
Enhancement of Farnesoid X Receptor Inhibits Migration, Adhesion and Angiogenesis through Proteasome Degradation and VEGF Reduction in Bladder Cancers

 , , ,
, , , 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Survival Rate and Expressions of FXR in Bladder Cancer Patients and Bladder Cancer Cell Lines
2.2. Overexpression of FXR Inhibited Survival and Colony Formation in T24 Cells
2.3. Overexpression of FXR Inhibited the Migration and Adhesion Abilities in TSGH8301 and T24 Cells
2.4. FXR Overexpression Inhibits Migratory and Adhesive Ability via Proteosome Degradation
2.5. Overexpression of FXR Inhibited the Invasive Ability in the T24 Cells
2.6. Proteosome Degradation Was Involved in FXR Overexpression-Decreased Tube Formation in T24 Cells
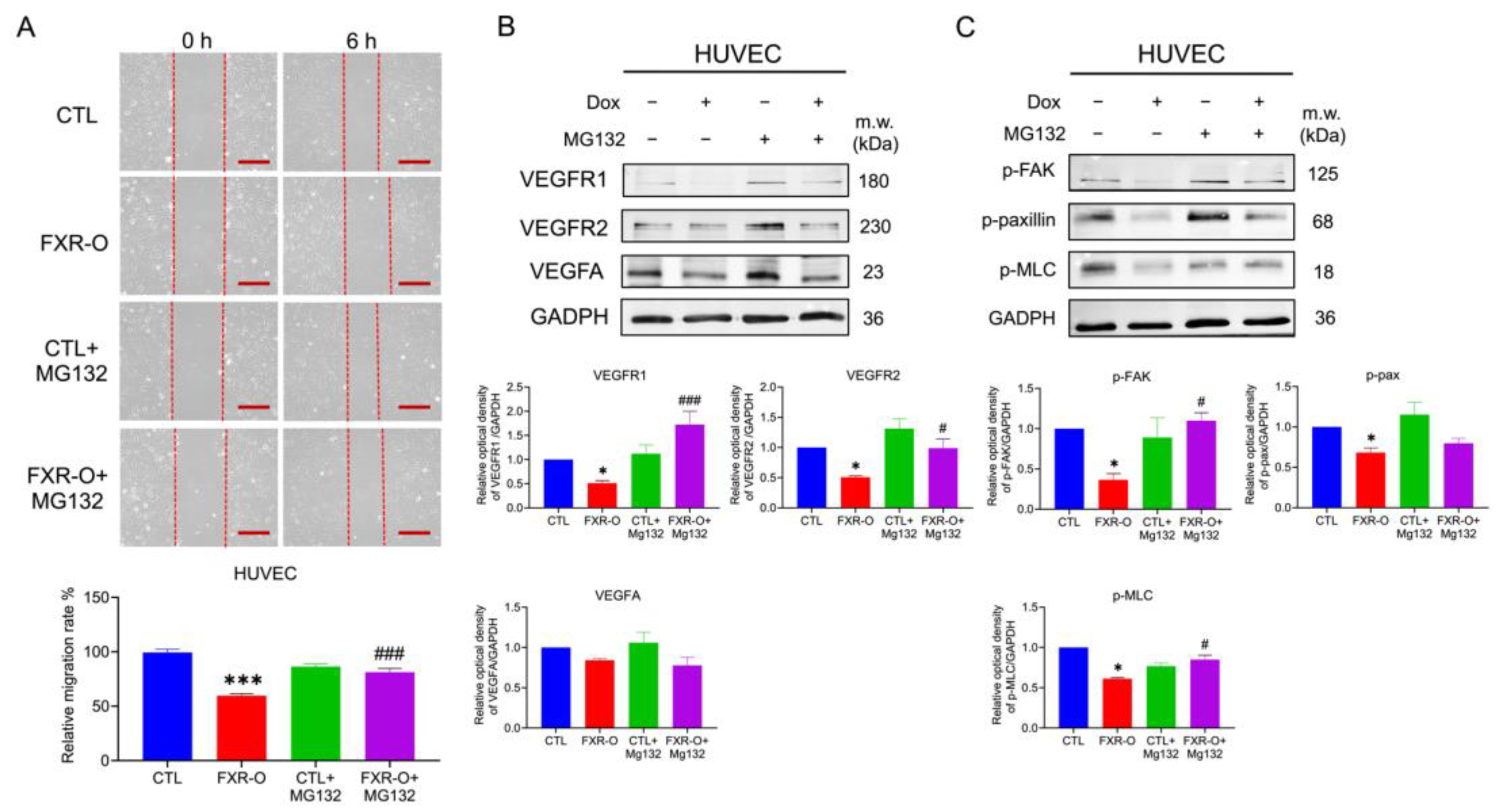
2.7. HUVECs Migratory Abilities Were Reduced by Proteasomal Degradation
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Plasmid Construction, Lentivirus Production and Doxycycline Induciable Overexpression
4.3. MTT Assays
4.4. Colony Formation Assays
4.5. Wound Healing Migration Assay
4.6. Adhesion Assays
4.7. Transwell Assays
4.8. Tube Formation Assays
4.9. Real-Time Polymerase Chain Reaction (RT-PCR)
4.10. Western Blotting
4.11. Enzyme-Linked Immunosorbent Assay (ELISA) for VEGF and MMP2
4.12. Human Active MMP9 Fluorokine Eassay
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- DeGeorge, K.C.; Holt, H.R.; Hodges, S.C. Bladder Cancer: Diagnosis and Treatment. Am. Fam. Physician 2017, 96, 507–514. [Google Scholar] [PubMed]
- Babjuk, M.; Burger, M.; Zigeuner, R.; Shariat, S.F.; van Rhijn, B.W.; Compérat, E.; Sylvester, R.J.; Kaasinen, E.; Böhle, A.; Redorta, J.P.; et al. EAU Guidelines on Non–Muscle-invasive Urothelial Carcinoma of the Bladder: Update 2013. Eur. Urol. 2013, 64, 639–653. [Google Scholar] [CrossRef] [PubMed]
- Crocetto, F.; di Zazzo, E.; Buonerba, C.; Aveta, A.; Pandolfo, S.D.; Barone, B.; Trama, F.; Caputo, V.F.; Scafuri, L.; Ferro, M.; et al. Kaempferol, Myricetin and Fisetin in Prostate and Bladder Cancer: A Systematic Review of the Literature. Nutrients 2021, 13, 3750. [Google Scholar] [CrossRef]
- Crocetto, F.; Barone, B.; Ferro, M.; Busetto, G.M.; La Civita, E.; Buonerba, C.; Di Lorenzo, G.; Terracciano, D.; Schalken, J.A. Liquid biopsy in bladder cancer: State of the art and future perspectives. Crit. Rev. Oncol. 2022, 170, 103577. [Google Scholar] [CrossRef]
- Chamie, K.; Litwin, M.S.; Bassett, J.C.; Daskivich, T.J.; Lai, J.; Hanley, J.M.; Konety, B.R.; Saigal, C.S.; the Urologic Diseases in America Project. Recurrence of high-risk bladder cancer: A population-based analysis. Cancer 2013, 119, 3219–3227. [Google Scholar] [CrossRef] [Green Version]
