
Intrauterine Inflammation Leads to Select Sex- and Age-Specific Behavior and Molecular Differences in Mice

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
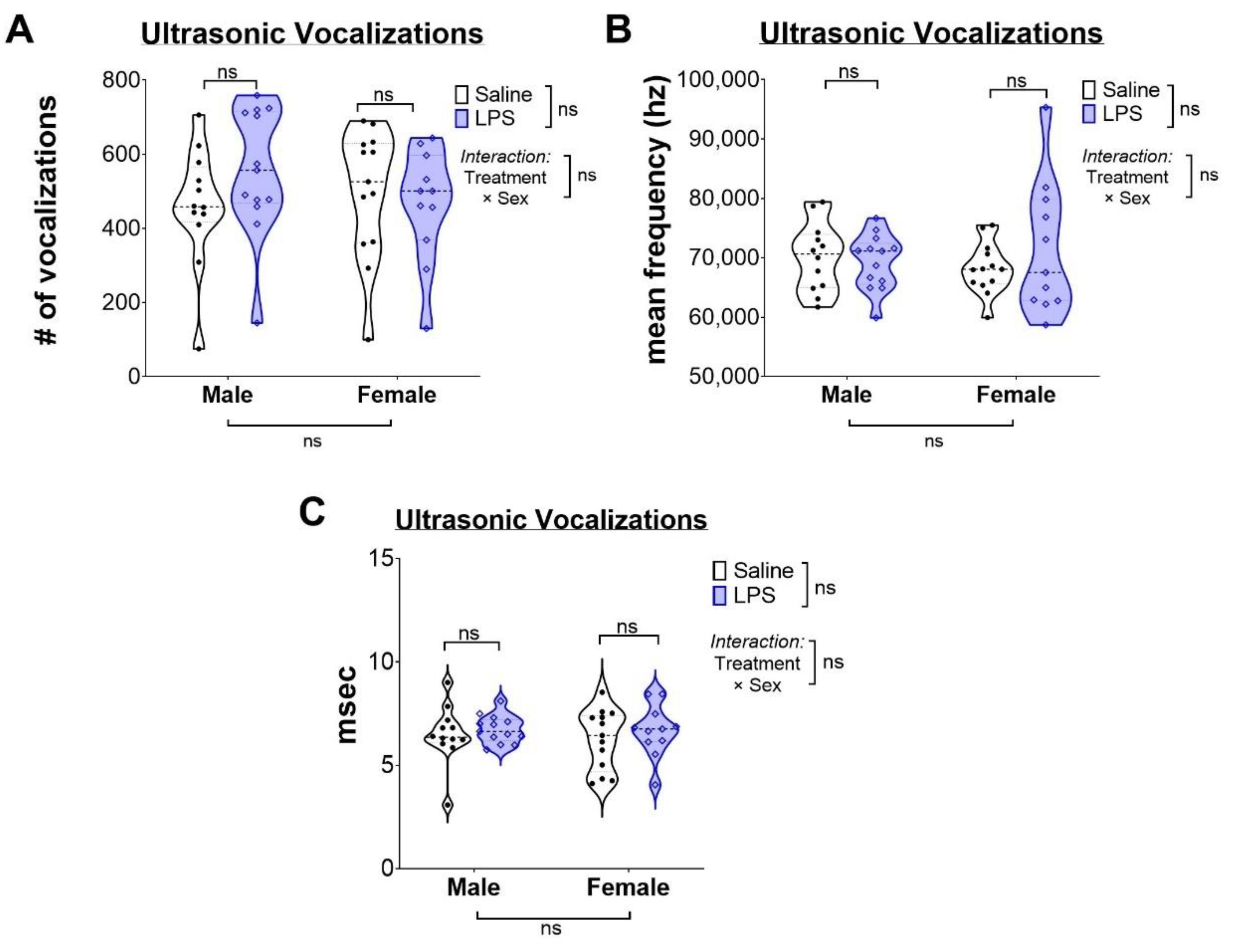
2.1. Intrauterine Inflammation Does Not Alter Ultrasonic Vocalizations in Neonatal Pups
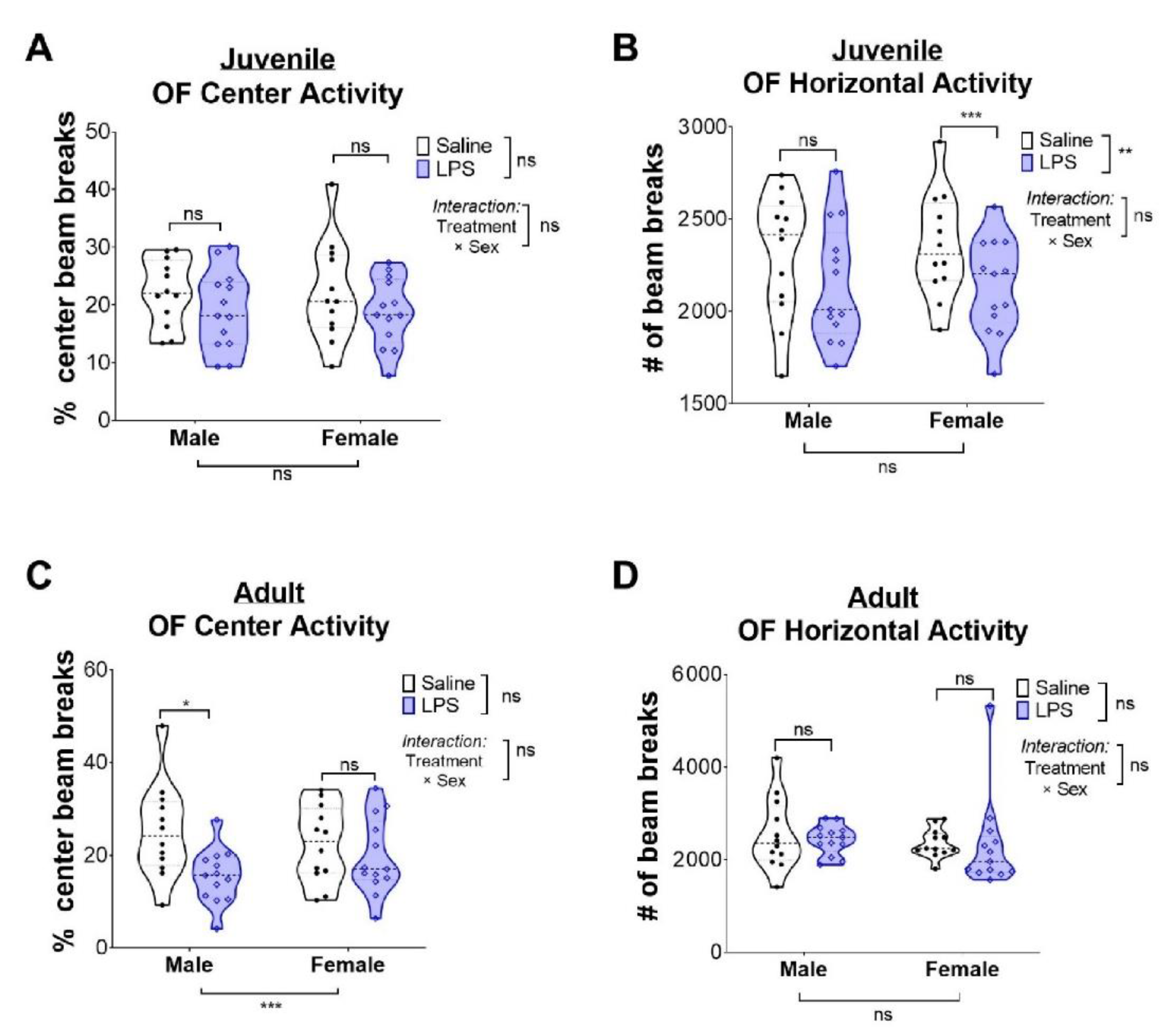
2.2. Intrauterine Inflammation Leads to Increased Anxiety-Related Behavior in Adult, but Not Juvenile, Males
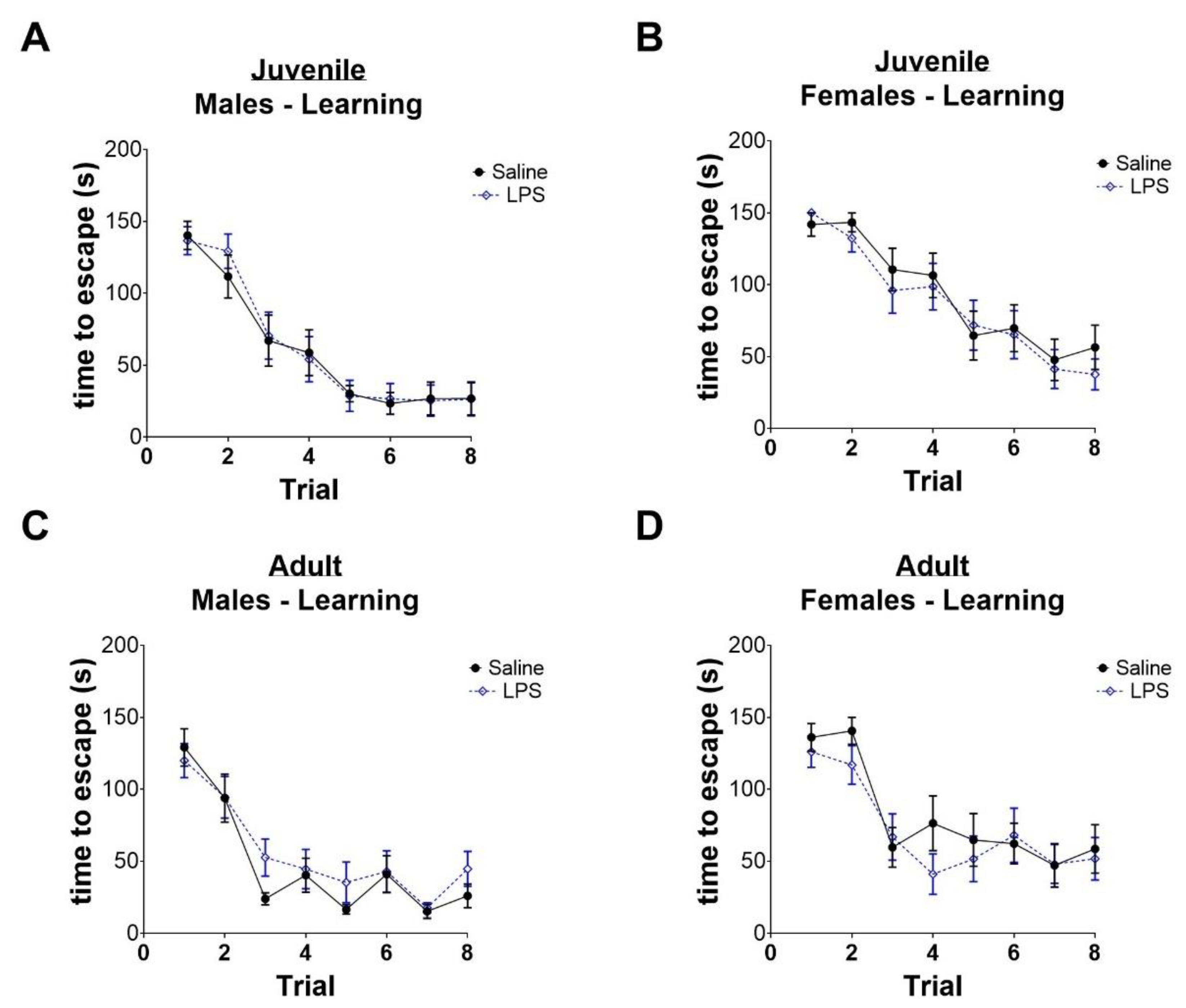
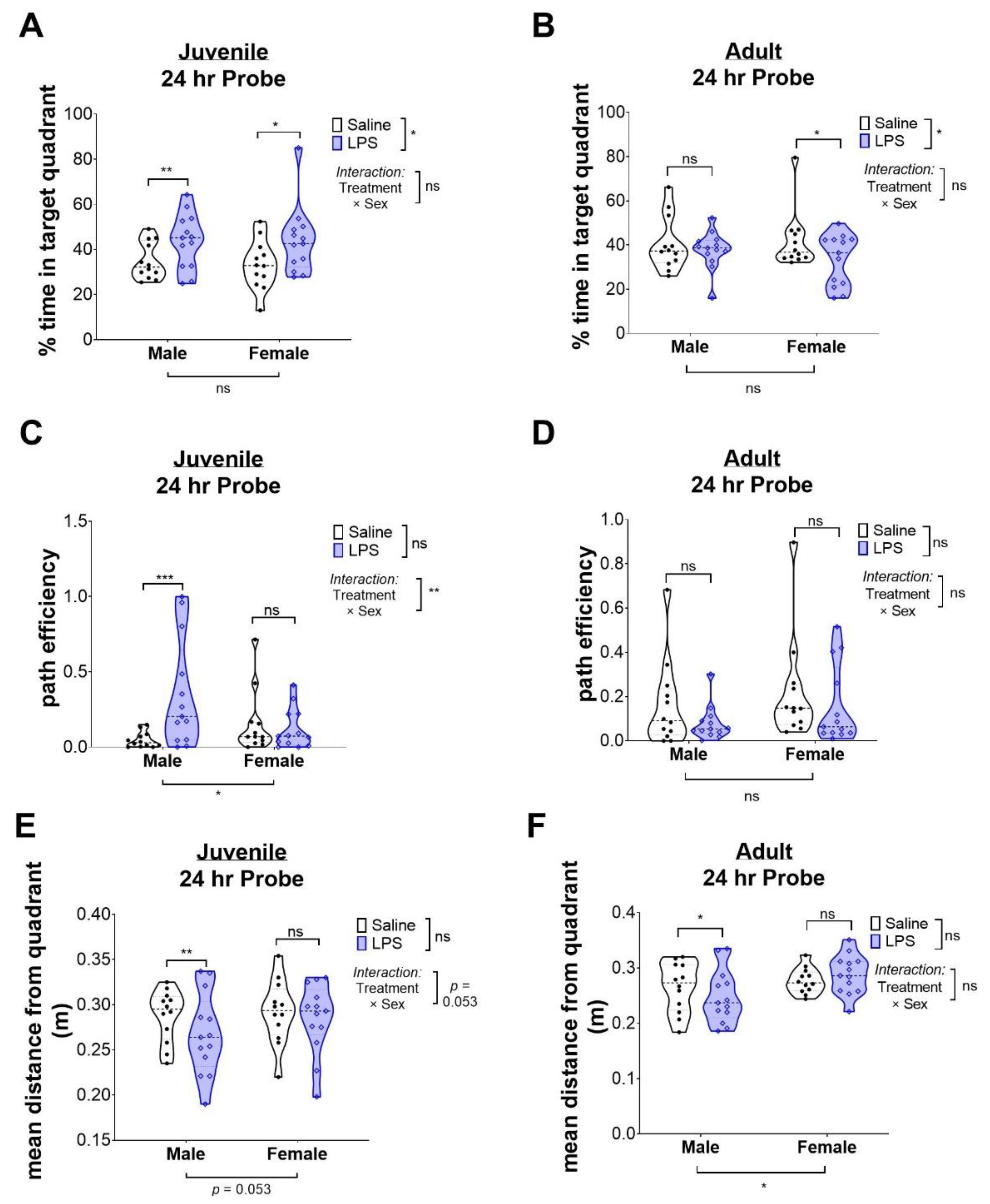
2.3. Intrauterine Exposure to Inflammation Is Not Associated with Improved Memory in Juvenile Mice, but Impaired Memory in Adult Female Mice
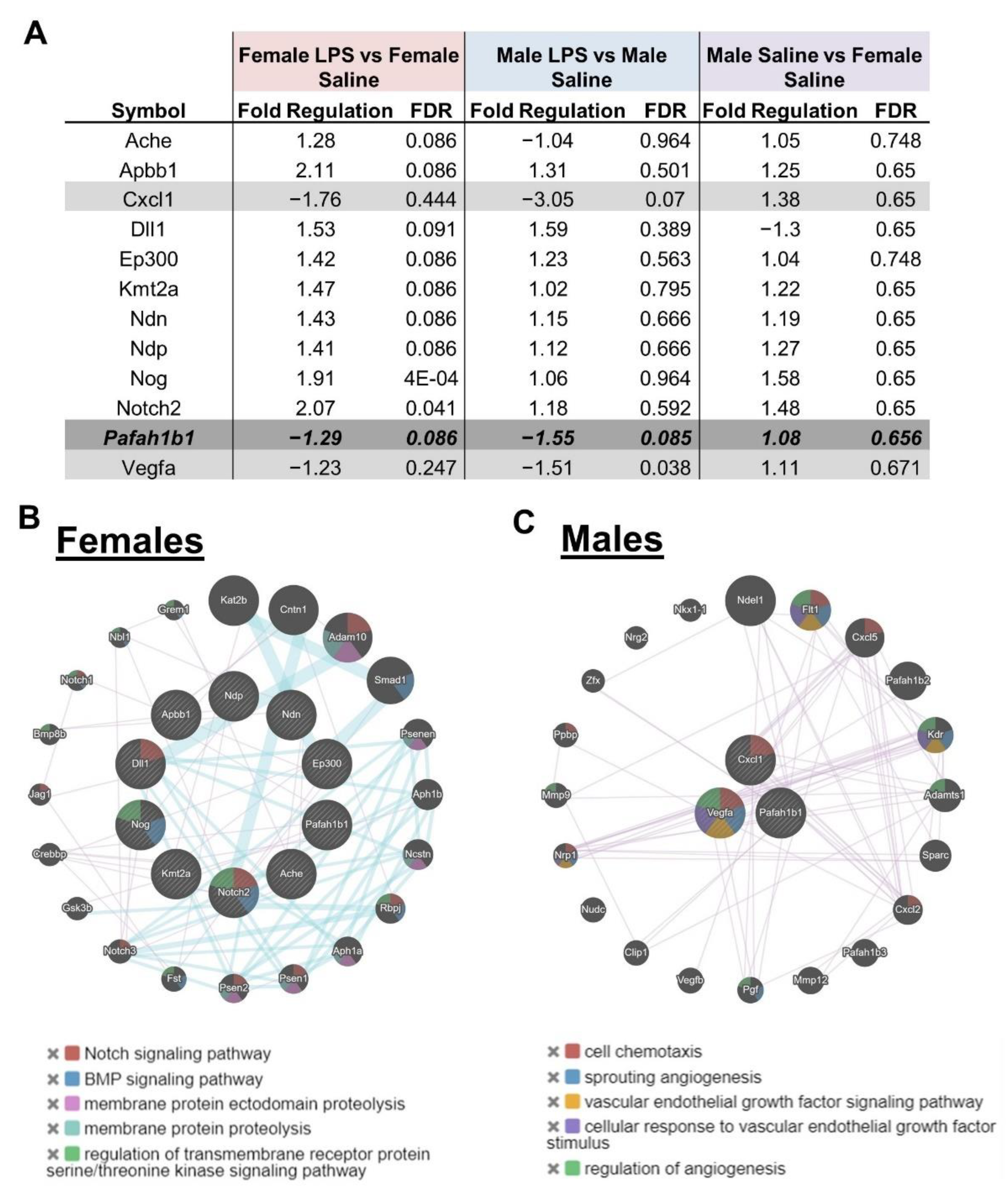
2.4. Intrauterine Exposure to Inflammation Leads to Sex-Dichotomous Differences in Gene Expression Related to Neurogenesis
3. Discussion
4. Materials and Methods
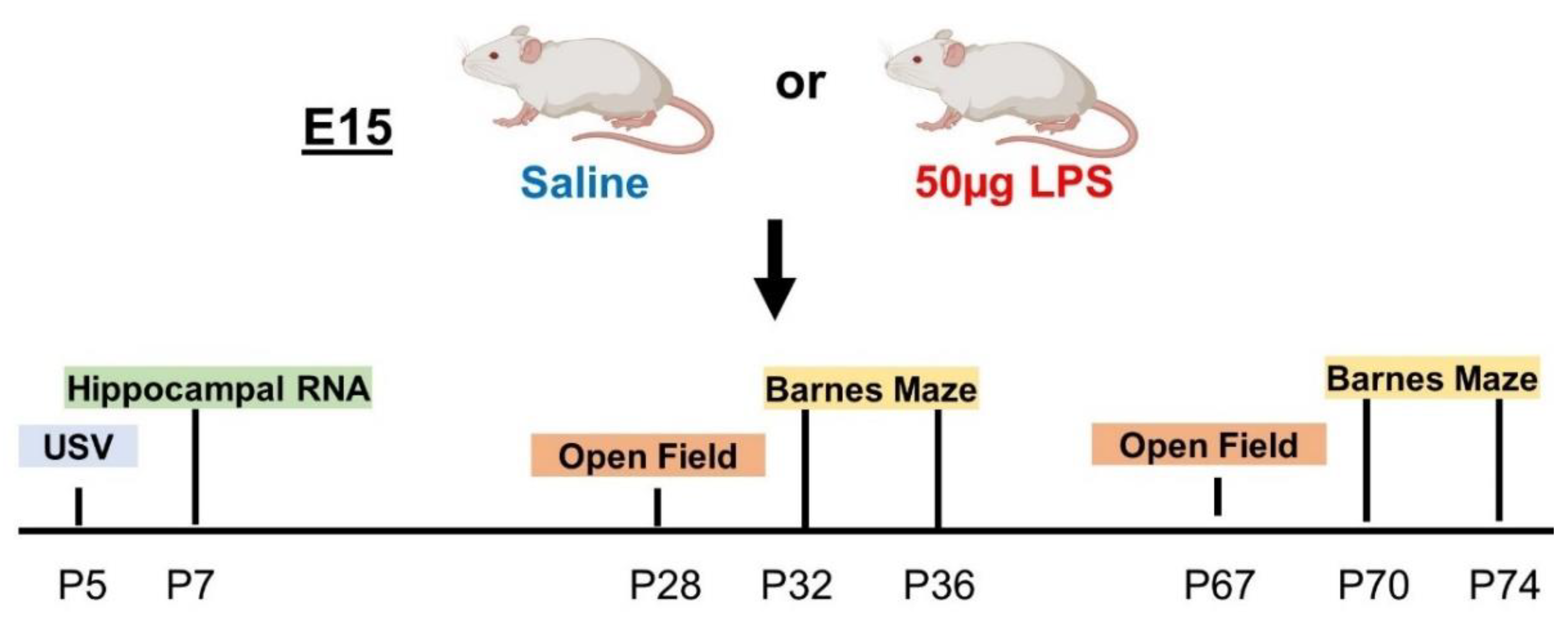
4.1. Mouse Model of Intrauterine Inflammation
4.2. Behavioral Testing
4.3. Hippocampal Dissections
4.4. Gene Expression Analysis Using Mouse Neurogenesis Arrays
4.5. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leung, M.P.; Thompson, B.; Black, J.; Dai, S.; Alsweiler, J.M. The effects of preterm birth on visual development. Clin. Exp. Optom. 2018, 101, 4–12. [Google Scholar] [CrossRef] [Green Version]
- Vieira, M.E.; Linhares, M.B. Developmental outcomes and quality of life in children born preterm at preschool- and school-age. J. Pediatr. 2011, 87, 281–291. [Google Scholar] [CrossRef] [Green Version]
- Allen, M.C.; Cristofalo, E.A.; Kim, C. Outcomes of preterm infants: Morbidity replaces mortality. Clin. Perinatol. 2011, 38, 441–454. [Google Scholar] [CrossRef]
- Fawke, J. Neurological outcomes following preterm birth. Semin. Fetal Neonatal. Med. 2007, 12, 374–382. [Google Scholar] [CrossRef]
- Dammann, O.; Leviton, A. Maternal intrauterine infection, cytokines, and brain damage in the preterm newborn. Pediatr. Res. 1997, 42, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Strunk, T.; Inder, T.; Wang, X.; Burgner, D.; Mallard, C.; Levy, O. Infection-induced inflammation and cerebral injury in preterm infants. Lancet Infect Dis. 2014, 14, 751–762. [Google Scholar] [CrossRef] [Green Version]
- Knuesel, I.; Chicha, L.; Britschgi, M.; Schobel, S.A.; Bodmer, M.; Hellings, J.A.; Toovey, S.; Prinssen, E.P. Maternal immune activation and abnormal brain development across CNS disorders. Nat. Rev. Neurol. 2014, 10, 643–660. [Google Scholar] [CrossRef]
- Wang, X.; Hellgren, G.; Lofqvist, C.; Li, W.; Hellstrom, A.; Hagberg, H.; Mallard, C. White matter damage after chronic subclinical inflammation in newborn mice. J. Child Neurol. 2009, 24, 1171–1178. [Google Scholar] [CrossRef]
