
microRNAs Are Abundant and Stable in Platelet-Rich Fibrin and Other Autologous Blood Products of Canines

 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
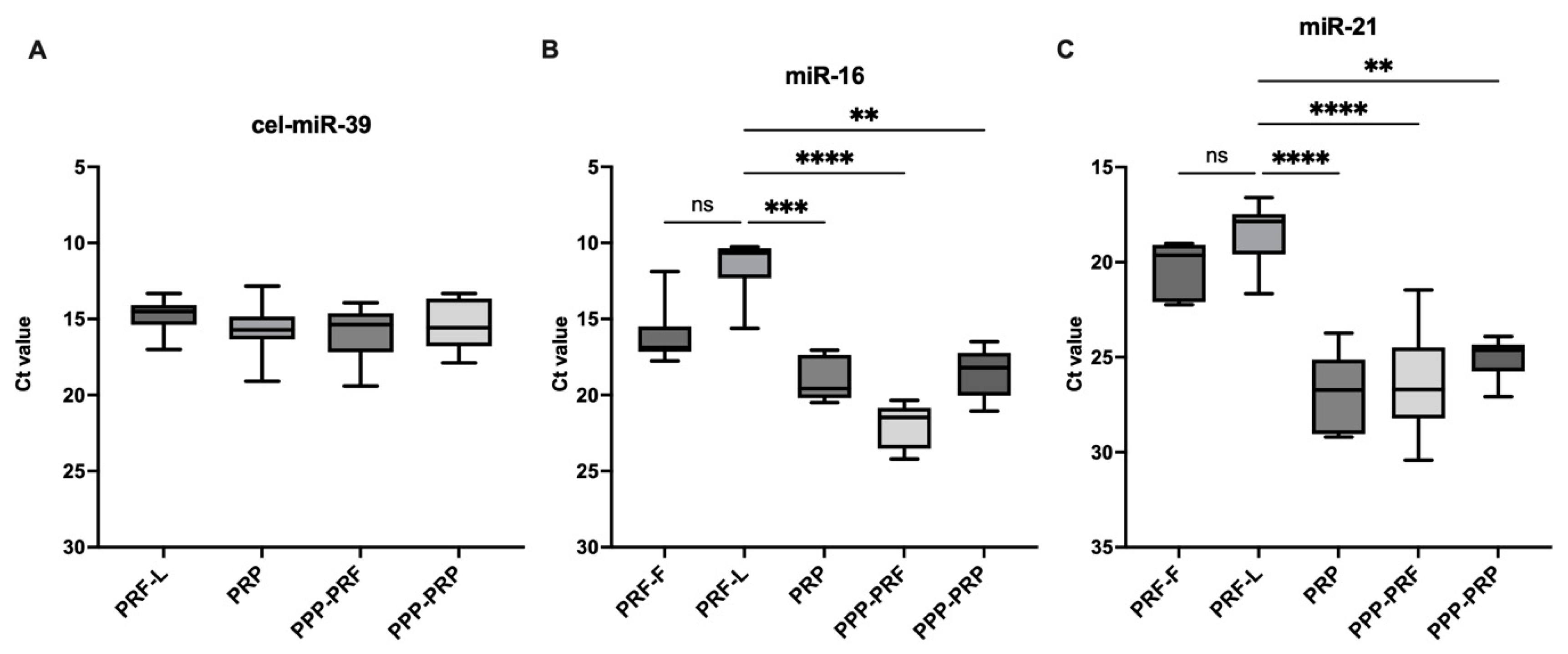
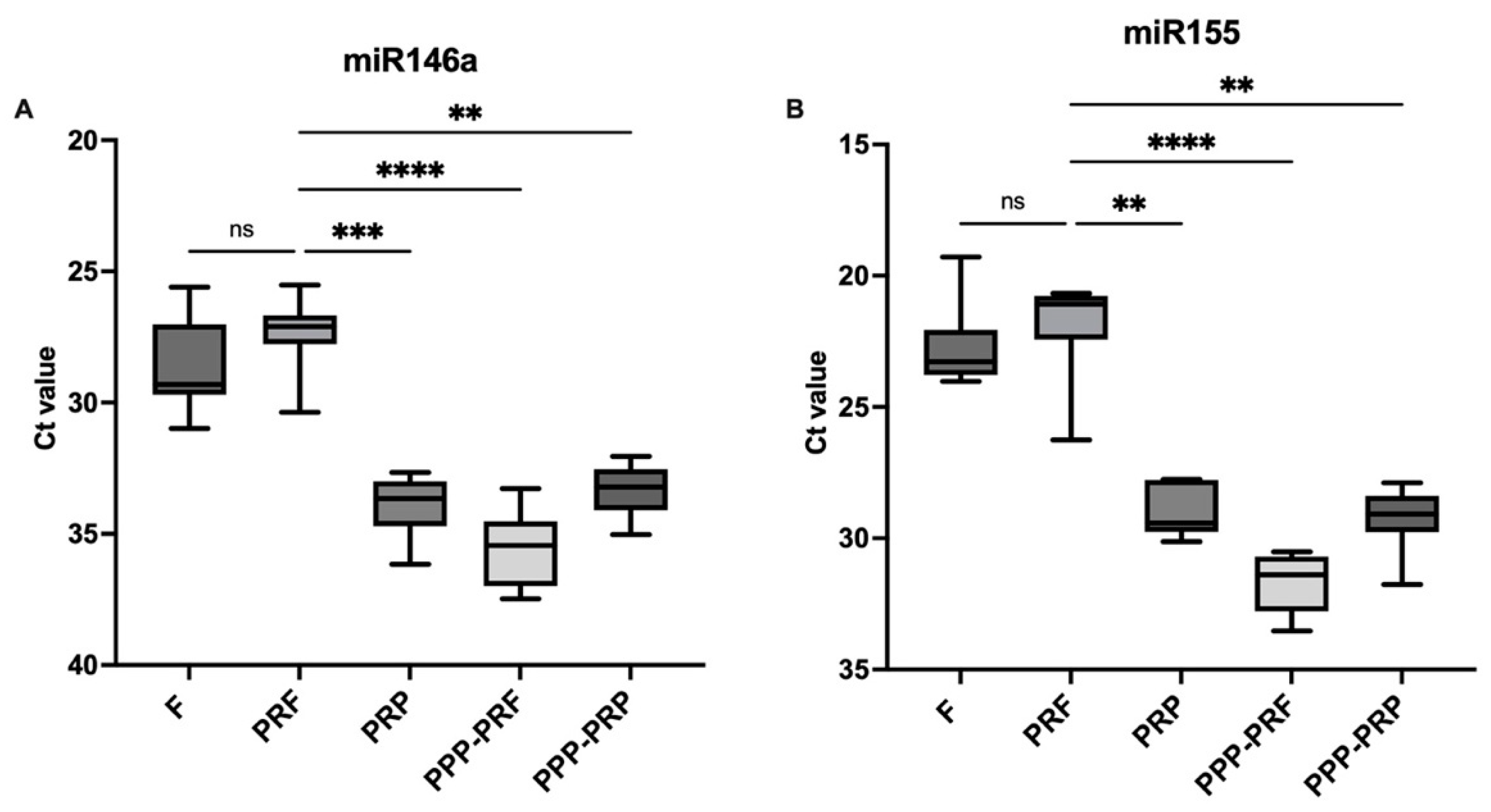
2.1. miRNAs Are More Abundant in PRF than Other Autologous Blood Products of Canines
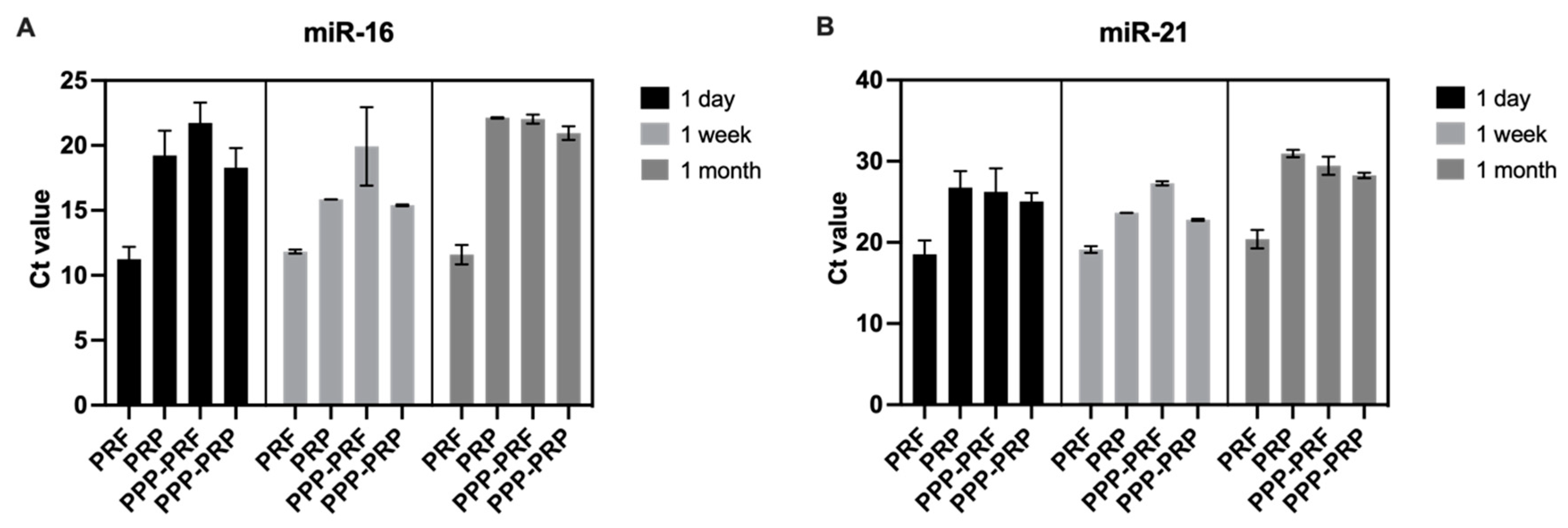
2.2. miRNAs Are Stable in Different Autologous Blood Products of Canines after Frozen at −30 °C
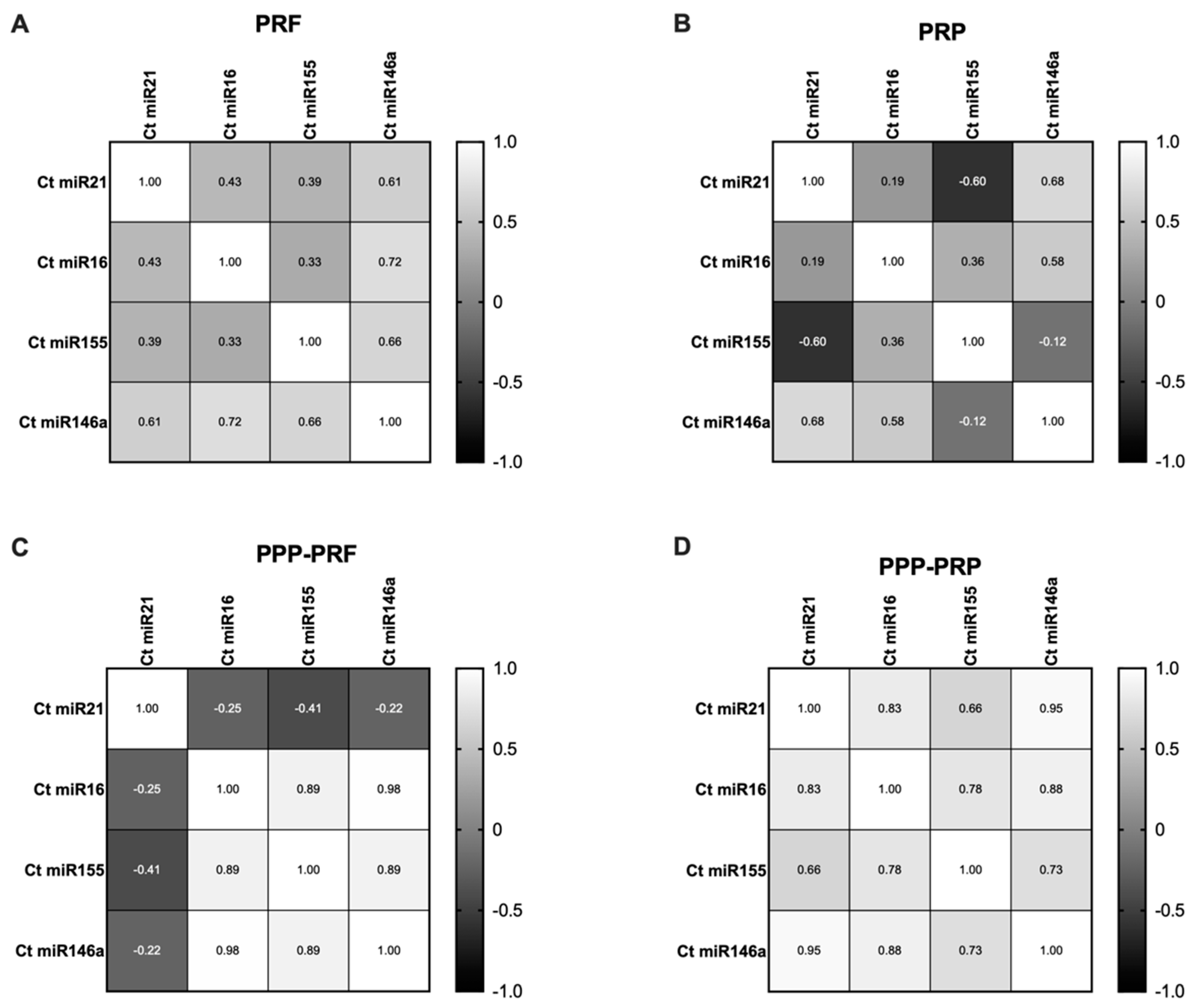
2.3. miRNAs Expression Status of Different Autologous Canine Blood Products
3. Discussion
4. Materials and Methods
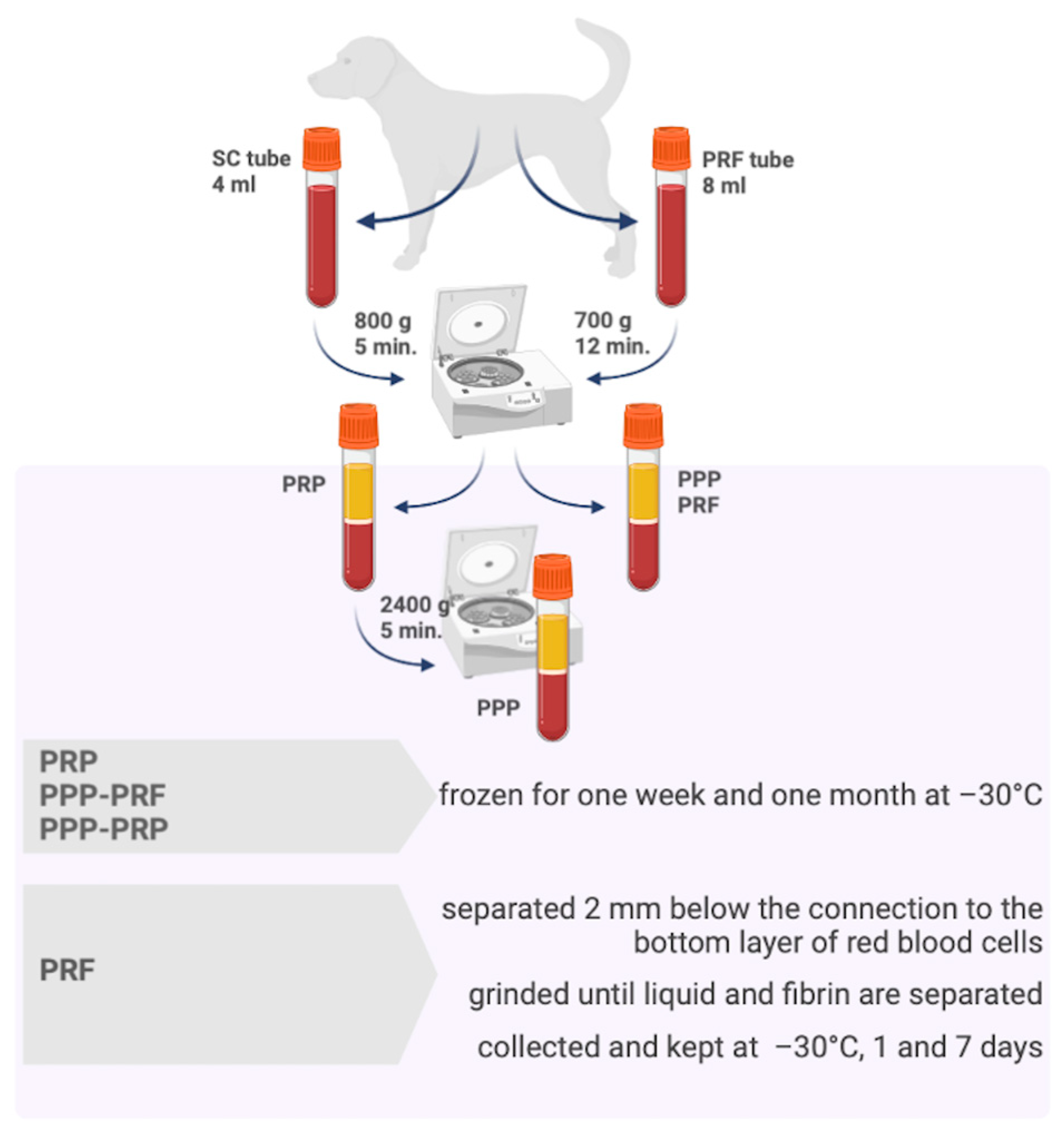
4.1. The Whole Blood Sample Collection and Preparation of PRF, PRP, and PPP
4.2. RNA Extraction
4.3. Quantitative Real-Time PCR
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arabaci, T.; Albayrak, M. Titanium-Prepared Platelet-Rich Fibrin Provides Advantages on Periodontal Healing: A Randomized Split-Mouth Clinical Study: T-PRF Contributes to Periodontal Healing. J. Periodontol. 2018, 89, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Barbu, H.M.; Andreescu, C.F.; Comaneanu, M.R.; Referendaru, D.; Mijiritsky, E. Maxillary Sinus Floor Augmentation to Enable One-Stage Implant Placement by Using Bovine Bone Substitute and Platelet-Rich Fibrin. BioMed Res. Int. 2018, 2018, 6562958. [Google Scholar] [CrossRef] [PubMed]