- Makishima, M.; Okamoto, A.Y.; Repa, J.J.; Tu, H.; Learned, R.M.; Luk, A.; Hull, M.V.; Lustig, K.D.; Mangelsdorf, D.J.; Shan, B. Identification of a Nuclear Receptor for Bile Acids. Science 1999, 284, 1362–1365. [Google Scholar] [CrossRef]
- Forman, B.M.; Goode, E.; Chen, J.; Oro, E.A.; Bradley, D.J.; Perlmann, T.; Noonan, D.J.; Burka, L.T.; McMorris, T.; Lamph, W.W.; et al. Identification of a nuclear receptor that is activated by farnesol metabolites. Cell 1995, 81, 687–693. [Google Scholar] [CrossRef] [Green Version]
- Lambert, G.; Amar, M.J.A.; Guo, G.; Brewer, H.B.; Gonzalez, F.J.; Sinal, C.J. The Farnesoid X-receptor Is an Essential Regulator of Cholesterol Homeostasis. J. Biol. Chem. 2003, 278, 2563–2570. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, M.; Houten, S.M.; Wang, L.; Moschetta, A.; Mangelsdorf, D.J.; Heyman, R.A.; Moore, D.D.; Auwerx, J. Bile acids lower triglyceride levels via a pathway involving FXR, SHP, and SREBP-1c. J. Clin. Investig. 2004, 113, 1408–1418. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Lee, F.Y.; Barrera, G.; Lee, H.; Vales, C.; Gonzalez, F.J.; Willson, T.M.; Edwards, P.A. Activation of the nuclear receptor FXR improves hyperglycemia and hyperlipidemia in diabetic mice. Proc. Natl. Acad. Sci. USA 2006, 103, 1006–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, S. Bile Acid Receptor Farnesoid X Receptor: A Novel Therapeutic Target for Metabolic Diseases. J. Lipid Atheroscler. 2017, 6, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Absil, L.; Journé, F.; Larsimont, D.; Body, J.J.; Tafforeau, L.; Nonclercq, D. Farnesoid X receptor as marker of osteotropism of breast cancers through its role in the osteomimetism of tumor cells. BMC Cancer 2020, 20, 640. [Google Scholar] [CrossRef] [PubMed]
- De Gottardi, A.; Touri, F.; Maurer, C.A.; Perez, A.; Maurhofer, O.; Ventre, G.; Bentzen, C.L.; Niesor, E.J.; Dufour, J.-F. The Bile Acid Nuclear Receptor FXR and the Bile Acid Binding Protein IBABP Are Differently Expressed in Colon Cancer. Am. J. Dig. Dis. 2004, 49, 982–989. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.-H.; Ro, E.J.; Yoon, J.-S.; Mizutani, T.; Kang, D.-W.; Park, J.-C.; Kim, T.I.; Clevers, H.; Choi, K.-Y. 5-FU promotes stemness of colorectal cancer via p53-mediated WNT/β-catenin pathway activation. Nat. Commun. 2020, 11, 5321. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Wang, B.; Chen, R.; Zhong, S.; Gao, F.; Zhang, Y.; Niu, Y.; Li, C.; Shi, G. The Nuclear Farnesoid X Receptor Reduces p53 Ubiquitination and Inhibits Cervical Cancer Cell Proliferation. Front. Cell Dev. Biol. 2021, 9, 583146. [Google Scholar] [CrossRef]