- Burd, I.; Balakrishnan, B.; Kannan, S. Models of fetal brain injury, intrauterine inflammation, and preterm birth. Am. J. Reprod. Immunol. 2012, 67, 287–294. [Google Scholar] [CrossRef]
- Wang, X.; Hagberg, H.; Zhu, C.; Jacobsson, B.; Mallard, C. Effects of intrauterine inflammation on the developing mouse brain. Brain Res. 2007, 1144, 180–185. [Google Scholar] [CrossRef]
- Lehnardt, S.; Lachance, C.; Patrizi, S.; Lefebvre, S.; Follett, P.L.; Jensen, F.E.; Rosenberg, P.A.; Volpe, J.J.; Vartanian, T. The toll-like receptor TLR4 is necessary for lipopolysaccharide-induced oligodendrocyte injury in the CNS. J. Neurosci. 2002, 22, 2478–2486. [Google Scholar] [CrossRef] [Green Version]
- Mallard, C.; Wang, X.; Hagberg, H. The role of Toll-like receptors in perinatal brain injury. Clin. Perinatol. 2009, 36, 763–772. [Google Scholar] [CrossRef]
- Wang, X.; Rousset, C.I.; Hagberg, H.; Mallard, C. Lipopolysaccharide-induced inflammation and perinatal brain injury. Semin Fetal Neonatal Med. 2006, 11, 343–353. [Google Scholar] [CrossRef]
- Yuan, T.M.; Sun, Y.; Zhan, C.Y.; Yu, H.M. Intrauterine infection/inflammation and perinatal brain damage: Role of glial cells and Toll-like receptor signaling. J. Neuroimmunol. 2010, 229, 16–25. [Google Scholar] [CrossRef]
- Bauman, M.D.; Van de Water, J. Translational opportunities in the prenatal immune environment: Promises and limitations of the maternal immune activation model. Neurobiol. Dis. 2020, 141, 104864. [Google Scholar] [CrossRef]
- Elovitz, M.A.; Wang, Z.; Chien, E.K.; Rychlik, D.F.; Phillippe, M. A new model for inflammation-induced preterm birth: The role of platelet-activating factor and Toll-like receptor-4. Am. J. Pathol. 2003, 163, 2103–2111. [Google Scholar] [CrossRef]
- Leitner, K.; Al Shammary, M.; McLane, M.; Johnston, M.V.; Elovitz, M.A.; Burd, I. IL-1 receptor blockade prevents fetal cortical brain injury but not preterm birth in a mouse model of inflammation-induced preterm birth and perinatal brain injury. Am. J. Reprod. Immunol. 2014, 71, 418–426. [Google Scholar] [CrossRef] [Green Version]
- Elovitz, M.A.; Brown, A.G.; Breen, K.; Anton, L.; Maubert, M.; Burd, I. Intrauterine inflammation, insufficient to induce parturition, still evokes fetal and neonatal brain injury. Int. J. Dev. Neurosci. 2011, 29, 663–671. [Google Scholar] [CrossRef] [Green Version]
- Elovitz, M.A.; Mrinalini, C.; Sammel, M.D. Elucidating the early signal transduction pathways leading to fetal brain injury in preterm birth. Pediatr. Res. 2006, 59, 50–55. [Google Scholar] [CrossRef] [Green Version]
- Burd, I.; Bentz, A.I.; Chai, J.; Gonzalez, J.; Monnerie, H.; Le Roux, P.D.; Cohen, A.S.; Yudkoff, M.; Elovitz, M.A. Inflammation-induced preterm birth alters neuronal morphology in the mouse fetal brain. J. Neurosci. Res. 2010, 88, 1872–1881. [Google Scholar] [CrossRef]
