H.; Pan, J.; Cai, R. Loss-of-Function Mutations in CsMLO1 Confer Durable Powdery Mildew Resistance in Cucumber (Cucumis sativus L.). Front Plant Sci 2015, 6, 1155. [Google Scholar] [CrossRef]
- Zhang, Y.; Waseem, M.; Zeng, Z.; Xu, J.; Chen, C.; Liu, Y.; Zhai, J.; Xia, R. MicroRNA482/2118, a miRNA superfamily essential for both disease resistance and plant development. New Phytol. 2021, 233, 2047–2057. [Google Scholar] [CrossRef]
- Richly, E.; Kurth, J.; Leister, D. thaliana Evolution. Mol. Biol. Evol. 2002, 19, 76–84. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Q.-H.; Fan, L.; Liu, Y.; Xu, H.; Llewellyn, D.; Wilson, I. miR482 Regulation of NBS-LRR Defense Genes during Fungal Pathogen Infection in Cotton. PLoS ONE 2013, 8, e84390. [Google Scholar] [CrossRef] [Green Version]
- Nie, J.; Wang, H.; Zhang, W.; Teng, X.; Yu, C.; Cai, R.; Wu, G. Characterization of lncRNAs and mRNAs Involved in Powdery Mildew Resistance in Cucumber. Phytopathology 2021, 111, 1613–1624. [Google Scholar] [CrossRef]
- Zhou, T.; Wang, Y.; Chen, J.-Q.; Araki, H.; Jing, Z.; Jiang, K.; Shen, J.; Tian, D. Genome-wide identification of NBS genes in japonica rice reveals significant expansion of divergent non-TIR NBS-LRR genes. Mol. Genet. Genom. 2004, 271, 402–415. [Google Scholar] [CrossRef] [PubMed]
- Andolfo, G.; Sanseverino, W.; Rombauts, S.; Peer, Y.; Bradeen, J.M.; Carputo, D.; Frusciante, L.; Ercolano, M.R. Overview of tomato (Solanum lycopersicum) candidate pathogen recognition genes reveals importantSolanum R locus dynamics. New Phytol. 2012, 197, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Jupe, F.; Pritchard, L.; Etherington, G.J.; MacKenzie, K.; Cock, P.J.; Wright, F.; Sharma, S.K.; Bolser, D.; Bryan, G.J.; Jones, J.D.; et al. Identification and localisation of the NB-LRR gene family within the potato genome. BMC Genom. 2012, 13, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.; Li, X.; Jiang, H.; Ma, W.; Miao, W.; Yamada, T.; Zhang, M. Systematic analysis and comparison of nucleotide-binding site disease resistance genes in maize. FEBS J. 2012, 279, 2431–2443. [Google Scholar] [CrossRef] [PubMed]
- Kuang, H.; Woo, S.-S.; Meyers, B.; Nevo, E.; Michelmore, R.W. Multiple Genetic Processes Result in Heterogeneous Rates of Evolution within the Major Cluster Disease Resistance Genes in Lettuce. Plant Cell 2004, 16, 2870–2894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chin, D.B.; Arroyo-Garcia, R.; Ochoa, O.E.; Kesseli, R.V.; Lavelle, D.O.; Michelmore, R.W. Recombination and spontaneous mutation at the major cluster of resistance genes in lettuce (Lactuca sativa). Genetics 2001, 157, 831–849. [Google Scholar] [CrossRef]
- Ma, Y.; Chhapekar, S.S.; Lu, L.; Oh, S.; Singh, S.; Kim, C.S.; Kim, S.; Choi, G.J.; Lim, Y.P.; Choi, S.R. Genome-wide identification and characterization of NBS-encoding genes in Raphanus sativus L. and their roles related to Fusarium oxysporum resistance. BMC Plant Biol. 2021, 21, 1–17. [Google Scholar] [CrossRef]