- Soares, C.S.; Barros, L.C.; Saraiva, V.; Gomez-Florit, M.; Babo, P.S.; Dias, I.R.; Reis, R.L.; Carvalho, P.P.; Gomes, M.E. Bioengineered Surgical Repair of a Chronic Oronasal Fistula in a Cat Using Autologous Platelet-Rich Fibrin and Bone Marrow with a Tailored 3D Printed Implant. J. Feline Med. Surg. 2018, 20, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Ghanaati, S.; Booms, P.; Orlowska, A.; Kubesch, A.; Lorenz, J.; Rutkowski, J.; Landes, C.; Sader, R.; Kirkpatrick, C.; Choukroun, J. Advanced Platelet-Rich Fibrin: A New Concept for Cell-Based Tissue Engineering by Means of Inflammatory Cells. J. Oral Implantol. 2014, 40, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Bahammam, M.A. Effect of Platelet-Rich Fibrin Palatal Bandage on Pain Scores and Wound Healing after Free Gingival Graft: A Randomized Controlled Clinical Trial. Clin. Oral Investig. 2018, 22, 3179–3188. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, E.; Flückiger, L.; Fujioka-Kobayashi, M.; Sawada, K.; Sculean, A.; Schaller, B.; Miron, R.J. Comparative Release of Growth Factors from PRP, PRF, and Advanced-PRF. Clin. Oral Investig. 2016, 20, 2353–2360. [Google Scholar] [CrossRef]
- Liu, J.; Lu, F.; Chen, H.; Bao, R.; Li, Z.; Lu, B.; Yu, K.; Dai, F.; Wu, D.; Lan, G. Healing of Skin Wounds Using a New Cocoon Scaffold Loaded with Platelet-Rich or Platelet-Poor Plasma. RSC Adv. 2017, 7, 6474–6485. [Google Scholar] [CrossRef] [Green Version]
