
Plant Alkaloid Tetrandrine Is a Nuclear Receptor 4A1 Antagonist and Inhibits Panc-1 Cell Growth In Vitro and In Vivo


 , , and
, , and 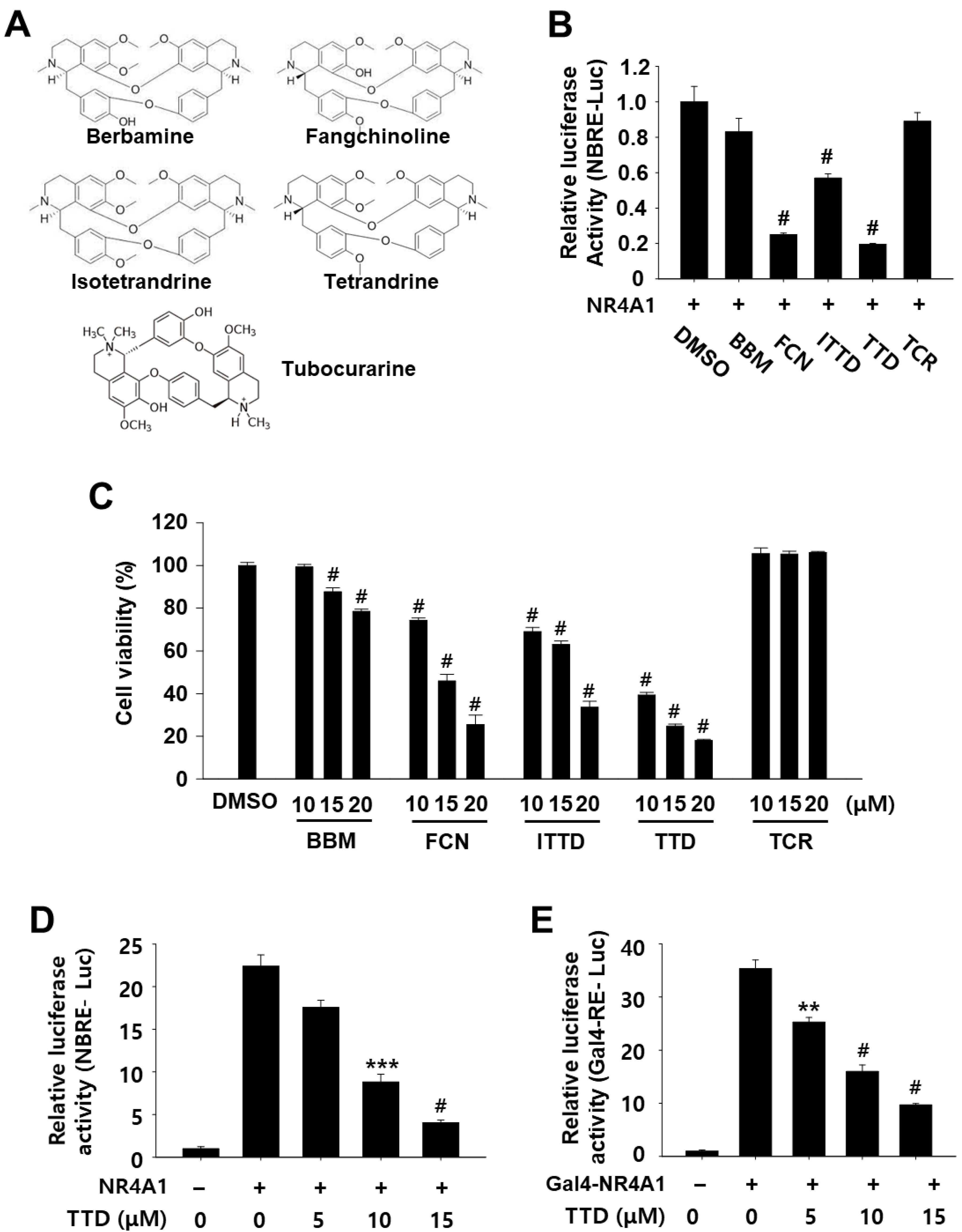{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
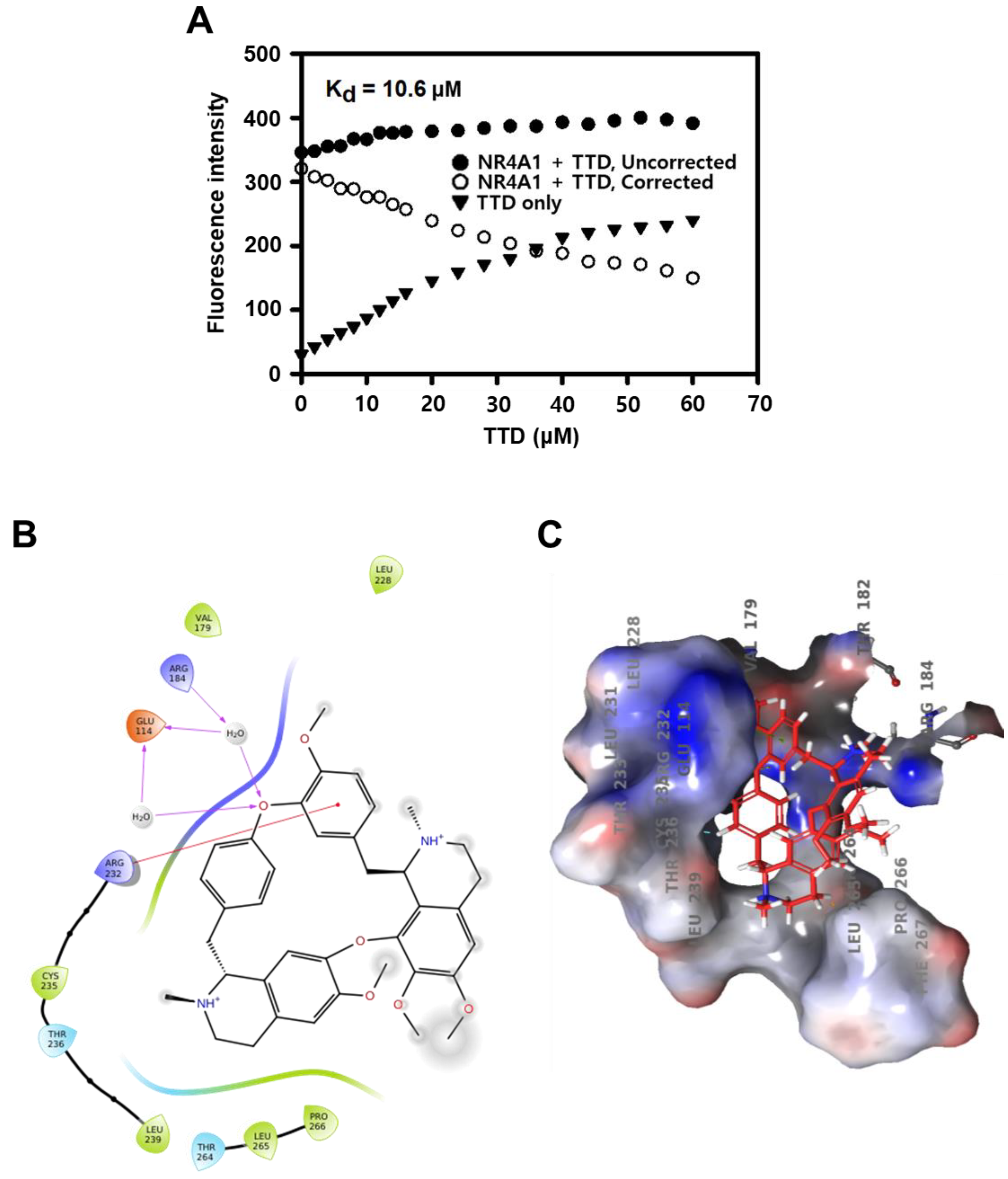
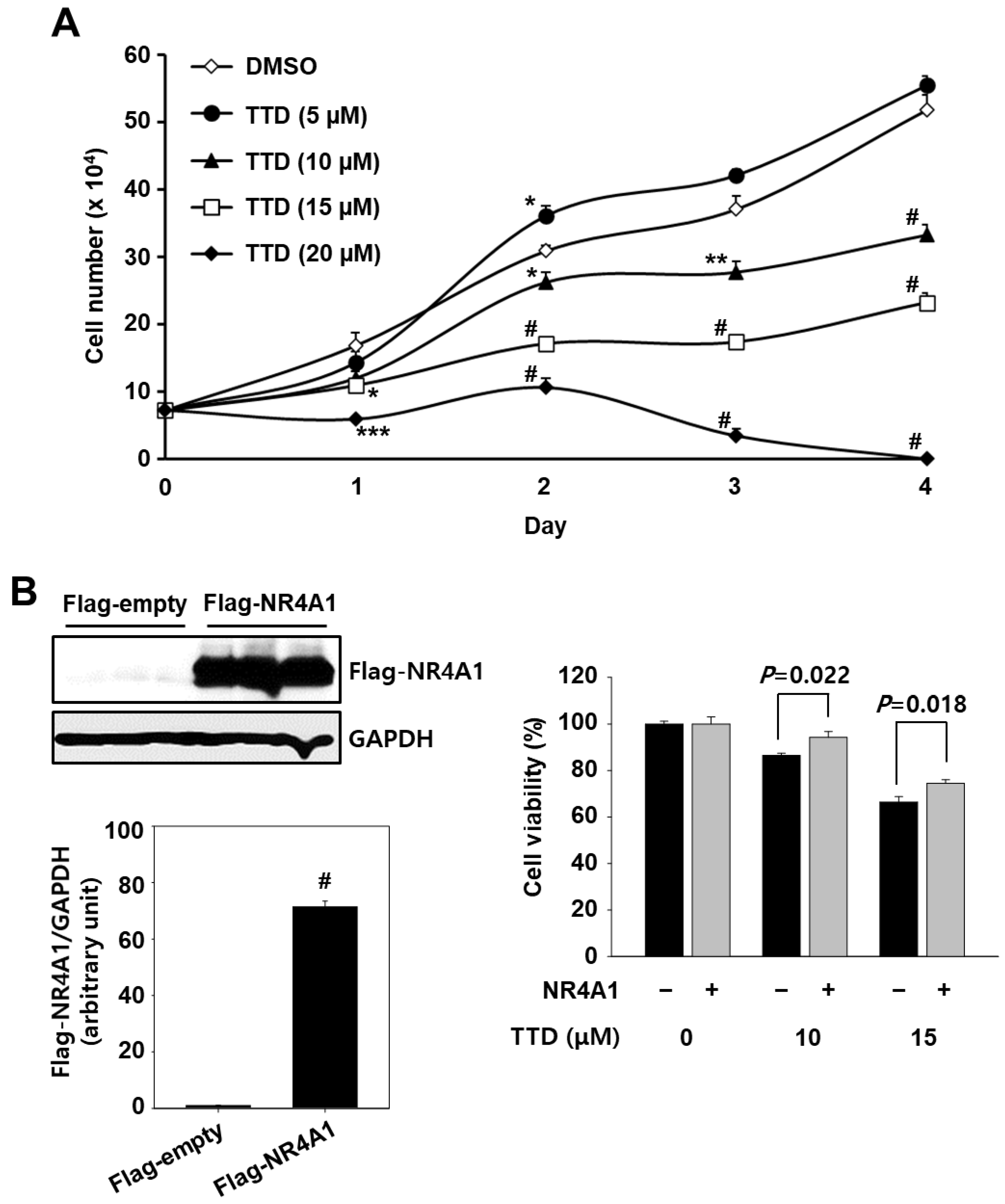
2.1. TTD Is a NR4A1 Antagonist and Decreases Panc-1 Cell Growth
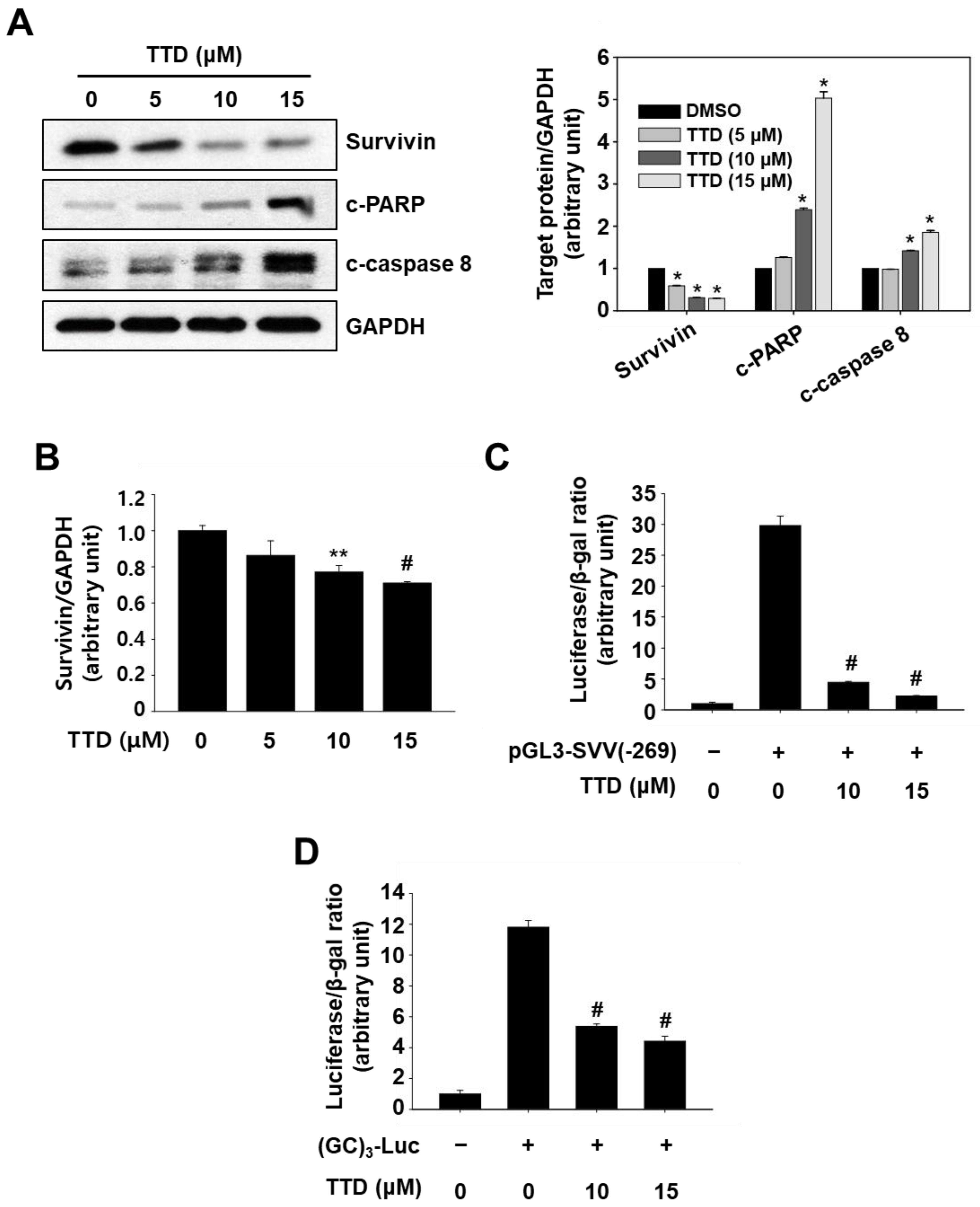
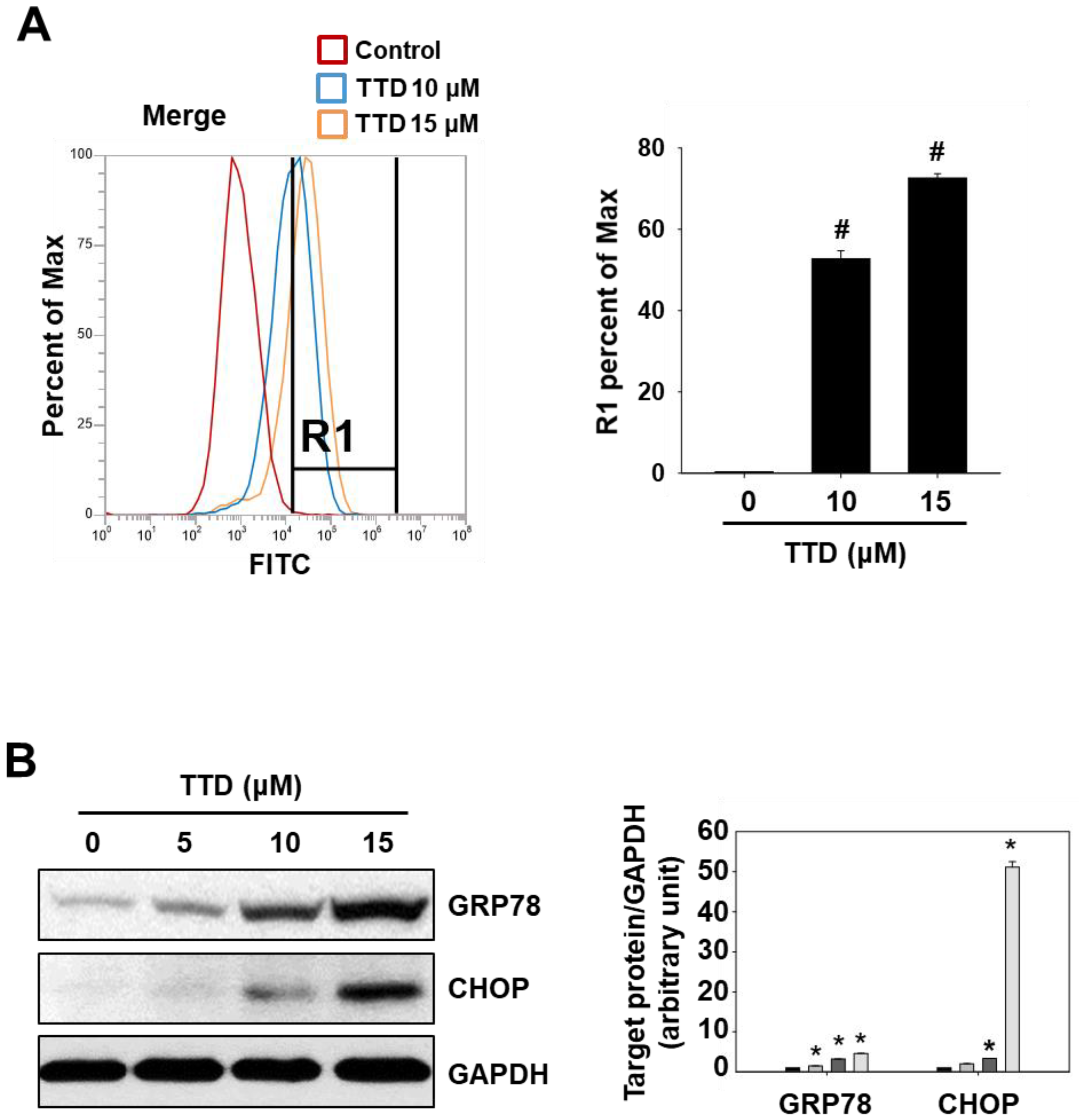
2.2. TTD Induces Apoptosis through Inhibition of NR4A1-Mediated Pro-Oncogenic Pathways
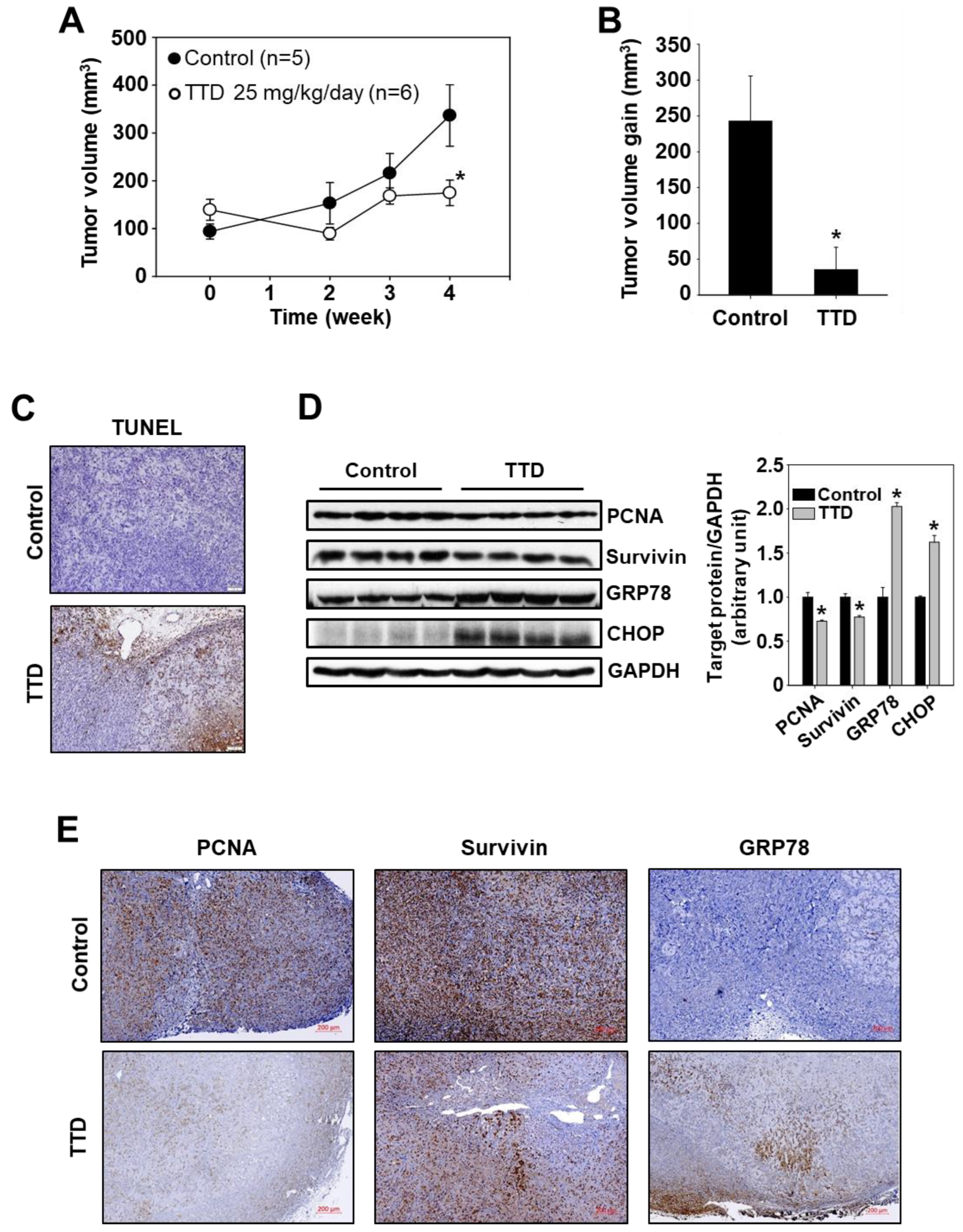
2.3. TTD Reduces Tumor Growth in a Xenograft Mouse Model of Panc-1 Cells
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Plasmids
4.2. Antibodies, Reagents, Quantitative Real-Time PCR, and Western Blot Analysis
4.3. Cell Proliferation Assay, Transfection, and Reporter Gene Assay
4.4. Direct Binding Assay
4.5. Computation-Based Molecular Modeling
4.6. Measurement of Intracellular Level of ROS
4.7. Animals and Experimental Design
4.8. Immunohistochemical Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Crean, D.; Murphy, E.P. Targeting NR4A nuclear receptors to control stromal cell inflammation, metabolism, angiogenesis, and tumorigenesis. Front. Cell Dev. Biol. 2021, 9, 589770. [Google Scholar] [CrossRef] [PubMed]