- Wang, T.; Jia, Z.-H.; Zhang, J.-Y.; Liu, M.; Guo, Z.-R.; Wang, G. Identification and Analysis of NBS-LRR Genes in Actinidia chinensis Genome. Plants 2020, 9, 1350. [Google Scholar] [CrossRef]
- Mohr, T.J.; Mammarella, N.D.; Hoff, T.; Woffenden, B.J.; Jelesko, J.G.; McDowell, J.M. The Arabidopsis Downy Mildew Resistance Gene RPP8 Is Induced by Pathogens and Salicylic Acid and Is Regulated by W Box cis Elements. Mol. Plant-Microbe Interact. 2010, 23, 1303–1315. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Zhang, Y.; Kuang, H.; Chen, J. Frequent loss of lineages and deficient duplications accounted for low copy number of disease resistance genes in Cucurbitaceae. BMC Genom. 2013, 14, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korves, T.; Bergelson, J. A novel cost of R gene resistance in the presence of disease. Am. Nat. 2004, 163, 489–504. [Google Scholar] [CrossRef]
- Ariga, H.; Katori, T.; Tsuchimatsu, T.; Hirase, T.; Tajima, Y.; Parker, J.E.; Alcazar, R.; Koornneef, M.; Hoekenga, O.; Lipka, A.E.; et al. NLR locus-mediated trade-off between abiotic and biotic stress adaptation in Arabidopsis. Nat. Plants 2017, 3, 17072. [Google Scholar] [CrossRef] [Green Version]
- Yuan, M.; Jiang, Z.; Bi, G.; Nomura, K.; Liu, M.; Wang, Y.; Cai, B.; Zhou, J.-M.; He, S.Y.; Xin, X.-F. Pattern-recognition receptors are required for NLR-mediated plant immunity. Nature 2021, 592, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Wu, G. Plant MicroRNAs and Development. J. Genet. Genom. 2013, 40, 217–230. [Google Scholar] [CrossRef]
- Wu, G.; Park, M.Y.; Conway, S.R.; Wang, J.-W.; Weigel, D.; Poethig, R.S. The Sequential Action of miR156 and miR172 Regulates Developmental Timing in Arabidopsis. Cell 2009, 138, 750–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Wu, J.; Zhang, Y.; Zhao, Y.; Xu, W.; Li, Y.; Xie, J. Genome-Wide Analysis of Coding and Non-coding RNA Reveals a Conserved miR164-NAC-mRNA Regulatory Pathway for Disease Defense in Populus. Front. Genet. 2021, 12, 668940. [Google Scholar] [CrossRef] [PubMed]
- Yao, S.; Yang, Z.; Yang, R.; Huang, Y.; Guo, G.; Kong, X.; Lan, Y.; Zhou, T.; Wang, H.; Wang, W.; et al. Transcriptional Regulation of miR528 by OsSPL9 Orchestrates Antiviral Response in Rice. Mol. Plant 2019, 12, 1114–1122. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Cao, Y.; Li, T.; Wang, X.; Chen, J.; He, H.; Yao, W.; Wu, J.; Zhang, H. MicroRNAs Are Involved in Maize Immunity Against Fusarium verticillioides Ear Rot. Genom. Proteom. Bioinform. 2020, 18, 241–255. [Google Scholar] [CrossRef]
- Xu, T.; Zhang, L.; Yang, Z.; Wei, Y.; Dong, T. Identification and Functional Characterization of Plant MiRNA Under Salt Stress Shed Light on Salinity Resistance Improvement Through MiRNA Manipulation in Crops. Front. Plant Sci. 2021, 12, 665439. [Google Scholar] [CrossRef]