- Doss, J.F.; Corcoran, D.L.; Jima, D.D.; Telen, M.J.; Dave, S.S.; Chi, J.-T. A Comprehensive Joint Analysis of the Long and Short RNA Transcriptomes of Human Erythrocytes. BMC Genom. 2015, 16, 952. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Yu, Y.; Niu, B.; Wang, D. Red Blood Cells as Potential Repositories of MicroRNAs in the Circulatory System. Front. Genet. 2020, 11, 442. [Google Scholar] [CrossRef]
- Ke, Y.; Zhao, W.; Xiong, J.; Cao, R. Downregulation of MiR-16 Promotes Growth and Motility by Targeting HDGF in Non-Small Cell Lung Cancer Cells. FEBS Lett. 2013, 587, 3153–3157. [Google Scholar] [CrossRef]
- Sa, F. Control of Immunoregulatory Molecules by MiRNAs in T Cell Activation. Front. Immunol. 2018, 9, 10. [Google Scholar]
- Niemiec, S.M.; Louiselle, A.E.; Hilton, S.A.; Dewberry, L.C.; Zhang, L.; Azeltine, M.; Xu, J.; Singh, S.; Sakthivel, T.S.; Seal, S.; et al. Nanosilk Increases the Strength of Diabetic Skin and Delivers CNP-MiR146a to Improve Wound Healing. Front. Immunol. 2020, 11, 590285. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.Y.; Kim, H.; Kwak, G.; Jo, S.D.; Cho, D.; Yang, Y.; Chan, I.; Jeong, J.H.; Kim, S.H. Development of MicroRNA-21 Mimic Nanocarriers for the Treatment of Cutaneous Wounds. Theranostics 2020, 10, 3240. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Schambach, F.; DeJong, C.S.; Hammond, S.M. Micro-RNA-155 Inhibits IFN-γ Signaling in CD4+ T Cells. Eur. J. Immunol. 