- Feng, Q.; Zhang, H.; Yao, D.; Zhang, X.; Chen, W.D.; Wang, Y.D. Activation of FXR suppresses esophageal squamous cell carcinoma through antagonizing ERK1/2 signaling pathway. Cancer Manag. Res. 2021, 13, 5907. [Google Scholar] [CrossRef]
- Su, H.; Ma, C.; Liu, J.; Li, N.; Gao, M.; Huang, A.; Wang, X.; Huang, W.; Huang, X. Downregulation of nuclear receptor FXR is associated with multiple malignant clinicopathological characteristics in human hepatocellular carcinoma. Am. J. Physiol. Liver Physiol. 2012, 303, G1245–G1253. [Google Scholar] [CrossRef] [Green Version]
- Ylänne, J.; Chen, Y.; O’Toole, E.T.; Loftus, J.C.; Takada, Y.; Ginsberg, M.H. Distinct functions of integrin alpha and beta subunit cytoplasmic domains in cell spreading and formation of focal adhesions. J. Cell Biol. 1993, 122, 223–233. [Google Scholar] [CrossRef]
- Janiszewska, M.; Primi, M.C.; Izard, T. Cell adhesion in cancer: Beyond the migration of single cells. J. Biol. Chem. 2020, 295, 2495–2505. [Google Scholar] [CrossRef] [Green Version]
- Guan, J.-L. Focal adhesion kinase in integrin signaling. Matrix Biol. 1997, 16, 195–200. [Google Scholar] [CrossRef]
- Crowe, D.L.; Ohannessian, A. Recruitment of focal adhesion kinase and paxillin to β1 integrin promotes cancer cell migration via mitogen activated protein kinase activation. BMC Cancer 2004, 4, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Li, N.; Zeng, Y.; Wang, X.; Li, J.; Su, H.; Gao, M.; Huang, X. Nuclear receptor FXR impairs SK-Hep-1 cell migration and invasion by inhibiting the Wnt/β-catenin signaling pathway. Oncol. Lett. 2020, 20, 1. [Google Scholar] [CrossRef]
- Yu, J.; Li, S.; Guo, J.; Xu, Z.; Zheng, J.; Sun, X. Farnesoid X receptor antagonizes Wnt/β-catenin signaling in colorectal tumorigenesis. Cell Death Dis. 2020, 11, 640. [Google Scholar] [CrossRef] [PubMed]
- Renner, G.; Noulet, F.; Mercier, M.-C.; Choulier, L.; Etienne-Selloum, N.; Gies, J.-P.; Lehmann, M.; Lelong-Rebel, I.; Martin, S.; Dontenwill, M. Expression/activation of α5β1 integrin is linked to the β-catenin signaling pathway to drive migration in glioma cells. Oncotarget 2016, 7, 62194–62207. [Google Scholar] [CrossRef] [Green Version]
- Lang, B.J.; Nguyen, L.; Nguyen, H.C.; Vieusseux, J.L.; Chai, R.C.C.; Christophi, C.; Fifis, T.; Kouspou, M.M.; Price, J.T. Heat stress induces epithelial plasticity and cell migration independent of heat shock factor 1. Cell Stress Chaperon 2012, 17, 765–778. [Google Scholar] [CrossRef] [Green Version]