- Medzikovic, L.; de Vries, C.J.M.; de Waard, V. NR4A nuclear receptors in cardiac remodeling and neurohormonal regulation. Trends Cardiovasc. Med. 2019, 29, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Mohan, H.M.; Aherne, C.M.; Rogers, A.; Baird, A.; Winter, D.C.; Murphy, E.P. Molecular pathways: The role of NR4A orphan nuclear receptors in cancer. Clin. Cancer Res. 2012, 18, 3223–3228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safe, S.; Karki, K. The paradoxical roles of orphan nuclear receptor 4A (NR4A) in cancer. Mol. Cancer Res. 2021, 19, 180–191. [Google Scholar] [CrossRef]
- Wu, L.; Chen, L. Characteristics of Nur77 and its ligands as potential anticancer compounds (Review). Mol. Med. Rep. 2018, 18, 4793–4801. [Google Scholar] [CrossRef]
- Lee, S.O.; Abdelrahim, M.; Yoon, K.; Chintharlapalli, S.; Papineni, S.; Kim, K.; Wang, H.; Safe, S. Inactivation of the orphan nuclear receptor TR3/Nur77 inhibits pancreatic cancer cell and tumor growth. Cancer Res. 2010, 70, 6824–6836. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-O.; Andey, T.; Jin, U.-H.; Kim, K.; Sachdeva, M.; Safe, S. The nuclear receptor TR3 regulates mTORC1 signaling in lung cancer cells expressing wild-type p53. Oncogene 2012, 31, 3265–3276. [Google Scholar] [CrossRef] [Green Version]