2010, 40, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Dou, H.; Li, X.; Zhao, X.; Li, Y.; Liu, D.; Ji, J.; Liu, F.; Ding, L.; Ni, Y.; et al. Exosomal MiR-146a Contributes to the Enhanced Therapeutic Efficacy of Interleukin-1β-Primed Mesenchymal Stem Cells Against Sepsis. Stem Cells 2017, 35, 1208–1221. [Google Scholar] [CrossRef] [Green Version]
- Muth, D.C.; Powell, B.H.; Zhao, Z.; Witwer, K.W. MiRNAs in Platelet-Poor Blood Plasma and Purified RNA Are Highly Stable: A Confirmatory Study. BMC Res. Notes 2018, 11, 273. [Google Scholar] [CrossRef]
- Baćević, M.; Brković, B.; Lambert, F.; Djukić, L.; Petrović, N.; Roganović, J. Leukocyte- and Platelet-Rich Fibrin as Graft Material Improves MicroRNA-21 Expression and Decreases Oxidative Stress in the Calvarial Defects of Diabetic Rabbits. Arch. Oral Biol. 2019, 102, 231–237. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, Y.; Choukroun, J.; Ghanaati, S.; Miron, R. Behavior of Gingival Fibroblasts on Titanium Implant Surfaces in Combination with Either Injectable-PRF or PRP. Int. J. Mol. Sci. 2017, 18, 331. [Google Scholar] [CrossRef] [Green Version]
- Krammer, T.L.; Mayr, M.; Hackl, M. MicroRNAs as Promising Biomarkers of Platelet Activity in Antiplatelet Therapy Monitoring. Int. J. Mol. Sci. 2020, 21, 3477. [Google Scholar] [CrossRef]