- Kang, J.K.; Chang, C.-H.; Nam, H.J.; Kim, S.-K.; Ahn, K.J.; Seok, H.; Park, S.J.; Kang, Y.J.; Jo, Y.S.; Shong, M.; et al. Downregulation of erythropoietin receptor by overexpression of phospholipase C-gamma 1 is critical for decrease on focal adhesion in transformed cells. Cell. Oncol. 2011, 34, 11–21. [Google Scholar] [CrossRef]
- Clyman, R.I.; Mauray, F.; Kramer, R.H. β1 and β3 integrins have different roles in the adhesion and migration of vascular smooth muscle cells on extracellular matrix. Exp. Cell Res. 1992, 200, 272–284. [Google Scholar] [CrossRef]
- Peng, Z.; Chen, J.; Drachenberg, C.B.; Raufman, J.-P.; Xie, G. Farnesoid X receptor represses matrix metalloproteinase 7 expression, revealing this regulatory axis as a promising therapeutic target in colon cancer. J. Biol. Chem. 2019, 294, 8529–8542. [Google Scholar] [CrossRef]
- Chen, Y.; Mathy, N.W.; Lu, H. The role of VEGF in the diagnosis and treatment of malignant pleural effusion in patients with non-small cell lung cancer (Review). Mol. Med. Rep. 2018, 17, 8019–8030. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.T.; Ung, T.T.; Kim, N.H.; Jung, Y.D. Role of bile acids in colon carcinogenesis. World J. Clin. Cases 2018, 6, 577–588. [Google Scholar] [CrossRef] [PubMed]
- Carmeliet, P.; Jain, R.K. Angiogenesis in cancer and other diseases. Nature 2000, 407, 249–257. [Google Scholar] [CrossRef]
- Wang, Z.; Yan, M.; Li, J.; Long, J.; Li, Y.; Zhang, H. Dual functions of STAT3 in LPS-induced angiogenesis of hepatocellular carcinoma. Biochim. Biophys. Acta 2019, 1866, 566–574. [Google Scholar] [CrossRef]
- Meng, L.; Wang, C.; Wang, Z.; Yin, T.; Liu, Z.; Qin, H.; Zhang, Y.; Gu, X.; Yu, X.; Jiang, L.; et al. Feixian Recipe inhibits pulmonary fibrosis by targeting pulmonary microvascular endothelial cells and VEGF/VEGFR2 signaling pathway. Tradit. Med. Mod. Med. 2018, 1, 59–67. [Google Scholar] [CrossRef]
- Buess, M.; Rajski, M.; Vogel-Durrer, B.M.; Herrmann, R.; Rochlitz, C. Tumor-Endothelial interaction links the CD44+/CD24- phenotype with poor prognosis in early-stage breast cancer. Neoplasia 2009, 11, 987–1002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, T.; Yu, H.; Xia, Q.; Ma, Y.; Yin, H.; Shen, Y.; Liu, X. Cross-talk mechanism between endothelial cells and hepatocellular carcinoma cells via growth factors and integrin pathway promotes tumor angiogenesis and cell migration. Oncotarget 2017, 8, 69577–69593. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.-H.; Lee, B.-H.; Choi, S.-H.; Kim, H.-J.; Won, K.J.; Lee, H.M.; Rhim, H.; Kim, H.-C.; Nah, S.