- Safe, S.; Jin, U.-H.; Hedrick, E.; Reeder, A.; Lee, S.-O. Minireview: Role of orphan nuclear receptors in cancer and potential as drug targets. Mol. Endocrinol. 2014, 28, 157–172. [Google Scholar] [CrossRef]
- Lee, S.-O.; Jin, U.-H.; Kang, J.H.; Kim, S.B.; Guthrie, A.S.; Sreevalsan, S.; Lee, J.-S.; Safe, S. The orphan nuclear receptor NR4A1 (Nur77) regulates oxidative and endoplasmic reticulum stress in pancreatic cancer cells. Mol. Cancer Res. 2014, 12, 527–538. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.-S.; Safe, S.; Lee, S.-O. Inactivation of the orphan nuclear receptor NR4A1 contributes to apoptosis induction by fangchinoline in pancreatic cancer cells. Toxicol. Appl. Pharmacol. 2017, 332, 32–39. [Google Scholar] [CrossRef]
- Lee, H.-S.; Kim, S.-H.; Kim, B.-M.; Safe, S.; Lee, S.-O. Broussochalcone A is a novel inhibitor of the orphan nuclear receptor NR4A1 and induces apoptosis in pancreatic cancer cells. Molecules 2021, 26, 2316. [Google Scholar] [CrossRef] [PubMed]
- Felix, J.P.; King, V.F.; Shevell, J.L.; Garcia, M.L.; Kaczorowski, G.J.; Bick, I.R.C.; Slaughter, R.S. Bis(benzylisoquinoline) analogs of tetrandrine block L-type calcium channels: Evidence for interaction at the diltiazem-binding site. Biochemistry 1992, 31, 11793–11800. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Peng, J.