- Willeit, P.; Zampetaki, A.; Dudek, K.; Kaudewitz, D.; King, A.; Kirkby, N.S.; Crosby-Nwaobi, R.; Prokopi, M.; Drozdov, I.; Langley, S.R.; et al. Circulating MicroRNAs as Novel Biomarkers for Platelet Activation. Circ. Res. 2013, 112, 595–600. [Google Scholar] [CrossRef] [Green Version]
- Nagalla, S.; Shaw, C.; Kong, X.; Kondkar, A.A.; Edelstein, L.C.; Ma, L.; Chen, J.; McKnight, G.S.; Dong, J.; Bray, P.F. Platelet MicroRNA-MRNA Coexpression Profiles Correlate with Platelet Reactivity. Blood J. Am. Soc. Hematol. 2011, 117, 5189–5197. [Google Scholar] [CrossRef] [PubMed]
- Caporali, A.; Emanueli, C. MicroRNA-503 and the Extended MicroRNA-16 Family in Angiogenesis. Trends Cardiovasc. Med. 2011, 21, 162–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hildreth, R.; Georgantas, R.W.; Patel, R.; Morisot, S.; Alder, J.; Civin, C.I. MicroRNA Hsa-Mir-16 Contributes to Regulation of Myeloid Differentiation of Human CD34+ Cells. Blood 2006, 108, 1343. [Google Scholar] [CrossRef]
- Calin, G.A.; Dumitru, C.D.; Shimizu, M.; Bichi, R.; Zupo, S.; Noch, E.; Aldler, H.; Rattan, S.; Keating, M.; Rai, K.; et al. Frequent Deletions and Down-Regulation of Micro- RNA Genes MiR15 and MiR16 at 13q14 in Chronic Lymphocytic Leukemia. Proc. Natl. Acad. Sci. USA 2002, 99, 15524–15529. [Google Scholar] [CrossRef] [Green Version]
- Ratajczak, J.; Vangansewinkel, T.; Gervois, P.; Merckx, G.; Hilkens, P.; Quirynen, M.; Lambrichts, I.; Bronckaers, A. Angiogenic Properties of ‘Leukocyte- and Platelet-Rich Fibrin’. Sci. Rep. 2018, 8, 14632. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Wu, W.; Zhang, L.; Dorset-Martin, W.; Morris, M.W.; Mitchell, M.E.; Liechty, K.W. The Role of MicroRNA-146a in the Pathogenesis of the Diabetic Wound-Healing Impairment. Diabetes 2012, 61, 2906–2912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto, N.R.; Ubilla, M.; Zamora, Y.; Del Rio, V.; Dohan Ehrenfest, D.M.; Quirynen, M. Leucocyte- and Platelet-Rich Fibrin (L-PRF) as a Regenerative Medicine Strategy for the Treatment of Refractory Leg Ulcers: A Prospective Cohort Study. Platelets 2018, 29, 468–475. [Google Scholar] [CrossRef] [Green Version]
- Luly, F.R.; Lévêque, M.; Licursi, V.; Cimino, G.; Martin-Chouly, C.; Théret, N.; Negri, R.; Cavinato, L.; Ascenzioni, F.; Del Porto, P. MiR-146a Is over-Expressed and Controls IL-6 Production in Cystic Fibrosis Macrophages. Sci. Rep. 2019, 9, 16259. [Google Scholar] [CrossRef] [Green Version]
- Kargarpour, Z.; Nasirzade, J.; Panahipour, L.; Miron, R.J.; Gruber, R. Platelet-Rich Fibrin Decreases the Inflammatory Response of Mesenchymal Cells. Int. J. Mol. Sci. 2021, 22, 11333. [Google Scholar] [CrossRef]
- Kaur, S.; Abu-Shahba, A.G.; Paananen, R.O.; Hongisto, H.; Hiidenmaa, H.; Skottman, H.; Seppänen-Kaijansinkko, R.; Mannerström, B. Small Non-Coding RNA Landscape of Extracellular Vesicles from Human Stem Cells. Sci. Rep. 2018, 8, 15503. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, A.K.; Mal, C. Underpinning MiRNA-MiRNA Co-Functional Interaction Patterns in the Metabolism of Oryza Sativa by Genome-Scale Network Analysis. Heliyon 2020, 6, e05496. [Google Scholar] [CrossRef]