- Makinson, R.; Lloyd, K.; Rayasam, A.; McKee, S.; Brown, A.; Barila, G.; Grissom, N.; George, R.; Marini, M.; Fabry, Z.; et al. Intrauterine inflammation induces sex-specific effects on neuroinflammation, white matter, and behavior. Brain Behav. Immun. 2017, 66, 277–288. [Google Scholar] [CrossRef]
- Rosenkrantz, T.S.; Hussain, Z.; Fitch, R.H. Sex Differences in Brain Injury and Repair in Newborn Infants: Clinical Evidence and Biological Mechanisms. Front. Pediatr. 2019, 7, 211. [Google Scholar] [CrossRef]
- Kokras, N.; Dalla, C. Preclinical sex differences in depression and antidepressant response: Implications for clinical research. J. Neurosci. Res. 2017, 95, 731–736. [Google Scholar] [CrossRef] [Green Version]
- Boivin, J.R.; Piekarski, D.J.; Wahlberg, J.K.; Wilbrecht, L. Age, sex, and gonadal hormones differently influence anxiety- and depression-related behavior during puberty in mice. Psychoneuroendocrinology 2017, 85, 78–87. [Google Scholar] [CrossRef]
- Bucci, D.J.; Chiba, A.A.; Gallagher, M. Spatial learning in male and female Long-Evans rats. Behav. Neurosci. 2021, 135, 4–7. [Google Scholar] [CrossRef]
- Thagard, A.S.; Slack, J.L.; Estrada, S.M.; Kazanjian, A.A.; Chan, S.; Burd, I.; Napolitano, P.G.; Ieronimakis, N. Long-term impact of intrauterine neuroinflammation and treatment with magnesium sulphate and betamethasone: Sex-specific differences in a preterm labor murine model. Sci. Rep. 2017, 7, 17883. [Google Scholar] [CrossRef] [Green Version]
- Yagi, S.; Galea, L.A.M. Sex differences in hippocampal cognition and neurogenesis. Neuropsychopharmacology 2019, 44, 200–213. [Google Scholar] [CrossRef] [Green Version]
- Culig, L.; Surget, A.; Bourdey, M.; Khemissi, W.; Le Guisquet, A.M.; Vogel, E.; Sahay, A.; Hen, R.; Belzung, C. Increasing adult hippocampal neurogenesis in mice after exposure to unpredictable chronic mild stress may counteract some of the effects of stress. Neuropharmacology 2017, 126, 179–189. [Google Scholar] [CrossRef]
- Lazarov, O.; Hollands, C. Hippocampal neurogenesis: Learning to remember. Prog. Neurobiol. 2016, 138–140, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Hill, A.S.; Sahay, A.; Hen, R. Increasing Adult Hippocampal Neurogenesis is Sufficient to Reduce Anxiety and Depression-Like Behaviors. Neuropsychopharmacology 2015, 40, 2368–2378. [Google Scholar] [CrossRef]
- Revest, J.M.; Dupret, D.; Koehl, M.; Funk-Reiter, C.; Grosjean, N.; Piazza, P.V.; Abrous, D.N. Adult hippocampal neurogenesis is involved in anxiety-related behaviors. Mol. Psychiatry 2009, 14, 959–967. [Google Scholar] [CrossRef] [Green Version]
- Caruso, A.; Ricceri, L.; Scattoni, M.L. Ultrasonic vocalizations as a fundamental tool for early and adult behavioral phenotyping of Autism Spectrum Disorder rodent models. Neurosci. Biobehav. Rev. 2020, 116, 31–43. [Google Scholar] [CrossRef]