-M.; Su, L.-D.; Wang, N.-Y.; Yu, Y.-J. Fangchinoline inhibits the proliferation of SPC-A-1 lung cancer cells by blocking cell cycle progression. Exp. Ther. Med. 2016, 11, 613–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Lu, Y.; Sun, P.; Feng, L.X.; Liu, M.; Hu, L.H.; Wu, W.Y.; Jiang, B.H.; Yang, M.; Qu, X.B.; et al. Inhibition on proteasome beta1 subunit might contribute to the anti-cancer effects of fangchinoline in human prostate cancer cells. PLoS ONE 2015, 10, e0141681. [Google Scholar]
- Liu, L.; Xu, Z.; Yu, B.; Tao, L.; Cao, Y. Berbamine inhibits cell proliferation and migration and induces cell death of lung cancer cells via regulating c-Maf, PI3K/Akt, and MDM2-p53 pathways. Evid.-Based Complementary Altern. Med. 2021, 2021, 5517143. [Google Scholar] [CrossRef] [PubMed]
- Luan, F.; He, X.; Zeng, N. Tetrandrine: A review of its anticancer potentials, clinical settings, pharmacokinetics and drug delivery systems. J. Pharm. Pharmacol. 2020, 72, 1491–1512. [Google Scholar] [CrossRef]
- Karki, K.; Mohankumar, K.; Schoeller, A.; Martin, G.; Shrestha, R.; Safe, S. NR4A1 ligands as potent inhibitors of breast cancer cell and tumor growth. Cancers 2021, 13, 2682. [Google Scholar] [CrossRef]