- Zhang, L.; Ding, H.; Jiang, H.; Wang, H.; Chen, K.; Duan, J.; Feng, S.; Wu, G. Regulation of cadmium tolerance and accumulation by miR156 in Arabidopsis. Chemosphere 2020, 242, 125168. [Google Scholar] [CrossRef] [PubMed]
- Chandran, V.; Wang, H.; Gao, F.; Cao, X.-L.; Chen, Y.-P.; Li, G.-B.; Zhu, Y.; Yang, X.-M.; Zhang, L.-L.; Zhao, Z.; et al. miR396-OsGRFs Module Balances Growth and Rice Blast Disease-Resistance. Front. Plant Sci. 2019, 9, 1999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Vries, S.; Kukuk, A.; von Dahlen, J.K.; Schnake, A.; Kloesges, T.; Rose, L.E. Expression profiling across wild and cultivated tomatoes supports the relevance of early miR482/2118 suppression for Phytophthora resistance. Proc. Biol. Sci. 2018, 285, 1873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, Y.; Han, J.; Zhou, G.; Xu, Y.; Ding, Y.; Shi, M.; Guo, C.; Wu, G. Silencing of miR156 confers enhanced resistance to brown planthopper in rice. Planta 2018, 248, 813–826. [Google Scholar] [CrossRef]
- Shivaprasad, P.V.; Chen, H.-M.; Patel, K.; Bond, D.; Santos, B.; Baulcombe, D.C. A MicroRNA Superfamily Regulates Nucleotide Binding Site–Leucine-Rich Repeats and Other mRNAs. Plant Cell 2012, 24, 859–874. [Google Scholar] [CrossRef] [Green Version]
- Meyers, B.C.; Kozik, A.; Griego, A.; Kuang, H.; Michelmore, R.W. Genome-Wide Analysis of NBS-LRR–Encoding Genes in Arabidopsis. Plant Cell 2003, 15, 809–834. [Google Scholar] [CrossRef] [Green Version]
- Fiumara, F.; Fioriti, L.; Kandel, E.R.; Hendrickson, W.A. Essential Role of Coiled Coils for Aggregation and Activity of Q/N-Rich Prions and PolyQ Proteins. Cell 2010, 143, 1121–1135. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Xu, X.; Shi, Y.; Qi, X.; Chen, X. Elucidation of the molecular responses of a cucumber segment substitution line carrying Pm5.1 and its recurrent parent triggered by powdery mildew by comparative transcriptome profiling. BMC Genom. 2017, 18, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Nie, J.; He, H.; Peng, J.; Yang, X.; Wang, Y.; Bie, B.; Zhao, J.; Wang, Y.; Si, L.; Pan, J.; et al. Identification and fine mapping of pm5.1: A recessive gene for powdery mildew resistance in cucumber (Cucumis sativus L.). Mol. Breed. 2015, 35, 7. [Google Scholar] [CrossRef]
- Dai, X.; Zhuang, Z.; Zhao, P.X. psRNATarget: A plant small RNA target analysis server (2017 release). Nucleic Acids Res. 2018, 46, W49–W54. [Google Scholar] [CrossRef] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Length of CDS (bp) | Number of Amino Acids | Number of Exons | Molecular Weight (kDa) | Theoretical pI | Subcellular Localization |
|---|---|---|---|---|---|---|
| Csa1G201260 Csa2G008000 | 2409 2469 | 803 823 | 7 5 | 90.94932 95.32828 | 5.72 6.80 | N/Cyto Cyto/N |