- Jensen, F.E.; Holmes, G.L.; Lombroso, C.T.; Blume, H.K.; Firkusny, I.R. Age-dependent changes in long-term seizure susceptibility and behavior after hypoxia in rats. Epilepsia 1992, 33, 971–980. [Google Scholar] [CrossRef]
- Simonet, J.C.; Sunnen, C.N.; Wu, J.; Golden, J.A.; Marsh, E.D. Conditional Loss of Arx from the Developing Dorsal Telencephalon Results in Behavioral Phenotypes Resembling Mild Human ARX Mutations. Cereb. Cortex 2015, 25, 2939–2950. [Google Scholar] [CrossRef] [Green Version]
- Yardeni, T.; Cristancho, A.G.; McCoy, A.J.; Schaefer, P.M.; McManus, M.J.; Marsh, E.D.; Wallace, D.C. An mtDNA mutant mouse demonstrates that mitochondrial deficiency can result in autism endophenotypes. Proc. Natl. Acad. Sci. USA 2021, 118, e2021429118. [Google Scholar] [CrossRef]
- Gawel, K.; Gibula, E.; Marszalek-Grabska, M.; Filarowska, J.; Kotlinska, J.H. Assessment of spatial learning and memory in the Barnes maze task in rodents-methodological consideration. Naunyn Schmiedebergs Arch. Pharmacol. 2019, 392, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Hester, M.S.; Tulina, N.; Brown, A.; Barila, G.; Elovitz, M.A. Intrauterine inflammation reduces postnatal neurogenesis in the hippocampal subgranular zone and leads to accumulation of hilar ectopic granule cells. Brain Res. 2018, 1685, 51–59. [Google Scholar] [CrossRef]
- Soucy-Giguere, L.; Gasse, C.; Giguere, Y.; Demers, S.; Bujold, E.; Boutin, A. Intra-amniotic inflammation and child neurodevelopment: A systematic review protocol. Syst. Rev. 2018, 7, 12. [Google Scholar] [CrossRef] [Green Version]
- Barroca, N.C.B.; Della Santa, G.; Suchecki, D.; Garcia-Cairasco, N.; Umeoka, E.H.L. Challenges in the use of animal models and perspectives for a translational view of stress and psychopathologies. Neurosci. Biobehav. Rev. 2022, 140, 104771. [Google Scholar] [CrossRef]
- Baharnoori, M.; Bhardwaj, S.K.; Srivastava, L.K. Neonatal behavioral changes in rats with gestational exposure to lipopolysaccharide: A prenatal infection model for developmental neuropsychiatric disorders. Schizophr. Bull. 2012, 38, 444–456. [Google Scholar] [CrossRef]
- Malkova, N.V.; Yu, C.Z.; Hsiao, E.Y.; Moore, M.J.; Patterson, P.H. Maternal immune activation yields offspring displaying mouse versions of the three core symptoms of autism. Brain Behav. Immun. 2012, 26, 607–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irie, F.; Badie-Mahdavi, H.; Yamaguchi, Y. Autism-like socio-communicative deficits and stereotypies in mice lacking heparan sulfate. Proc. Natl. Acad. Sci. USA 2012, 109, 5052–5056. [Google Scholar] [CrossRef] [Green Version]