- Han, Y.-H.; Cao, X.; Lin, B.; Lin, F.; Kolluri, S.K.; Stebbins, J.; Reed, J.C.; I Dawson, M.; Zhang, X.-K. Regulation of Nur77 nuclear export by c-Jun N-terminal kinase and Akt. Oncogene 2006, 25, 2974–2986. [Google Scholar] [CrossRef] [Green Version]
- Zhan, Y.; Du, X.; Chen, H.; Liu, J.; Zhao, B.; Huang, D.; Li, G.; Xu, Q.; Zhang, M.; Weimer, B.C.; et al. Cytosporone B is an agonist for nuclear orphan receptor Nur77. Nat. Chem. Biol. 2008, 4, 548–556. [Google Scholar] [CrossRef]
- Bellik, Y.; Boukraa, L.; Alzahrani, H.A.; Bakhotmak, B.A.; Abdellah, F.; Hammoudi, S.M.; Iguer-Ouada, M. Molecular mechanism underlying anti-inflammatory and anti-allergic activities of phytochemicals: An update. Molecules 2012, 18, 322–353. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Jin, Q.; Wu, Y.L.; Sun, P.; Jiang, S.; Zhang, Y.; Zhang, D.Q.; Zhang, Y.J.; Lian, L.H.; Nan, J.X. Tetrandrine regulates hepatic stellate cell activation via TAK1 and NF-κB signaling. Int. Immunopharmacol. 2016, 36, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Capuzzi, S.J.; Sun, W.; Muratov, E.N.; Martinez-Romero, C.; He, S.; Zhu, W.; Li, H.; Tawa, G.; Fisher, E.G.; Xu, M.; et al. Computer-aided discovery and characterization of novel Ebola virus inhibitors. J. Med. Chem. 2018, 61, 3582–3594. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.L.; Wu, Z.Z.; Chen, L.X.; Wu, B.X.; Chen, L.L.; Qin, G.C.; Gui, B.; Zhou, J.Y. Neuroprotective effects of tetrandrine against vascular dementia. Neural. Regen. Res. 2016, 11, 454–459. [Google Scholar] [PubMed]