- Lai, Y.; Lai, Y.; Rahman, M.M.; Chen, H.; Husna, A.A.; Fujikawa, T.; Ando, T.; Kitahara, G.; Koiwa, M.; Kubota, C.; et al. Bovine Milk Transcriptome Analysis Reveals MicroRNAs and RNU2 Involved in Mastitis. FEBS J. 2020, 287, 1899–1918. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Lai, Y.; Husna, A.A.; Chen, H.; Tanaka, Y.; Kawaguchi, H.; Hatai, H.; Miyoshi, N.; Nakagawa, T.; Fukushima, R.; et al. Aberrantly Expressed SnoRNA, SnRNA, PiRNA and TRFs in Canine Melanoma. Vet. Comp. Oncol. 2020, 18, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Lai, Y.-C.; Husna, A.A.; Chen, H.; Tanaka, Y.; Kawaguchi, H.; Miyoshi, N.; Nakagawa, T.; Fukushima, R.; Miura, N. Micro RNA Transcriptome Profile in Canine Oral Melanoma. Int. J. Mol. Sci. 2019, 20, 4832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing Real-Time PCR Data by the Comparative CT Method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Binderup, H.G.; Madsen, J.S.; Heegaard, N.H.H.; Houlind, K.; Andersen, R.F.; Brasen, C.L. Quantification of MicroRNA Levels in Plasma—Impact of Preanalytical and Analytical Conditions. PLoS ONE 2018, 13, e0201069. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Dogs (n = 5) |
|---|---|
| Age, y/o | |
| 0–8 (adult) | 5 |
| 9–17 (senior) | 0 |
| Sex | |
| Female, n (%) | 3 (60) |