-Y. Effects of gintonin on the proliferation, migration, and tube formation of human umbilical-vein endothelial cells: Involvement of lysophosphatidic-acid receptors and vascular-endothelial-growth-factor signaling. J. Ginseng Res. 2016, 40, 325–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bijnsdorp, I.V.; Capriotti, F.; Kruyt, A.E.F.; Losekoot, N.; Fukushima, M.; Griffioen, A.W.; Thijssen, V.L.; Peters, G.J. Thymidine phosphorylase in cancer cells stimulates human endothelial cell migration and invasion by the secretion of angiogenic factors. Br. J. Cancer 2011, 104, 1185–1192. [Google Scholar] [CrossRef]
- Jang, J.P.; Han, J.M.; Jung, H.J.; Osada, H.; Jang, J.H.; Ahn, J.S. Anti-Angiogenesis Effects Induced by Octaminomycins A and B against HUVECs. J. Microbiol. Biotechnol. 2018, 28, 1332–1338. [Google Scholar] [CrossRef]
- Gege, C.; Hambruch, E.; Hambruch, N.; Kinzel, O.; Kremoser, C. Nonsteroidal FXR Ligands: Current Status and Clinical Applications. Handb. Exp. Pharm. 2019, 256, 167–205. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lai, C.-R.; Wang, H.-H.; Chang, H.-H.; Tsai, Y.-L.; Tsai, W.-C.; Lee, C.-R.; Changchien, C.-Y.; Cheng, Y.-C.; Wu, S.-T.; Chen, Y. Enhancement of Farnesoid X Receptor Inhibits Migration, Adhesion and Angiogenesis through Proteasome Degradation and VEGF Reduction in Bladder Cancers. Int. J. Mol. Sci. 2022, 23, 5259. https://doi.org/10.3390/ijms23095259
Lai C-R, Wang H-H, Chang H-H, Tsai Y-L, Tsai W-C, Lee C-R, Changchien C-Y, Cheng Y-C, Wu S-T, Chen Y. Enhancement of Farnesoid X Receptor Inhibits Migration, Adhesion and Angiogenesis through Proteasome Degradation and VEGF Reduction in Bladder Cancers. International Journal of Molecular Sciences. 2022; 23(9):5259. https://doi.org/10.3390/ijms23095259
Chicago/Turabian StyleLai, Chien-Rui, Hisao-Hsien Wang, Hsin-Han Chang, Yu-Ling Tsai, Wen-Chiuan Tsai, Chen-Ray Lee, Chih-Ying Changchien, Yu-Chen Cheng, Sheng-Tang Wu, and Ying Chen. 2022. "Enhancement of Farnesoid X Receptor Inhibits Migration, Adhesion and Angiogenesis through Proteasome Degradation and VEGF Reduction in Bladder Cancers" International Journal of Molecular Sciences 23, no. 9: 5259. https://doi.org/10.3390/ijms23095259
APA StyleLai, C. -R., Wang, H. -H., Chang, H. -H., Tsai, Y. -L., Tsai, W. -C., Lee, C. -R., Changchien, C. -Y., Cheng, Y. -C., Wu, S. -T., & Chen, Y. (2022). Enhancement of Farnesoid X Receptor Inhibits Migration, Adhesion and Angiogenesis through Proteasome Degradation and VEGF Reduction in Bladder Cancers. International Journal of Molecular Sciences, 23(9), 5259. https://doi.org/10.3390/ijms23095259