- Bhagya, N.; Chandrashekar, K.R.; Prabhu, A.; Rekha, P.D. Tetrandrine isolated from Cyclea peltata induces cytotoxicity and apoptosis through ROS and caspase pathways in breast and pancreatic cancer cells. In Vitro Cell Dev. Biol. Anim. 2019, 55, 331–340. [Google Scholar]
- Zhang, H.; Xie, B.; Zhang, Z.; Sheng, X.; Zhang, S. Tetrandrine suppresses cervical cancer growth by inducing apoptosis in vitro and in vivo. Drug Des. Dev. Ther. 2018, 13, 119–127. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Zhao, L.; Zhao, F.; Yang, G.; Wang, J.J. Tetrandrine suppresses lung cancer growth and induces apoptosis, potentially via the VEGF/HIF-1α/ICAM-1 signaling pathway. Oncol. Lett. 2018, 15, 7433–7437. [Google Scholar] [CrossRef] [Green Version]
- Lien, J.-C.; Lin, M.-W.; Chang, S.-J.; Lai, K.-C.; Huang, A.-C.; Yu, F.-S.; Chung, J.-G. Tetrandrine induces programmed cell death in human oral cancer CAL 27 cells through the reactive oxygen species production and caspase-dependent pathways and associated with beclin-1-induced cell autophagy. Environ. Toxicol. 2017, 32, 329–343. [Google Scholar] [CrossRef]