- Choi, G.B.; Yim, Y.S.; Wong, H.; Kim, S.; Kim, H.; Kim, S.V.; Hoeffer, C.A.; Littman, D.R.; Huh, J.R. The maternal interleukin-17a pathway in mice promotes autism-like phenotypes in offspring. Science 2016, 351, 933–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez de Cossio, L.; Guzman, A.; van der Veldt, S.; Luheshi, G.N. Prenatal infection leads to ASD-like behavior and altered synaptic pruning in the mouse offspring. Brain Behav. Immun. 2017, 63, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Schwartzer, J.J.; Careaga, M.; Onore, C.E.; Rushakoff, J.A.; Berman, R.F.; Ashwood, P. Maternal immune activation and strain specific interactions in the development of autism-like behaviors in mice. Transl. Psychiatry 2013, 3, e240. [Google Scholar] [CrossRef] [Green Version]
- Rocha-Gomes, A.; Teixeira, A.E.; de Oliveira, D.G.; Santiago, C.M.O.; da Silva, A.A.; Riul, T.R.; Lacerda, A.C.R.; Mendonca, V.A.; Rocha-Vieira, E.; Leite, H.R. LPS tolerance prevents anxiety-like behavior and amygdala inflammation of high-fat-fed dams’ adolescent offspring. Behav. Brain Res. 2021, 411, 113371. [Google Scholar] [CrossRef]
- Gumusoglu, S.B.; Stevens, H.E. Maternal Inflammation and Neurodevelopmental Programming: A Review of Preclinical Outcomes and Implications for Translational Psychiatry. Biol. Psychiatry 2019, 85, 107–121. [Google Scholar] [CrossRef]
- Hava, G.; Vered, L.; Yael, M.; Mordechai, H.; Mahoud, H. Alterations in behavior in adult offspring mice following maternal inflammation during pregnancy. Dev. Psychobiol. 2006, 48, 162–168. [Google Scholar] [CrossRef]
- Smith, S.E.; Li, J.; Garbett, K.; Mirnics, K.; Patterson, P.H. Maternal immune activation alters fetal brain development through interleukin-6. J. Neurosci. 2007, 27, 10695–10702. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Meng, X.H.; Ning, H.; Zhao, X.F.; Wang, Q.; Liu, P.; Zhang, H.; Zhang, C.; Chen, G.H.; Xu, D.X. Age- and gender-dependent impairments of neurobehaviors in mice whose mothers were exposed to lipopolysaccharide during pregnancy. Toxicol. Lett. 2010, 192, 245–251. [Google Scholar] [CrossRef]
- Shi, L.; Fatemi, S.H.; Sidwell, R.W.; Patterson, P.H. Maternal influenza infection causes marked behavioral and pharmacological changes in the offspring. J. Neurosci. 2003, 23, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Meyer, U.; Nyffeler, M.; Engler, A.; Urwyler, A.; Schedlowski, M.; Knuesel, I.; Yee, B.K.; Feldon, J. The time of prenatal immune challenge determines the specificity of inflammation-mediated brain and behavioral pathology. J. Neurosci. 