| Male, n (%) | 2 (40) |
| Weight, kg | |
| 0–10, n (%) | 1 (20) |
| 11–20, n (%) | 4 (80) |
| Breed | |
| Mix | 3 |
| Shiba | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jasineviciute, I.; Hasan, M.N.; Grigas, J.; Pautienius, A.; Stankevicius, A.; Zymantiene, J.; Miura, N. microRNAs Are Abundant and Stable in Platelet-Rich Fibrin and Other Autologous Blood Products of Canines. Int. J. Mol. Sci. 2023, 24, 770. https://doi.org/10.3390/ijms24010770
Jasineviciute I, Hasan MN, Grigas J, Pautienius A, Stankevicius A, Zymantiene J, Miura N. microRNAs Are Abundant and Stable in Platelet-Rich Fibrin and Other Autologous Blood Products of Canines. International Journal of Molecular Sciences. 2023; 24(1):770. https://doi.org/10.3390/ijms24010770
Chicago/Turabian StyleJasineviciute, Indre, Md Nazmul Hasan, Juozas Grigas, Arnoldas Pautienius, Arunas Stankevicius, Judita Zymantiene, and Naoki Miura. 2023. "microRNAs Are Abundant and Stable in Platelet-Rich Fibrin and Other Autologous Blood Products of Canines" International Journal of Molecular Sciences 24, no. 1: 770. https://doi.org/10.3390/ijms24010770
APA StyleJasineviciute, I., Hasan, M. N., Grigas, J., Pautienius, A., Stankevicius, A., Zymantiene, J., & Miura, N. (2023). microRNAs Are Abundant and Stable in Platelet-Rich Fibrin and Other Autologous Blood Products of Canines. International Journal of Molecular Sciences, 24(1), 770. https://doi.org/10.3390/ijms24010770