- Singh, K.; Dong, Q.; TimiriShanmugam, P.S.; Koul, S.; Koul, H.K. Tetrandrine inhibits deregulated cell cycle in pancreatic cancer cells: Differential regulation of p21Cip1/Waf1, p27Kip1 and cyclin D1. Cancer Lett. 2018, 425, 164–173. [Google Scholar] [CrossRef]
- Song, J.; Xu, J.; Guo, J.; Shang, Y.; Wang, J.; Wang, T. The enhancement of Tetrandrine to gemcitabine-resistant PANC-1 cytochemical sensitivity involves the promotion of PI3K/Akt/mTOR-mediated apoptosis and AMPK-regulated autophagy. Acta Histochem. 2021, 123, 151769. [Google Scholar] [CrossRef]
- Bishayee, A.; Sethi, G. Bioactive natural products in cancer prevention and therapy: Progress and promise. Semin. Cancer Biol. 2016, 40–41, 1–3. [Google Scholar] [CrossRef]
- Hwang, E.-Y.; Yu, M.-H.; Jung, Y.-S.; Lee, S.-P.; Shon, J.H.; Lee, S.-O. Defatted safflower seed extract inhibits adipogenesis in 3T3-L1 preadipocytes and improves lipid profiles in C57BL/6J ob/ob mice fed a high-fat diet. Nutr. Res. 2016, 36, 995–1003. [Google Scholar] [CrossRef] [PubMed]
- Hedrick, E.; Lee, S.-O.; Kim, G.; Abdelrahim, M.; Jin, U.-H.; Safe, S.; Abudayyeh, A. Nuclear receptor 4A1 (NR4A1) as a drug target for renal cell adenocarcinoma. PLoS ONE 2015, 10, e0128308. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.-S.; Kim, D.H.; Lee, I.-S.; Park, J.