| Csa2G012670 | 2439 | 813 | 5 | 92.46041 | 7.93 | N |
| Csa2G014830 | 5403 | 1801 | 7 | 207.11112 | 5.57 | N |
| Csa2G074130 | 2745 | 915 | 2 | 105.95585 | 8.29 | N |
| Csa2G075440 | 4056 | 1352 | 4 | 152.92906 | 6.35 | N/Cyto |
| Csa2G096930 | 2913 | 971 | 1 | 110.91379 | 6.20 | N/Cyto |
| Csa2G403680 | 3966 | 1322 | 4 | 151.81125 | 6.54 | N |
| Csa2G433340 | 2898 | 966 | 2 | 111.92962 | 8.38 | N/Cyto |
| Csa2G433370 | 3579 | 1193 | 4 | 136.25234 | 6.98 | N |
| Csa2G435460 | 2721 | 907 | 2 | 104.05611 | 7.23 | N |
| Csa3G172400 | 3444 | 1148 | 1 | 131.34413 | 7.38 | N/Cyto |
| Csa3G684170 | 3267 | 1089 | 1 | 125.61899 | 6.10 | Cyto/N |
| Csa3G814390 | 2409 | 803 | 1 | 93.33195 | 9.15 | N |
| Csa3G814400 | 2466 | 822 | 1 | 94.42609 | 7.10 | N |
| Csa3G815400 | 2676 | 892 | 1 | 103.11633 | 6.43 | N |
| Csa3G822360 | 3159 | 1053 | 2 | 120.24954 | 6.27 | N |
| Csa4G015840 | 3261 | 1087 | 1 | 124.35665 | 6.84 | Cyto/N |
| Csa4G015850 | 3240 | 1080 | 1 | 123.31024 | 7.36 | N/Cyto |
| Csa4G016360 | 2874 | 958 | 2 | 109.57171 | 7.09 | N |
| Csa4G016430 | 2520 | 840 | 3 | 96.12670 | 6.14 | Cyto/N |
| Csa4G016460 | 2841 | 947 | 2 | 108.23233 | 7.33 | N |
| Csa4G638480 | 2463 | 821 | 5 | 94.24655 | 6.58 | N |
| Csa5G165310 | 879 | 293 | 1 | 32.98491 | 6.80 | Cyto |
| Csa5G266890 | 3141 | 1047 | 1 | 119.3791 | 6.04 | Cyto/N |
| Csa5G647550 | 3213 | 1071 | 6 | 121.78672 | 6.70 | N |
| Csa5G647580 | 1143 | 381 | 3 | 44.46930 | 8.70 | N/Cyto |
| Csa5G647590 | 1884 | 628 | 7 | 71.53999 | 5.97 | N |
| Csa6G375730 | 2751 | 917 | 2 | 106.78455 | 8.21 | N |
| Csa6G490170 | 2454 | 818 | 5 | 93.59007 | 6.05 | N |
| Csa7G239020 | 3621 | 1207 | 2 | 138.19007 | 7.59 | N/Cyto |
| Csa7G420890 | 1665 | 555 | 2 | 64.71685 | 7.61 | N |
| Csa7G425940 | 3060 | 1020 | 5 | 116.40068 | 6.86 | N |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Yuan, Q.; Wu, Y.; Zhang, J.; Nie, J. Genome-Wide Identification and Characterization of the CC-NBS-LRR Gene Family in Cucumber (Cucumis sativus L.). Int. J. Mol. Sci. 2022, 23, 5048. https://doi.org/10.3390/ijms23095048
Zhang W, Yuan Q, Wu Y, Zhang J, Nie J. Genome-Wide Identification and Characterization of the CC-NBS-LRR Gene Family in Cucumber (Cucumis sativus L.). International Journal of Molecular Sciences. 2022; 23(9):5048. https://doi.org/10.3390/ijms23095048
Chicago/Turabian StyleZhang, Wanlu, Qi Yuan, Yiduo Wu, Jing Zhang, and Jingtao Nie. 2022. "Genome-Wide Identification and Characterization of the CC-NBS-LRR Gene Family in Cucumber (Cucumis sativus L.)" International Journal of Molecular Sciences 23, no. 9: 5048. https://doi.org/10.3390/ijms23095048
APA StyleZhang, W., Yuan, Q., Wu, Y., Zhang, J., & Nie, J. (2022). Genome-Wide Identification and Characterization of the CC-NBS-LRR Gene Family in Cucumber (Cucumis sativus L.). International Journal of Molecular Sciences, 23(9), 5048. https://doi.org/10.3390/ijms23095048