2006, 26, 4752–4762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, U. Prenatal poly(i:C) exposure and other developmental immune activation models in rodent systems. Biol. Psychiatry 2014, 75, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Chen, C.J.; Yan, X.X.; Li, Z.; Deng, X.H. Early-life lipopolysaccharide exposure potentiates forebrain expression of NLRP3 inflammasome proteins and anxiety-like behavior in adolescent rats. Brain Res. 2017, 1671, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, E.Y.; McBride, S.W.; Chow, J.; Mazmanian, S.K.; Patterson, P.H. Modeling an autism risk factor in mice leads to permanent immune dysregulation. Proc. Natl. Acad. Sci. USA 2012, 109, 12776–12781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Depino, A.M. Early prenatal exposure to LPS results in anxiety- and depression-related behaviors in adulthood. Neuroscience 2015, 299, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Dada, T.; Rosenzweig, J.M.; Al Shammary, M.; Firdaus, W.; Al Rebh, S.; Borbiev, T.; Tekes, A.; Zhang, J.; Alqahtani, E.; Mori, S.; et al. Mouse model of intrauterine inflammation: Sex-specific differences in long-term neurologic and immune sequelae. Brain Behav. Immun. 2014, 38, 142–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cristancho, A.G.; Gadra, E.C.; Samba, I.M.; Zhao, C.; Ouyang, M.; Magnitsky, S.; Huang, H.; Viaene, A.N.; Anderson, S.A.; Marsh, E.D. Deficits in Seizure Threshold and Other Behaviors in Adult Mice without Gross Neuroanatomic Injury after Late Gestation Transient Prenatal Hypoxia. Dev. Neurosci. 2022, 44, 246–265. [Google Scholar] [CrossRef]
- Eichenbaum, H.; Sauvage, M.; Fortin, N.; Komorowski, R.; Lipton, P. Towards a functional organization of episodic memory in the medial temporal lobe. Neurosci. Biobehav. Rev. 2012, 36, 1597–1608. [Google Scholar] [CrossRef] [Green Version]
- Gaesser, B.; Spreng, R.N.; McLelland, V.C.; Addis, D.R.; Schacter, D.L. Imagining the future: Evidence for a hippocampal contribution to constructive processing. Hippocampus 2013, 23, 1150–1161. [Google Scholar] [CrossRef]
- Schacter, D.L.; Addis, D.R.; Buckner, R.L. Remembering the past to imagine the future: The prospective brain. Nat. Rev. Neurosci. 2007, 8, 657–661. [Google Scholar] [CrossRef] [PubMed]
- Aimone, J.B.; Li, Y.; Lee, S.W.; Clemenson, G.D.; Deng, W.; Gage, F.H. Regulation and function of adult neurogenesis: From genes to cognition. Physiol. Rev. 2014, 94, 991–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Y.; Arruda-Carvalho, M.; Wang, J.; Janoschka, S.R.; Josselyn, S.A.; Frankland, P.W.; Ge, S. Optical controlling reveals time-dependent roles for adult-born dentate granule cells. Nat. Neurosci. 2012, 15, 1700–1706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohman, R.A.; Rhodes, J.S. Neurogenesis, inflammation and behavior. Brain Behav. Immun. 2013, 27, 22–32. [Google Scholar] [CrossRef] [Green Version]
- Lucassen, P.J.; Oomen, C.A.; Naninck, E.F.; Fitzsimons, C.P.; van Dam, A.M.; Czeh, B.; Korosi, A. Regulation of Adult Neurogenesis and Plasticity by (Early) Stress, Glucocorticoids, and Inflammation. Cold Spring Harb. Perspect. Biol. 2015, 7, a021303. [Google Scholar] [CrossRef] [Green Version]