-H.; Martin, G.; Safe, S.; Kim, K.-J.; Kim, J.-H.; Jang, B.I.; Lee, S.-O. Plant Alkaloid Tetrandrine Is a Nuclear Receptor 4A1 Antagonist and Inhibits Panc-1 Cell Growth In Vitro and In Vivo. Int. J. Mol. Sci. 2022, 23, 5280. https://doi.org/10.3390/ijms23095280
Lee H-S, Kim DH, Lee I-S, Park J-H, Martin G, Safe S, Kim K-J, Kim J-H, Jang BI, Lee S-O. Plant Alkaloid Tetrandrine Is a Nuclear Receptor 4A1 Antagonist and Inhibits Panc-1 Cell Growth In Vitro and In Vivo. International Journal of Molecular Sciences. 2022; 23(9):5280. https://doi.org/10.3390/ijms23095280
Chicago/Turabian StyleLee, Hyo-Seon, Dae Hwan Kim, In-Seon Lee, Ji-Hyun Park, Gregory Martin, Stephen Safe, Keuk-Jun Kim, Joung-Hee Kim, Byung Ik Jang, and Syng-Ook Lee. 2022. "Plant Alkaloid Tetrandrine Is a Nuclear Receptor 4A1 Antagonist and Inhibits Panc-1 Cell Growth In Vitro and In Vivo" International Journal of Molecular Sciences 23, no. 9: 5280. https://doi.org/10.3390/ijms23095280
APA StyleLee, H. -S., Kim, D. H., Lee, I. -S., Park, J. -H., Martin, G., Safe, S., Kim, K. -J., Kim, J. -H., Jang, B. I., & Lee, S. -O. (2022). Plant Alkaloid Tetrandrine Is a Nuclear Receptor 4A1 Antagonist and Inhibits Panc-1 Cell Growth In Vitro and In Vivo. International Journal of Molecular Sciences, 23(9), 5280. https://doi.org/10.3390/ijms23095280