- Kitabatake, Y.; Sailor, K.A.; Ming, G.L.; Song, H. Adult neurogenesis and hippocampal memory function: New cells, more plasticity, new memories? Neurosurg. Clin. N. Am. 2007, 18, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Lieberwirth, C.; Pan, Y.; Liu, Y.; Zhang, Z.; Wang, Z. Hippocampal adult neurogenesis: Its regulation and potential role in spatial learning and memory. Brain Res. 2016, 1644, 127–140. [Google Scholar] [CrossRef]
- Kelley, M.H.; Wu, W.W.; Lei, J.; McLane, M.; Xie, H.; Hart, K.D.; Pereira, L.; Burd, I.; Maylie, J. Functional changes in hippocampal synaptic signaling in offspring survivors of a mouse model of intrauterine inflammation. J. Neuroinflammation 2017, 14, 180. [Google Scholar] [CrossRef] [Green Version]
- Ng, P.C.; Wong, S.P.; Chan, I.H.; Lam, H.S.; Lee, C.H.; Lam, C.W. A prospective longitudinal study to estimate the "adjusted cortisol percentile" in preterm infants. Pediatr. Res. 2011, 69, 511–516. [Google Scholar] [CrossRef] [Green Version]
- Watterberg, K.L.; Scott, S.M.; Naeye, R.L. Chorioamnionitis, cortisol, and acute lung disease in very low birth weight infants. Pediatrics 1997, 99, E6. [Google Scholar] [CrossRef]
- Kim, J.J.; Diamond, D.M. The stressed hippocampus, synaptic plasticity and lost memories. Nat. Rev. Neurosci. 2002, 3, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Almela, M.; Hidalgo, V.; van der Meij, L.; Pulopulos, M.M.; Villada, C.; Salvador, A. A low cortisol response to acute stress is related to worse basal memory performance in older people. Front Aging Neurosci. 2014, 6, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boksa, P. Effects of prenatal infection on brain development and behavior: A review of findings from animal models. Brain Behav. Immun. 2010, 24, 881–897. [Google Scholar] [CrossRef] [PubMed]
- Giovanoli, S.; Notter, T.; Richetto, J.; Labouesse, M.A.; Vuillermot, S.; Riva, M.A.; Meyer, U. Late prenatal immune activation causes hippocampal deficits in the absence of persistent inflammation across aging. J. Neuroinflammation 2015, 12, 221. [Google Scholar] [CrossRef] [Green Version]
- Golan, H.M.; Lev, V.; Hallak, M.; Sorokin, Y.; Huleihel, M. Specific neurodevelopmental damage in mice offspring following maternal inflammation during pregnancy. Neuropharmacology 2005, 48, 903–917. [Google Scholar] [CrossRef]
- Khan, D.; Fernando, P.; Cicvaric, A.; Berger, A.; Pollak, A.; Monje, F.J.; Pollak, D.D. Long-term effects of maternal immune activation on depression-like behavior in the mouse. Transl. Psychiatry 2014, 4, e363. [Google Scholar] [CrossRef] [Green Version]
- Solek, C.M.; Farooqi, N.; Verly, M.; Lim, T.K.; Ruthazer, E.S. Maternal immune activation in neurodevelopmental disorders. Dev Dyn 2018, 247, 588–619. [Google Scholar] [CrossRef]
- Glass, R.; Norton, S.; Fox, N.; Kusnecov, A.W. Maternal immune activation with staphylococcal enterotoxin A produces unique behavioral changes in C57BL/6 mouse offspring. Brain Behav. Immun. 2019, 75, 12–25. [Google Scholar] [CrossRef]
- Jin, J.; Kim, S.N.; Liu, X.; Zhang, H.; Zhang, C.; Seo, J.S.; Kim, Y.; Sun, T. miR-17-92 Cluster Regulates Adult Hippocampal Neurogenesis, Anxiety, and Depression. Cell Rep. 2016, 16, 1653–1663. [Google Scholar] [CrossRef] [Green Version]
- Goncalves, J.T.; Schafer, S.T.; Gage, F.H. Adult Neurogenesis in the Hippocampus: From Stem Cells to Behavior. Cell 2016, 167, 897–914. [Google Scholar] [CrossRef]
- Hofer, M.A.; Shair, H.N.; Masmela, J.R.; Brunelli, S.A. Developmental effects of selective breeding for an infantile trait: The rat pup ultrasonic isolation call. Dev. Psychobiol. 2001, 39, 231–246. [Google Scholar] [CrossRef] [PubMed]
- Spijker, S. Dissection of Rodent Brain Regions. In Neuroproteomics (Neuromethods); Li, K., Ed.; Humana Press: Totowa, NJ, USA, 2011. [Google Scholar]
- Warde-Farley, D.; Donaldson, S.L.; Comes, O.; Zuberi, K.; Badrawi, R.; Chao, P.; Franz, M.; Grouios, C.; Kazi, F.; Lopes, C.T.; et al. The GeneMANIA prediction server: Biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res. 2010, 38, W214-20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halekoh, U.; Højsgaard, S.; Yan, J. The R Package gee pack for Generalized Estimating Equations. J. Stat. Softw. 2006, 15, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cristancho, A.G.; Tulina, N.; Brown, A.G.; Anton, L.; Barila, G.; Elovitz, M.A. Intrauterine Inflammation Leads to Select Sex- and Age-Specific Behavior and Molecular Differences in Mice. Int. J. Mol. Sci. 2023, 24, 32. https://doi.org/10.3390/ijms24010032
Cristancho AG, Tulina N, Brown AG, Anton L, Barila G, Elovitz MA. Intrauterine Inflammation Leads to Select Sex- and Age-Specific Behavior and Molecular Differences in Mice. International Journal of Molecular Sciences. 2023; 24(1):32. https://doi.org/10.3390/ijms24010032
Chicago/Turabian StyleCristancho, Ana G., Natalia Tulina, Amy G. Brown, Lauren Anton, Guillermo Barila, and Michal A. Elovitz. 2023. "Intrauterine Inflammation Leads to Select Sex- and Age-Specific Behavior and Molecular Differences in Mice" International Journal of Molecular Sciences 24, no. 1: 32. https://doi.org/10.3390/ijms24010032
APA StyleCristancho, A. G., Tulina, N., Brown, A. G., Anton, L., Barila, G., & Elovitz, M. A. (2023). Intrauterine Inflammation Leads to Select Sex- and Age-Specific Behavior and Molecular Differences in Mice. International Journal of Molecular Sciences, 24(1), 32. https://doi.org/10.3390/ijms24010032